Introduction
High altitudes create an environment with lower
atmospheric pressure, compared with that at sea level. It is
reported that there are ~17,000,000 individuals living 3,500 m
above sea level worldwide and encounter hypoxia, which causes
deficiency of red blood cells or hemoglobin, and can reduce the
ability of the body to transfer oxygen to tissues (1–3).
Furthermore, exposure to high altitude can cause digestive system
disease, resulting in severe damage to the intestinal tract
(4).
MicroRNAs (miRNAs), a set of endogenous small
non-coding RNAs with a length of 19–24 nt, exert their functions
through translational inhibition or the degradation of target mRNAs
(5). It has been demonstrated that
miRNAs are key in regulating genes, pathways and various biological
networks (5). Circulating miRNAs
are being intensively investigated for their involvement in various
pathogenic processes and may serve as potential diseases biomarkers
(6,7). The concentrations of circulating
miRNA can be affected by factors, including age and gender, and
environmental factors, including living conditions, residence and
altitude (8). However, the
expression profile and mechanism of miRNAs under hypoxia at high
altitudes remain to be fully elucidated. The advent of genome-wide
technologies, including gene expression microarrays, has made it
possible to obtain a comprehensive view of miRNA alterations
associated with high altitude hypoxia, and the use of
bioinformatics enables analysis of the pathways of the altered
miRNAs. A previous study (9) found
that miRNAs were significantly altered in the Tibetan population,
compared with the Nanjing Han population, which suggested that
miRNAs may function as important regulators in high altitude
hypoxic conditions.
The aim of the present study was to use microarray
analysis to examine alterations in the plasma miRNAs of rats
exposed to hypobaric hypoxia for different durations. In addition,
miRNA functions and pathways were analyzed using Gene Ontology
terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway
analysis. The results may assist in identifying novel targets to
treat high altitude hypoxia-associated disease and provide
biomarkers for diagnosis and prognosis.
Materials and methods
Animals and hypoxic exposure
Adult male Sprague-Dawley rats (n=24) weighing
~180–200 g were purchased from the Academy of Military Medical
Sciences (Beijing, China). The animals were maintained in the
animal house facility of the Key Laboratory of Disaster and
Emergency Rescue Medicine in People's Liberation Army (Tianjin,
China) under a standard environment (altitude 0 m, temperature
25±1°C, humidity 50±10%) with a 12 h light/dark cycle, and were
provided with food and water for 7 days. To investigate the effects
of acute hypobaric hypoxia exposure, the 24 male Sprague-Dawley
rats were randomly divided into four groups (I to IV; n=6). Group I
served as the normoxia group maintained in a standard environment.
Groups II, III and IV served as hypoxia groups, in which the rats
were exposed to simulated hypobaric hypoxia for 24, 48 and 72 h,
respectively, at 25,000 ft (8,000 m; 7.5 mm Hg; 35.4 kPa) in a
specially designed animal decompression chamber in which altitude
was maintained by reducing the ambient barometric pressure, and
temperature and humidity were precisely controlled. The airflow in
the chamber was 2 l/min. The temperature and humidity were
maintained at 20±2°C and 60±10%, respectively. The rate of ascent
to altitude was maintained at a rate of 300 m/min and it took 25–30
min to reach the desired altitude. A slow rate of ascent in
altitude with a gradual decrease in ambient pressure was used, as
this is less likely to induce decompression-induced gas bubbles
during exposure. Experiments and animal care were approved by the
ethics committee of the Affiliated Hospital of the Logistics
University of Chinese People's Armed Police Force (Tianjin,
China).
Blood sample collection
Blood was collected from the inferior cava vein of
rats in the normoxia (control) group, and the rats in groups I–IV
immediately following 24, 48 and 72 h exposure to hypoxia. To
prepare plasma, anticoagulants (EDTA, heparin or sodium citrate)
were added to the blood samples immediately following collection of
the blood to prevent clotting. EDTA plasma (10 ml) contained ~1.7
mg potassium EDTA; heparin plasma (5 ml) containing heparin 1 vial;
sodium citrate plasma, (10 ml) containing 1 ml 0.118 mol/l (3.2%)
citrate solution. The specimens were then centrifuged at 1,500 g
for 10 min at 4°C to avoid hemolysis, and were then decanted and
transferred into RNAase-free Eppendorf tubes as aliquots. The
plasma samples were stored at −80°C until further analysis.
Arterial blood measurements
Arterial blood gases were determined using the
i-STAT system with the CG8+ cartridge (Abbott Point of Care, Inc.,
Princeton, NJ, USA). The arterial blood samples (0.5 ml) were
collected and the following blood parameters were determined: pH,
partial pressure of arterial carbon dioxide (PCO2),
partial pressure of arterial oxygen (PO2), total carbon
dioxide (TCO2, base excess (BE), saturation of arterial
blood oxygen (SaO2), bicarbonate concentration
(HCO3−). The TCO2 was calculated
as follows: TCO2 (mmol/l)=HCO3− +
0.03 PCO2.
Lung wet-to-dry weight (W/D)
ratio
Upon termination of each experiment, the animals
were sacrificed with a lethal dose of i.v. sodium pentobarbital.
The lungs were isolated following chest opening, and the right
superior lobe of the hemi-lung was excised and blotted on filter
paper to remove adherent blood, with extra pulmonary tissue
dissected. The wet weight was determined, followed by drying in an
oven at 50°C for 72 h when a constant weight was achieved. The
water content of the tissue, which was used as an index of
pulmonary edema formation, was calculated as wet weight minus dry
weight, expressed as mg water per mg dry tissue.
Lung histology
The remainder of right middle lobe lung was immersed
in formalin, embedded in paraffin, cut into 6 µm sections and
stained with hematoxylin and eosin for histological analysis. The
severity of lung injury was inspected using light microscopy (Nikon
Eclipse 50i; Nikon Instruments, Inc., Tokyo, Japan) (10).
RNA isolation
TRIzol was used for total RNA extraction according
to the manufacturer's protocol (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Small RNAs (<200 nt) were
separated from the total RNA using mirVana miRNA purification
columns (Ambion; Thermo Fisher Scientific, Inc.) for microarray
analysis and reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) analysis according to the manufacturer's
protocol. The quality and quantity of each RNA preparation were
determined using a Nanodrop ND-1000 spectrophotometer (Agilent
Technologies, Inc., Santa Clara, CA, USA).
Expression of miRNAs
Each total RNA sample (700 ng) was labeled and
hybridized using a FlashTag™ Biotin HSR RNA labeling kit
(manufactured for Affymetrix, Inc., Santa Clara, CA, USA by
Genisphere LLC, Hatfield, PA, USA). Total RNA was labeled using
poly A polymerase. Biotin-labeled RNAs were hybridized for 16–18 h
at 45°C on an Affymetrix miRNA v2.0 array. GeneChips were washed
and stained in the Affymetrix Fluidics Station 450, and were then
scanned using the Affymetrix GeneChip Scanner 3000 7G. The data
were analyzed using the robust multi-array analysis-detection above
background (RMA-DABG) normalization method, implemented in
Affymetrix® expression console software (version
1.2.0.20; Affymetrix; Thermo Fisher Scientific, Inc.). The
normalized and log-transformed intensity values were analyzed using
Expression Console (Affymetrix; Thermo Fisher Scientific, Inc.).
Fold change filters were set requiring genes to be present in ≥200%
of the controls to be considered an upregulated miRNA and <50%
of controls to be considered a downregulated miRNA.
RT-qPCR analysis
To validate the fold change results of the miRNA
arrays, six miRNAs, comprising three upregulated (miR-25-5p,
miR-451-5p and miR-466b-5p) and three downregulated (miR-214-3p,
miR-140-3p and let-7a-5p) miRNAs, were randomly selected and
examined using RT-qPCR analysis. The RT reaction was performed
using mature miRNA-specific primer sets (Applied Biosystems; Thermo
Fisher Scientific, Inc.) and an microRNA reverse transcription kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. PCR was performed using the 7500 Fast
Real Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.) and the SYBR Premix Ex Taq™ kit (Takara
Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer's protocol. The thermocycling conditions were as
follows: 95°C for 10 min, followed by 35 cycles of 95°C for 15 sec
and 60°C for 1 min. Melting curve analyses were subsequently
performed. The fold change for each miRNA was calculated using the
2−ΔΔCq method (11)
with U6 small nuclear RNA as the endogenous control. All reactions
were performed in triplicate for each sample. The PCR primer
sequences were as follows: The universal reverse primer was:
CCAGTGCAGGGTCCGAGGT; and the forward primer sequences were as
follows: miR-25-5p, TGCGGAGGCGGAGACUUGGG; miR-451-5p,
TGCGGAAACCGUUACCAUUA; miR-466b-5p, TGCGGUGAUGUGUGUGUACA;
miR-214-3p, TGCGGACAGCAGGCACAGAC; miR-140-3p, TGCGGUACCACAGGGUAGAA;
let-7a-5p, TGCGGUGAGGUAGUAGGUUG; and U6,
TGCGGGTGCTCGCTTCGGCAGC.
MicroRNA target prediction
The miRNA target sites were predicted by
computer-aided algorithms obtained from TargetScan (version 7.1;
www.targetscan.org/). The miRNA
sequences were downloaded from the miRBase website (http://www.mirbase.org).
Gene ontology (GO) analysis
Based on the GO database (http://www.geneontology.org), the significant GO terms
of the upregulated miRNA targeted genes were analyzed using the
Database for Annotation, Visualization and Integrated Discovery
(http://david.abcc.ncifcrf.gov/home.jsp) (12). Fisher's exact test and the
Chi-square test were used to classify the GO categories, and the
false discovery rate (FDR) was calculated to correct the P-values
using the Benjamini and Hochberg procedure (13). The P-values of each differentially
expressed gene in all the GO terms were calculated. P<0.05 was
considered to indicate a statistically significant difference.
Pathway analysis
Pathway analysis was performed to determine the
significant pathways of the differentially expressed genes,
according to the KEGG Orthology Based Annotation System (http://kobas.cbi.pku.edu.cn/), in which pathway
enrichment can be analyzed by the KEGG pathway, pathway interaction
database, BioCyc, Reactome and Protein ANalysis THrough
Evolutionary Relationships. Fisher's exact test and a χ2
test were used to select the significant pathway, and the threshold
of significance was defined by the P-value and false discovery rate
(13).
Statistical analysis
Numerical data are presented as the mean ± standard
deviation. Differences between means were analyzed using Student's
t-test. For comparison of multiple groups, one-way analysis of
variance followed by Tukey's post-test was performed. All
statistical analyses were performed using SPSS 13.0 software (SPSS,
Inc., Chicago, IL, USA).
Results
Blood gas analysis
The results of the blood gases analysis confirmed
that, compared with control group, PCO2 and lactate were
markedly increased in the high altitude hypoxia group, whereas
PO2, BE, HCO3−, TCO2
and SaO2 were significantly decreased at 24, 48 and 72 h
in a time-dependent manner (P<0.01). The pH level showed
marginal decrease with no statistically significant difference
(Table I).
 | Table I.Measurements of arterial blood gas
parameters. |
Table I.
Measurements of arterial blood gas
parameters.
|
|
| High altitude
hypoxia (h) |
|---|
|
|
|
|
|---|
| Parameter | Control | 24 | 48 | 72 |
|---|
| pH | 7.396±0.03 | 7.369±0.03 | 7.291±0.04 | 7.263±0.03 |
| PCO2
(mmHg) | 34.4±5.01 | 37.280±5.23 |
39.475±2.46a |
50.5±2.89a–c |
| PO2
(mmHg) | 88.7±3.98 |
61.67±7.12a |
53.33±2.16a,b |
41.17±1.47a–c |
| BE (mmol/l) | 1.67±0.52 |
−8±1.22a |
−8.67±1.51a |
−9.33±0.82a |
|
HCO3− (mmol/l) | 30.717±0.86 |
19.083±1.21a |
18.825±1.33a |
16.25±0.49a |
| TCO2
(mmHg) | 32.67±1.51 |
20±1.41a |
19.17±1.94a |
17.5±1.58a |
| SaO2
(%) | 91.67±1.86 |
67.5±1.8a |
57±2.37a,b |
47.8±2.48a–c |
| Lac (mmol/l) | 0.982±0.36 | 1.488±0.72 | 1.537±0.74 | 1.635±0.81 |
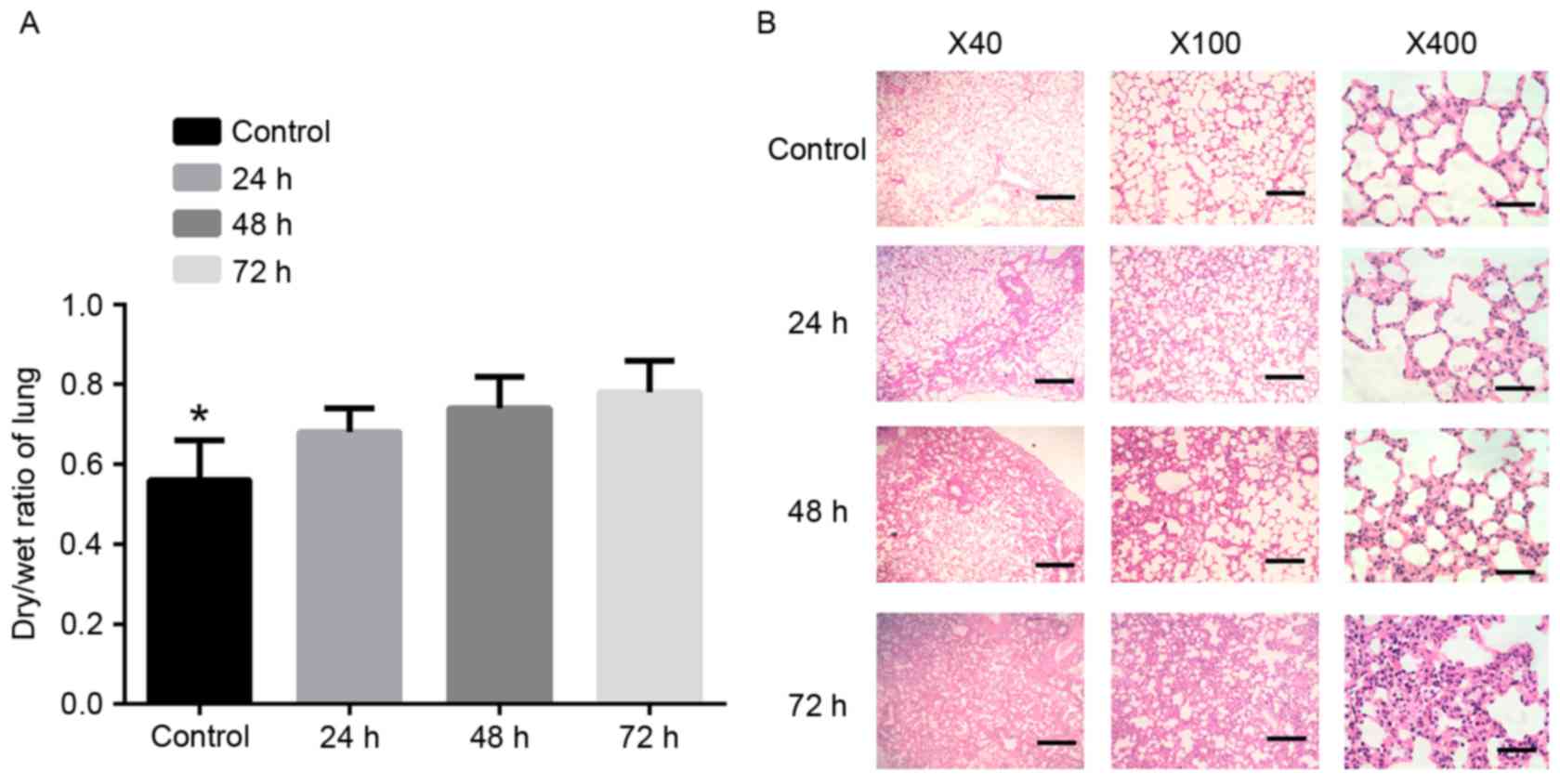
Lung W/D ratio and histology
The W/D lung weight ratio was measured to evaluate
fluid accumulation in the lung specimens (14). Compared with the normal control
groups, the W/D of lung tissues in the hypoxia groups were
significantly upregulated at 24, 48 and 72 h (P<0.01). The W/D
of the lung tissues at 48 and 72 h were higher, compared with that
at 24 h. However, no significant difference in the lung W/D ratio
was observed between the 48 and 72 h time points (Fig. 1A). Using light microscopy, a
detailed examination of lung pathology was performed. Compared with
the control groups, the pulmonary interstitium showed marginal
dilations or hyperemia in the 24 h group. In the 48 h group, the
pulmonary interstitium exhibited increased dilations and
thickening. Blood stasis was also identified in the alveolar space.
The tissues in the 72 h group exhibited marked thickening of the
pulmonary interstitium and visible pink exudant in the alveolar
space. Hyperemia and expansion of the alveolar capillary were also
observed (Fig. 1B).
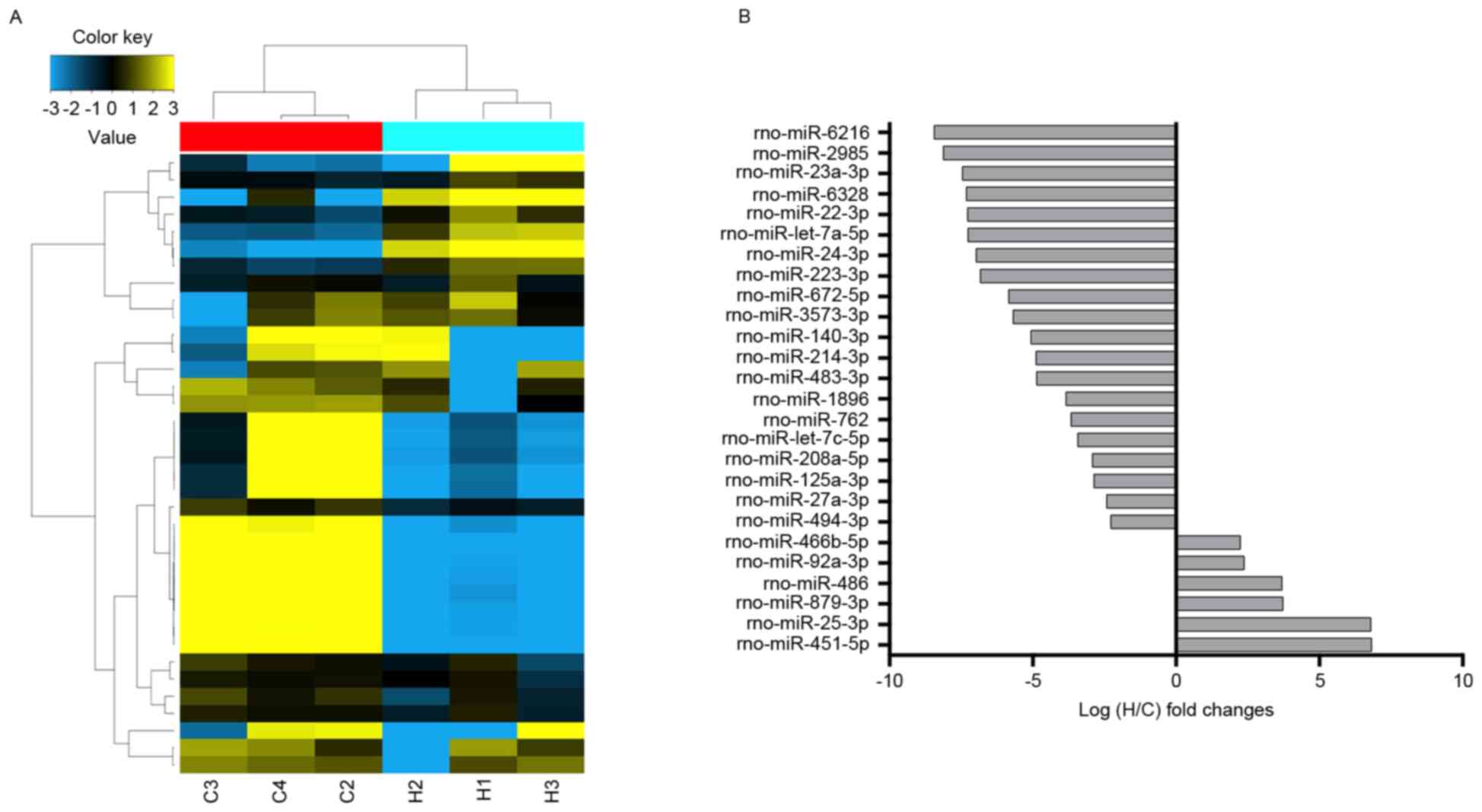
Overview of the miRNAs profiles
From the miRNAs expression profiles, differentially
expressed miRNAs were identified between the high-altitude hypoxia
environment and normal control samples. The miRNA expression
profiles were determined by calculating the log FC in the
high-altitude hypoxia group/normal group. Among a set of
differentially expressed miRNAs, six miRNAs were significantly
upregulated and 20 miRNAs were significantly downregulated in the
high-altitude hypoxia group, compared with the control group.
miR-451-5p, miR-25-3p, miR-879-3p, miR-486, miR-92a-3p and
miR-466b-5p were the identified upregulated miRNAs, whereas
miR-6216, miR-2985, miR-23a-3p, miR-6328, miR-22-3p and let-7a-5p
were the top six significantly downregulated miRNAs (Fig. 2 and Table II).
| Table II.List of miRNAs with altered
expression in high altitude hypoxia. |
Table II.
List of miRNAs with altered
expression in high altitude hypoxia.
| miRNA | Fold change |
|---|
| Upregulated |
|
|
rno-miR-25-3p | 6.796382 |
|
rno-miR-451-5p | 6.811991 |
|
rno-miR-466b-5p | 2.243331 |
|
rno-miR-486 | 3.692567 |
|
rno-miR-879-3p | 3.723761 |
|
rno-miR-92a-3p | 2.373798 |
| Downregulated |
|
|
rno-let-7a-5p | −7.25955 |
|
rno-let-7c-5p | −3.42339 |
|
rno-miR-125a-3p | −2.86652 |
|
rno-miR-140-3p | −5.06208 |
|
rno-miR-1896 | −3.84228 |
|
rno-miR-208a-5p | −2.92098 |
|
rno-miR-214-3p | −4.88104 |
|
rno-miR-22-3p | −7.2768 |
|
rno-miR-223-3p | −6.82643 |
|
rno-miR-23a-3p | −7.45505 |
|
rno-miR-24-3p | −6.97121 |
|
rno-miR-27a-3p | −2.41551 |
|
rno-miR-2985 | −8.11875 |
|
rno-miR-3573-3p | −5.68544 |
|
rno-miR-483-3p | −4.86634 |
|
rno-miR-494-3p | −2.27509 |
|
rno-miR-6216 | −8.4358 |
|
rno-miR-6328 | −7.31127 |
|
rno-miR-672-5p | −5.84322 |
|
rno-miR-762 | −3.6636 |
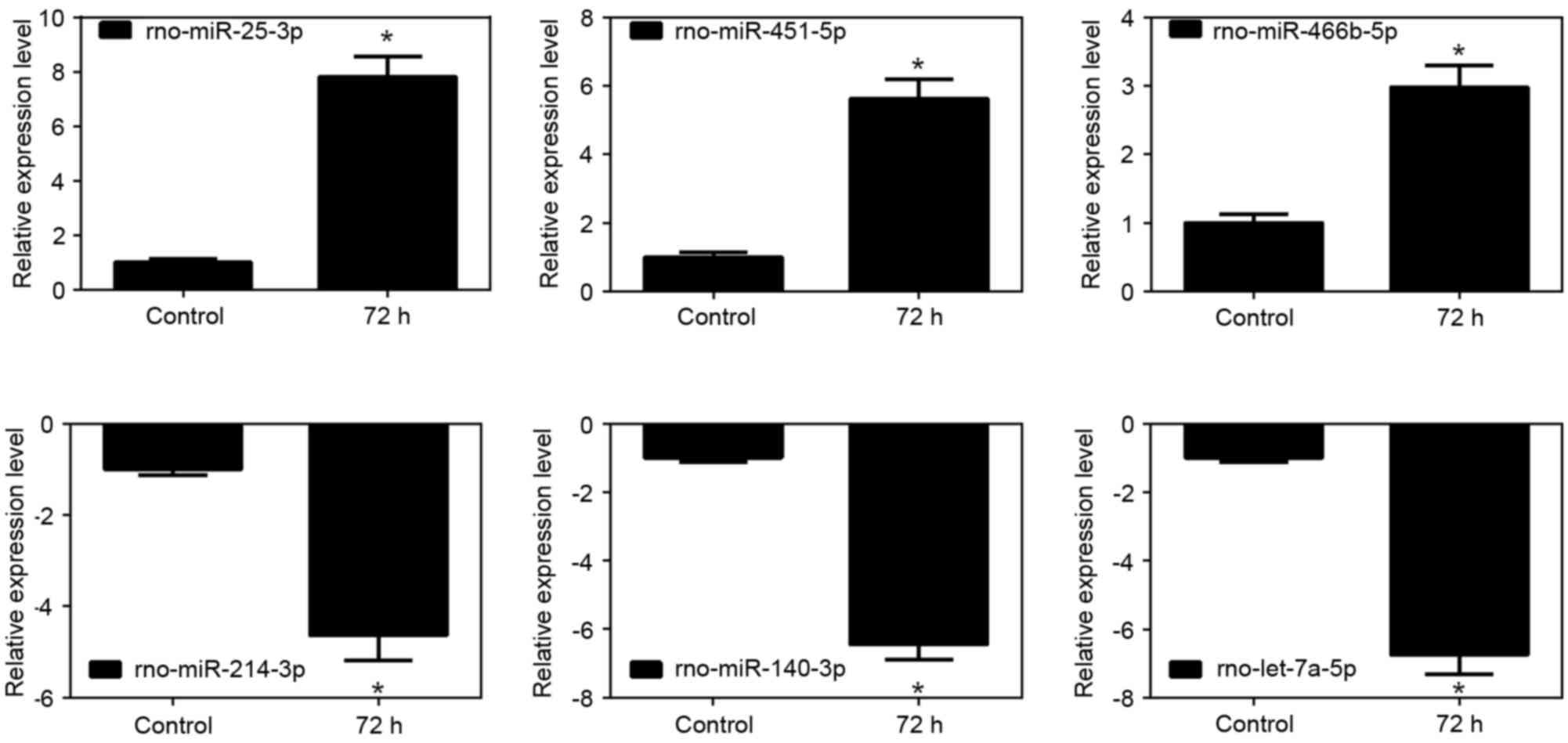
Validation of the microarray analysis
data using RT-qPCR analysis
The relative expression changes of the randomly
selected miRNAs were analyzed using RT-qPCR analysis. The results
of the miR-25-5p, miR-451-5p and miR-466b-5p upregulated miRNAs,
and the miR-214-3p, miR-140-3p and let-7a-5p the downregulated
miRNAs were generally consistent with the microarray analysis
results, as shown in the histograms in Fig. 3 (P<0.05).
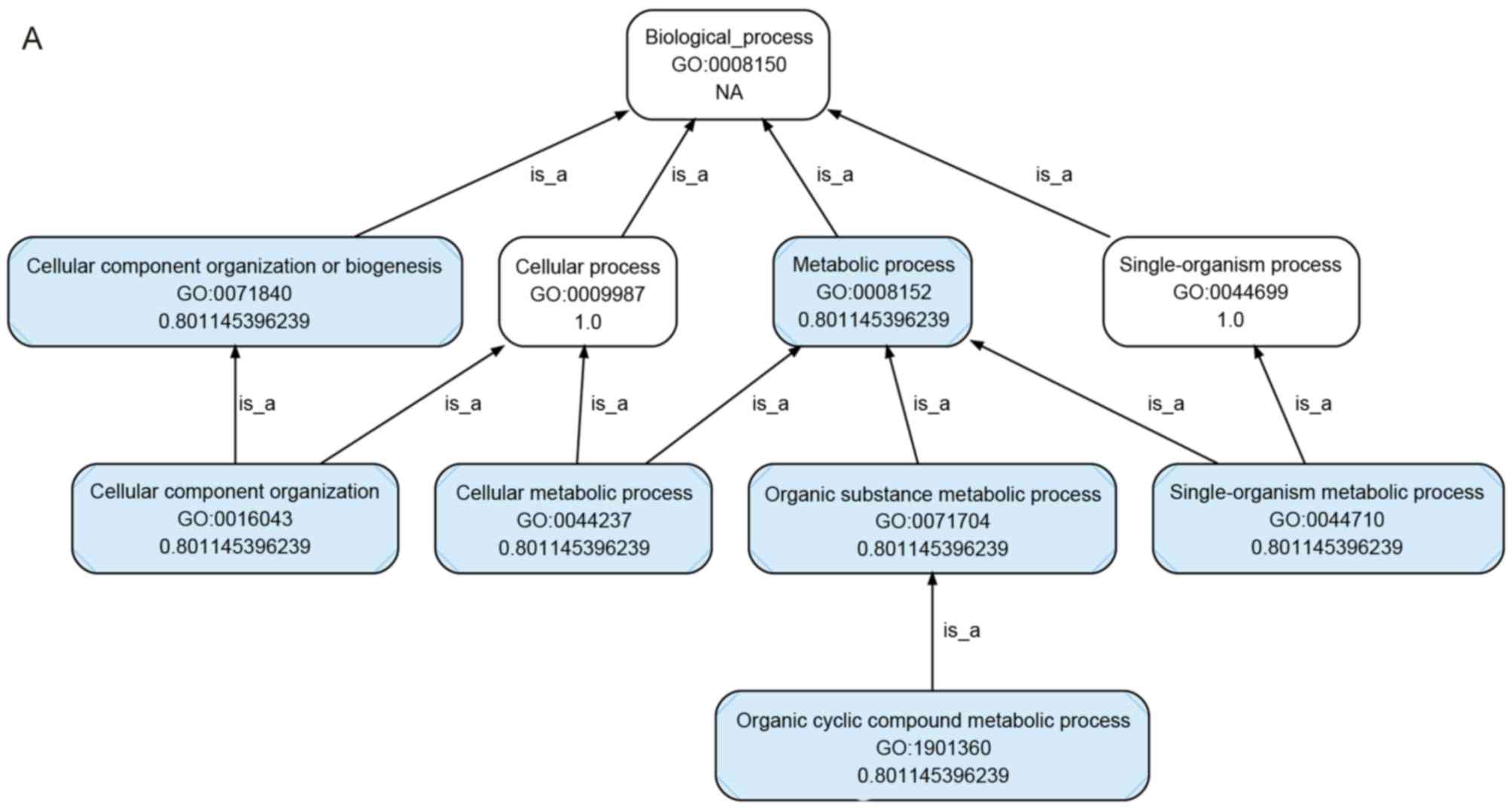
Microarray-based GO analysis
For GO analysis, the target mRNAs for the six
upregulated miRNAs were predicted using TargetScan (data not
shown). As shown in Fig. 4A-D, GO
analysis showed that the targets were involved in several
biological processes, and the top five gene-associated processes
were cellular process, single-organism process, metabolic process,
biological process and regulation of biological process (Fig. 4A). Several genes were involved in
the cellular component, the top five of which were cell, cell part,
organelle, membrane and organelle part (Fig. 4B). Several genes were also involved
in molecular function, the top five of which were binding,
catalytic activity, molecular transducer activity, transporter
activity and molecular function regulator (Fig. 4C and D). These results supported
the hypothesis that these biological processes, cellular components
and molecular functions are important in rats exposed to high
altitude hypoxia.
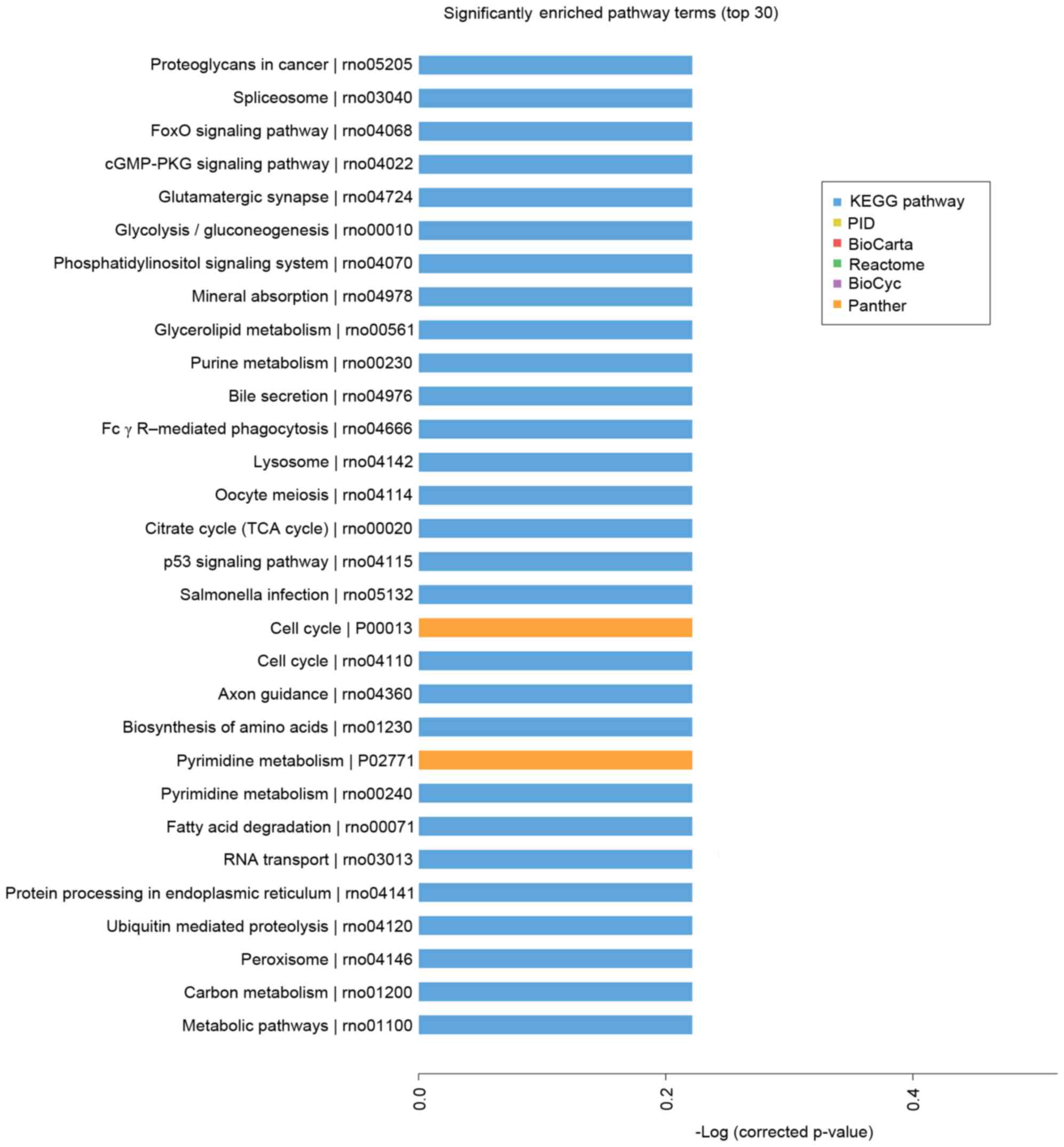
Microarray-based pathway analysis
Pathway-based analysis assists in further
understanding the biological functions of genes. In the present
study, KEGG pathway enrichment analysis was performed to identify
significantly enriched metabolic pathways or signal transduction
pathways in the differentially expressed genes. Using pathway
analysis, which considered the relative change direction and fold
change, and had a the threshold of significance of P<0.05, the
top 30 significant pathways were identified (Fig. 5). The pathways targeted by
upregulated miRNAs with the highest enrichment included the
proteoglycans in cancer, spliceosome, gluamatergic synapse,
glycolysis/gluconeogenesis, Foxo, cGMP-PKG and p53 signaling
pathways, which suggested that these pathways may be involved in
the response to a high altitude hypoxic environment.
Discussion
Understanding the clinical relevance of miRNA
expression patterns in the environment during exposure to high
altitude is necessary to circumvent the therapeutic challenges
faced in clinical management. The present study used bioinformatics
methods to screen the differently expressed miRNAs, and then
analyzed the functions and pathways of the upregulated miRNAs. In
the present study, a total of 26 aberrantly expressed miRNAs were
identified in the blood of rats exposed to high altitude, compared
to the samples from rats in normal conditions. Subsequently,
RT-qPCR analysis was used to confirm the miRNA array results. To
measure the concentrations of miRNAs, three upregulated and three
downregulated miRNAs were selected for RT-qPCR analysis, the
results of which were in accordance with the miRNA array assays.
Among the differently expressed miRNAs, a subset of the altered
miRNAs (miR-24, miR-25, miR-486, miR-451 and miR-92a) has also been
associated with the hypoxic response in mammalian cells in other
studies (15–19), which suggested that a high altitude
hypoxic environment has a marked effect on human plasma miRNA
patterns. The functional investigation of miRNAs, which respond
during hypoxia, may assist in uncovering the molecular basis of
hypoxic acclimatization and elucidate the complexity of hypoxic
response pathways in humans. The present study also found that the
expression of rno-miR-23a-3p was similar with the that reported in
a study by Yan et al (9),
which showed that human miR-23a-3p was downregulated in the Tibetan
population, compared with the Nanjing Han population. This
suggested the species conservation of this miRNA and indicated its
importance for further investigation.
GO is widely recognized as a premier tool for
molecular organization and functional annotation (20). Using the criteria of P<0.05 to
identify significant GO terms, the present study revealed that the
predicted targets of the six upregulated miRNAs were associated
with biological processes, including cellular process,
single-organism process, metabolic process, biological process and
regulation of biological process; cellular components, including
cell, cell part, organelle, membrane and organelle part; and
molecular functions, including binding, catalytic activity,
molecular transducer activity, transporter activity and molecular
function regulator. The above GO terms have also been well
represented in Triplophysa dalaica (21).
Pathway analysis can reveal distinct biological
processes and identify the significant pathways, which dysregulated
mRNAs are involved in. This enables a comprehensive understanding
of the interactions of genes, their functions and the association
between upstream and downstream genes, and can identify genes,
which may be regulated by miRNAs. The appearance of the pathways in
proteoglycans in cancer, spliceosome, gluamatergic synapse,
glycolysis/gluconeogenesis, Foxo, cGMP-PKG and p53 signaling
pathways confirmed their concordance with GO terms and their
critical role in high altitude hypoxia. A significant adaptation
was identified as an increased evolutionary rate and positive
selection of genes involved in the hypoxic response and energy
metabolism, analogous to those observed in other organisms in Tibet
(22). It was previously reported
that a conversion from oxidative glucose metabolism to glycolysis
compensated for insufficient levels of oxygen in hypoxic conditions
(23–25). The present study also showed the
enhanced role of glycolysis. The FOXO subfamily of Forkhead
transcription factors has a role in evolutionary conservation in
cellular adaptation to stress stimuli, including hypoxic conditions
(26). A previous study by Wang
et al (27) showed that
FOXO1 may be essential in adaptation to high altitudes. p53, as an
upstream mediator of p21, may suppress hypoxic human lung
fibroblast proliferation and pulmonary arterial remodeling by
interacting with hypoxia-inducible factor-1 under hypoxia (28,29).
A previous study showed that p53 gene deficiency with a decreased
expression of p21 promoted hypoxia-induced pulmonary hypertension
in mice (30). cGMP-dependent
protein kinase (PKG) is a critical enzyme involved in the
regulation of vascular contractility. Impaired PKG-mediated
signaling has been found to be responsible for reduced
cGMP-mediated pulmonary hypertension and vasodilatation following
acute and chronic hypoxia (31–33).
As these pathways have been identified to be involved in various
hypoxic environments, together with the results of the present
study, these pathways may be critical in high altitude hypoxic
conditions.
In conclusion, the results of the present study
identified six upregulated miRNAs and 20 downregulated miRNAs from
two platforms. As upregulated miRNAs may better serve as
biomarkers, the six upregulated miRNAs were used to perform GO and
pathway analysis, which identified that Foxo, cGMP-PKG and p53 may
be critical in the study model of hypoxia. Based on the integrated
analysis of transcriptome features, these results may provide an
important contribution to future investigations aimed at
characterizing the role of specific miRNAs in the pathogenesis of
high altitude hypoxia-induced diseases, and contribute to improving
diagnosis and treatment.
Acknowledgements
Not applicable.
Funding
This study was funded by the Science and technology
planning project of Tianjin City (grant no. 14ZCDZSY00033), the
Opening fund Key laboratory of Disaster & Emergency Rescue
Medicine in People's Liberation Army (grant no. JY1402), the
Central laboratory opening fund of Logistics University of Chinese
People's Armed Police Forces (grant no. 2015ZXKF01), the Technical
project of logistics equipment of Logistics University of Chinese
People's Armed Police Forces (grant no. WHZ201507), the National
Natural Science Foundation of China (grant no. 81772018), the
Natural Science Foundation of Tianjin City (grant no.
17JCZDJC35400) and the Opening fund of Affiliated Hospital of
Logistics University of Chinese People's Armed Police Force (grant
no. WYKFZ201603).
Availability of data and materials
The analyzed datasets generated during the study are
available from the corresponding author on reasonable request.
Authors' contributions
FC, RJW and GZL performed the experiments. YZ, SY
and XYC analyzed the data. YFL and SKH designed the experiment. SKH
drafted the manuscript and revised it critically to produce the
final approval of the version to be published.
Ethics approval and consent to
participate
Experiments and animal care were approved by the
ethics committee of the Affiliated Hospital of the Logistics
University of Chinese People's Armed Police Force (Tianjin,
China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sherpa LY, Deji, Stigum H,
Chongsuvivatwong V, Thelle DS and Bjertness E: Obesity in Tibetans
aged 30–30 living at different altitudes under the north and south
faces of Mt. Everest. Int J Environ Res Public Health. 7:1670–80.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Windsor JS and Rodway GW: Rodway, Heights
and haematology: The story of haemoglobin at altitude. Postgrad Med
J. 83:148–151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peyssonnaux C, Nizet V and Johnson RS:
Role of the hypoxia inducible factors HIF in iron metabolism. Cell
Cycle. 7:28–32. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Recavarren-Arce S, Ramirez-Ramos A, Gilman
RH, Chinga-Alayo E, Watanabe-Yamamoto J, Rodriguez-Ulloa C, Miyagui
J, Passaro DJ and Eza D: Severe gastritis in the Peruvian Andes.
Histopathology. 46:374–379. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mitchell PS, Parkin RK, Kroh EM, Fritz BR,
Wyman SK, Pogosova-Agadjanyan EL, Peterson A, Noteboom J, O'Briant
KC, Allen A, et al: Circulating microRNAs as stable blood-based
markers for cancer detection. Proc Natl Acad Sci USA.
105:10513–10518. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen X, Ba Y, Ma L, Cai X, Yin Y, Wang K,
Guo J, Zhang Y, Chen J, Guo X, et al: Characterization of microRNAs
in serum: A novel class of biomarkers for diagnosis of cancer and
other diseases. Cell Res. 18:997–1006. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Meder B, Backes C, Haas J, Leidinger P,
Stähler C, Großmann T, Vogel B, Frese K, Giannitsis E, Katus HA, et
al: Influence of the confounding factors age and sex on microRNA
profiles from peripheral blood. Clin Chem. 60:1200–1208. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yan Y, Shi Y, Wang C, Guo P, Wang J, Zhang
CY and Zhang C: Influence of a high-altitude hypoxic environment on
human plasma microRNA profiles. Sci Rep. 5:151562015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Su X, Song Y, Jiang J and Bai C: The role
of aquaporin-1 (AQP1) expression in a murine model of
lipopolysaccharide-induced acute lung injury. Respir Physiol
Neurobiol. 142:1–11. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:P32003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Dupuy D, Bertin N, Hidalgo CA, Venkatesan
K, Tu D, Lee D, Rosenberg J, Svrzikapa N, Blanc A, Carnec A, et al:
Genome-scale analysis of in vivo spatiotemporal promoter activity
in Caenorhabditis elegans. Nat Biotechnol. 25:663–668. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Michel RP, Hakim TS, Smith TT and Poulsen
RS: Quantitative morphology of permeability lung edema in dogs
induced by alpha-naphthylthiourea. Lab Invest. 49:412–419.
1983.PubMed/NCBI
|
|
15
|
Kulshreshtha R, Ferracin M, Wojcik SE,
Garzon R, Alder H, Agosto-Perez FJ, Davuluri R, Liu CG, Croce CM,
Negrini M, et al: A microRNA signature of hypoxia. Mol Cell Biol.
27:1859–1867. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liang H, Studach L, Hullinger RL, Xie J
and Andrisani OM: Down-regulation of RE-1 silencing transcription
factor (REST) in advanced prostate cancer by hypoxia-induced
miR-106b~25. Exp Cell Res. 320:188–199. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Caruso P, MacLean MR, Khanin R, McClure J,
Soon E, Southgate M, MacDonald RA, Greig JA, Robertson KE, Masson
R, et al: Dynamic changes in lung microRNA profiles during the
development of pulmonary hypertension due to chronic hypoxia and
monocrotaline. Arterioscler Thromb Vasc Biol. 30:716–723. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shi XF, Wang H, Xiao FJ, Yin Y, Xu QQ, Ge
RL and Wang LS: MiRNA-486 regulates angiogenic activity and
survival of mesenchymal stem cells under hypoxia through modulating
Akt signal. Biochem Biophys Res Commun. 470:670–677. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang B, Zhou M, Li C, Zhou J, Li H, Zhu
D, Wang Z, Chen A and Zhao Q: MicroRNA-92a inhibition attenuates
hypoxia/reoxygenation-induced myocardiocyte apoptosis by targeting
Smad7. PLoS One. 9:e1002982014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lovering RC, Camon EB, Blake JA and Diehl
AD: Access to immunology through the Gene Ontology. Immunology.
125:154–160. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Y, Yang L, Wu B, Song Z and He S:
Transcriptome analysis of the plateau fish (Triplophysa dalaica):
Implications for adaptation to hypoxia in fishes. Gene.
565:211–220. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang Y, Wang L, Han J, Tang X, Ma M, Wang
K, Zhang X, Ren Q, Chen Q and Qiu Q: Comparative transcriptomic
analysis revealed adaptation mechanism of Phrynocephalus
erythrurus, the highest altitude Lizard living in the Qinghai-Tibet
Plateau. BMC Evol Bio. 15:1012015. View Article : Google Scholar
|
|
23
|
Butler PJ and Jones DR: Physiology of
diving of birds and mammals. Physiol Rev. 77:837–899. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Denko NC: Hypoxia, HIF1 and glucose
metabolism in the solid tumour. Nat Rev Cancer. 8:705–713. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Semenza GL: Hypoxia-inducible factors in
physiology and medicine. Cell. 148:399–408. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bakker WJ, Harris IS and Mak TW: FOXO3a is
activated in response to hypoxic stress and inhibits HIF1-induced
apoptosis via regulation of CITED2. Mol Cell. 28:941–953. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang B, Zhang YB, Zhang F, Lin H, Wang X,
Wan N, Ye Z, Weng H, Zhang L, Li X, et al: On the origin of
Tibetans and their genetic basis in adapting high-altitude
environments. PLoS One. 6:e170022011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu J, Liu XW and Kim HR: Platelet-derived
growth factor (PDGF) receptor-alpha-activated c-Jun NH2-terminal
kinase-1 is critical for PDGF-induced p21WAF1/CIP1 promoter
activity independent of p53. J Biol Chem. 278:49582–49588. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mizuno S, Bogaard HJ, Voelkel NF, Umeda Y,
Kadowaki M, Ameshima S, Miyamori I and Ishizaki T: Hypoxia
regulates human lung fibroblast proliferation via p53-dependent and
-independent pathways. Respir Res. 10:172009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mizuno S, Bogaard HJ, Kraskauskas D,
Alhussaini A, Gomez-Arroyo J, Voelkel NF and Ishizaki T: p53 Gene
deficiency promotes hypoxia-induced pulmonary hypertension and
vascular remodeling in mice. Am J Physiol Lung Cell Mol Physiol.
300:L753–L761. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gao Y, Dhanakoti S, Trevino EM, Sander FC,
Portugal AM and Raj JU: Effect of oxygen on cyclic GMP-dependent
protein kinase-mediated relaxation in ovine fetal pulmonary
arteries and veins. Am J Physiol Lung Cell Mol Physiol.
285:L611–L618. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gao Y, Portugal AD, Negash S, Zhou W,
Longo LD and Usha Raj J: Role of Rho kinases in PKG-mediated
relaxation of pulmonary arteries of fetal lambs exposed to chronic
high altitude hypoxia. Am J Physiol Lung Cell Mol Physiol.
292:L678–L684. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jernigan NL and Resta TC: Chronic hypoxia
attenuates cGMP-dependent pulmonary vasodilation. Am J Physiol Lung
Cell Mol Physiol. 282:L1366–L1375. 2002. View Article : Google Scholar : PubMed/NCBI
|