Introduction
Myocardial ischemia is a major cause of sudden
mortality worldwide, which arises from decreased coronary blood
flow to the heart due to atherosclerosis or thrombosis (1,2).
Restoration of coronary blood flow to the ischemia-injured
myocardium in time, which is termed reperfusion, is the most common
therapeutic strategy; however, sudden reperfusion to the
ischemia-injured myocardium may induce oxidative stress and cell
apoptosis, thus further exacerbating myocardial injury, which is
known as myocardial ischemia/reperfusion (I/R) injury (3,4).
Myocardial I/R injury is a complex pathophysiological process
associated with a variety of mechanisms, including oxidative
stress, cell apoptosis and inflammatory responses (5–7). The
particular mechanisms underlying the development of myocardial I/R
injury are unclear; however, it has been reported that excessive
production of reactive oxygen species (ROS) is a primary factor
that activates a variety of molecular cascades of apoptosis,
thereby aggravating myocardial I/R injury (8,9).
Therefore, inhibition of ROS production or the scavenging of free
radicals may be potential therapeutic strategy to attenuate I/R
injury.
Melatonin (N-acetyl-5-methoxytryptamine) is an
endogenous circadian hormone that is mainly produced by the pineal
gland (10). Melatonin and its
derivatives possess numerous beneficial effects, including
anti-oxidation and anti-apoptotic properties (11,12).
Additionally, melatonin has powerful anti-oxidant properties,
including scavenging of superfluous free radicals and enhancing the
synthesis of intracellular antioxidative enzymes (13). Previous studies have demonstrated
that melatonin could protect the myocardium against I/R injury by
regulating oxidative stress and apoptosis (14,15);
however, the specific mechanism by which melatonin exerts its
protective effects on the myocardium against I/R injury requires
further investigation.
Nuclear factor erythroid 2-related factor 2 (Nrf2)
is a member of the basic region-leucine zipper transcription
factors (16). Nrf2 is expressed
in a variety of organs, and is involved in the regulation of
oxidative stress by promoting the transcription and expression of
anti-oxidative proteins and phase II detoxifying enzymes, including
heme oxygenase 1 (HO-1), sestrin2 and superoxide dismutase (SOD)
(17). Under normal conditions,
Nrf2 is predominantly expressed in the cell cytoplasm and is bound
to Kelch-like ECH-associated protein 1 (Keap1), an adaptor protein
that facilitates Nrf2 ubiquitination and degradation (18,19).
In the presence of oxidative stress, Nrf2 disassociates from Keap1
and translocates to the nucleus; subsequent binding to antioxidant
response elements (AREs) regulates the expression of target genes
(18,19). Recent studies have suggested that
Nrf2 signaling was associated with the cleavage process of
myocardial I/R injury (20,21)
The Nrf2 signaling pathway was reported to protect against
anoxia/reoxygenation-induced apoptosis of H9c2 cells (22); however, whether Nrf2 signaling is
involved in the protective effects of melatonin against myocardial
I/R injury remains unknown.
Therefore, the present study aimed to investigate
whether melatonin exerts protective effects against myocardial I/R
injury and whether these protective effects are associated with
Nrf2 signaling via a H9c2 cardiomyocyte-simulated I/R injury in
vitro model.
Materials and methods
Cell culture and treatment
The H9c2 cell line Tiancheng Technology (Shanghai,
China), was cultured in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and 1% (v/v)
penicillin/streptomycin at 37°C with 5% CO2.
Simulated ischemia reperfusion (SIR) treatment was
performed as described (23). The
process of SIR treatment was as follows: H9c2 cells (70%
confluence) were plated into glucose- and serum-free DMEM in a
hypoxia airtight gas chamber containing with 95% N2 and
5% CO2 gas mixture at 37°C for 12 h. Subsequently, the
medium was replaced with normal DMEM and cells were cultured for 6
h at 37°C with 5% CO2. H9c2 cells were first treated
with different concentrations of melatonin (50, 100 and 200
µmol/l), respectively for 1 h to select the suitable concentration
of melatonin.
Subsequently, H9c2 cells were randomly divided into
the following groups: Control group, H9c2 cells were treated with
1% ethanol in DMEM without SIR treatment for 1 h; Mel group, cells
were pretreated with melatonin (100 µmol/l) for 1 h; SIR group,
cells were subjected to SIR treatment as aforementioned; SIR + Mel
group, cells were pretreated with melatonin (100 µmol/l) for 1 h
and then were subjected to SIR treatment and the SIR + Mel + Nrf2
small interfering RNA (siRNA) group, in which cells were
transfected with Nrf2 siRNA for 48 h and pretreated with melatonin
(100 µmol/l) for 1 h prior to SIR treatment, Additionally, the SIR
+ Nrf2 siRNA group constituted cells transfected with Nrf2 specific
siRNA for 48 h, which were then subject to SIR treatment. Melatonin
was dissolved in ethanol and further diluted with DMEM to a
concertation of 100 mmol/l (1% ethanol contained in DMEM) prior to
application.
Determination of cell viability
Cell viability was detected using Cell Counting
kit-8 (CCK-8) assay (Jiancheng Bioengineering Institute, Nanjing,
Jiangsu, China). Briefly, H9c2 cells were seeded in 96-well plates
at a density of 1×104 cells per well. Following the
various treatments described above, 10 µl CCK-8 reagent was added
to 100 µl DMEM medium in each well and then cells were incubated
for 2 h at 37°C. The absorbance was measured using a microplate
reader at 450 nm and the results were presented as percentages of
the control values.
Caspase-3 activity analysis
Caspase-3 activity was measured with a caspase-3
Activity Assay kit (Beyotime Institute of Biotechnology, Haimen,
China). Cell lysates were prepared and centrifuged at 12,000 × g
for 10 min at 4°C with lysis buffer (included in kit). The protein
concentration was determined by Bradford's assay and the caspase-3
activity assay was conducted according to the manufacturer's
protocols. The absorbance was measured at 405 nm with a microtiter
plate reader.
Quantitative measurement of free
radical production
The production of intracellular ROS was measured
using a Reactive Oxygen Species Assay kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocols. Following
the various treatments, cells (90% confluence) were treated with
dichloro-dihydro-fluorescein diacetate (10 µM) at 37°C for 15 min.
Following three washes with phosphate-buffered saline (PBS), the
fluorescence was measured with an FSX100 microscope (Olympus
Corporation, Tokyo, Japan) and the fluorescence intensity was
analyzed with ImageJ 1.6.0 software (National Institutes of Health,
Bethesda, MD, USA).
Measurement of malondialdehyde (MDA),
SOD activity and glutathione (GSH) levels
The MDA content, and the activity levels of SOD and
GSH were spectrophotometrically assayed using an MDA Assay, SOD
Activity Assay and GSH Activity Assay kits (Jiancheng
Bioengineering Institute), respectively according to the
manufacturer's protocols. The absorbance of MDA, SOD, and GSH was
spectrophotometrically measured at 532, 550 and 420 nm respectively
with a SpectraMax M5 spectrophotometer (Molecular Devices, LLC,
Sunnyvale, CA, USA).
siRNA transient transfection
For Nrf2 silencing experiments, H9c2 cells (50%
confluence) were transfected with Nrf2-specific siRNA or scramble
siRNA (20 µM) with Micropoly-transfecter™ cell reagent (Micropoly
Biotech, Nantong, China) according to the manufacturer's protocols.
The particular sequences of Nrf2 siRNA were: Sense,
5′-GAGGAUGGGAAACCUUACUTT-3′ and antisense,
5′-AGUAAGGUUUCCCAUCCUCTT-3′. The sequences of the scramble siRNA
were: Sense, 5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′. At 48 h after transfection, the medium
was replaced and cells were utilized for further experiments.
Preparation of protein lysates
H9c2 cells were lysed with radioimmunoprecipitation
assay buffer (Heart Biological Ltd., Hartlepool, UK) on ice for 30
min, and then centrifuged at 12,000 × g for 10 min at 4°C to remove
the insoluble materials. To detect nuclear Nrf2 expression, the
nuclear protein fraction was extracted using a Nuclear and
Cytoplasmic Protein Extraction kit (Yeasen; Shanghai Shengsheng
Biotechnology Co., Ltd., Shanghai, China) according to the
manufacturer's protocols. Following the various treatments, H9c2
cells were harvested and centrifuged at 1,000 × g for 5 min at 4°C;
the supernatant was subsequently discarded. The precipitate was
lysed with the cytoplasmic protein extraction reagent on ice for 30
min and centrifuged at 12,000 × g for 10 min at 4°C. The
supernatant was removed and the pellet was further lysed with the
nuclear protein extraction reagent on ice for 30 min. The nuclear
protein samples and the whole protein samples were mixed with 5X
SDS-PAGE loading buffer and then heated to 100°C for 10 min. In
total, 30 µg protein lysate was separated by SDS-PAGE (Hart
Biological Ltd.) and immunoblotted with specific antibodies.
Western blot analysis
The concentration of the protein samples were
measured via a bicinchoninic acid protein assay. A total of 30 µg
protein was separated by 10% SDS-PAGE and transferred to
polyvinylidene difluoride membranes. The membranes were incubated
with 10% skimmed milk for 1 h at 37°C and then incubated with
specific primary antibodies overnight at 4°C. The next day, the
membranes were washed three times with Tris-buffered saline with 1%
Tween-20 and incubated with appropriate secondary antibodies for 1
h at room temperature. The bands were detected by chemiluminescence
using the FluorChem FC2 system (Alpha Innotech, San Leandro, CA,
USA) and the results were analyzed with Quantity One 4.62 software
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Caspase-3
(sc-271759; 1:1,000), cleaved caspase-3 (sc-271759; 1:1,000), Nrf2
(sc-30915; 1:1,000), HO-1 (sc-390991; 1:1,000), lamin A (sc-71481;
1:1,000) and β-actin (sc-70319; 1:1,000) were purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). Goat anti-rabbit/mouse
IgG secondary antibody (BA1054 and BA1051; Wuhan Boster Biological
Technology, Ltd., Wuhan, China) were diluted at 1:3,000. Western
blot analysis was repeated three times.
Measurement of apoptosis
Flow cytometry was performed to determine the levels
of apoptosis of cells subject to various treatments using Annexin
V-PI Apoptosis Assay kit (BioVision, Inc., Milpitas, CA, USA)
according to the manufacture's protocols. Briefly, H9c2 cells were
seeded in 6-well plates at 5×104 cells per well.
Following the aforementioned treatments, cells were trypsinized and
resuspended at 5×105 cells/ml in ice-cold PBS. The cells
were washed three times with saline and then were transferred to 1
ml binding buffer (included in kit), and then 5 µl Annexin V-Cy5
and 10 µl propidium iodide (PI) were added. Following thorough
mixing, the stained cells were incubated for 15 min in the dark.
The samples were measured with a FACScan flow cytometer (BD
FACSAria; BD Biosciences, Franklin Lakes, NJ, USA). For each
sample, 1×104 cells were analyzed. The data of early
apoptosis cells (Annexin-Cy5+/PI−) were
collected for each analysis. The data were analyzed with FlowJo
7.6.1 software (FlowJo LLC, Ashland, OR, USA).
Statistical analysis
All experiment data were expressed as the mean ±
standard error (n=8 per group). Comparisons between groups were
conducted with using one-way analysis of variance followed by a
Bonferroni post-hoc test using GraphPad Prism 6 software (La Jolla,
CA, USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Melatonin attenuates SIR-induced
apoptosis of H9c2 cells
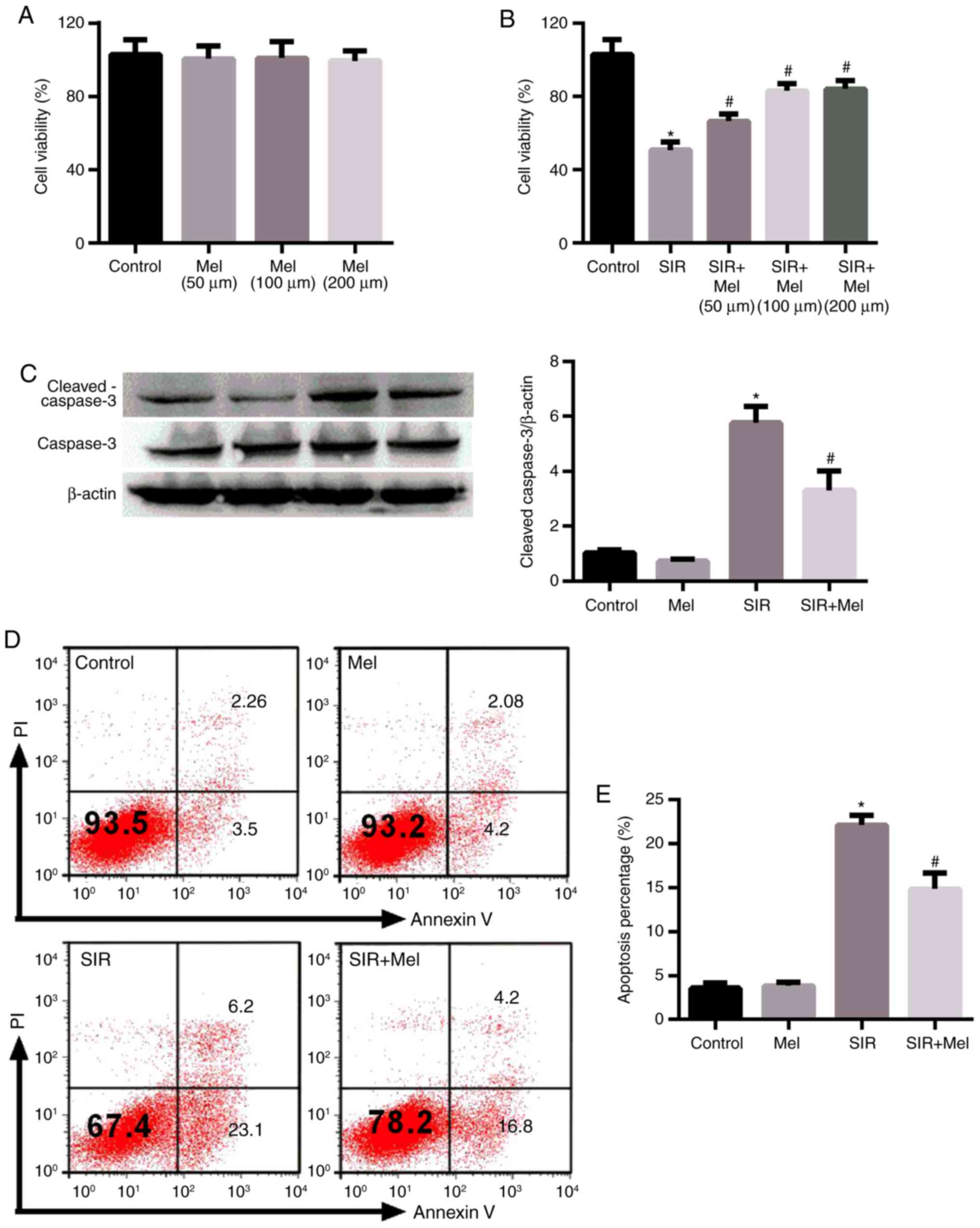
To investigate the effects of melatonin on cell
apoptosis following SIR, cell viability was analyzed. Various
concentrations of melatonin (50, 100 and 200 µmol/l) alone
exhibited no significant effects on the viability of H9c2 cells, as
determined by a CCK-8 assay (Fig.
1A). Conversely, SIR treatment significantly decreased the
viability of H9c2 cells compared with in the control; melatonin
significantly increased the cell viability of SIR-injured H9c2
compared with SIR treatment alone (Fig. 1B). The pro-survival effects of
melatonin did not reveal a significant difference between
treatments with 100 and 200 µmol/l. Therefore, 100 µmol/l was
selected to investigate the protective effects of melatonin in the
present study. To determine the effects of melatonin on cell
apoptosis, western blot analysis of cleaved caspase 3 level and
flow cytometry were conducted. As presented in Fig. 1C-E, consistent with the results of
the cell viability experiments (Fig.
1A and B), SIR treatment significantly promoted cell apoptosis
compared with in the control and melatonin significantly reversed
this alteration induced by SIR.
Melatonin reverses SIR-induced
oxidative stress in H9c2 cells
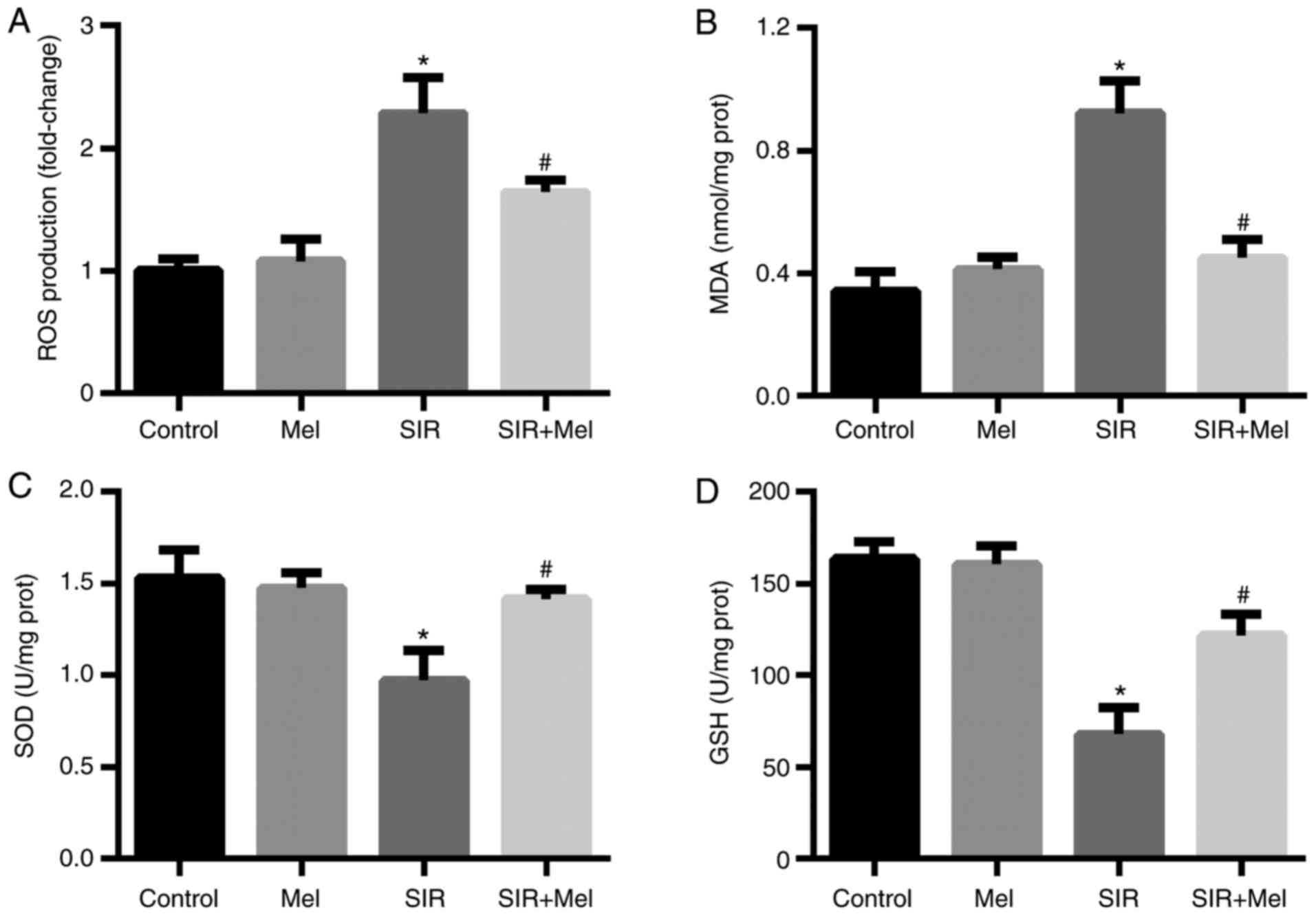
Additionally, the present study further investigated
the effects of melatonin on oxidative stress in H9c2 cells
subjected to SIR. The results revealed that SIR treatment induced a
significant increase in ROS production and MDA content compared
with in the control group (Fig. 2A and
B), while the levels of SOD and GSH were significantly
decreased following SIR compared with the control group (Fig. 2C and D). Pretreatment with
melatonin significantly reduced the effects of SIR on oxidative
stress (Fig. 2).
Melatonin attenuates SIR-induced
apoptosis and oxidative stress via activation of the Nrf2 signaling
pathway
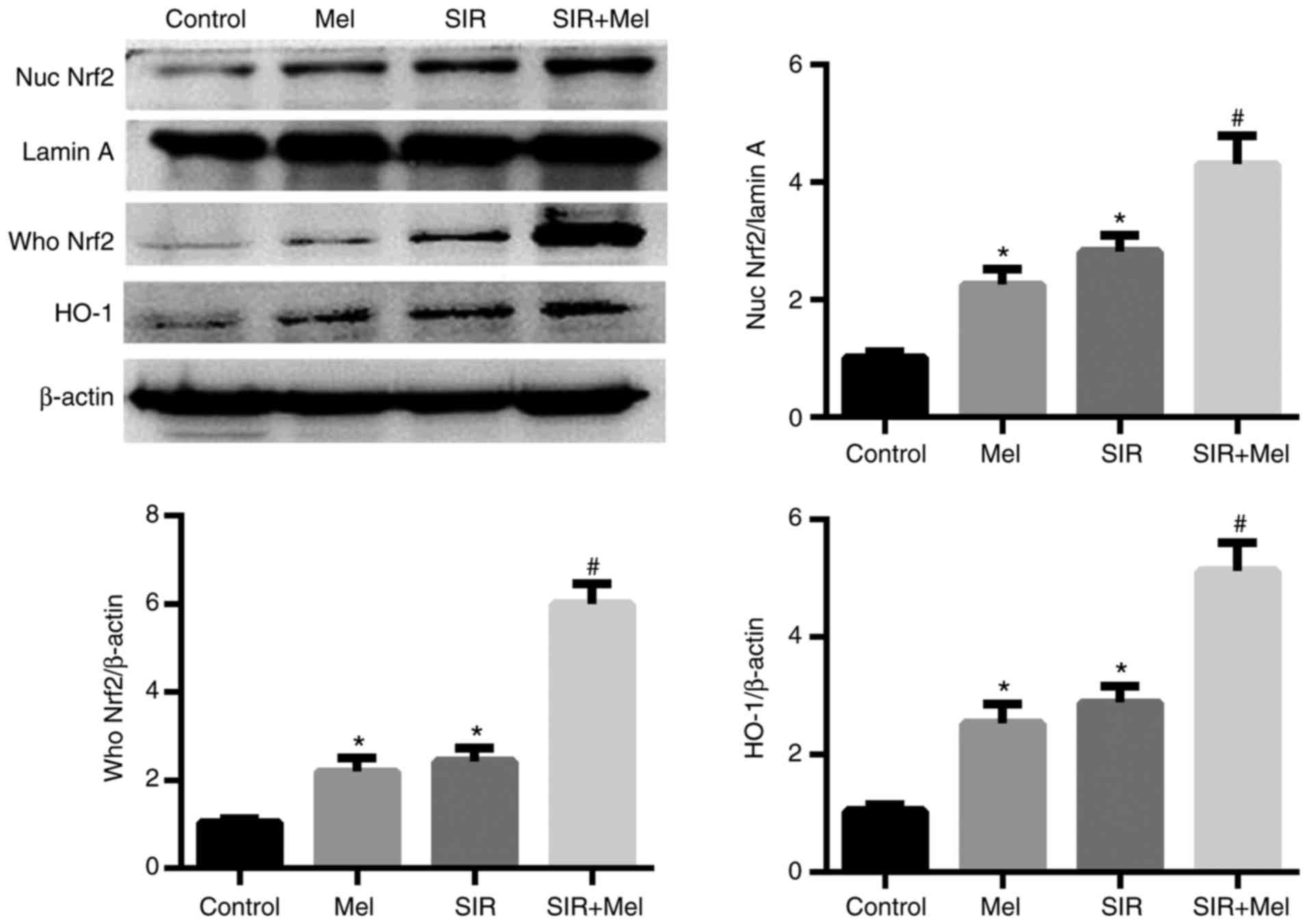
To investigate whether melatonin attenuates
SIR-induced apoptosis and oxidative stress via the Nrf2 signaling
pathway, the whole cell and nuclear lysates of the cell groups were
prepared for immunoblotting. As presented in Fig. 3, SIR induced a significant increase
in Nrf2 expression levels within the nuclear and whole H9c2 cell
lysates. Compared with in the SIR group, pretreatment with
melatonin significantly increased the nuclear and whole cell
expression levels of Nrf2. In addition, the present study analyzed
the expression levels of HO-1, an important target gene of Nrf2
(24). Consistent with that of
Nrf2, the expression levels of HO-1 were significantly increased
when subjected to SIR, which further increased in response to
pretreatment with melatonin. Interestingly, the results of
treatment with melatonin alone also induced a significant increase
in the expression levels of nuclear and whole lysate Nrf2, and HO-1
(Fig. 3).
Inhibition of the Nrf2 signaling
pathway eliminates the anti-apoptotic effects of melatonin on H9c2
cells following SIR
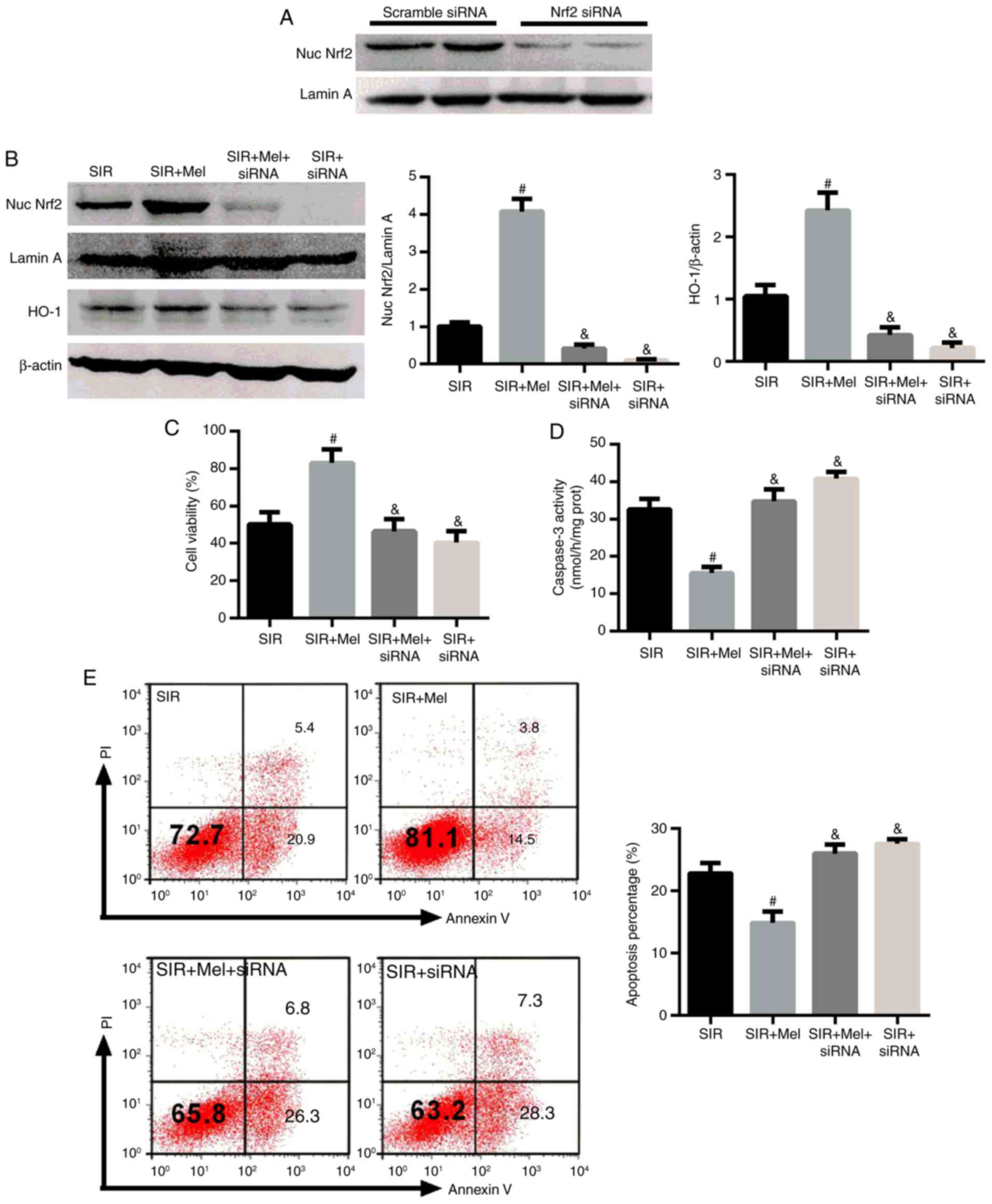
To further verify the role of Nrf2 in the protective
effects of melatonin on SIR treated H9c2 cells, a specific siRNA
targeting Nrf2 was employed for the knockdown of its expression.
Initially, the knockdown capacity of Nrf2 siRNA was determined. As
presented in Fig. 4A, the
expression levels of nuclear Nrf2 were markedly reduced in response
to Nrf2 siRNA; melatonin was suggested to increase the expression
levels of nuclear Nrf2 and HO-1 (Fig.
3). However, the effects of melatonin on the expression levels
of nuclear Nrf2, and its target gene HO-1, were also significantly
reduced by Nrf2 siRNA (Fig. 4B).
In addition, when the Nrf2 signaling pathway was inhibited by Nrf2
siRNA, cell viability significantly decreased and the caspase-3
activity of H9c2 cells was significantly elevated compared with in
the SIR + Mel group (Fig. 4C and
D). Cell apoptosis was also analyzed by flow cytometry
(Fig. 4E). This indicated that the
effects of melatonin on cell viability and apoptosis were abolished
when Nrf2 signaling was inhibited. The data from the Control and
Melatonin groups were not analyzed to avoid any overlap as escribed
previously (25). These results
confirmed that Nrf2 signaling may serve a crucial role in the
protective effects of melatonin on SIR-induced apoptosis in H9c2
cells.
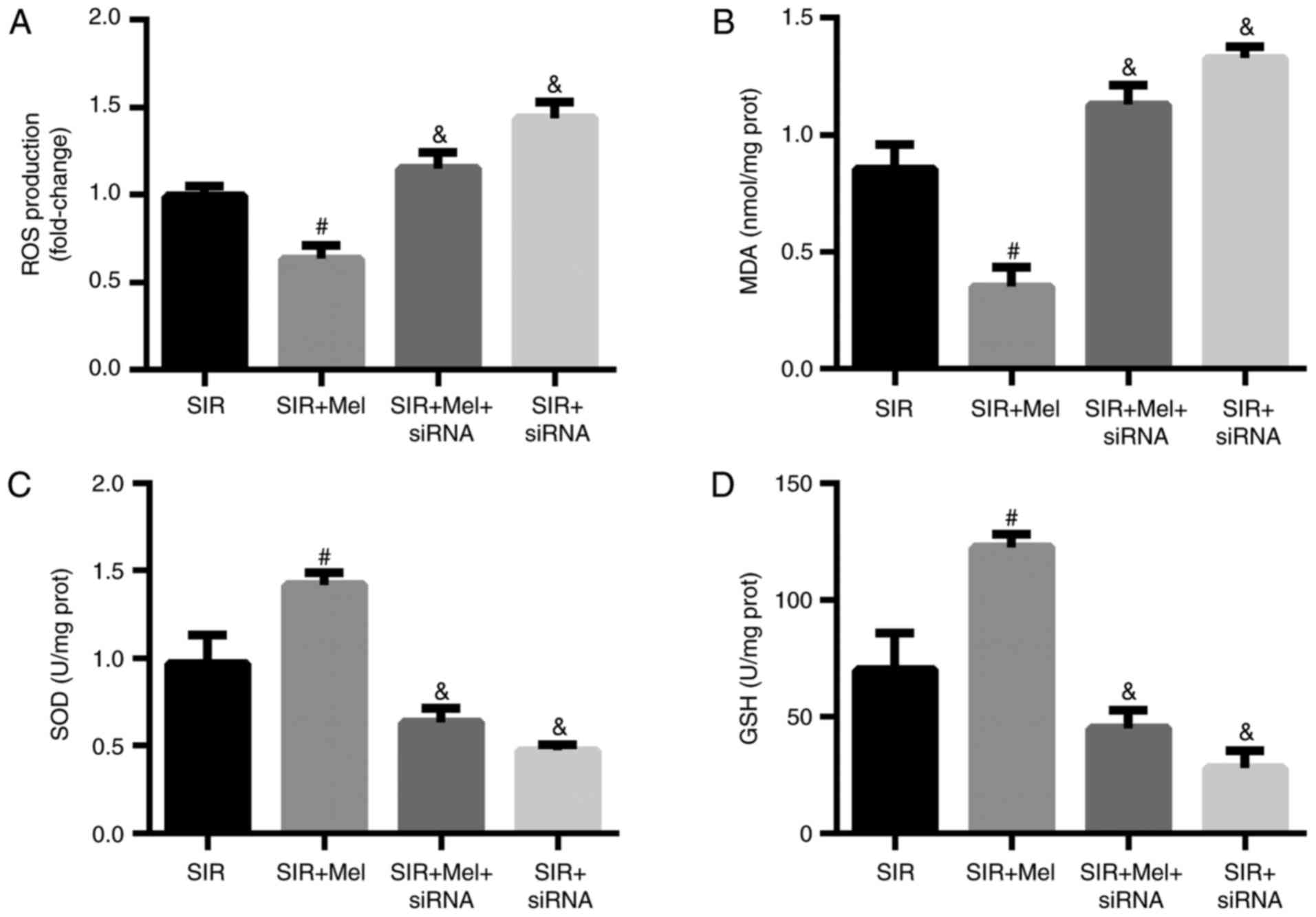
Inhibition of the Nrf2 signaling
pathway eliminates the anti-oxidative effects of melatonin on H9c2
cells following SIR
The present study investigated alterations in
oxidative stress within H9c2 cells expressing Nrf2 siRNA. The
results revealed that Nrf2 siRNA significantly attenuated the
anti-oxidative effects of melatonin on SIR-treated H9c2 cells as
demonstrated by elevated ROS production and MDA content, decreased
SOD activity and GSH expression (Fig.
5). These results suggested that Nrf2 signaling may serve a
crucial role in mitigating SIR-induced oxidative stress in H9c2
cells.
Discussion
Melatonin is a multifunctional and ubiquitous
hormone that exhibits potent cardioprotective effects due to its
notable antioxidant ability (14).
Studies have demonstrated that the administration of melatonin may
alleviate several cardiovascular diseases, including myocardial I/R
injury (26), atherosclerosis
(27) and hypertension (28); however, the specific mechanism by
which melatonin protects the myocardium against I/R injury requires
further investigation. In the present study, H9c2 cardiomyocytes
subjected to SIR treatment were used to mimic in vivo
myocardial I/R injury. Consistent with a previous study (4), SIR induced significant apoptosis and
oxidative stress; melatonin pretreatment may have protected H9c2
cells against SIR-induced apoptosis and oxidative stress as
reported in the present study. To the best of our knowledge, the
present study is the first to demonstrate that the protective
effects of melatonin on SIR-induced H9c2 cells may occur via the
activation of the Nrf2 signaling pathway.
It has been reported that myocardial I/R injury
resulted in cellular oxidative stress responses, which further led
to lipid peroxidation, Ca2+ overload, protein oxidation
and myocardial apoptosis (29).
Additionally, it has been demonstrated that melatonin has notable
myocardial protective effects due to its potent anti-oxidant and
anti-apoptotic properties (15).
In accordance with these studies, the present study revealed that
SIR treatment significantly reduced the cell viability and enhanced
the extent of apoptosis of H9c2 cells. Furthermore, oxidative
stress was investigated in the present study, which indicated that
SIR induced significant increases in ROS and MDA production, but
decreased SOD activity and GSH expression; however, pretreatment
with melatonin revealed that the effects of SIR treatment in H9c2
cells were all reversed. These results indicated melatonin may
protect H9c2 cells from I/R injury via inhibition of oxidative
stress and apoptosis.
The Nrf2 signaling pathway serves a crucial role in
reducing oxidative stress-induced injury (30,31).
Under normal conditions, Nrf2 is mainly located in the cytoplasm;
under conditions of oxidative stress, Nrf2 translocates to the
nucleus, binds to AREs and then initiates the activation of the
endogenous anti-oxidative response to inhibit oxidative stress
(32,33). Activation of Nrf2 signaling has
been reported to increase cell anti-oxidative ability (34). HO-1 is a target gene of Nrf2, which
is involved in regulating cellular ROS production and scavenging
(35,36). Activation of Nrf2-HO-1 signaling
may alleviate myocardial I/R injury (37). In the present study, SIR treatment
was observed to enhance the expression of Nrf2 in the nucleus and
the whole cell, which may suggest increased nuclear translocation.
As the expression of Nrf2 in whole lysate and nucleus was not
directly compared, the increased nuclear translocation of Nrf2 may
be likely; however, further investigation is required. However,
pretreatment with melatonin further promoted Nrf2 translocation, as
well as the expression of the downstream gene, HO-1, which encodes
an enzyme involved in heme metabolism. In the present study, only
the expression levels of HO-1 were analyzed; however, analysis of
the activity of HO-1 may improve understanding of its role in Nrf2
signaling. In addition, the present study observed that
pretreatment with melatonin alone without SIR also promoted Nrf2
nuclear transport, which further demonstrated the protective effect
of melatonin. These results indicated that the protective effects
of melatonin on SIR-induced H9c2 cells may occur via activation of
the Nrf2 signaling pathway.
To further verify the effects of Nrf2 on SIR-induced
H9c2 cells pretreated with melatonin, a specific siRNA targeting
Nrf2 was employed in the present study. The inhibition of Nrf2
signaling eliminated the anti-apoptotic and anti-oxidative effects
of melatonin on H9c2 cells in the present study. These results
further suggested that the protective effects of melatonin on
SIR-induced H9c2 cells may be dependent on the Nrf2 signaling
pathway.
However, there were limitations of the present study
as whether melatonin exhibited its protective effects on myocardial
I/R injury via activation of the Nrf2 signaling pathway was not
investigated in vivo. Thus, the mechanism underlying the
effects of melatonin in activating the Nrf2 signaling pathway
requires further study.
In conclusion, the present study proposed that
melatonin may protect H9c2 cells against I/R injury by reducing
apoptosis and oxidative stress, and this effect may be mediated via
the activation of the Nrf2 signaling pathway, suggesting that
melatonin may be considered as a potential therapeutic target for
the prevention of myocardial I/R injury.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
YZ and HZ designed the study. YZ and HW conducted
the experiments. BQ and FG performed the statistical analysis. HW
and SM contributed to the interpretation of the experimental
results, produced the figures and wrote the manuscript. HZ revised
and edited the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhang X, Hu H, Luo J, Deng H, Yu P, Zhang
Z, Zhang G, Shan L and Wang Y: A novel
danshensu-tetramethylpyrazine conjugate DT-010 provides
cardioprotection through the PGC-1α/Nrf2/HO-1 pathway. Biol Pharm
Bull. 40:1490–1498. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hu T, Wei G, Xi M, Yan J, Wu X, Wang Y,
Zhu Y, Wang C and Wen A: Synergistic cardioprotective effects of
Danshensu and hydroxysafflor yellow A against myocardial
ischemia-reperfusion injury are mediated through the Akt/Nrf2/HO-1
pathway. Int J Mol Med. 38:83–94. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang C, He S, Li Y, Li F, Liu Z, Liu J
and Gong J: Bisoprolol protects myocardium cells against
ischemia/reperfusion injury by attenuating unfolded protein
response in rats. Sci Rep. 7:118592017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu J, Sui H, Zhao J and Wang Y: Osmotin
protects H9c2 cells from simulated ischemia-reperfusion injury
through AdipoR1/PI3K/AKT signaling pathway. Front Physiol.
8:6112017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sinning C, Westermann D and Clemmensen P:
Oxidative stress in ischemia and reperfusion: Current concepts,
novel ideas and future perspectives. Biomark Med. 11:11031–11040.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lim SH and Lee J: Xyloglucan intake
attenuates myocardial injury by inhibiting apoptosis and improving
energy metabolism in a rat model of myocardial infarction. Nutr
Res. 45:19–29. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Toldo S, Marchetti C, Mauro AG, Chojnacki
J, Mezzaroma E, Carbone S, Zhang S, Van Tassell B, Salloum FN and
Abbate A: Inhibition of the NLRP3 inflammasome limits the
inflammatory injury following myocardial ischemia-reperfusion in
the mouse. Int J Cardiol. 209:215–220. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu SZ, Tao LY, Wang JN, Xu ZQ, Wang J, Xue
YJ, Huang KY, Lin JF, Li L and Ji KT: Amifostine pretreatment
attenuates myocardial ischemia/reperfusion injury by inhibiting
apoptosis and oxidative stress. Oxid Med Cell Longev.
2017:41308242017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu YF, Chu YY, Zhang XZ, Zhang M, Xie FG,
Zhou M, Wen HH and Shu AH: TGFβ1 protects myocardium from apoptosis
and oxidative damage after ischemia reperfusion. Eur Rev Med
Pharmacol Sci. 21:1551–1558. 2017.PubMed/NCBI
|
|
10
|
Baltatu OC, Amaral FG, Campos LA and
Cipolla-Neto J: Melatonin, mitochondria and hypertension. Cell Mol
Life Sci. 74:3955–3964. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo Y, Sun J, Li T, Zhang Q, Bu S, Wang Q
and Lai D: Melatonin ameliorates restraint stress-induced oxidative
stress and apoptosis in testicular cells via NF-κB/iNOS and
Nrf2/HO-1 signaling pathway. Sci Rep. 7:95992017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang Z, Zhou F, Dou Y, Tian X, Liu C, Li
H, Shen H and Chen G: Melatonin alleviates intracerebral
hemorrhage-induced secondary brain injury in rats via suppressing
apoptosis, inflammation, oxidative stress, DNA damage, and
mitochondria injury. Transl Stroke Res. 9:74–91. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bai XZ, He T, Gao JX, Liu Y, Liu JQ, Han
SC, Li Y, Shi JH, Han JT, Tao K, et al: Melatonin prevents acute
kidney injury in severely burned rats via the activation of SIRT1.
Sci Rep. 6:321992016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu L, Gong B, Duan W, Fan C, Zhang J, Li
Z, Xue X, Xu Y, Meng D, Li B, et al: Melatonin ameliorates
myocardial ischemia/reperfusion injury in type 1 diabetic rats by
preserving mitochondrial function: Role of AMPK-PGC-1α-SIRT3
signaling. Sci Rep. 7:413372017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu L, Sun Y, Cheng L, Jin Z, Yang Y, Zhai
M, Pei H, Wang X, Zhang H, Meng Q, et al: Melatonin
receptor-mediated protection against myocardial
ischemia/reperfusion injury: Role of SIRT1. J Pineal Res.
57:228–238. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lian Y, Xia X, Zhao H and Zhu Y: The
potential of chrysophanol in protecting against high fat-induced
cardiac injury through Nrf2-regulated anti-inflammation,
anti-oxidant and anti-fibrosis in Nrf2 knockout mice. Biomed
Pharmacother. 93:1175–1189. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dludla PV, Muller CJ, Joubert E, Louw J,
Essop MF, Gabuza KB, Ghoor S, Huisamen B and Johnson R: Aspalathin
protects the heart against hyperglycemia-induced oxidative damage
by Up-regulating Nrf2 expression. Molecules. 22:pii: E129. 2017.
View Article : Google Scholar
|
|
18
|
Bai Y, Chen Q, Sun YP, Wang X, Lv L, Zhang
LP, Liu JS, Zhao S and Wang XL: Sulforaphane protection against the
development of doxorubicin-induced chronic heart failure is
associated with Nrf2 Upregulation. Cardiovasc Ther. 35:2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zeng C, Zhong P, Zhao Y, Kanchana K, Zhang
Y, Khan ZA, Chakrabarti S, Wu L, Wang J and Liang G: Curcumin
protects hearts from FFA-induced injury by activating Nrf2 and
inactivating NF-κB both in vitro and in vivo. J Mol Cell Cardiol.
79:1–12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li W, Wu M, Tang L, Pan Y, Liu Z, Zeng C,
Wang J, Wei T and Liang G: Novel curcumin analogue 14p protects
against myocardial ischemia reperfusion injury through
Nrf2-activating anti-oxidative activity. Toxicol Appl Pharmacol.
282:175–183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cui G, Luk SC, Li RA, Chan KK, Lei SW,
Wang L, Shen H, Leung GP and Lee SM: Cytoprotection of baicalein
against oxidative stress-induced cardiomyocytes injury through the
Nrf2/Keap1 pathway. J Cardiovasc Pharmacol. 65:39–46. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen RC, Sun GB, Wang J, Zhang HJ and Sun
XB: Naringin protects against anoxia/reoxygenation-induced
apoptosis in H9c2 cells via the Nrf2 signaling pathway. Food Funct.
6:1331–1344. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Han H, Hu J, Yan Q, Zhu J, Zhu Z, Chen Y,
Sun J and Zhang R: Bone marrow-derived mesenchymal stem cells
rescue injured H9c2 cells via transferring intact mitochondria
through tunneling nanotubes in an in vitro simulated
ischemia/reperfusion model. Mol Med Rep. 13:1517–1524. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu J, Li C, Li Z, Yang C, Lei L, Ren W, Su
Y and Chen C: Protective effects of oxymatrine against
lipopolysaccharide/D-galactosamine-induced acute liver failure
through oxidative damage, via activation of Nrf2/HO-1 and
modulation of inflammatory TLR4-signaling pathways. Mol Med Rep.
17:1907–1912. 2018.PubMed/NCBI
|
|
25
|
Zhai M, Li B, Duan W, Jing L, Zhang B,
Zhang M, Yu L, Liu Z, Yu B, Ren K, et al: Melatonin ameliorates
myocardial ischemia reperfusion injury through SIRT3-dependent
regulation of oxidative stress and apoptosis. J Pineal Res.
63:2017. View Article : Google Scholar
|
|
26
|
Yeung HM, Hung MW and Fung ML: Melatonin
ameliorates calcium homeostasis in myocardial and
ischemia-reperfusion injury in chronically hypoxic rats. J Pineal
Res. 45:373–382. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hu ZP, Fang XL, Fang N, Wang XB, Qian HY,
Cao Z, Cheng Y, Wang BN and Wang Y: Melatonin ameliorates vascular
endothelial dysfunction, inflammation, and atherosclerosis by
suppressing the TLR4/NF-κB system in high-fat-fed rabbits. J Pineal
Res. 55:388–398. 2013.PubMed/NCBI
|
|
28
|
Shao G, Zhang S, Nie J, Li J and Tong J:
Effects of melatonin on mechanisms involved in hypertension using
human umbilical vein endothelial cells. J Toxicol Environ Health A.
80:1342–1348. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo Y, Li Z, Shi C, Li J, Yao M and Chen
X: Trichostatin A attenuates oxidative stress-mediated myocardial
injury through the FoxO3a signaling pathway. Int J Mol Med.
40:999–1008. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Satta S, Mahmoud AM, Wilkinson FL,
Alexander Yvonne M and White SJ: The role of Nrf2 in cardiovascular
function and disease. Oxid Med Cell Longev. 2017:92372632017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ding Y, Chen M, Wang M, Li Y and Wen A:
Posttreatment with 11-Keto-β-boswellic acid ameliorates cerebral
ischemia-reperfusion injury: Nrf2/HO-1 pathway as a potential
mechanism. Mol Neurobiol. 52:1430–1439. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tang J, Li B, Liu C, Li Q, Wang L, Min J,
Hu M, Hong S and Hong L: Mechanism of mechanical trauma-induced
extracellular matrix remodeling of fibroblasts in association with
Nrf2/ARE signaling suppression mediating TGF-β1/Smad3 signaling
inhibition. Oxid Med Cell Longev. 2017:85243532017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ding Y, Zhang B, Zhou K, Chen M, Wang M,
Jia Y, Song Y, Li Y and Wen A: Dietary ellagic acid improves
oxidant-induced endothelial dysfunction and atherosclerosis: Role
of Nrf2 activation. Int J Cardiol. 175:508–514. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jin Y, Huang ZL, Li L, Yang Y, Wang CH,
Wang ZT and Ji LL: Quercetin attenuates toosendanin-induced
hepatotoxicity through inducing the Nrf2/GCL/GSH antioxidant
signaling pathway. Acta Pharmacol Sin. 19–Jun;2018.(Epub ahead of
print). View Article : Google Scholar
|
|
35
|
Zhou Y, Zhang J, Lei B, Liang W, Gong J,
Zhao C, Yu J, Li X, Tang B and Yuan S: DADLE improves hepatic
ischemia/reperfusion injury in mice via activation of the Nrf2/HO1
pathway. Mol Med Rep. 16:6214–6221. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ding Y, Chen M, Wang M, Wang M, Zhang T,
Park J, Zhu Y, Guo C, Jia Y, Li Y and Wen A: Neuroprotection by
acetyl-11-keto-β-Boswellic acid, in ischemic brain injury involves
the Nrf2/HO-1 defense pathway. Sci Rep. 4:70022014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee MH, Han MH, Lee DS, Park C, Hong SH,
Kim GY, Hong SH, Song KS, Choi IW, Cha HJ and Choi YH: Morin exerts
cytoprotective effects against oxidative stress in C2C12 myoblasts
via the upregulation of Nrf2-dependent HO-1 expression and the
activation of the ERK pathway. Int J Mol Med. 39:399–406. 2017.
View Article : Google Scholar : PubMed/NCBI
|