Introduction
Doxorubicin (DOX) is one of the most effective
chemotherapy drugs for the treatment of hematological malignancies
and several solid tumors including lymphomas, leukemia and breast
cancer (1). However, DOX has
limited clinical use as it induces severe cardiotoxicity, that
leads to irreversible cardiac injuries such as heart failure and
left ventricular dysfunction (2).
DOX has a high affinity for binding to phospholipids in cardiolipin
in the inner mitochondrial membrane (3). Therefore, the heart is susceptible to
DOX-associated injury as it contains a greater number of
mitochondria and fewer antioxidative enzymes than other organs. The
mechanism underlying DOX-induced cardiotoxicity is multifactorial
(4). It is primarily associated
with the production of reactive oxygen species (ROS), apoptosis of
cardiac cells, mitochondrial damage, impairment of cardiac energy
metabolism and defects in the adenosine monophosphate-activated
protein kinase (AMPK) signaling pathway (5,6).
Cordyceps sinensis (CS), a widely used
traditional Chinese medicine (TCM), is the dried stroma and
sporophore of Cordyceps sinensis (Berk.) Sacc. Which
parasitizes on the larvae of Hepialidae. CS has been used
extensively in TCM to treat coughs, shortness of breath and to
improve quality of life. It is now commonly used as a health
product in China (7), as it has
broad pharmacological effects associated with anti-oxidative stress
(OS), immunoregulatory, antisenescent, hypoglycemic, hypolipidemic
and antitumor activities (8). As
the primary sources of CS are restricted by the controlled natural
conditions required, only an extremely limited amount of natural CS
can be produced each year. Thus, it is very expensive and so, it is
not widely available to general patients. Fermented CS, a
substitute for natural CS, is produced by purifying and
artificially fermenting the fungus isolated from fresh Qinghai CS.
Fermented CS has been used extensively in supplementing medical
treatments in traditional Chinese medicine, particularly in the
adjuvant therapy of cardiac arrhythmia and chronic renal
insufficiency (4). The chemical
components and pharmacological effects of fermented CS are similar
to those of natural CS, including anti-hyperglycemic actions
(9), antitumor activity (10), anti-inflammation (11) and anti-OS (4).
Therefore, the aim of the study was to evaluate the
protective effects of fermented CS against DOX-induced
cardiotoxicity and the potential underlying mechanisms. The results
suggested that fermented CS attenuated DOX-induced cardiotoxicity
in rats by reducing mitochondrial damage and oxidative stress,
depressing phosphodiesterase activities, upregulating the AMPK and
cyclic (c)AMP signaling pathways, and ameliorating energy
metabolism, myocardial damage, cardiac dysfunction and acute heart
failure.
Materials and methods
Herbal materials and reagents
Fermented CS is produced by artificial fermentation,
and CS mycelia, the raw material, is isolated from natural CS.
Fermented CS is listed in the Pharmacopoeia of the People's
Republic of China (2015 edition) (12), its chemical components are similar
to those of natural CS (4).
Fermented CS is produced by purifying and artificially fermenting
the fungus isolated from fresh Qinghai CS. It was supplied by
Hangzhou Zhongmei Huadong Pharmaceutical Co., Ltd. (batch no.
1410170; Zhejiang, China). Fermented CS contains 14.8% protein,
39.4% carbohydrate, 6.63% fat, 2.95% ash, 1.4% cordycepin, 6.4%
H2O, 9.23% amino acid and 0.21% adenosine. The adenosine
content in fermented CS was analyzed by high-performance liquid
chromatography (HPLC; UltiMate 3000 HPLC system; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). The mobile phase consisted of
acetonitrile and water (containing 0.04 mol/l
KH2PO4). A linear gradient elution program
was applied as follows: 0–15 min 100% water, 15–45 min 0–15%
acetonitrile, 100–85% water. The flow rate was 1.0 ml/min and the
injection volume was 2 µl. The reference substance of adenosine and
fermented CS were individually dissolved in 0.5% phosphoric acid to
obtain a solution of 12 µg/ml. The detection wavelength was 223 nm
and the chromatographic column used was the Ultimate AQ-C18
(4.6×150 mm, 3 µm, Welch Materials, Inc., Shanghai, China) with the
injection volume of 10 µl with 0.2 µm precolumn inline filter
maintained at 30°C. The adenosine content in 0.5 g fermented CS was
quantitated as 1.07 mg, which was in accordance with the
requirements of the Pharmacopoeia of People's Republic of China
(2015 edition). The DOX Hydrochloride injection was obtained from
Shenzhen Main Luck Pharmaceuticals Inc. (Guangdong, China).
Captopril was supplied by Shanghai Hengshan Pharmaceuticals Co.,
Ltd. (Shanghai, China).
Animals
Male Sprague-Dawley rats (n=84; weight, 180–200 g; 8
weeks old) were supplied by Shanghai SLAC Laboratory Animal Co.,
Ltd. (Shanghai, China). Rats were kept in the Laboratory Animal
Center of Shanghai Traditional Chinese Medicine (Shanghai, China;
temperature 22–24°C, humidity 40±5% and 12 h light/dark cycle) with
free access to food and water. All animals were used in accordance
with the guidelines of the National Institutes of Health (NIH
Publication no. 85-23, revised 1996) Guide for the Care and Use of
Laboratory Animals (13), and were
approved by the Animal Care and Use Committee of Shanghai
University of Traditional Chinese Medicine (approval no.
SZY2014029).
Experimental design
A total of 84 rats were randomly assigned to 7
groups which included the normal control group (n=10), the DOX
control group (n=15), the DOX+captopril (0.05 g/kg) group (n=13),
the 3 DOX+CS groups [0.75 (n=13), 1.5 (n=14) and 3 g/kg (n=14) of
CS], and the CS (1.5 g/kg) control group (n=5).
Rats in the DOX control group, and the captopril and
CS treatment groups were administered 6 doses of DOX (2.50
mg/kg/dose, via an intraperitoneal injection) at 48 h intervals in
order to achieve a total dose of 15 mg/kg (487 mg/m2)
which is above the threshold of DOX-induced cardiomyopathy (370
mg/m2) (14). Rats in
the normal control and CS control groups received 0.9% NaCl via an
intraperitoneal injection (solvent of DOX) in place of DOX. The
volume of injection was 1.0 ml/kg each time.
Rats in the normal control and DOX control groups
were intragastrically administered drinking water only, and the
rats in the remaining 5 groups were administered with the
corresponding drugs [captopril (0.05 g/kg) or CS (0.75, 1.5 or 3
g/kg)] in a volume of 10 ml/kg, starting 2 days prior to initial
DOX administration and continuing for 23 days.
A total of 24 h following the last injection of DOX,
blood samples (~500 µl each) were collected from the tail vein to
measure the levels of cardiac enzymes in serum. Following the
aforementioned intragastric drug administration for 23 days, the
surviving animals (n=68) were fasted for 12 h. The number of
surviving rats in each group was recorded to calculate the
mortality. Subsequently, rats were anesthetized to measure blood
pressure and electrocardiogram (ECG). Once blood pressure and ECG
measurements were performed, the rats were sacrificed under
anesthesia (urethane, i.p. injection, 1 g/kg). Blood samples were
also collected following ECG and blood pressure measurement and
centrifuged at 4°C and 2,325 × g for 10 min to recover the serum.
The cardiac tissues were excised and the tissues of the left
ventricle were separated. The upper part of left ventricle was
fixed in 10% buffered neutral formalin solution (22–24°C, 1 week)
for histopathological analysis. The serum and the remaining cardiac
tissues were stored at −80°C for subsequent experiments.
Recording blood pressure and ECG
Blood pressure, including systolic blood pressure
(SBP), diastolic blood pressure (DBP) and mean arterial pressure
(MAP), and ECG (position II) were recorded using a multi-channel
biological signal analysis system (RM6240C; Chengdu Instrument
Factory, Chengdu, China). Heart rate, and the duration of the QRS
complex and Sα-T segment were measured. QRS complexes and the Sα-T
segment have been reported to be reliable ECG parameters for the
assessment of DOX-induced cardiotoxicity (15).
Assessment of general data and cardiac
weight indexes
Once treatments and subsequent experiments were
completed, total body weight (BW) and heart weight (HW) were
measured following fasting for 12 h. Following sacrifice, the heart
was removed and the left ventricle was separated from the adipose
tissue and atria once blood samples were collected. The left
ventricle weight (LVW) and HW were recorded, and then the LVW index
(LVWI; the LVW/BW ratio) and the HW index (HWI; the HW/BW ratio)
were calculated. The aforementioned fixed part of left ventricle
was dehydrated in 70% ethanol, cleared in xylene and then embedded
in paraffin. Sections (4-µm thick) were prepared and stained with
hematoxylin-eosin (H&E) at room temperature for 1 day. A light
microscope (UB202i; Chongqing COIC Industrial Co., Ltd., Chongqing,
China) was used to capture the images of each sample.
Endogenous antioxidant enzymes and
malondialdehyde (MDA)
Once serum was recovered from blood samples
(centrifugation at 4°C and 2,325 × g for 10 min), total superoxide
dismutase (T-SOD) activity, glutathione peroxidase (GSH-Px)
activity and MDA content in serum were assayed using T-SOD assay
kit (hydroxylamine method), MDA assay kit (TBA method) and assay
kit (colorimetric method) kit, respectively (Nanjing Jiancheng
Bioengineering Institute, Nanjing, China). All assays were
performed according to the manufacturer's instructions.
Measurement of serum cardiac
enzymes
Blood samples were collected 24 h following the last
DOX injection, serum was recovered and then the activities of
creatine kinase (CK), lactate dehydrogenase (LDH) and aspartate
aminotransferase (AST) in serum were analyzed using the Hitachi
7080 Biochemical Analyzer (Hitachi, Ltd., Tokyo, Japan). CK assay
kit, LDH assay kit and AST assay kit were obtained from Shino-Test
Corporation (Tokyo, Japan). All kits were performed according to
the manufacturer's instructions.
Measurement of brain natriuretic
peptide (BNP)
Serum BNP content was determined by ELISA using the
aforementioned blood samples. The rat BNP ELISA kit (cat. no.
JL15563) was purchased from Shanghai Jianglai Bioengineering
Institute Co., Ltd. (Shanghai, China).
Measurement of adenosine
5′-triphosphate (ATP) ase and CK activities in myocardial
mitochondria
Mitochondria were recovered from cardiac tissues by
differential centrifugation (2,267 × g for 15 min at 4°C, followed
by 10,000 × g for 20 min at 4°C). The activities of mitochondrial
Na+K+-ATPase,
Ca2+Mg2+-ATPase and CK were measured using
Na+K+-ATPase assay kit and
Ca2+Mg2+-ATPase assay kit, respectively.
(Nanjing Jiancheng Bioengineering Institute Co., Ltd). All kits
were used according to the manufacturer's instructions.
Measurement of adenine nucleotides and
phosphocreatine (PCr) contents
ATP, adenosine 5′-diphosphate (ADP), AMP and PCr
contents in cardiac tissues were assayed by HPLC (UltiMate 3000
HPLC system; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Chromatographic separation was achieved on an Ultimate AQ C18
column (4.6×150 mm, 3 µm; Welch Materials Co., Ltd., Shanghai,
China) with 0.2 µm precolumn inline filter maintained at 15°C. The
mobile phase consisted of (A) water (containing 10 mmol/l
KH2PO4 and K2HPO4, pH
6.2) and (B) methanol. A linear gradient elution program was
applied as follows: 0–2 min 100% A, 2–8.5 min 100–30% A and 0–30%
B, 8.5–11 min 30% A and 70% B, 11–14 min 100% A. The flow rate was
1.0 ml/min and the injection volume was 2 µl. Detected wavelength:
210 nm. Total adenine nucleotides (ATP+ADP and +AMP), the PCr/ATP
ratio, the ATP/ADP ratio and the AMP/ATP ratio were also
calculated.
Myocardial mRNA expression of AMPKα2,
peroxisome proliferator-activated receptor α (PPARα) and peroxisome
proliferator-activated receptor γ coactivator-1α (PCG-1α)
The mRNA expression levels of AMPKα2, PPARα and
PCG-1α were determined by reverse transcription-quantitative
polymerase chain reaction. Total RNA was extracted from cardiac
tissues by the Fresh Animal Tissue and Cell Sample Total RNA
Extraction kit (cat. no. LN-0108A; Shanghai Novland
Bio-pharmaceutical Tech. Co., Ltd., Shanghai, China) and
centrifuged at 12,000 × g for 10 min at 4°C. Universal Reverse
Transcription kit (Shanghai Novland Bio-pharmaceutical Tech. Co.,
Ltd., cat. no. LR-0103A) was used for reverse transcription and
then real-time PCR kit (cat. no. LK-0101A; SYBR Green, Shanghai
Novland Bio-pharmaceutical Tech. Co., Ltd., Shanghai, China) was
used for qPCR. The temperature protocol for RT-PCR is 42°C for 30
min and 85°C for 10 min. The following thermocycling conditions
were used for PCR: Initial denaturation at 95°C for 3 min; 40
cycles of 95°C for 12 sec, 60°C for 40 sec and 72°C extension for
30 sec. The method of quantification is according to the references
(16). The following primers were
used: AMPKα2 forward, 5-AAACACGGGAGGGTTGAAGAGG-3 and reverse,
5-ATGTGCCTGTGACAGTAGTCCACG-3; PPARα forward,
5-CATCTGTCCTCTCTCCCCACTTG-3 and reverse,
5-CCGAGGGACTGAGAAATCTCTTG-3; PCG-1α forward,
5-AAACCCTGCCATTGTTAAGACCG-3 and reverse,
5-CGTCTTTGTGGCTTTTGCTGTTG-3; and β-actin forward,
5-CGTAAAGACCTCTATGCCAACA-3 and reverse, 5-GGAGGAGCAATGATCTTGATCT-3.
The mRNA expression levels were determined using the MX3000P qPCR
System (Agilent Technologies GmbH, Waldbronn, Germany) and the
results were normalized to those of β-actin.
Measurement of myocardial cyclic AMP
(cAMP) and activity of serum phosphodiesterases (PDEs)
cAMP contents in cardiac tissues were assayed by
HPLC, performed as previously described for adenine nucleotides.
The activities of serum PDEs selected groups were assayed using
PDEs assay kit (Shanghai Jianglai Bioengineering Institute Co.,
Ltd). In the 0.75 g/kg CS or captopril groups, the cAMP content
were unaltered and therfore PDEs activity was not measured in the
two groups.
Statistical analysis
All values were expressed as the mean ± standard
error mean (n=9). The results were analyzed using SPSS v18.0
software (SPSS, Inc., Chicago, IL, USA). A one-way analysis of
variance followed by Dunnett's post hoc test were performed for
multiple comparisons. Mann-Whitney U rank-sum test was used as an
alternate test for variance heterogeneity. P<0.05 was considered
to indicate a statistically significant difference.
Results
General data and cardiac weight
indexes
The DOX-treated control rats exhibited 40% mortality
when comparing the original number of rats (n=15) to those that
survived treatment (n=9), and when compared with the normal control
group, a reduction in BW, and an increase in LVWI and HWI were
observed (P<0.01). When compared with the DOX control group,
rats treated with DOX and captopril or CS (3 g/kg) showed a
significant decrease in LVWI and HWI (P<0.05 and P<0.01). In
the DOX + 1.5 g/kg CS group, treatment did not result in mortality
as all 14 rats survived. In addition, there were no marked
differences in the cardiac weight indexes between the normal
control group and the CS control group (Table I).
 | Table I.Effect of fermented Cordyceps
sinensis on mortality and cardiac weight indexes in
doxorubicin-treated rats. |
Table I.
Effect of fermented Cordyceps
sinensis on mortality and cardiac weight indexes in
doxorubicin-treated rats.
| Groups | No. of rats
pre-treatment | No. of treatment
induced mortalities | No. of surviving
rats post-treatment | Mortality (%) | BW (g) | LVWI (mg/g) | HWI (mg/g) |
|---|
| Normal control | 10 | 0 | 10 | 0 | 286.2±6.93 | 2.22±0.05 | 2.79±0.05 |
| DOX control | 15 | 6 | 9 | 40.00a |
208.3±12.50b |
2.50±0.09b |
3.11±0.10b |
| DOX plus
captopril | 13 | 3 | 10 | 23.08 | 224.8±8.25 |
2.18±0.05d |
2.73±0.06d |
| DOX plus 0.75 g/kg
CS | 13 | 5 | 8 | 38.46 | 216.3±6.36 | 2.33±0.07 | 2.90±0.08 |
| DOX plus 1.50 g/kg
CS | 14 | 0 | 14 | 0c | 211.0±7.64 | 2.40±0.05 | 3.00±0.08 |
| DOX plus 3.00 g/kg
CS | 14 | 2 | 12 | 14.29 | 219.0±7.53 |
2.26±0.06d |
2.85±0.08c |
| CS control | 5 | 0 | 5 | 0 | 292.0±11.14 | 2.21±0.04 | 2.76±0.05 |
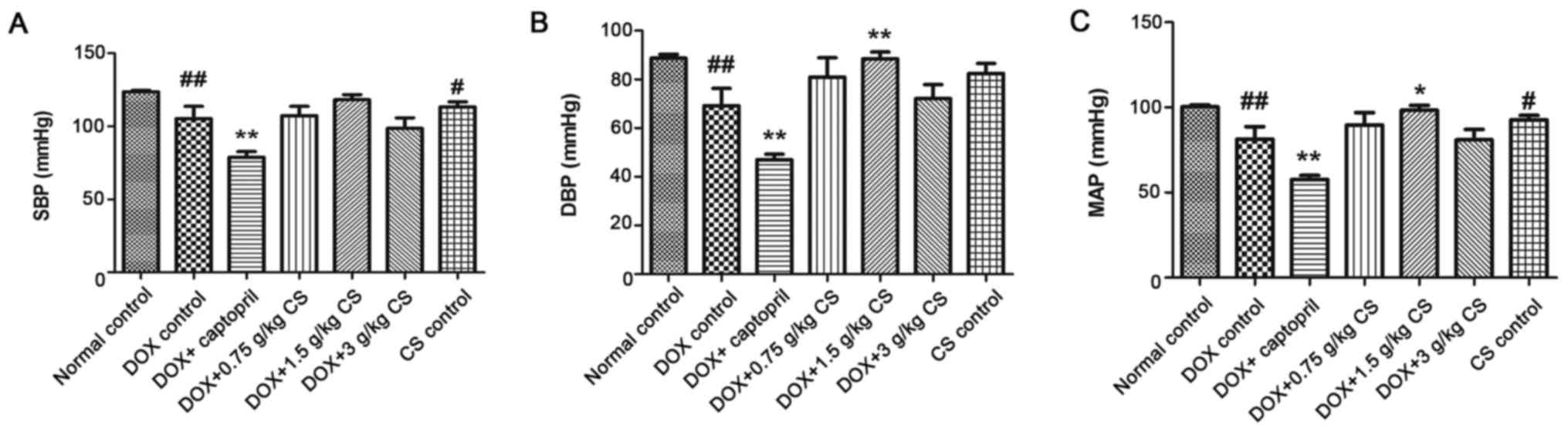
Blood pressure
In the DOX control group, rat SBP, DBP and MAP were
significantly reduced when compared with the normal control group
(P<0.01). In addition, combined treatment with captopril
significantly decreased SBP, DBP and MAP when compared with the DOX
control group (P<0.01). The DBP and MAP of the DOX + 1.5 g/kg CS
group were significantly increased when compared with the DOX
control group (P<0.05 and P<0.01). Furthermore, when compared
with the normal control group, rat SBP and MAP were significantly
lower in CS control group (P<0.05; Fig. 1).
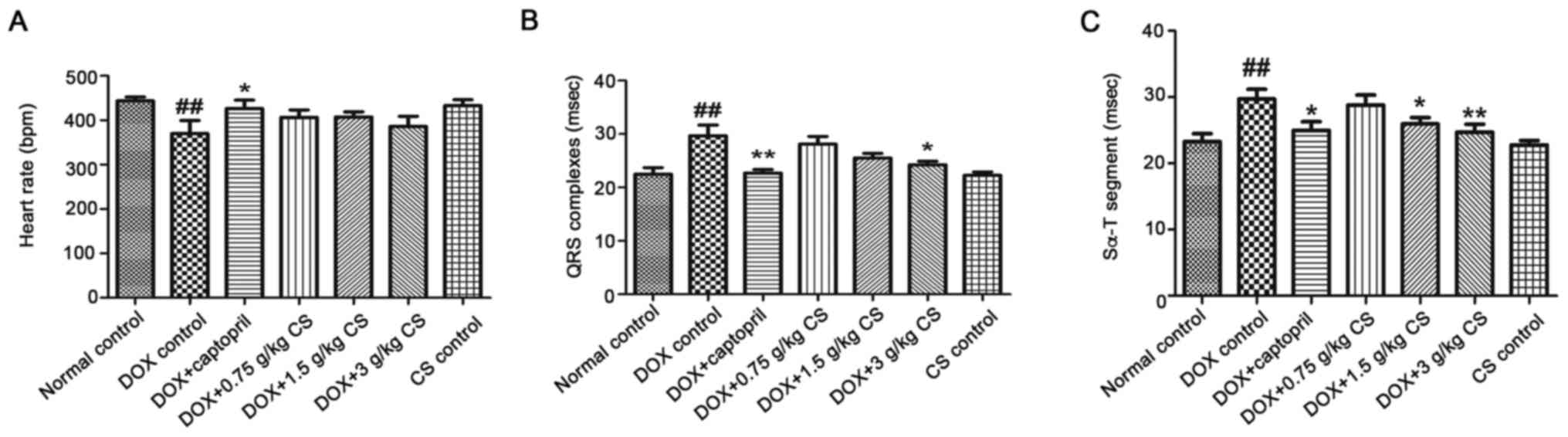
Evaluation of ECG
The DOX control group exhibited a significantly
reduced heart rate, and a prolonged QRS complex and Sα-T segment,
when compared with the normal control group (P<0.01). When
compared with the DOX control group, rats co-treated with captopril
had significantly increased heart rates (P<0.05), and decreased
QRS complex and Sα-T segment durations (P<0.05 and P<0.01).
In addition, rats co-treated with 1.5 g/kg CS had decreased Sα-T
segment durations (P<0.05), and rats co-treated with 3 g/kg CS
showed markedly shorter QRS complex and Sα-T segment durations
(P<0.05 and P<0.01). However, no differences were observed
for all ECG measurements the between normal control and CS control
groups (Fig. 2).
Serum activities of CK, LDH, and
AST
The activities of CK and AST in the DOX control
group were significantly increased when compared with the normal
control group (P<0.05 and P<0.01). The activities of CK, LDH
and AST in the DOX + captopril or CS (1.5 and 3 g/kg) groups were
markedly lower than those of the DOX control group (P<0.05 and
P<0.01). In addition, CK activity was significantly decreased
when comparing the DOX + captopril or CS (0.75 g/kg) groups with
the DOX control group (P<0.05). When compared with normal
control group, rats in CS control group exhibited a significant
decrease in CK, LDH and AST activities (P<0.05 and P<0.01;
Fig. 3).
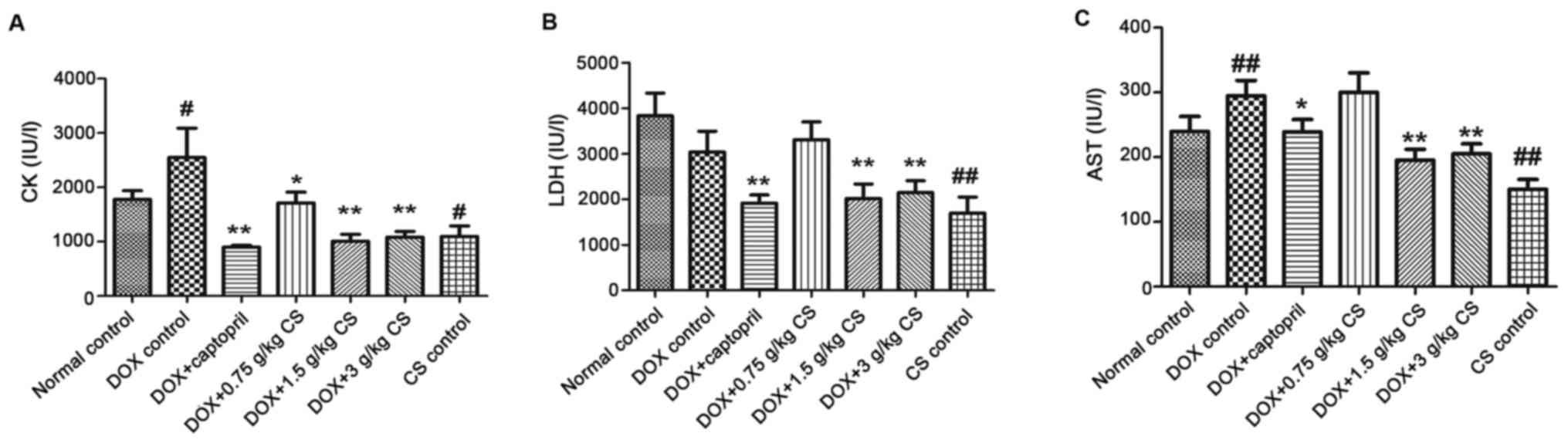 | Figure 3.Effect of fermented CS on the
activities of CK, LDH and AST in the serum of DOX-treated rats.
Blood samples were collected from rats treated with DOX alone, or
in combination with 0.05 g/kg captopril, or with 0.75, 1.5 or 3
g/kg CS in order to measure the levels of (A) CK, (B) LDH, and (C)
AST in rat serum. Data are presented as the mean ± standard
deviation. #P<0.05 and ##P<0.01 vs. the
normal control group; *P<0.05 and **P<0.01 vs. the DOX
control group. CS, Cordyceps sinensis; DOX, doxorubicin; CK,
creatine kinase; LDH, lactate dehydrogenase; AST, aspartate
aminotransferase. |
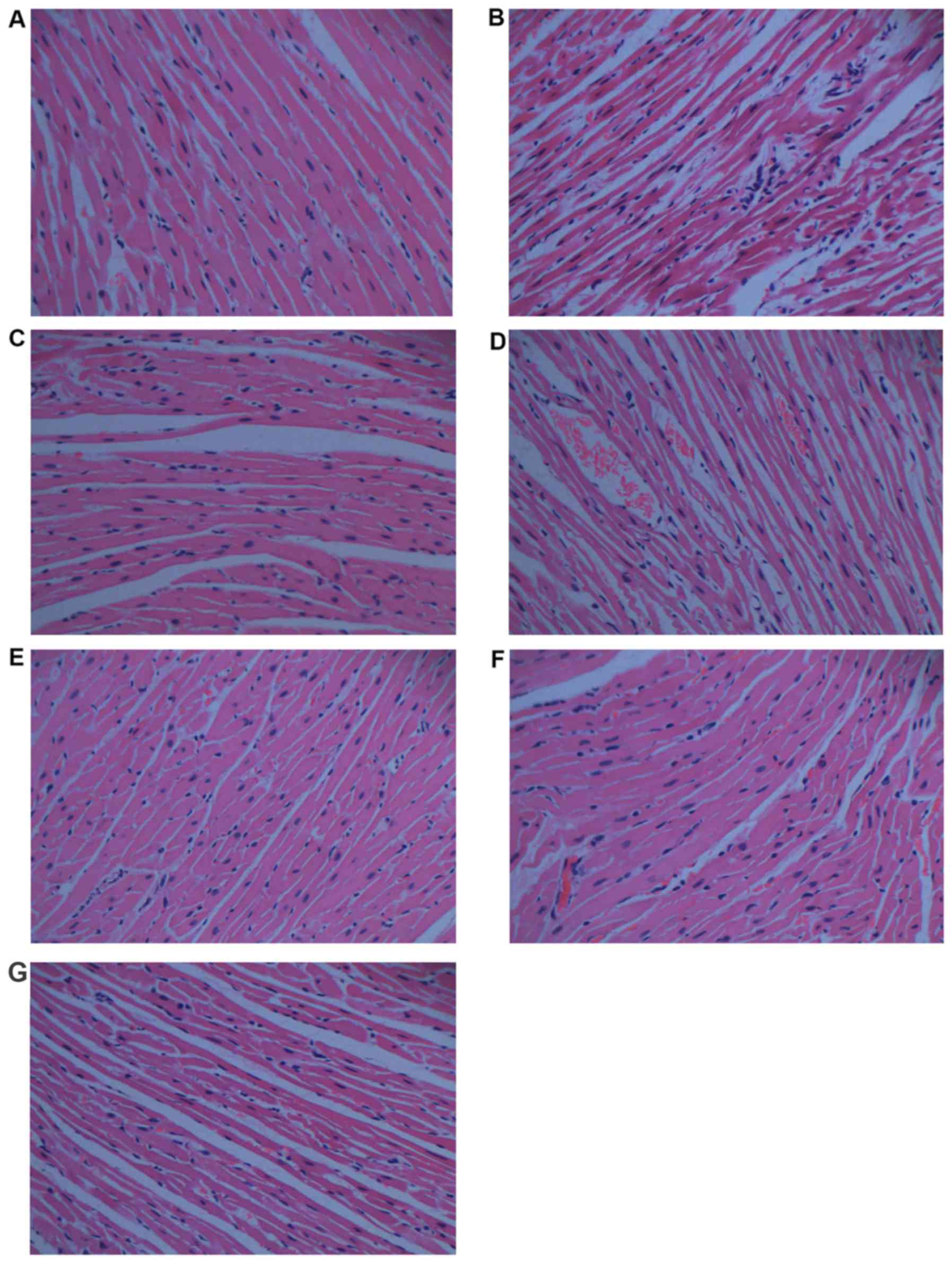
Histopathological analysis
Histopathological analysis is shown in Fig. 4. When compared with the normal
control group (Fig. 4A), the
histological section of the cardiac tissue in rats treated with DOX
(Fig. 4B) shows widespread
vacuolization, marked myocardial degeneration, and some necrotic
cardiomyocytes. DOX treatment with 0.75 g/kg CS (Fig. 4D) and with 1.5 g/kg CS (Fig. 4E) also induced vacuolization and
moderate myocardial degeneration. The DOX + captopril group
(Fig. 4C) and DOX + 3 g/kg CS
group (Fig. 4F) presented only
moderate vacuolization. In addition, no marked differences were
observed in the CS control group (Fig.
4G).
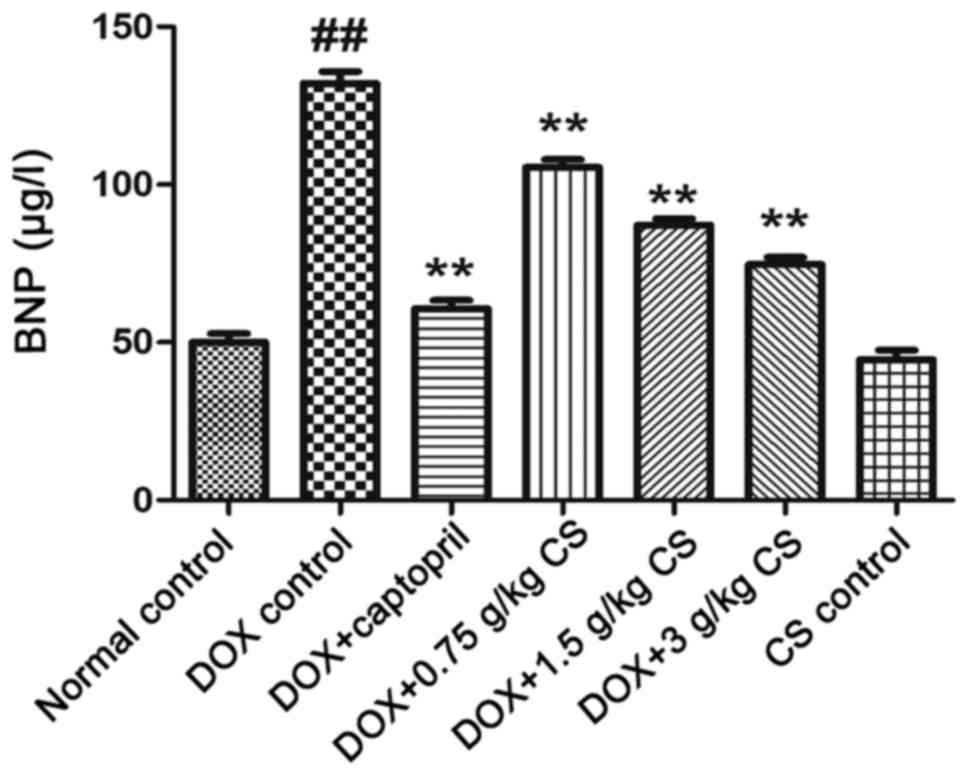
Serum BNP content
The BNP content of the DOX control group was
significantly higher than that of the normal control group
(P<0.01). Co-treatment with DOX and captopril or CS (0.75, 1.5
and 3 g/kg) significantly reduced the BNP content when compared
with DOX treatment alone (P<0.01). No marked difference in BNP
content was observed between the normal control and CS control
groups (Fig. 5).
Serum activities of T-SOD and GSH-Px,
and MDA content
The DOX control treated rats exhibited significantly
decreased T-SOD and GSH-Px activities, and increased MDA content
when compared with the normal control (P<0.01). In rats
co-treated with captopril or CS (0.75, 1.5 and 3 g/kg), the T-SOD
and GSH-Px activities were increased, and MDA content was
significantly decreased, when compared with those observed in the
DOX control group (P<0.05 and P<0.01). There were no marked
differences in the serum activities of T-SOD and GSH-Px, and MDA
content between the normal control and CS control groups (Fig. 6).
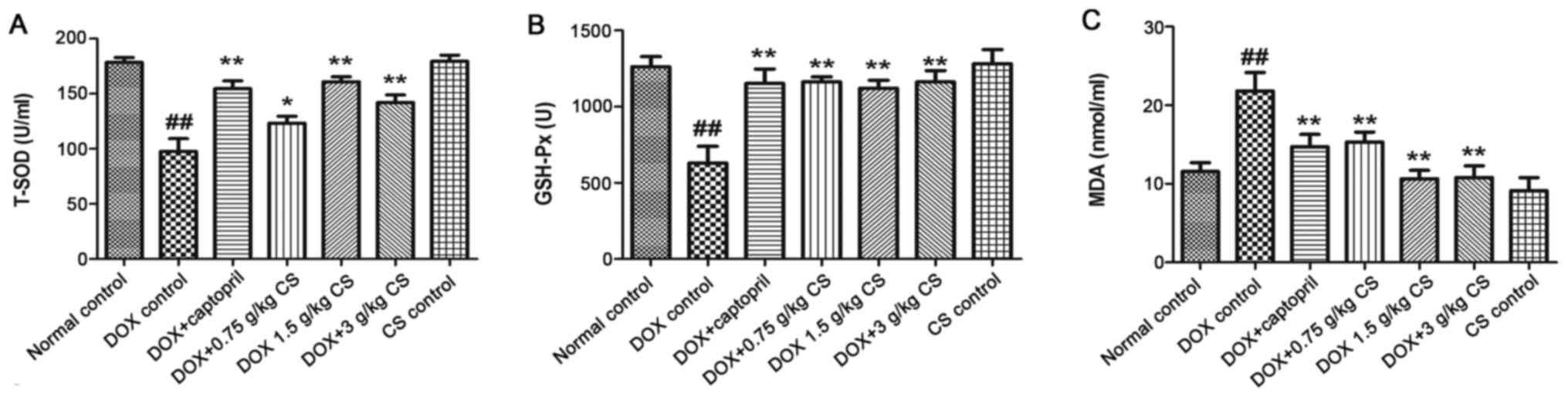 | Figure 6.Effect of fermented CS on T-SOD and
GSH-Px activities, and MDA content in DOX-treated rats. Serum was
recovered from blood samples in order to measure (A) T-SOD and (B)
GSH-Px activities, and (C) MDA content in rats treated with DOX
alone, or in combination with 0.05 g/kg captopril, or with 0.75,
1.5 or 3 g/kg CS. Data are presented as the mean ± standard
deviation. ##P<0.01 vs. the normal control group;
*P<0.05 and **P<0.01 vs. the DOX control group. CS,
Cordyceps sinensis; DOX, doxorubicin; T-SOD, total
superoxide dismutase; GSH-Px, glutathione peroxidase; MDA,
malondialdehyde. |
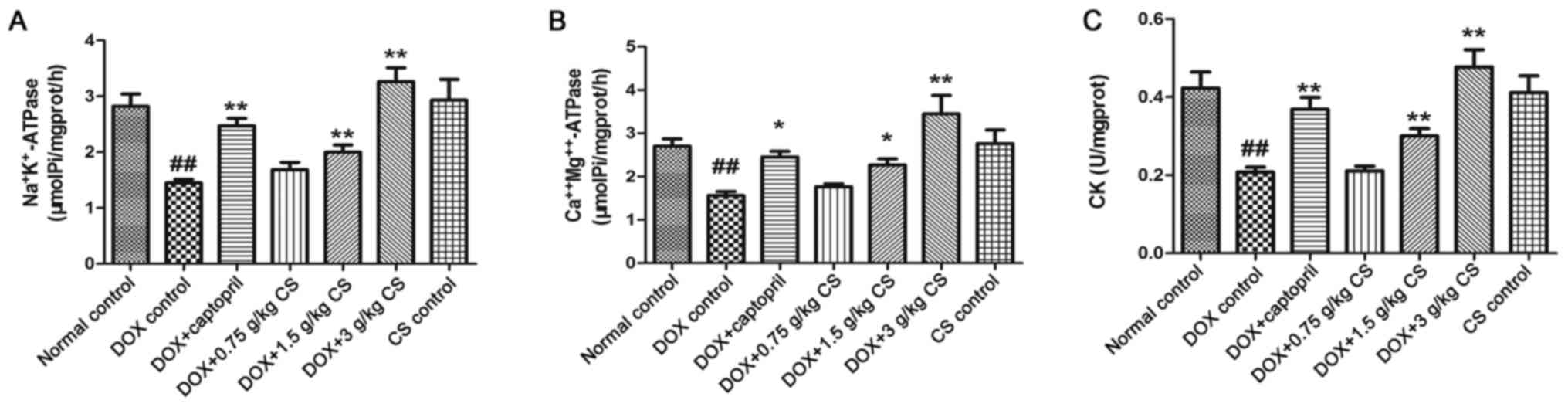
Activities of
Na+K+-ATPase,
Ca2+Mg2+-ATPase and CK in myocardial
mitochondria
In the DOX control group, the rats exhibited
significantly reduced mitochondrial activities of
Na+K+-ATPase,
Ca2+Mg2+-ATPase and CK (P<0.01). When
compared with the DOX control group, the mitochondrial activities
of Na+K+-ATPase,
Ca2+Mg2+-ATPase and CK in the DOX + captopril
or CS (1.5 and 3 g/kg) groups were significantly increased
(P<0.05 and P<0.01). However, no marked differences in CK or
ATPase activities were observed between the normal control and CS
control groups (Fig. 7).
Adenine nucleotides (ATP, ADP and AMP)
and PCr contents
In the DOX control group, the ADP content and the
PCr/ATP and AMP/ATP ratios were significantly reduced (P<0.05),
and the ATP/ADP ratio was significantly increased, when compared
with the normal control group (P<0.01). When compared with the
DOX control group, rats co-treated with captopril presented
significantly increased ADP and total adenine nucleotides
(ATP+ADP+AMP fold-change) contents (P<0.05 and P<0.01), and a
decreased ATP/ADP ratio (P<0.05). Rats co-treated with 0.75 g/kg
CS exhibited increased PCr/ATP and AMP/ATP ratios (P<0.05) and a
reduced ATP/ADP ratio (P<0.01). Co-treatment with 1.5 g/kg CS
increased ADP, AMP and total adenine nucleotides content, and the
AMP/ATP ratio (P<0.05 and P<0.01). In addition, DOX + 3 g/kg
CS increased the ATP, ADP, AMP and total adenine nucleotides
contents, and the AMP/ATP ratio (P<0.05 and P<0.01). When
compared with the normal control group, rats in the CS control
group had significantly increased ATP and AMP contents, and ATP/ADP
and AMP/ATP ratios; however, the PCr/ATP ratio was significantly
decreased (P<0.01; Fig. 8). No
differences were observed across the groups in PCr content.
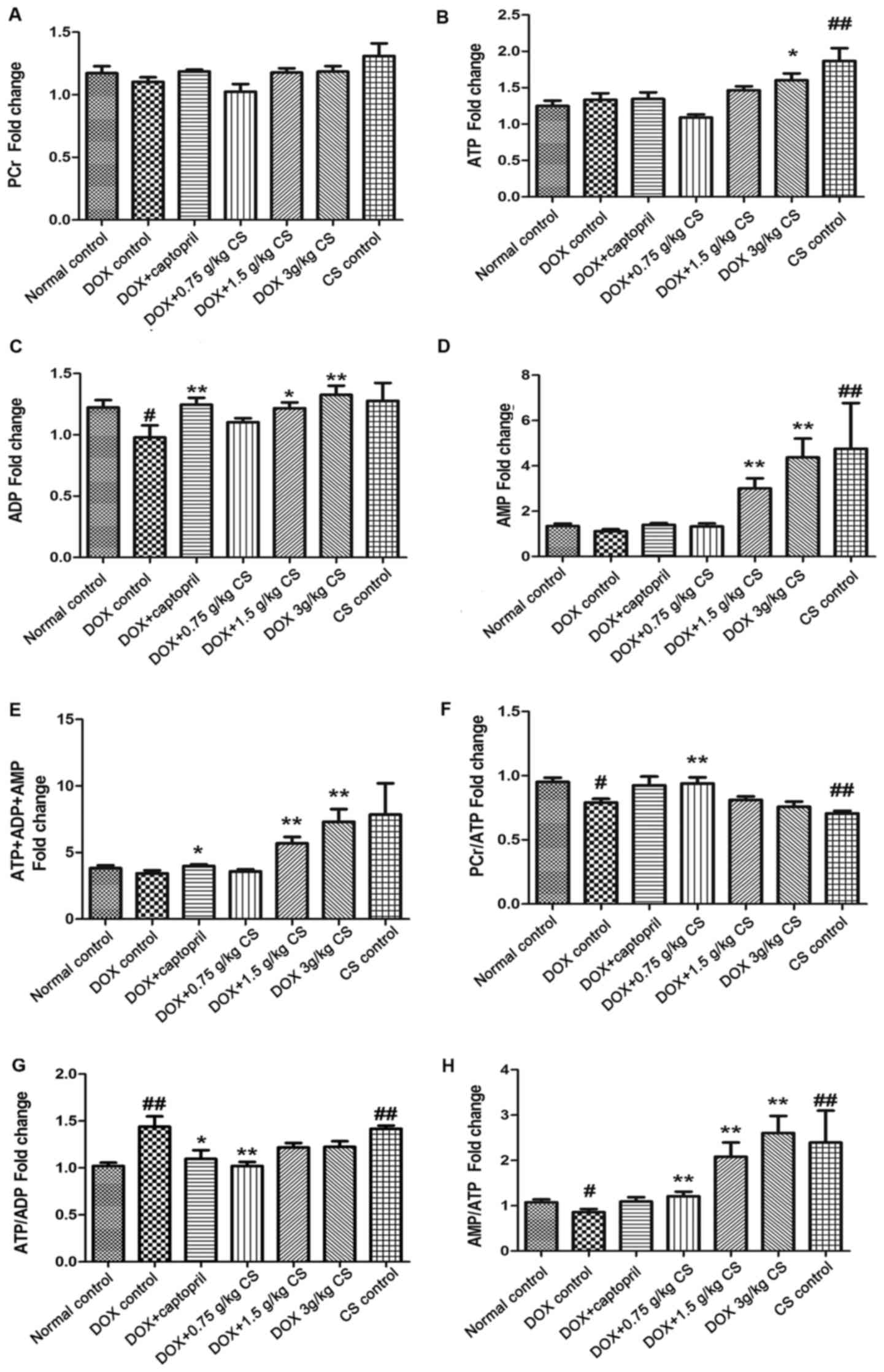 | Figure 8.Effect of fermented CS on adenine
nucleotides and PCr contents in DOX-treated rats. Cardiac tissues
were collected from DOX treated rats to measure the (A) PCr, (B)
ATP, (C) ADP, (D) AMP and (E) ATP+ADP+AMP (total adenine
nucleotides) contents, and the (F) PCr/ATP ratio, (G) ATP/ADP ratio
and (H) AMP/ATP ratio via high-performance liquid chromatography.
The contents of adenine nucleotides and PCr in each group were
expressed as the fold change of the normal control group. Data are
presented as the mean ± standard deviation. #P<0.05
and ##P<0.01 vs. the normal control group; *P<0.05
and **P<0.01 vs. the DOX control group. CS, Cordyceps
sinensis; DOX, doxorubicin; PCr, phosphocreatine; ATP adenosine
triphosphate; ADP, adenosine diphosphate; AMP, adenosine
monophosphate. |
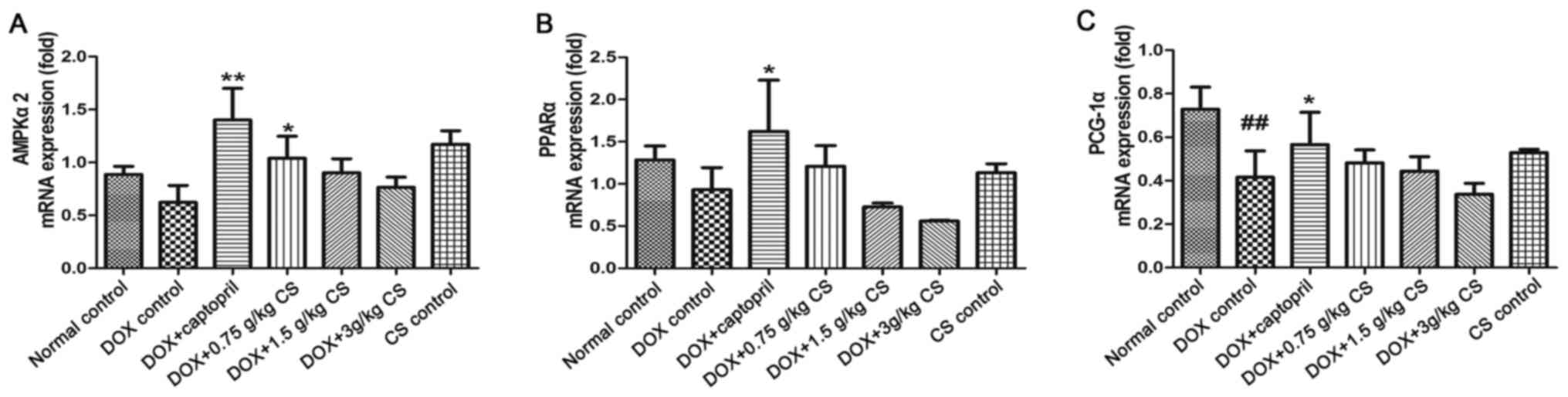
mRNA expression of AMPKα2, PPARα and
PCG-1α
When compared to the normal control group, PCG-1α
mRNA expression was significantly reduced in the DOX control
treated rats (P<0.01). Rats co-treated with captopril had
significantly increased AMPKα2, PPARα and PCG-1α mRNA expression
when compared with the levels presented by the DOX control group
(P<0.05 and P<0.01). In the DOX + 0.75 g/kg CS treatment
group, the AMPKα2 mRNA expression was significantly greater than
that of the DOX control group (P<0.05). However, no marked
difference in mRNA expression was observed between the normal
control and CS control groups (Fig.
9).
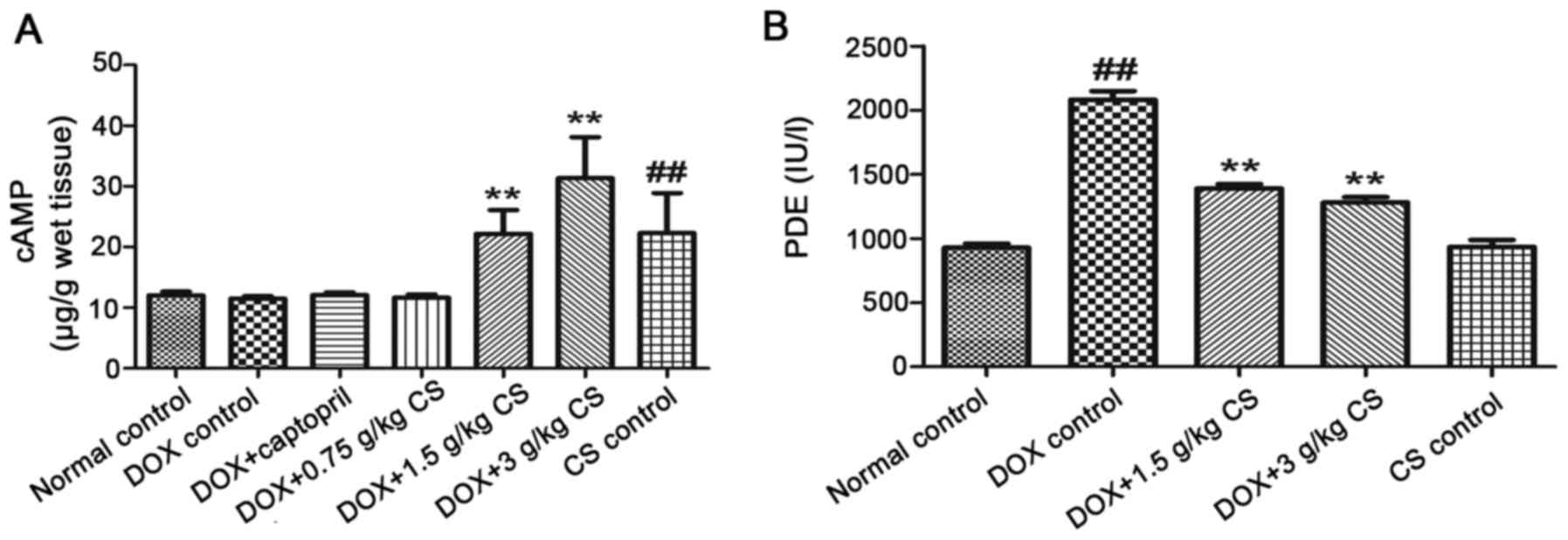
Myocardial cAMP contents and serum PDE
activity
When compared with the normal control group, the DOX
control treated rats exhibited an increase in the activity of serum
PDEs (P<0.01). In CS (1.5 and 3 g/kg) treated rats, the content
of myocardial cAMP and serum PDE activity were significantly lower
than that observed in the DOX control group (P<0.01). In
addition, when compared with the normal control group, rats in the
CS control group had significantly increased myocardial cAMP
content (P<0.01; Fig. 10).
Discussion
DOX-induced cumulative cardiotoxicity is the
greatest problem to overcome in order to improve clinical
application. DOX can induce cardiac and systemic toxicity (17), while the cardiomyopathy induced by
DOX serves a key role in the mortality of experimental animals
following treatment with DOX (18,19).
The experimental model used in the present study has been reported
previously (19). CS, a TCM, has a
variety of pharmacological effects. A previous study demonstrated
that the extracts from the mycelia of CS have cardiovascular
protective and antioxidant activities (20). It has also been demonstrated that
oral administration of CS significantly attenuated liver and
cardiac-associated injuries in rats with anti-chronic kidney
disease (21). In the present
study, rats were treated with a cumulative dose of 15 mg/kg of DOX
to mimic the DOX-induced cumulative cardiotoxicity observed in
clinical treatments. This dose is above the threshold at which
doxorubicin cardiomyopathy is expected to clinically occur
(14). It was observed that 40% of
the rats in the DOX control group, succumbed to treatment prior to
the termination of the experiment, which is similar to the results
of Kelishomi et al (17).
There was quite a low mortality in DOX + 3 g/kg CS treated animals,
and none of rats succumbed in the DOX + 1.5 g/kg CS treated. This
result indicated that treatment with fermented CS may enhance the
survival of DOX-treated rats. In addition, a significant decrease
in BW was observed in the DOX control group. This reduction may be
attributable to an inhibition of protein synthesis, mucositis and a
reduction in food intake due to DOX treatment (18).
DOX-induced cardiomyopathy is a transition between
myocardial hypertrophy and heart failure (22). It is a severe adverse reaction of
DOX, and is usually accompanied by myocardial hypertrophy. In
clinical treatments, myocardial hypertrophy is associated with
mortality in patients (23).
Myocardial hypertrophy is primarily evaluated by measuring HWI and
LVWI in animals (24). In the
present study, an increase in HWI and LVWI was observed following
DOX treatment which demonstrated that myocardial hypertrophy is
induced by DOX; this result is in accordance with a previous study
involving DOX-treated rats (22).
In the DOX + 3 g/kg CS group, the increase in the HWI and LVWI was
attenuated. This result indicated that a high dose of fermented CS
may contribute to the inhibition of myocardial hypertrophy,
potentially leading to the attenuation of DOX-induced
cardiomyopathy.
Systolic and diastolic functioning of heart are
closely associated with cardiac dysfunction. Previous studies have
reported that DOX treatment induced diastolic dysfunction, leading
to alterations in systolic function (25,26).
These alterations included a reduction in BP. In the present study,
a significant decrease in SBP, DBP and MAP was observed in
DOX-treated rats. This decrease is similar to results observed
previously (27), and is
potentially caused by DOX-induced myofibril disruption (28). Rats treated with DOX + 1.5 g/kg CS
had significantly elevated DBP and MAP, suggesting that fermented
CS may improve systolic and diastolic function, and may have a
potential role in therapies for DOX-induced cardiac
dysfunction.
ECGs can reflect the electrical activity of the
heart and identify damage to the myocardium in specific areas. It
is also one of the most classic methods applied to diagnose heart
injuries (29). Larsen et
al (30) reported that ECG
analysis can be used to evaluate DOX-induced cardiotoxicity in
clinical practice. According to previous studies, the duration of
the QRS complexes and Sα-T segment are the most reliable ECG
parameters in the assessment of DOX-induced cardiotoxicity in rats
(14,31). The duration of the transmembrane
action potential is easily affected by DOX treatment, thus, all ECG
alterations are accompanied by a prolongation of action potential
duration (31). In the present
study, the prolongation of the duration of the QRS complexes and
Sα-T segment, and a reduction in heart rate, were observed
following DOX treatment. This result is in agreement with previous
research (14). In rats treated
with 3 g/kg CS, there was a decrease in the prolongation of QRS
complexes and Sα-T segment, and rats treated with 1.5 g/kg CS also
exhibited a reduction in the prolongation of the Sα-T segment.
These results demonstrated that fermented CS may be able to
attenuate DOX-induced cardiotoxicity.
The cardiac enzymes, CK, LDH and AST, are abundant
in cardiac muscle, particularly CK and LDH. In clinical settings,
an elevation of cardiac enzymes in serum is considered a vital
biomarker of myocardial damage, indicating a number of cardiac
diseases, such as myocarditis and myocardial infarction (32). Detecting the activities of these
enzymes in serum is essential for assessing myocardial damage. DOX
treatment can increase the activities of cardiac enzymes in serum
during clinical therapy (33). The
increase in serum cardiac enzymes in DOX-treated rats, due to
DOX-induced cardiac injury, has been reported previously (34). Histological changes also serve an
important role in evaluating DOX cardiotoxicity. In the present
study, the increase in CK and AST in the serum and the microscopic
findings indicated that DOX-induced myocardial damage. In the
majority of the CS treated animals, the secretion of LDH, AST and
CK, and the histological changes observed (including vacuolization,
myocardial degeneration and necrotic cardiomyocytes) were
significantly attenuated, indicating that fermented CS may be able
to inhibit DOX-induced myocardial necrosis, leading to cardiac
protection from DOX-induced myocardial damage.
As a sensitive biomarker of heart failure, BNP is
secreted by cardiomyocytes and is frequently used in emergency
settings (35). During heart
failure, a large quantity of BNP is released into the blood; due to
this, it can be used for diagnosing heart failure induced by DOX
(36). In the present study, DOX
treatment significantly increased serum BNP, thereby demonstrating
that acute heart failure may be caused by DOX. Rats treated with CS
exhibited a decrease in serum BNP, indicating that fermented CS may
be able to protect the heart from DOX-induced cardiotoxicity and
acute heart failure.
OS, the basis of a number of diseases, is considered
to be a direct factor of many types of cardiac damage (37), such as cardiomyopathy and cardiac
dysfunction. It is generally believed that OS serves a vital role
in the mechanism underlying DOX-induced cardiotoxicity (6,27).
In some previous studies, DOX promoted the production of ROS and
MDA (6), and inhibited the
activities of SOD and GSH-Px (6),
resulting in damage to myocardial mitochondria (38). As a final product of
lipoperoxidation, MDA is a reliable biomarker for evaluating the
degree of oxidative damage (39).
SOD and GSH-Px are involved in a critical defense mechanism that
protects the body from the harmful effects of ROS (40). Thus, the degree of OS is assessed
by measuring the activities of T-SOD and GSH-Px, and MDA content.
In the present study, the decrease in the activities of T-SOD and
GSH-Px, and the increase in MDA content demonstrated that DOX
treatment induced OS. An elevation in the activities of T-SOD and
GSH-Px, and a reduction in MDA content were observed in the DOX +
CS (0.75, 1.5 and 3 g/kg) groups. These results indicated that
fermented CS may significantly inhibit the oxidative damage caused
by treatment with DOX, particularly by improving the activities of
antioxidant enzymes. It can be speculated that the anti-OS effect
of fermented CS may serve an important role in protecting the heart
against DOX-induced cardiotoxicity.
ATPases, containing
Na+K+-ATPase and
Ca2+Mg2+-ATPase, are primarily found in
mitochondria. These enzymes can maintain the cell membrane
potential, catalyze the decomposition of ATP into ADP and release
energy (41). They are also
important for muscle tissues, such as cardiac muscle, as a decline
in mitochondrial ATPases can lead to energy impairment.
Mitochondrial CK is a modulator of the energy reservoir, that
converts creatine to phosphocreatine (41). A reduction in mitochondrial CK
activity can affect energy metabolism and the electron transport
chain (42). Cardiac energy
impairment is one of the main characteristics of DOX-induced
mitochondrial ultrastructural damage (1). In the present study, the reduction in
the activities of mitochondrial ATPases and CK demonstrated that
DOX treatment induced mitochondrial ultrastructural damage in the
heart, which is in agreement with a previous study (22). Rats treated with CS exhibited an
increase in the mitochondrial activities of ATPases and CK. This
result indicated that fermented CS may significantly inhibit
DOX-induced mitochondrial ultrastructural damage, and may have a
positive effect in therapies for cardiac energy impairment induced
by DOX as well as other causes.
Cardiac energy metabolism is associated with a
number of cardiac diseases, such as heart failure and heart
remodeling. Due to the high energy requirements, a large amount of
ATP is transported to cardiac muscles every day (43). ATP is predominantly produced by
carbohydrate oxidation and fatty acid (FA) oxidation from ADP or
AMP, as well as other donors. Thus, total adenine nucleotides (ATP,
ADP and AMP) are important in energy metabolism (44). In the process of cardiac energy
metabolism, FAs are the primary substrates that provide ATP under
normal conditions; however, glucose is frequently used during heart
failure (43). The ATP/ADP ratio
serves an important role in indicating the balance of FA and
glucose utilization. During heart failure, mitochondrial damage
frequently leads to a low production of mitochondrial ATP and a
decrease in the ATP/ADP ratio that favors enhanced glycolysis
(45). However, in a previous
study investigating heart failure, the ATP/ADP ratio was observed
to be increased (46). The
decrease in glucose utilization is potentially due to an impairment
of the expression of associated genes (47). PCr is able to donate phosphate
groups to ADP to form ATP under anaerobic conditions. By contrast,
excess ATP can be used to produce PCr from creatine; a reduction in
PCr will accelerate the depletion of ATP (48), particularly during early heart
failure. Thus, the PCr/ATP ratio is a key feature used to determine
the extent of energy impairment. AMPK is mainly regulated by AMP,
and an increase in the AMP/ATP ratio can activate the expression of
AMPK (49). In some previous
studies, DOX induced a number of alterations in cardiac energy
metabolism (1,22,42).
These alterations mainly included a reduction in the PCr/ATP ratio
and a depletion of total adenine nucleotides. In the present study,
an increase in the ATP/ADP ratio of DOX-treated rats was observed,
similar to the results reported by Amorim et al (46). A reduction in ADP, the PCr/ATP
ratio and the AMP/ATP ratio were also identified, demonstrating
that DOX treatment induced cardiac energy impairment. In addition,
CS control rats exhibited an increase in the AMP/ATP and ATP/ADP
ratios, which indicated that fermented CS may potentially increase
AMPK expression and FA utilization in the healthy heart. In CS
treated animals, the levels of total adenine nucleotides, PCr/ATP
ratio and AMP/ATP ratio were significantly improved, indicating
that fermented CS may be able to attenuate the cardiac energy
impairment induced by DOX; this improvement may also be associated
with the AMPK signaling pathway.
AMPK, an important energy regulator and sensor, is a
heterotrimer consisting of α, β and γ subunits; AMPKα2 is the main
subunit in cardiac muscle. AMPK can increase energy production,
maintain myocardial energy homeostasis and inhibit apoptosis, thus,
a previous study suggested that it may be considered as a potential
therapeutic target in a number of cardiac diseases including heart
failure, myocardial infarction and cardiac ischemia (50). PPARα, a major regulator of lipid
metabolism, is predominantly found in the liver, followed by the
heart and kidneys (51). PPARα can
promote the utilization, catabolism and uptake of FAs; a reduction
in cardiac PPARα serves an important role in decreasing FA
utilization (52). The expression
of PPARα is dependent on AMPK (53). Agonists of PPARα are able to
activate AMPK in order to increase energy production, such as via
fatty acid oxidation (53).
PCG-1α, a regulator of energy metabolism and mitochondrial function
(54), can activate oxidative
phosphorylation. The gene silencing of PCG-1α will inhibit
oxidative phosphorylation, downregulate the mRNA expression of
mitochondria and accelerate heart failure (55). Previous studies have reported that
DOX induces a reduction in the genetic expression of AMPKα2, PPARα
and PCG-1α, leading to an impairment of energy metabolism (4,56).
In the present study, a significant decrease in PCG-1α mRNA
expression indicated that DOX treatment induced an impairment in
energy metabolism; DOX treatment also decreased the mRNA expression
of AMPKα2 and PPARα to certain extent. In DOX + 0.75 g/kg CS group,
an increase in AMPKα2 mRNA expression was observed, while the mRNA
expression of PPARα and PCG-1α was marginally improved. These
results revealed that fermented CS may potentially contribute
towards the regulation of the AMPK signaling pathway. In addition,
fermented CS may improve cardiac energy metabolism, as indicated by
a significant increase in the mitochondrial capacities of ATPases
combined with improved total adenine nucleotides contents and
PCr/ATP ratio. It may also be involved in the regulation of the
AMPK signaling pathway.
cAMP, a second messenger derived from ATP, can
activate protein kinases, and regulate glucagon and adrenaline,
thereby improving energy metabolism, and preventing myocardial
ischemia and myocardial damage (57). The cAMP signaling pathway serves an
important role in a number of biological processes, and is
converted into AMP by PDE enzymes (58). PDEs include a group of enzymes that
dephosphorylate cAMP and cyclic guanosine monophosphate (cGMP)
(59). PDEs are important
regulators of these second messenger molecules; inhibition of PDEs
can enhance the effects of biological processes mediated by cGMP or
cAMP (60). Therefore, PDEs and
inhibitors of PDEs are important in clinical therapies (61). PDE activities are known to be
increased under specific conditions, such as during ischemia and
spinal cord injury (61). In the
present study, an increase in serum PDE activities was associated
with DOX treatment in rats. Rats co-treated with CS exhibited a
significant increase in myocardial cAMP content and a marked
decrease in serum PDEs. CS control rats also presented a
significant increase in myocardial cAMP content. These results
revealed that fermented CS may be able to increase myocardial cAMP,
potentially induced via the inhibition of PDE activities. This may
be one of the effective targets of fermented CS, which is
contributed to an improvement in energy metabolism and ameliorated
heart failure. Additional studies are required to evaluate whether
or not fermented CS is a PDE inhibitor.
In conclusion, fermented CS ameliorated the
cardiotoxicity induced by DOX in rats. The results demonstrated
that CS treatment was able to inhibit myocardial hypertrophy and
myocardial damage, improve the antioxidant enzyme system and energy
metabolism, and inhibit PDE activities. Its potential effects on
the upregulation of the AMPK and cAMP signaling pathways were also
revealed. Therefore, fermented CS may be a promising adjuvant of
DOX for clinical application. It may also be a candidate for the
prevention of a number of cardiac diseases. However, further
studies are required to confirm the accurate targets of fermented
CS in the AMPK and/or cAMP signaling pathways.
Acknowledgements
The present study was supported by the Shanghai
University of Traditional Chinese Medicine Budget Project (grant
no. 2014YSN07). The authors would also like to thank Professor
Chang-Xun Chen (Department of Pharmacology, School of Pharmacy,
Shanghai University of Traditional Chinese Medicine, Shanghai,
China) for his helpful suggestions and guidance.
References
|
1
|
Octavia Y, Tocchetti CG, Gabrielson KL,
Janssens S, Crijns HJ and Moens AL: Doxorubicin-induced
cardiomyopathy: From molecular mechanisms to therapeutic
strategies. J Mol Cell Cardiol. 52:1213–1225. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carvalho C, Santos RX, Cardoso S, Correia
S, Oliveira PJ, Santos MS and Moreira PI: Doxorubicin: The good,
the bad and the ugly effect. Curr Med Chem. 16:3267–3285. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parker MA, King V and Howard KP: Nuclear
magnetic resonance study of doxorubicin binding to cardiolipin
containing magnetically oriented phospholipid bilayers. Biochim
Biophys Acta. 1514:206–216. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wu R, Gao JP, Wang HL, Gao Y, Wu Q and Cui
XH: Effects of fermented Cordyceps sinensis on oxidative stress in
doxorubicin treated rats. Pharmacogn Mag. 11:724–731. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tokarska-Schlattner M, Wallimann T and
Schlattner U: Alterations in myocardial energy metabolism induced
by the anti-cancer drug doxorubicin. C R Biol. 329:657–668. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lebrecht D, Setzer B, Ketelsen UP,
Haberstroh J and Walker UA: Time-dependent and tissue-specific
accumulation of mtDNA and respiratory chain defects in chronic
doxorubicin cardiomyopathy. Circulation. 108:2423–2429. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li SP, Li P, Lai CM, Gong YX, Kan KK, Dong
TT, Tsim KW and Wang YT: Simultaneous determination of ergosterol,
nucleosides and their bases from natural and cultured Cordyceps by
pressurised liquid extraction and high-performance liquid
chromatography. J Chromatogr A. 1036:239–243. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhu JS, Halpern GM and Jones K: The
scientific rediscovery of an ancient Chinese herbal medicine:
Cordyceps sinensis: Part I. J Altern Complement Med. 4:289–303.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lo HC, Hsu TH, Tu ST and Lin KC:
Anti-hyperglycemic activity of natural and fermented Cordyceps
sinensis in rats with diabetes induced by nicotinamide and
streptozotocin. Am J Chin Med. 34:819–832. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ji J, Liu J, Liu H and Wang Y: Effects of
fermented mushroom of cordyceps sinensis, rich in selenium, on
uterine cervix cancer. Evid Based Complement Alternat Med.
2014:1731802014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin XX, Xie QM, Shen WH and Chen Y:
Effects of fermented Cordyceps powder on pulmonary function in
sensitized guinea pigs and airway inflammation in sensitized rats.
Zhongguo Zhong Yao Za Zhi. 26:622–625. 2001.(In Chinese).
PubMed/NCBI
|
|
12
|
National Pharmacopoeia Committee:
Pharmacopoeia of the people's Republic of China. China Medical
Science Press; Beijing: pp. 832–833. 2015
|
|
13
|
Council NR: Guide for the care and use of
laboratory animals. The National Academies Press; Washington, DC:
1996
|
|
14
|
Xiao J, Sun GB, Sun B, Wu Y, He L, Wang X,
Chen RC, Cao L, Ren XY and Sun XB: Kaempferol protects against
doxorubicin-induced cardiotoxicity in vivo and in vitro.
Toxicology. 292:53–62. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Danesi R, Del Tacca M and Soldani G:
Measurement of the S alpha T segment as the most reliable
electrocardiogram parameter for the assessment of
adriamycin-induced cardiotoxicity in the rat. J Pharmacol Methods.
16:251–259. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kelishomi RB, Ejtemaeemehr S, Tavangar SM,
Rahimian R, Mobarakeh JI and Dehpour AR: Morphine is protective
against doxorubicin-induced cardiotoxicity in rat. Toxicology.
243:96–104. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Herman EH, Zhang J, Chadwick DP and
Ferrans VJ: Comparison of the protective effects of amifostine and
dexrazoxane against the toxicity of doxorubicin in spontaneously
hypertensive rats. Cancer Chemother Pharmacol. 45:329–334. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rossi F, Filippelli W, Russo S, Filippelli
A and Berrino L: Cardiotoxicity of doxorubicin: Effects of drugs
inhibiting the release of vasoactive substances. Pharmacol Toxicol.
75:99–107. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan XF, Zhang ZM, Yao HY, Guan Y, Zhu JP,
Zhang LH, Jia YL and Wang RW: Cardiovascular protection and
antioxidant activity of the extracts from the mycelia of Cordyceps
sinensis act partially via adenosine receptors. Phytother Res.
27:1597–1604. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu X, Zhong F, Tang XL, Lian FL, Zhou Q,
Guo SM, Liu JF, Sun P, Hao X, Lu Y, et al: Cordyceps
sinensis protects against liver and heart injuries in a rat
model of chronic kidney disease: A metabolomic analysis. Acta
Pharmacol Sin. 35:697–706. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ahmed LA and El-Maraghy SA: Nicorandil
ameliorates mitochondrial dysfunction in doxorubicin-induced heart
failure in rats: Possible mechanism of cardioprotection. Biochem
Pharmacol. 86:1301–1310. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schillaci G, Verdecchia P, Porcellati C,
Cuccurullo O, Cosco C and Perticone F: Continuous relation between
left ventricular mass and cardiovascular risk in essential
hypertension. Hypertension. 35:580–586. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuzman JA, O'Connell TD and Gerdes AM:
Rapamycin prevents thyroid hormone-induced cardiac hypertrophy.
Endocrinology. 148:3477–3484. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Almeida AL and dos Santos Júnior EG:
Subclinical ventricular dysfunction detected by speckle-tracking
two years after use of anthracycline-reply. Arq Bras Cardiol.
105:2072015.PubMed/NCBI
|
|
26
|
Tassan-Mangina S, Codorean D, Metivier M,
Costa B, Himberlin C, Jouannaud C, Blaise AM, Elaerts J and
Nazeyrollas P: Tissue Doppler imaging and conventional
echocardiography after anthracycline treatment in adults: Early and
late alterations of left ventricular function during a prospective
study. Eur J Echocardiogr. 7:141–146. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ozdogan K, Taskin E and Dursun N:
Protective effect of carnosine on adriamycin-induced oxidative
heart damage in rats. Anadolu Kardiyol Derg. 11:3–10. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Weinberg LE and Singal PK: Refractory
heart failure and age-related differences in adriamycin-induced
myocardial changes in rats. Can J Physiol Pharmacol. 65:1957–1965.
1987. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo X, Xue M, Li CJ, Yang W, Wang SS, Ma
ZJ, Zhang XN, Wang XY, Zhao R, Chang BC and Chen LM: Protective
effects of triptolide on TLR4 mediated autoimmune and inflammatory
response induced myocardial fibrosis in diabetic cardiomyopathy. J
Ethnopharmacol. 193:333–344. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Larsen RL, Jakacki RI, Vetter VL, Meadows
AT, Silber JH and Barber G: Electrocardiographic changes and
arrhythmias after cancer therapy in children and young adults. Am J
Cardiol. 70:73–77. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Villani F, Galimberti M, Monti E, Cova D,
Lanza E, Rozza-Dionigi A, Favalli L and Poggi P: Effect of ICRF-187
pretreatment against doxorubicin-induced delayed cardiotoxicity in
the rat. Toxicol Appl Pharmacol. 102:292–299. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bagai A, Schulte PJ, Granger CB, Mahaffey
KW, Christenson RH, Bell G, Lopes RD, Green CL, Lincoff AM,
Armstrong PW and Roe MT: Prognostic implications of creatine
kinase-MB measurements in ST-segment elevation myocardial
infarction patients treated with primary percutaneous coronary
intervention. Am Hear J. 168(503–511): e22014.
|
|
33
|
Pongprot Y, Sittiwangkul R, Charoenkwan P
and Silvilairat S: Use of cardiac markers for monitoring of
doxorubixin-induced cardiotoxicity in children with cancer. J
Pediatr Hematol Oncol. 34:589–595. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Abdel-Raheem IT, Taye A and Abouzied MM:
Cardioprotective effects of nicorandil, a mitochondrial potassium
channel opener against doxorubicin-induced cardiotoxicity in rats.
Basic Clin Pharmacol Toxicol. 113:158–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Desai AS, Lewis EF, Li R, Solomon SD,
Assmann SF, Boineau R, Clausell N, Diaz R, Fleg JL, Gordeev I, et
al: Rationale and design of the treatment of preserved cardiac
function heart failure with an aldosterone antagonist trial: A
randomized, controlled study of spironolactone in patients with
symptomatic heart failure and preserved ejection fraction. Am Hear
J. 162(966–972): e102011.
|
|
36
|
Zhang Y, Chen Y, Zhang M, Tang Y, Xie Y,
Huang X and Li Y: Doxorubicin induces sarcoplasmic reticulum
calcium regulation dysfunction via the decrease of SERCA2 and
phospholamban expressions in rats. Cell Biochem Biophys.
70:1791–1798. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Forgione MA, Cap A, Liao R, Moldovan NI,
Eberhardt RT, Lim CC, Jones J, Goldschmidt-Clermont PJ and Loscalzo
J: Heterozygous cellular glutathione peroxidase deficiency in the
mouse: Abnormalities in vascular and cardiac function and
structure. Circulation. 106:1154–1158. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sharma M, Kishore K, Gupta SK, Joshi S and
Arya DS: Cardioprotective potential of ocimum sanctum in
isoproterenol induced myocardial infarction in rats. Mol Cell
Biochem. 225:75–83. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Traverso N, Menini S, Maineri EP,
Patriarca S, Odetti P, Cottalasso D, Marinari UM and Pronzato MA:
Malondialdehyde, a lipoperoxidation-derived aldehyde, can bring
about secondary oxidative damage to proteins. J Gerontol A Biol Sci
Med Sci. 59:B890–B895. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Anwar S, Khan MA, Sadaf A and Younus H: A
structural study on the protection of glycation of superoxide
dismutase by thymoquinone. Int J Biol Macromol. 69:476–481. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Martin SS and Senior AE: Membrane
adenosine triphosphatase activities in rat pancreas. Biochim
Biophys Acta. 602:401–418. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Tokarska-Schlattner M, Zaugg M, Zuppinger
C, Wallimann T and Schlattner U: New insights into
doxorubicin-induced cardiotoxicity: The critical role of cellular
energetics. J Mol Cell Cardiol. 41:389–405. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Azevedo PS, Minicucci MF, Santos PP, Paiva
SA and Zornoff LA: Energy metabolism in cardiac remodeling and
heart failure. Cardiol Rev. 21:135–140. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Brautbar N, Baczynski R, Carpenter C,
Moser S, Geiger P, Finander P and Massry SG: Impaired energy
metabolism in rat myocardium during phosphate depletion. Am J
Physiol. 242:F699–F704. 1982.PubMed/NCBI
|
|
45
|
Maldonado EN and Lemasters JJ: ATP/ADP
ratio, the missed connection between mitochondria and the Warburg
effect. Mitochondrion. 19:78–84. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Amorim PA, Nguyen TD, Schwarzer M, Mohr
FW, Schrepper A and Doenst T: Myocardial infarction in rats causes
partial impairment in insulin response associated with reduced
fatty acid oxidation and mitochondrial gene expression. J Thorac
Cardiovasc Surg. 140:1160–1167. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kato T, Niizuma S, Inuzuka Y, Kawashima T,
Okuda J, Tamaki Y, Iwanaga Y, Narazaki M, Matsuda T, Soga T, et al:
Analysis of metabolic remodeling in compensated left ventricular
hypertrophy and heart failure. Circ Hear Fail. 3:420–430. 2010.
View Article : Google Scholar
|
|
48
|
Brennan JP, Southworth R, Medina RA,
Davidson SM, Duchen MR and Shattock MJ: Mitochondrial uncoupling,
with low concentration FCCP, induces ROS-dependent cardioprotection
independent of KATP channel activation. Cardiovasc Res. 72:313–321.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gowans GJ and Hardie DG: AMPK: A cellular
energy sensor primarily regulated by AMP. Biochem Soc Trans.
42:71–75. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gao S, Li H, Feng XJ, Li M, Liu ZP, Cai Y,
Lu J, Huang XY, Wang JJ, Li Q, et al: α-Enolase plays a
catalytically independent role in doxorubicin-induced cardiomyocyte
apoptosis and mitochondrial dysfunction. J Mol Cell Cardiol.
79:92–103. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Braissant O, Foufelle F, Scotto C, Dauca M
and Wahli W: Differential expression of peroxisome
proliferator-activated receptors (PPARs): Tissue distribution of
PPAR-alpha, -beta and -gamma in the adult rat. Endocrinology.
137:354–366. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Karbowska J, Kochan Z and Smolenski RT:
Peroxisome proliferator-activated receptor alpha is downregulated
in the failing human heart. Cell Mol Biol Lett. 8:49–53.
2003.PubMed/NCBI
|
|
53
|
Chen WL, Chen YL, Chiang YM, Wang SG and
Lee HM: Fenofibrate lowers lipid accumulation in myotubes by
modulating the PPARα/AMPK/FoxO1/ATGL pathway. Biochem Pharmacol.
84:522–531. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu Z, Puigserver P, Andersson U, Zhang C,
Adelmant G, Mootha V, Troy A, Cinti S, Lowell B, Scarpulla RC and
Spiegelman BM: Mechanisms controlling mitochondrial biogenesis and
respiration through the thermogenic coactivator PGC-1. Cell.
98:115–124. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Finck BN and Kelly DP: PGC-1 coactivators:
Inducible regulators of energy metabolism in health and disease. J
Clin Invest. 116:615–622. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Tang DX, Zhao HP, Pan CS, Liu YY, Wei XH,
Yang XY, Chen YY, Fan JY, Wang CS, Han JY and Li PP: QiShenYiQi
Pills, a compound chinese medicine, ameliorates doxorubicin-induced
myocardial structure damage and cardiac dysfunction in rats. Evid
Based Complement Altern Med. 2013:4805972013. View Article : Google Scholar
|
|
57
|
Perera RK and Nikolaev VO:
Compartmentation of cAMP signalling in cardiomyocytes in health and
disease. Acta Physiol (Oxf). 207:650–662. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Meier S, Andressen KW, Aronsen JM,
Sjaastad I, Hougen K, Skomedal T, Osnes JB, Qvigstad E, Levy FO and
Moltzau LR: PDE3 inhibition by C-type natriuretic peptide-induced
cGMP enhances cAMP-mediated signaling in both non-failing and
failing hearts. Eur J Pharmacol. 812:174–183. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Levy FO: Cardiac PDEs and crosstalk
between cAMP and cGMP signalling pathways in the regulation of
contractility. Naunyn Schmiedebergs Arch Pharmacol. 386:665–670.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Guo L, Luo L, Ju R, Chen C, Zhu L, Li J,
Yu X, Ye C and Zhang D: Carboxyamidotriazole: A novel inhibitor of
both cAMP-phosphodiesterases and cGMP-phosphodiesterases. Eur J
Pharmacol. 746:14–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Li LX, Cheng YF, Lin HB, Wang C, Xu JP and
Zhang HT: Prevention of cerebral ischemia-induced memory deficits
by inhibition of phosphodiesterase-4 in rats. Metab Brain Dis.
26:37–47. 2011. View Article : Google Scholar : PubMed/NCBI
|