Introduction
Autophagy is a basic cellular maintenance mechanism,
by which unnecessary proteins and organelles are degraded in order
to maintain cellular survival and homeostasis under various
conditions of intracellular stress (1). Autophagy is becoming a focus of
research in endothelial cells in regards to atherosclerosis
prevention. Endothelial cell injury is a major step in the
pathological progression of atherosclerosis. Autophagy may occur to
protect the cells following this injury (2). When autophagy fails or is inhibited,
the breakdown of endothelium integrity facilitates local lipid
deposition, resulting in atherogenesis, plaque instability, acute
vascular occlusion and even sudden mortality (3,4). The
mechanism of autophagy regulation is complex, and is thought to be
linked to multiple signaling pathways (5–7). The
phosphatidylinositol-3 kinase (PI3K)/RAC-α serine/threonine-protein
kinase (Akt)/mechanistic target of rapamycin (mTOR) signaling
pathway is an essential regulator in cellular proliferation,
survival, metabolism and autophagy (8). In addition, this pathway may be
involved in the induction of autophagy in atherosclerosis (9,10).
However, the molecular mechanisms that control endothelial cell
autophagy in the development of atherosclerosis remain to be fully
elucidated.
MicroRNAs (miRNAs/miRs) are small 19–24 nucleotide
non-coding RNAs that function in the post-transcriptional
regulation of target gene expression (11). Previous research has revealed that
miRNAs are essential for autophagy regulation in atherosclerosis
(12–14). Previously, miR-155 has been
reported to have a pivotal role in the function of endothelial cell
apoptosis regulation, vascular smooth muscle migration, lipid
metabolism regulation and the inflammatory response, thus affecting
the incidence of atherosclerosis through physiological and
pathological processes (15);
however, the association between miR-155 and autophagy in
endothelial cells has not been fully elucidated. Therefore, the the
present study aimed to investigate the role of miR-155 in
autophagy.
Our previous study demonstrated that miR-155
promotes oxidized low-density lipoprotein (ox-LDL)-induced
autophagy in human umbilical vein endothelial cells (HUVECs)
(16). The present study
investigated whether miR-155 regulated vascular endothelial cell
autophagy via the PI3K/Akt/mTOR signaling pathway. Bioinformatic
analyses were conducted to predict target genes for miR-155 in the
PI3K/Akt/mTOR signaling pathway and it was revealed that PI3K
catalytic subunit a (PIK3CA) and Ras homolog enriched in brain
(Rheb) were direct targets of miR-155. The results suggested that
miR-155 promoted autophagy, and that this may have occurred through
the targeting of the PI3K/Akt/mTOR pathway in vascular endothelial
cells.
Materials and methods
Cell culture
HUVECs and 293T cells were purchased from the
Shanghai Institute for Biological Sciences, Chinese Academy of
Sciences (Shanghai, China). According to the supplier's protocols,
cells were cultured in Dulbecco's modified Eagle's medium (Hyclone;
GE Healthcare Life Sciences, Logan, UT, USA) supplemented with 10%
fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) and 1% antimycotic-antibiotic solution (Beijing Solarbio
Science & Technology Co., Ltd., Beijing, China) in a humidified
atmosphere of 5% CO2 at 37°C.
Cell treatment
miR-155 mimics, inhibitor and negative controls
(NCs) were purchased from Guangzhou RiboBio Co., Ltd. (Guangzhou,
China). Cells were transfected with 50 nmol/l miR-155 mimics,
miR-155 inhibitor or NCs for 6 h, using Lipofectamine®
2000 (Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol; complete medium containing FBS was then
added for 24 h. In order to induce autophagy, HUVECs were
stimulated with 100 µg/ml ox-LDL (Guangzhou Yiyuan Biotech Co.,
Ltd., Guangzhou, China) for 12 h. They were subsequently treated
with 0.5 µmol/l bafilomycin A1 for 12 h at 37°C to block
cytolysosome and lysosome fusion. HUVECs were divided into seven
experimental groups: i) Control; ii) ox-LDL + miR-155 inhibitor;
iii) ox-LDL + miR-155 mimic; iv) ox-LDL + bafilomycin A1; v)
ox-LDL; vi) ox-LDL + miR-155 inhibitor-NC; and vii) ox-LDL +
miR-155 mimic-NC.
Transmission electron microscopy
(TEM)
HUVECs were plated in 10 cm petri dishes in a
humidified atmosphere of 5% CO2 at 37°C for 24 h. Cells
at 70–80% confluence were used for experiments. The cells were
trypsinized, washed once with PBS and harvested by centrifugation
at 20,000 × g at 4°C for 10 min. The cell pellet (0.5 cm) was fixed
in 10 ml ice-cold 2.5% glutaraldehyde in 0.1M sodium cacodylate
buffer (pH 7.2) at 4°C overnight. Samples were sent to the
laboratory of Qingdao University (Qingdao, China), and subsequently
post-fixed in PBS with 1% osmium tetroxide for 1 h at 4°C, followed
by dehydration in graded ethanol (70% for 20 min, 96% for 20 min,
and 100% ethanol for 20 min twice) and propylene oxide. Cells were
embedded in epoxy resin. Sectioned grids were stained with 2%
uranyl acetate in 50% methanol for 10 min and lead citrate for 7
min at 4°C. Images were captured on a transmission electron
microscope (JEM-1220; JEOL, Ltd., Tokyo, Japan) at an accelerating
voltage of 80 kV and a magnification of ×40,000. A total of 10
field of view were randomly selected to observe autophagosomes in
each group.
Confocal microscopy
Cells (3×104 cells per well) were
cultured on glass coverslips in a 24-well culture plate covered
with cell culture medium (DMEM with 10% FBS and 1%
antimycotic-antibiotic solution) in a humidified atmosphere of 5%
CO2 at 37°C. Next, the cells on coverslips were fixed
using 4% paraformaldehyde (Beijing Solarbio Science &
Technology Co., Ltd.) for 15 min at 4°C and permeabilized with 1%
Triton X-100 (Beijing Solarbio Science & Technology Co., Ltd.)
for 10 min. Following washing of the coverslips, cells were blocked
with 3% bovine serum albumin (BSA; Beyotime Institute of
Biotechnology, Haimen, China) to prevent non-specific antibody
binding and incubated at 4°C overnight with 1% BSA in PBS 0.5%
Tween containing antibodies against microtubule-associated protein
light chain 3 (LC3; cat. no. 3868S; 1:100; Cell Signaling
Technology, Inc., Danvers, MA, USA). Cells were subsequently washed
twice with PBS 0.5% Tween and incubated with fluorescein
isothiocyanate-conjugated goat anti-rabbit IgG secondary antibodies
(1:200; BIOSS, Beijing, China) in 1% BSA in the dark for 1 h at
37°C. The cells were washed in PBS and nuclei were stained with 10
µg/ml of DAPI (Shanghai Yeasen Biotechnology Co., Ltd., Shanghai,
China) for 5 min in the dark. Cells were washed again and antifade
mounting medium (Beyotime institute of Biotechnology) was added
prior to mounting on glass slides for examination. Images were
acquired using a laser-scanning confocal imaging system
(magnification, ×25,000; TCS SP5; Leica Microsystems GmbH, Wetzlar,
Germany).
Western blot analysis
Protein extracts were prepared from cells using
radioimmunoprecipitation assay lysis buffer containing protease and
phosphatase inhibitors (Beyotime Institute of Biotechnology).
Following centrifugation at 4°C for 5 min at 30,000 × g the
supernatant was collected and protein concentrations in the lysates
were measured using a bicinchoninic acid protein assay kit
(Beyotime Institute of Biotechnology). Total protein (50 mg) was
separated by 10% SDS-PAGE and transferred onto polyvinylidene
difluoride membranes, which were blocked with 5% skimmed milk
powder in Tris-buffered saline with 0.1% Tween 20 (Beijing Solarbio
Science & Technology Co., Ltd.) for 2 h at 4°C. The membranes
were incubated with the following primary antibodies at 4°C
overnight: Rabbit anti-LC3B (cat. no. ab48394; Abcam), rabbit
anti-PI3K (cat. no. 4257; Cell Signaling Technology, Inc.), rabbit
anti-phosphorylated (p)-PI3K (cat. no. 5538; Cell Signaling
Technology, Inc.), rabbit anti-Rheb (cat. no. 13879; Cell Signaling
Technology, Inc.), rabbit anti-Akt (cat. no. 4691S; Cell Signaling
Technology, Inc.), rabbit anti-p-Akt (cat. no. 4060; Cell Signaling
Technology, Inc.), rabbit anti-70 kDa ribosomal protein S6 kinase 1
(p70s6k; cat. no. 2708; Cell Signaling Technology, Inc.), rabbit
anti-p-p70s6k (cat. no. ab1314362; Abcam) and rabbit anti-GAPDH
(cat. no. CW0101; Beijing ComWin Biotech Co., Ltd., Beijing, China)
antibodies. All primary antibodies were diluted to 1:500. Following
washing with 0.1% TBST, membranes were treated with a horseradish
peroxidase-conjugated goat anti-rabbit immunoglobulin G secondary
antibody (cat. no. CW0101, 1:2,000; Beijing ComWin Biotech Co.,
Ltd.) for 2 h at 37°C. The signals were detected with enhanced
chemiluminescence kit (EMD Millipore, Billerica, MA, USA). The
relative densities of LC3, PI3K, p-PI3K, Rheb, Akt, p-Akt, p70s6k
and p-p70s6k were determined by normalization to the density value
of GAPDH in the same blot. Relative band intensity was analyzed
using ImageJ 1.8.0 software (National Institutes of Health,
Bethesda, MD, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocol. miRNA was reverse transcribed into
cDNA (25°C for 10 min, 42°C for 50 min, followed by 70°C for 15
min) using an miRNA cDNA synthesis kit (Beijing Aidiab
Biotechnologies Co., Ltd., Beijing, China) and qPCR was performed
with a SYBR Green miRNA qPCR assay kit (Beijing ComWin Biotech Co.,
Ltd., Beijing, China). mRNA was reverse transcribed (25°C for 10
min, 42°C for 50 min, followed by 70°C for 15 min) using a
PrimeScript™ RT reagent kit (Beijing Aidiab Biotechnologies Co.,
Ltd.,) and qPCR was performed with SYBR® Premix Ex Taq™
(Takara Bio, Inc., Otsu, Japan). The housekeeping gene GAPDH was
used as an internal control; the internal control employed for the
normalization of miRNA expression levels was U6 (Sangon Biotech
Co., Ltd. Shanghai, China). PI3K, Rheb and GAPDH primers were
purchased from Sangon Biotech Co., Ltd. The mature miR-155 primer
(qRT-PCR Primer Set, cat. no. 132, Sanger Registry ID Human
has-miR-155, Sanger Accession no. Human MIMAT0000646, mature
sequence UUAAUGCUAAUCGUGAUAGGGGU) was purchased from Takara Bio,
Inc. The sequences of each specific primer were as follows: PI3K
forward, 5′-GACTTTGCGACAAGACTGCC-3′ and reverse,
5′-AATCTGAAGCAGCGCCTGAA-3′; Rheb forward,
5′-AGCTTTGGCAGAATCTTGGA-3′ and reverse, 5′-CACATCACCGAGCATGAAGA-3′;
GAPDH forward, 5′-AGAAGGCTGGGGCTCATTTG-3′ and reverse,
5′-AGAAGGCTGGGGCTCATTTG-3; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and
reverse, 5′-AACGCTTCACGAATTTGCGT-3′. A total of three independent
experiments were performed under the same experimental conditions.
All reactions ran at 95°C for 2 min, followed by 40 cycles at 94°C
for 15 sec and 60°C for 1 min and extension 75°C for 30 sec. The
relative mRNA and miRNA expression levels were quantified by
calculating the ΔCq value: Cq value (target gene)-Cq value
(housekeeping gene). The relative expression levels were calculated
using the 2-∆∆Cq method (17).
Bioinformatics analysis to predict
target genes for miR-155
The present study used bioinformatics tools to
predict target genes for miR-155, including miRanda (omictools.com/miranda-tool), TargetScan
(www.targetscan.org), miRBase (www.mirbase.org) and PicTar (pictar.mdc-berlin.de). miR-155 was determined to be
closely associated with the PI3K/Akt/mTOR signaling pathway. The
gene PIK3CA, encoding the p110 catalytic protein subunit of PI3K,
as well as Rheb, were identified as potential target genes of
miR-155.
Plasmid construction and dual
luciferase reporter assay
Human miR-155-5p overexpression plasmid, negative
control plasmid, PIK3CA, and Rheb 3′-untranslated region (UTR)
reporter plasmids were designed and purchased from Shanghai
Genechem Co., Ltd. (Shanghai, China). PIK3CA-wild type (wt),
PIK3CA-mutant (mut), Rheb-wt and Rheb-mut reporters were
successfully constructed using molecular cloning technology. For
the construction of the PIK3CA- and Rheb-expressing plasmid, the
3′-UTRs of PIK3CA and Rheb, which contained the putative miR-155
binding sites, were amplified using PCR (conducted by Shanghai
Genechem Co., Ltd.) and the product inserted into pGL3 luciferase
plasmids at the NotI and XhoI restriction sites. The
reporter vector was termed wt. A point mutation (mut) was
incorporated into the binding sites of the 3′-UTR in the PIK3CA and
Rheb genes to generate a mutant reporter vector. 293T cells grown
in 24-well plates were co-transfected using Lipofectamine 2000 with
10 nM negative controls or miR-155 mimics, and 500 ng
dual-luciferase reporter plasmid per well, which was either
PIK3CA-wt, PIK3CA-mut, Rheb-wt, Rheb-mut or empty vector.
Renilla relative luciferase activity was measured 48 h
post-transfection using a dual-luciferase reporter assay system
according to the manufacturer's protocol (Promega Corporation,
Madison, WI, USA). Each transfection was repeated three times.
Statistical analysis
Statistical analyses were performed with SPSS
software version 16.0 (SPSS, Inc., Chicago, IL, USA). Differences
between groups were assessed using one-way analysis of variance
followed by Fisher's Least Significant Difference for multiple
comparisons. The results were expressed as the mean ± standard
deviation of three independent experiments. P<0.05 was
considered to indicate a statistically significant difference.
Results
Overexpression of miR-155 induces
autophagy
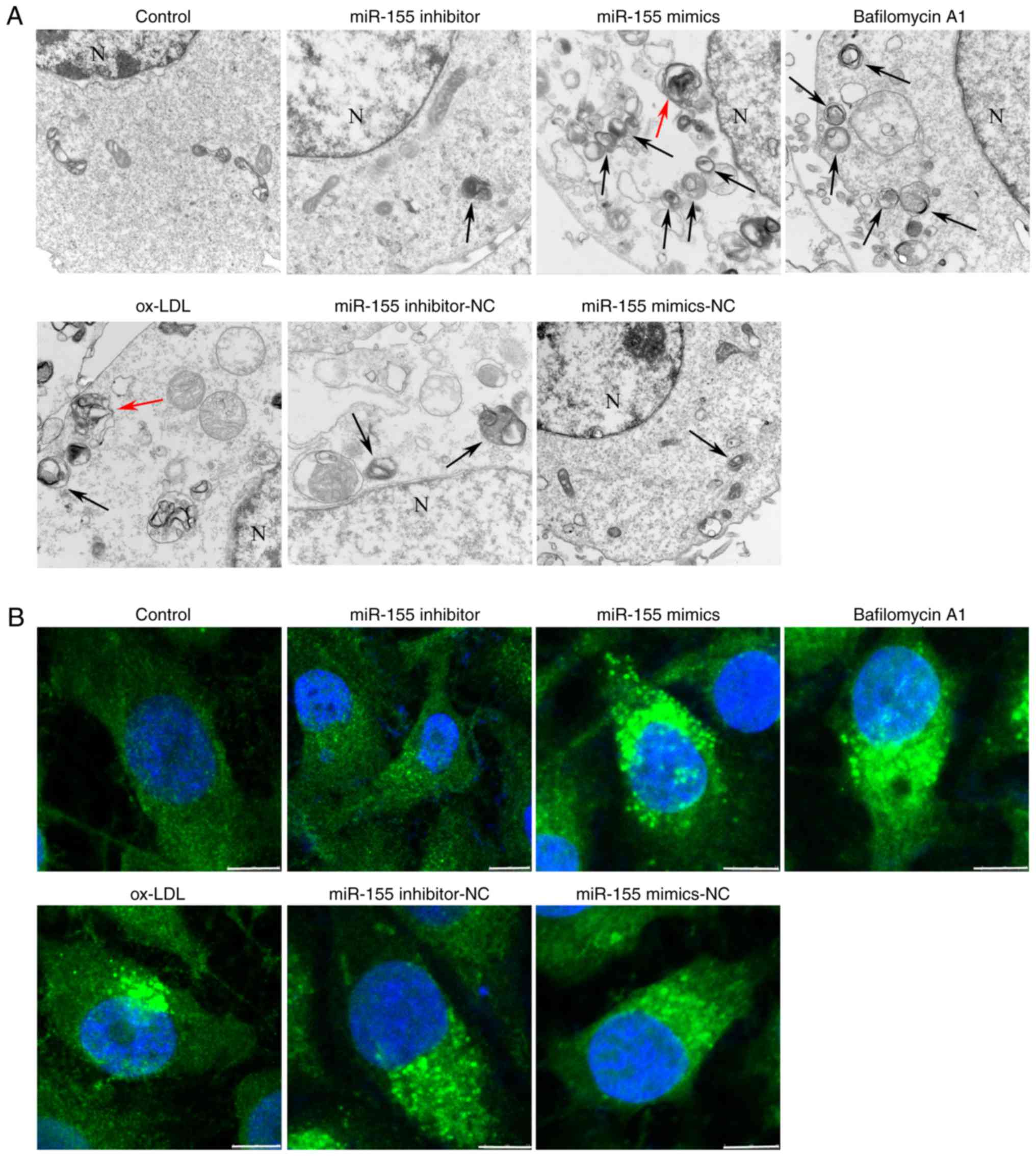
To explore the role of miR-155 in autophagy, the
present study used TEM to evaluate autophagosome and autolysosome
accumulation. The TEM results demonstrated that the average number
of autophagic vacuoles and autolysosomes in the control group was
2–3. It was observed that inhibition of the expression of miR-155
with miR-155 inhibitors resulted in the suppressed formation of
autophagic vacuoles and autolysosomes, compared with the control
mimic (NC) group. The average number of autophagic vacuoles and
autolysosomes was 0–1 in the miR-155 inhibitor group. In the
miR-155 mimic groups, cells displayed a higher number of
autolysosomes and autophagic vesicles than the NC group, and the
average number of autolysosomes and autophagic vacuoles was 5–7
(Fig. 1A). Following this, laser
confocal microscopy was performed to detect LC3 puncta accumulation
in ox-LDL-stimulated HUVECs. LC3 puncta appear in the cytoplasm and
reflect the recruitment of LC3 proteins to autophagosomes (18). The fluorescence intensity of LC3
was reduced in miR-155 inhibitor-transfected cells, compared with
NC cells. Conversely, transfection of HUVECs with miR-155 mimics
induced an increase in the fluorescence intensity, compared with NC
cells (Fig. 1B). To further
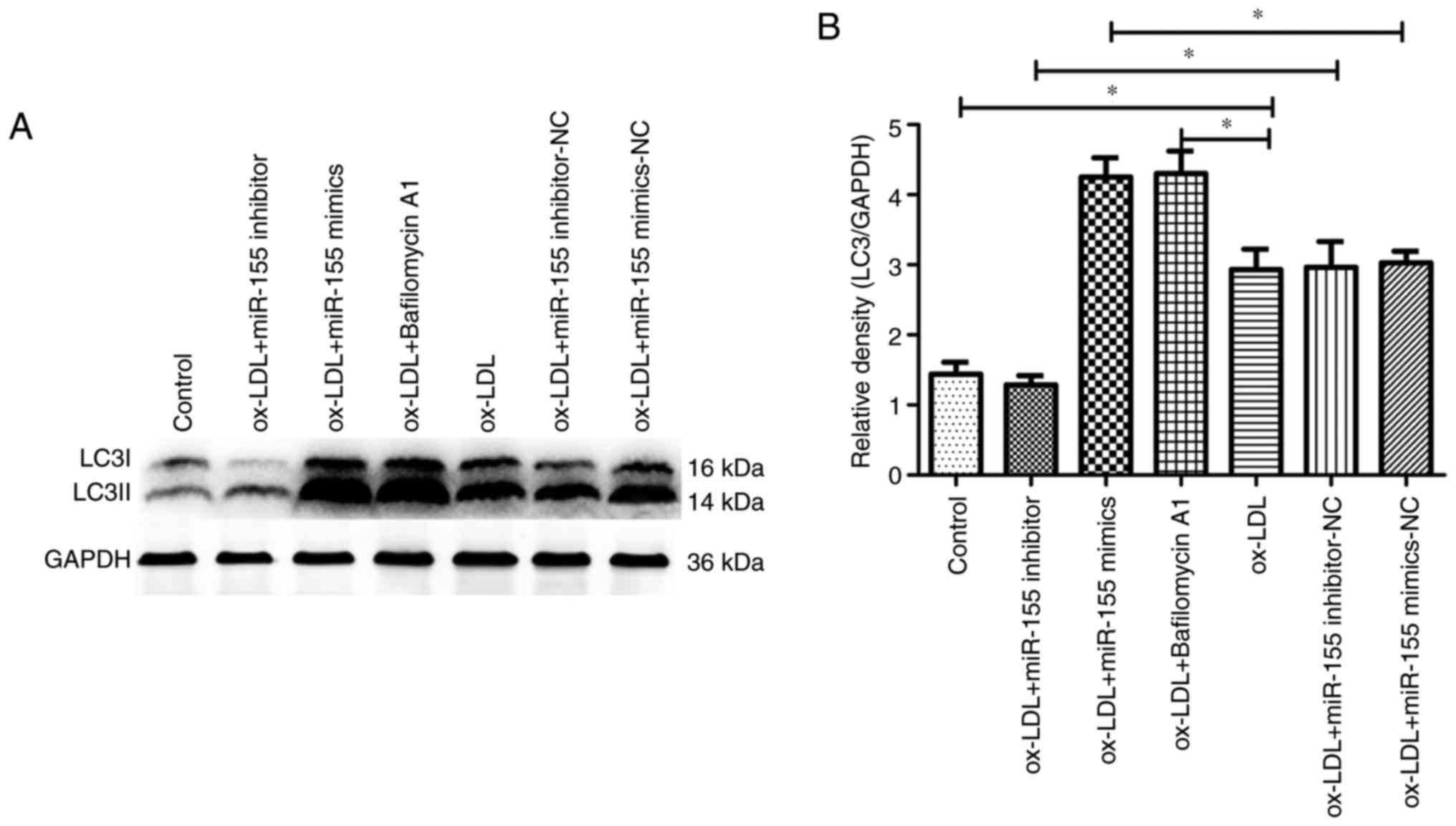
confirm the promotion of autophagy by miR-155, the conversion of
LC3-I to LC3-II (via the ratio of LCII to LC1) was examined through
western blotting (Fig. 2A). It was
observed that LC3-II expression levels were higher in the
miR-155-mimic group compared with the control group, and that these
autophagic markers were inhibited in HUVECs transfected with
miR-155-inhibitors, compared with the control group (Fig. 2B), which was in accordance with
results from electron microscopy and confocal microscopy.
Lysomotropic bafilomycin A1 prevents lysosome and autophagosome
fusion, and is often used for measurement of autophagic flux
(19). When cells were treated
with bafilomycin A1, autophagic activity was significantly
increased compared with the ox-LDL group, and the average number of
autolysosomes and autophagic vacuoles was 5–6 (Figs. 1 and 2). Taken together, these results
suggested that miR-155 efficiently promoted autophagy in vascular
endothelial cells.
Overexpression of miR-155 promotes
autophagy by suppressing the activation of the PI3K/Akt/mTOR
signaling pathway in HUVECs
The efficiency of miR-155 transfection in HUVECs was
examined. RT-qPCR data revealed that miR-155 expression was
significantly increased in the miR-155 mimic group, compared with
the corresponding negative control group. Furthermore, miR-155
levels were significantly decreased compared with the corresponding
negative control group following miR-155 inhibitor transfection
(Fig. 3A). The results were
repeatedly verified as stable following the exclusion of
interference from other factors under the same conditions, such as
the transfection reagent and transfection conditions. Our previous
study achieved similar results (16). To investigate the role of the
PI3K/Akt/mTOR signaling pathway and miR-155 in HUVECs, RT-qPCR and
western blotting was performed to detect the mRNA and protein
expression levels of key members of the PI3K/Akt/mTOR signaling
pathway. The results demonstrated that overexpression of miR-155
inhibited Akt (Fig. 3B) and p70s6k
(Fig. 3C) mRNA expression,
compared with the controls. Furthermore, Akt and p70s6k mRNA
expression significantly increased in the miR-155 inhibitor group.
In addition, western blot analysis was performed to determine the
relative protein expression of total/p-Akt and total/p-p70s6k
(Fig. 3D). Densitometry confirmed
that similar, significant effects were observed in the levels of
Akt (Fig. 3E) and p70s6k (Fig. 3F) activity in response to miR-155
mimics and inhibitors. Therefore, the data verified that miR-155
promoted autophagy by negatively regulating the expression
PI3K/Akt/mTOR signaling pathway proteins in HUVECs.
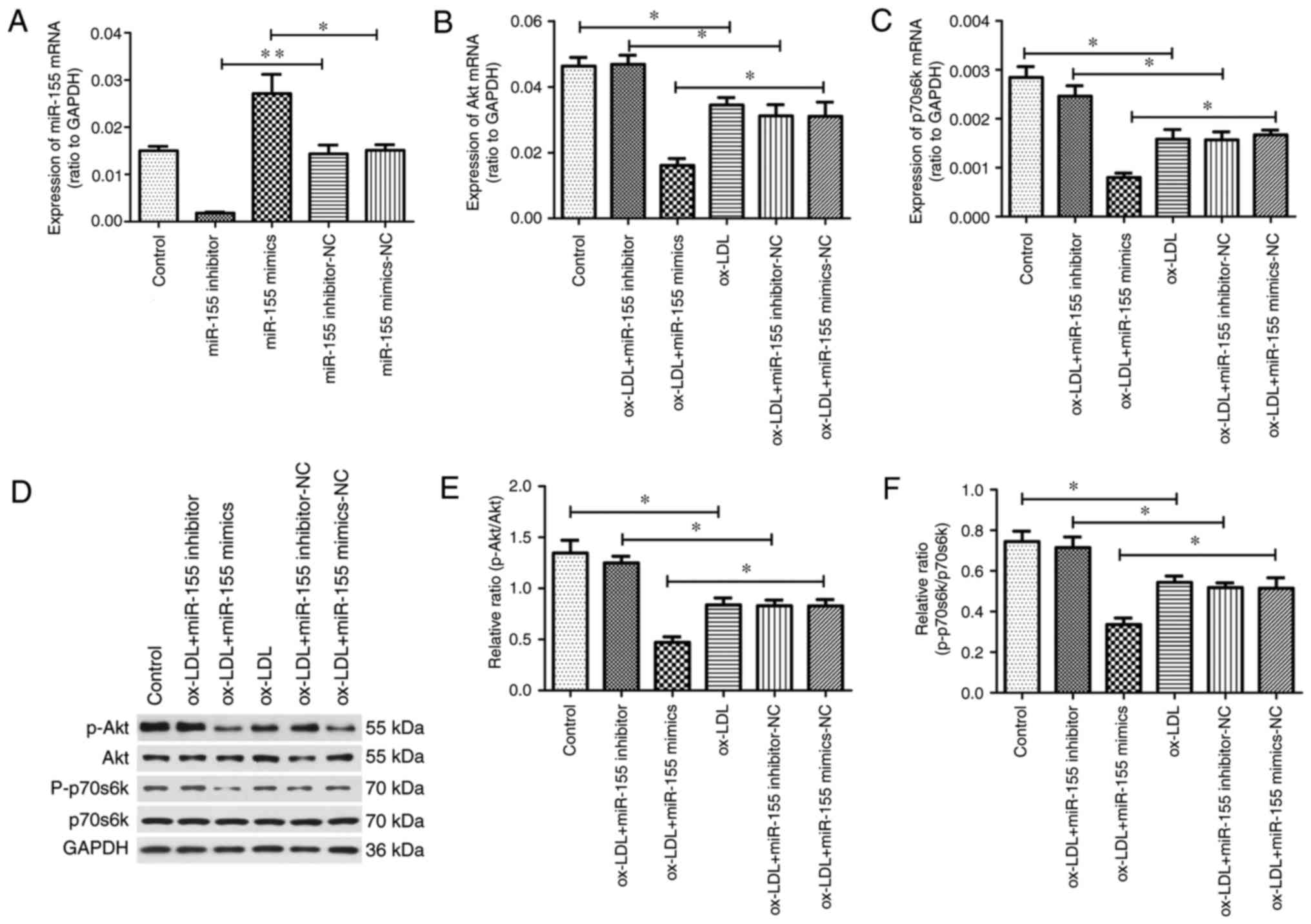 | Figure 3.miR-155 suppresses the activation of
the PI3K/Akt/mTOR signaling pathway. RT-qPCR was performed to
analyze (A) miR-155 (U6 as a normalization internal control), (B)
Akt and (C) p70s6k expression. (D) Representative image of western
blot analysis to determine total Akt, p-Akt, p70s6k, p-p70s6k and
GAPDH protein expression in ox-LDL-treated human umbilical vein
endothelial cells transfected with miR-155 mimics, inhibitors or
NCs. Quantification of western blotting results as a relative ratio
of (E) p-Akt/Akt and (F) p-p70s6k/p70s6k expression. Data are
expressed as the mean ± standard deviation (n=3). *P<0.05;
**P<0.01. miR-155, microRNA-155; ox-LDL, oxidized low-density
lipoprotein; NC, negative control; p, phosphorylated; Akt, RAC-α
serine/threonine-protein kinase; p70s6k, 70 kDa ribosomal protein
S6 kinase 1. |
miR-155 promotes autophagy by direct
interaction with the 3′-UTRs of PIK3CA and Rheb
An association between miR-155 and the PI3K/Akt/mTOR
signaling pathway was demonstrated in the present study; however,
the present study aimed to elucidate the mechanism underlying this.
Bioinformatics analyses were performed to predict potential target
genes for miR-155. PIK3CA and Rheb were identified due to the
existence of the evolutionarily conserved miR-155 binding site in
the 3′-UTR of these genes (Fig.
4A). In order to confirm these genes as targets of miR-155, a
luciferase reporter plasmid was constructed that contained the
human PIK3CA and Rheb mRNA 3′-UTR binding sites, downstream of the
firefly luciferase reporter gene (the reporter vector was named
wild-type). A point mutation was also incorporated into the PIK3CA
and Rheb gene 3′-UTR binding sites to generate a mutant reporter
vector.
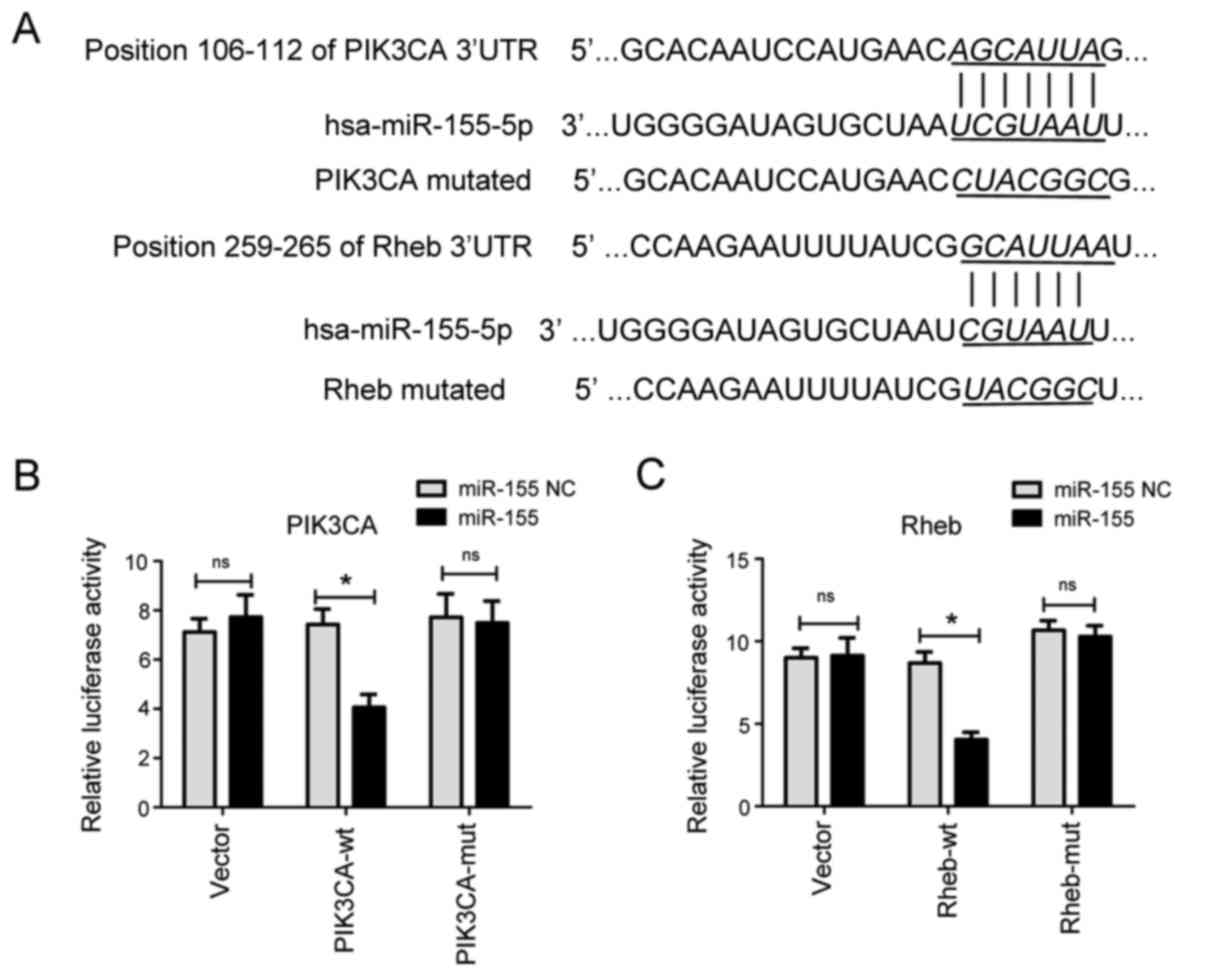 | Figure 4.miR-155 targets PIK3CA and Rheb. (A)
The predicted binding site of miR-155 in the 3′-UTR of PIK3CA and
Rheb. 293T cells were transfected with a reporter vector containing
the wt or mut (B) PIK3CA and (C) Rheb 3′-UTR, along with NC or
miR-155 mimics. Firefly luciferase activity was normalized to
Renilla luciferase. The results are presented as the mean ±
standard deviation, according to three independent experiments, n=3
*P<0.05. miR-155, microRNA-155; NC, negative control; UTR,
untranslated region; wt, wild-type; mut, mutant; Rheb, Ras homolog
enriched in brain; PIK3CA, phosphatidylinositol-3 kinase subunit
α. |
miR-155 mimic or NC was transfected with 500 ng
wild-type or mutated reporter plasmids. Co-expression of PIK3CA or
Rheb 3′-UTR constructs with miR-155 mimics was established in 293
cells. Luciferase activity was determined 24 h post-transfection.
The luciferase reporter assay indicated that miR-155 significantly
reduced the relative luciferase activity of the reporter vector
containing the wild-type 3′-UTR of PIK3CA (Fig. 4B) or Rheb (Fig. 4C), compared with the negative
control. Conversely, miR-155 did not significantly inhibit
luciferase activity in cells expressing mutant PIK3CA and Rheb
3′-UTR, or empty vector. These results suggested that miR-155
directly bound to the 3′-UTR of PIK3CA and Rheb, and inhibited
luciferase activity. Therefore, PIK3CA and Rheb were verified as
target genes of miR-155.
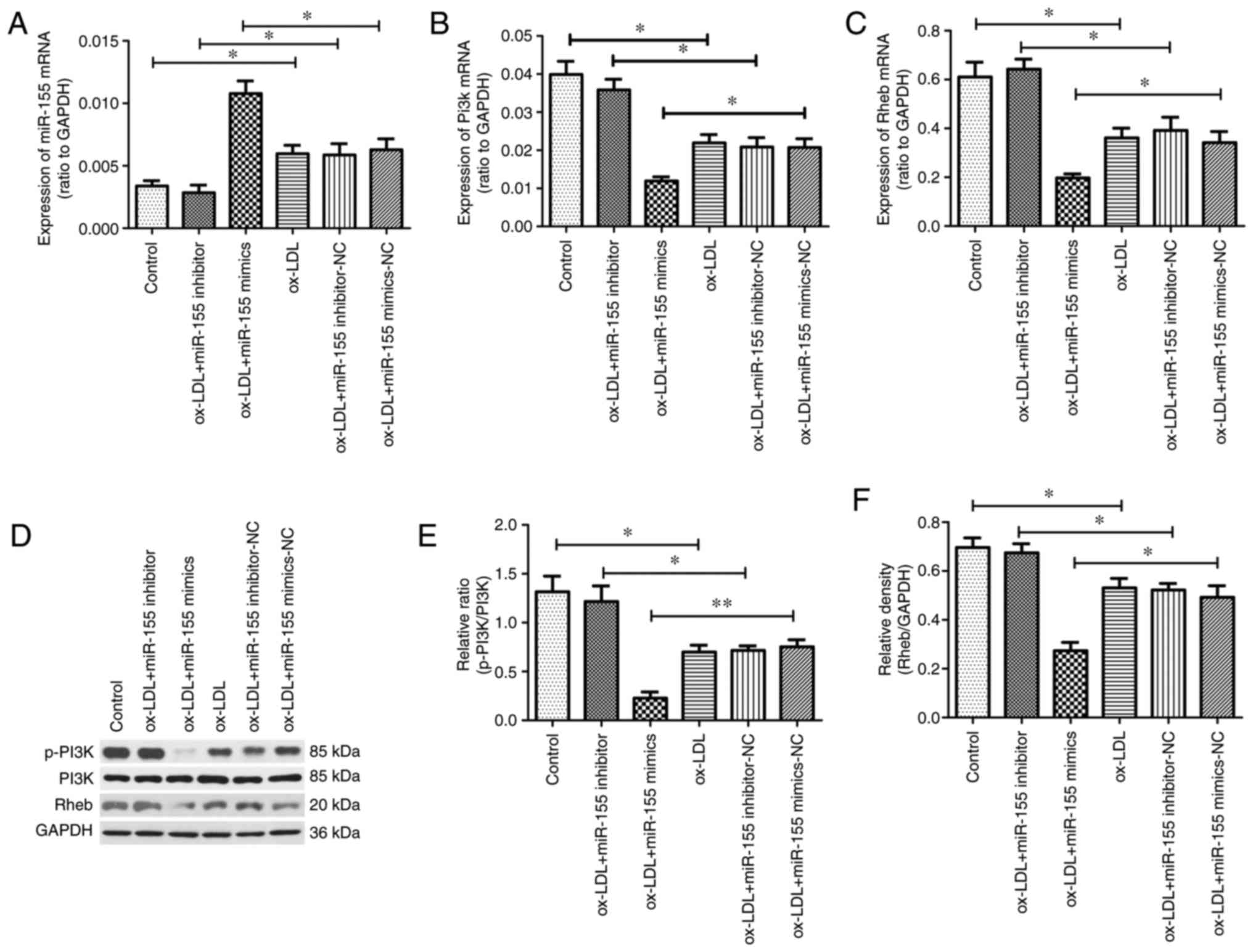
miR-155 expression was subsequently detected in
cells stimulated with ox-LDL. Ox-LDL treatment alone significantly
increased miR-155 expression compared with the control. In
addition, treatment with ox-LDL and miR-155 mimics resulted in
significantly higher miR-155 expression, compared with the ox-LDL
and miR-155 mimic NC group (Fig.
5A). In addition, the levels of PI3K (Fig. 5B) and Rheb (Fig. 5C) mRNA were significantly
downregulated in HUVECs transfected with miR-155 mimics compared
with the corresponding NC group. However, in cells stimulated with
ox-LDL, miR-155 inhibitor transfection increased PI3K and Rheb mRNA
expression to levels comparable with the control group.
Furthermore, western blot analysis was performed to detect the
protein expression of Rheb, PI3K and p-PI3K (Fig. 5D). Transfection of miR-155 mimics
suppressed the protein levels of p-PI3K (Fig. 5E) and Rheb (Fig. 5F) compared with the control group;
however, miR-155 inhibitors did not result in a significant
reduction in protein levels. In addition, total PI3K expression
remained unchanged. The different effects observed at the mRNA and
proteins were thought to be due to two potential reasons. It may be
a dynamic effect; one mRNA converts to protein more efficiently
within a certain time, so protein downregulation is not
significant. On the other hand, this may be due to a negative
feedback loop in the cell. In this process, following
miRNA-mediated protein downregulation, the cell may increase mRNA
production due to the cell requirements, or other regulatory
processes (20–25). These results verified that miR-155
suppressed the expression of PI3K and Rheb in HUVECs. Therefore, it
is possible that miR-155 regulated autophagy by targeting PI3K and
Rheb in atherosclerosis.
Discussion
The present study explored the underlying mechanisms
of miR-155-regulated autophagy in vascular endothelial cells, using
an in vitro model of HUVECs stimulated with ox-LDL. The
primary findings of the current study were that: i) Overexpression
of miR-155 upregulated autophagic activity and inhibition of
miR-155 expression reduced autophagic activity; ii) overexpression
of miR-155 promoted autophagy by suppressing the activation of the
PI3K/Akt/mTOR signaling pathway; and iii) dual luciferase reporter
assays confirmed PIK3CA and Rheb as target genes of miR-155.
Therefore, it was established that miR-155-mediated inhibition of
PI3K/Akt/mTOR signaling regulated autophagy, and that this occurred
through targeting of PI3K and Rheb. This mechanism may be involved
in the development of atherosclerosis.
miR-155 is one of the most extensively studied
miRNAs, and has been demonstrated to be involved in regulating the
cellular autophagy process. Chen et al (26) reported that an increase in miR-155
expression levels upregulates anticancer drug-induced autophagy in
osteosarcoma cells. In addition, Liu et al (27) suggested that miR-155 alleviates
septic lung injury by inducing autophagy via inhibition of TGF-β
activated kinase 1 (MAP3K7) binding protein 2. The results of the
present study demonstrated that overexpression of miR-155
significantly increased autophagic activity in ox-LDL-induced
HUVECs; however, suppression of miR-155 expression inhibited
autophagic activity. These results implied that miR-155 may have an
important role in the regulation of vascular endothelial cell
autophagy, which makes miR-155 a potential target in
atherosclerosis therapy.
PI3K/Akt/mTOR signaling is regarded as a classic
autophagic signaling pathway, which transmits signals from the cell
membrane to the nucleus and activates multiple cellular events
(28). Previously, the
PI3K/Akt/mTOR signaling pathway has been reported to serve an
important role in the regulation of autophagy in atherosclerosis
(9,10). PI3K activates the phosphorylation
of Akt, followed by the activation of mTOR to finally trigger the
inhibition of autophagy. P70s6k is a key downstream target of mTOR,
and an essential target for the regulation of protein translation
and additional metabolic processes in cell growth (29). Activation of mTOR complex 1
triggers a cascade of anabolic processes for cell growth,
proliferation and autophagy, primarily mediated by p70s6k (30). To verify whether miR-155 regulates
autophagy via the PI3K/Akt/mTOR pathway, the present study
performed RT-qPCR and western blot analysis in ox-LDL-stimulated
HUVECs to determine the expression of key proteins of the
PI3K/Akt/mTOR signaling pathway, including Akt, p-Akt, p70s6k and
p-p70s6k. It was demonstrated that Akt and p70s6k expression was
repressed when miR-155 mimics were transfected into ox-LDL-treated
HUVECs; however, inhibition of miR-155 increased the activation of
Akt and p70s6k. The results of the present study therefore revealed
that miR-155 downregulated Akt and p70s6k at both the mRNA and
protein level in HUVECs. The PI3K/Akt/mTOR signaling pathway
negatively regulates autophagy (31), as evidenced by the upregulation of
autophagy induced by the inhibition of Akt and p70s6k in the preset
study. This suggests that miR-155 promotes autophagy by suppressing
the PI3K/Akt/mTOR pathway, and this may be involved in the
regulation of atherosclerosis.
The present study focused on the PI3K/Akt/mTOR
signaling pathway in atherosclerosis, and investigated whether
miR-155 regulated autophagy in vascular endothelial cells by
targeting key proteins of the PI3K/Akt/mTOR signaling pathway.
Bioinformatics tools predicted that PIK3CA and downstream protein
Rheb were potential target genes of miR-155. A dual luciferase
reporter assay demonstrated that miR-155 downregulated PIK3CA and
Rheb by directly targeting the 3′-UTR region of these genes. The
inhibitory effect of miR-155 on the expression of PI3K and Rheb
protein and mRNA was also analyzed by western blot analysis and
RT-qPCR assays, respectively. These results again revealed the role
of miR-155 in the PI3K/Akt/mTOR pathway, and suggested that PI3K
and Rheb are direct targets of miR-155 that may be involved in the
pathogenesis of atherosclerosis. Huang et al (32) observed that miR-155 activates the
PI3K-Akt signaling pathway by targeting PI3K regulatory subunit
p85α in diffuse large B-cell lymphoma. A study by Wan et al
(33) suggested that
hypoxia-induced expression of miR-155 occurs in human cervical
cancer and nasopharyngeal cancer cells, which suppresses the
expression of multiple target genes in the mTOR signaling pathway,
including Rheb, RPTOR independent companion of MTOR complex 2 and
ribosomal protein S6 kinase B2. In addition, knockdown of
endogenous miR-155 expression inhibits hypoxia-induced autophagy
(33). Wang et al (34) demonstrated that miR-155 promotes
autophagy by targeting Rheb to eliminate intracellular
mycobacteria. These studies provide evidence that miR-155 activates
autophagy through the PI3K/Akt/mTOR signaling pathway. Further
study is required to determine whether other autophagy-associated
pathways also participate in this process.
In conclusion, the present study demonstrated that
miR-155 downregulated the activity of the PI3K/Akt/mTOR signaling
pathway via targeting PI3K and Rheb, to promote autophagy in
ox-LDL-induced HUVECs. Therefore, miR-155 may be a potential
therapeutic target in atherosclerosis. The present study identified
a novel role for miR-155 in the regulation of endothelial cell
autophagy during atherosclerosis, and provided insight into the
future development of innovative atherosclerosis therapies that may
function by targeting miR-155.
Acknowledgements
We sincerely thank the staff of Central Laboratory
of the Affiliated Hospital of Qingdao University (Qingdao, China)
and all members of our research group for general support.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81571112 and
81641046), the Shandong Province Natural Science Foundation (grant
no. 2015GSF118172) and the Municipal Scientific and Technological
Project of Qingdao City (grant no. 15-9-2-86-nsh).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XP, AM and SYa designed the experiments; SYi, JM,
HPe YD, SL, WL and XB performed experiments; SYi and AM analyzed
the data; SYi and SYa wrote the paper.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Affiliated Hospital of Qingdao University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhong LX, Zhang Y, Wu ML, Liu YN, Zhang P,
Chen XY, Kong QY, Liu J and Li H: Resveratrol and STAT inhibitor
enhance autophagy in ovarian cancer cells. Cell Death Discov.
2:150712016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tang F, Yang TL, Zhang Z, Li XG, Zhong QQ,
Zhao TT and Gong L: MicroRNA-21 suppresses ox-LDL-induced human
aortic endothelial cells injuries in atherosclerosis through
enhancement of autophagic flux: Involvement in promotion of
lysosomal function. Exp Cell Res. 359:374–383. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ding Z, Liu S, Wang X, Dai Y, Khaidakov M,
Romeo F and Mehta JL: LOX-1, oxidant stress, mtDNA damage,
autophagy, and immune response in atherosclerosis. Can J Physiol
Pharmacol. 92:524–530. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ding WX: Uncoupling AMPK from autophagy: A
foe that hinders the beneficial effects of metformin treatment on
metabolic syndrome-associated atherosclerosis? Focus on ‘glucose
and palmitate uncouple AMPK from autophagy in human aortic
endothelial cells’. Am J Physiol Cell Physiol. 308:C246–C248. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shafique E, Choy WC, Liu Y, Feng J,
Cordeiro B, Lyra A, Arafah M, Yassin-Kassab A, Zanetti AV, Clements
RT, et al: Oxidative stress improves coronary endothelial function
through activation of the pro-survival kinase AMPK. Aging (Albany
NY). 5:515–530. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Perrotta I and Aquila S: The role of
oxidative stress and autophagy in atherosclerosis. Oxid Med Cell
Longev. 2015:1303152015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mollace V, Gliozzi M, Musolino V, Carresi
C, Muscoli S, Mollace R, Tavernese A, Gratteri S, Palma E, Morabito
C, et al: Oxidized LDL attenuates protective autophagy and induces
apoptotic cell death of endothelial cells: Role of oxidative stress
and LOX-1 receptor expression. Int J Cardiol. 184:152–158. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Butler DE, Marlein C, Walker HF, Frame FM,
Mann VM, Simms MS, Davies BR, Collins AT and Maitland NJ:
Inhibition of the PI3K/AKT/mTOR pathway activates autophagy and
compensatory Ras/Raf/MEK/ERK signalling in prostate cancer.
Oncotarget. 8:56698–56713. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jiang Y, Kou J, Han X, Li X, Zhong Z, Liu
Z, Zheng Y, Tian Y and Yang L: ROS-dependent activation of
autophagy through the PI3K/Akt/mTOR pathway is induced by
hydroxysafflor yellow a-sonodynamic therapy in THP-1 macrophages.
Oxid Med Cell Longev. 2017:85191692017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhai C, Cheng J, Mujahid H, Wang H, Kong
J, Yin Y, Li J, Zhang Y, Ji X and Chen W: Selective inhibition of
PI3K/Akt/mTOR signaling pathway regulates autophagy of macrophage
and vulnerability of atherosclerotic plaque. PLoS One.
9:e905632014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gupta R, Arkatkar T, Keck J, Koundinya GK,
Castillo K, Hobel S, Chambers JP, Yu JJ, Guentzel MN, Aigner A, et
al: Antigen specific immune response in Chlamydia muridarum genital
infection is dependent on murine microRNAs-155 and −182.
Oncotarget. 7:64726–64742. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang T, Tian F, Wang J, Jing J, Zhou SS
and Chen YD: Endothelial cell autophagy in atherosclerosis is
regulated by miR-30-mediated translational control of ATG6. Cell
Physiol Biochem. 37:1369–1378. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang B, Zhong Y, Huang D and Li J:
Macrophage autophagy regulated by miR-384-5p-mediated control of
Beclin-1 plays a role in the development of atherosclerosis. Am J
Transl Res. 8:606–614. 2016.PubMed/NCBI
|
|
14
|
Ouimet M, Ediriweera H, Afonso MS,
Ramkhelawon B, Singaravelu R, Liao X, Bandler RC, Rahman K, Fisher
EA, Rayner KJ, et al: microRNA-33 regulates macrophage autophagy in
atherosclerosis. Arterioscler Thromb Vasc Biol. 37:1058–1067. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chistiakov DA, Orekhov AN and Bobryshev
YV: Chemokines and relevant microRNAs in the atherogenic process.
Mini Rev Med Chem. 18:597–608. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang Z, Pan X, Yang S, Ma A, Wang K, Wang
Y, Li T and Liu S: miR-155 promotes ox-LDL-induced autophagy in
human umbilical vein endothelial cells. Mediators Inflamm.
2017:91748012017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang L, Chen M, Yang J and Zhang Z: LC3
fluorescent puncta in autophagosomes or in protein aggregates can
be distinguished by FRAP analysis in living cells. Autophagy.
9:756–769. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lee WK, Probst S, Santoyo-Sánchez MP,
Al-Hamdani W, Diebels I, von Sivers JK, Kerek E, Prenner EJ and
Thévenod F: Initial autophagic protection switches to disruption of
autophagic flux by lysosomal instability during cadmium stress
accrual in renal NRK-52E cells. Arch Toxicol. 91:3225–3245. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao Q, Liu H, Yao C, Shuai J and Sun X:
Effect of dynamic interaction between microRNA and transcription
factor on gene expression. Biomed Res Int. 2016:26762822016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Olaru AV, Selaru FM, Mori Y, Vazquez C,
David S, Paun B, Cheng Y, Jin Z, Yang J, Agarwal R, et al: Dynamic
changes in the expression of MicroRNA-31 during inflammatory bowel
disease-associated neoplastic transformation. Inflamm Bowel Dis.
17:221–231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
O'Donnell KA, Wentzel EA, Zeller KI, Dang
CV and Mendell JT: c-Myc-regulated microRNAs modulate E2F1
expression. Nature. 435:839–843. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yamakuchi M and Lowenstein CJ: MiR-34,
SIRT1 and p53: The feedback loop. Cell Cycle. 8:712–715. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu H, Lin H, Zhang L, Sun Q, Yuan G,
Zhang L, Chen S and Chen Z: miR-145 and miR-143 regulate
odontoblast differentiation through targeting Klf4 and Osx genes in
a feedback loop. J Biol Chem. 288:9261–9271. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wei X, Cheng X, Peng Y, Zheng R, Chai J
and Jiang S: STAT5a promotes the transcription of mature
mmu-miR-135a in 3T3-L1 cells by binding to both miR-135a-1 and
miR-135a-2 promoter elements. Int J Biochem Cell Biol. 77:109–119.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen L, Jiang K, Jiang H and Wei P:
miR-155 mediates drug resistance in osteosarcoma cells via inducing
autophagy. Exp Ther Med. 8:527–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu F, Nie C, Zhao N, Wang Y, Liu Y, Li Y,
Zeng Z, Ding C, Shao Q, Qing C, et al: MiR-155 alleviates septic
lung injury by inducing autophagy via inhibition of transforming
growth factor-β-activated binding protein 2. Shock. 48:61–68. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zeng S, Song H, Chen Y, Xie W and Zhang L:
B7-H4-mediated immunoresistance is supressed
by PI3K/Akt/mTOR pathway inhibitors. Mol Biol.
50:887–894. 2016. View Article : Google Scholar
|
|
29
|
Cheng KY and Hao M: Mammalian target of
rapamycin (mTOR) regulates transforming growth factor-β1
(TGF-β1)-induced epithelial-mesenchymal transition via decreased
pyruvate kinase M2 (PKM2) expression in cervical cancer cells. Med
Sci Monit. 23:2017–2028. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nivon M, Richet E, Codogno P, Arrigo AP
and Kretz-Remy C: Autophagy activation by NFkappaB is essential for
cell survival after heat shock. Autophagy. 5:766–783. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shintani T and Klionsky DJ: Autophagy in
health and disease: A double-edged sword. Science. 306:990–995.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang X, Shen Y, Liu M, Bi C, Jiang C,
Iqbal J, McKeithan TW, Chan WC, Ding SJ and Fu K: Quantitative
proteomics reveals that miR-155 regulates the PI3K-AKT pathway in
diffuse large B-cell lymphoma. Am J Pathol. 181:26–33. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wan G, Xie W, Liu Z, Xu W, Lao Y, Huang N,
Cui K, Liao M, He J, Jiang Y, et al: Hypoxia-induced MIR155 is a
potent autophagy inducer by targeting multiple players in the MTOR
pathway. Autophagy. 10:70–79. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang J, Yang K, Zhou L, Minhaowu, Wu Y,
Zhu M, Lai X, Chen T, Feng L, Li M, et al: MicroRNA-155 promotes
autophagy to eliminate intracellular mycobacteria by targeting
Rheb. PLoS Pathog. 9:e10036972013. View Article : Google Scholar : PubMed/NCBI
|