Introduction
Glioma is one of the most common primary central
nervous system tumors (1).
Characterized by strong proliferation and intensive invasion,
glioma cells always infiltrate into the surrounding normal brain
tissues, which makes glioma difficult to eradicate completely by
traditional surgical resection (2). Thus, comprehensive therapy, including
radiotherapy and chemotherapy, is used in order to delay the
recurrence and prolong survival rates (3,4).
However, the prognosis of glioma remains poor, mainly due to its
complex pathogenesis (5). To date,
no effective treatment for glioma has been identified, and the
search for new treatment options is thus necessary. Illuminating
the possible molecular mechanism is also an important aspect for
the treatment of glioma.
Shikonin, an active naphthoquinone extracted from
the traditional Chinese herb Zicao, was originally used in
traditional medicine as an ointment to treat wound healing, due to
its anti-inflammatory properties (6,7).
Recently, an increasing number of studies have indicated that
shikonin may exert anti-tumor functions in cancer cells, including
glioma (8–10). Marked inhibition of proliferation
and the induction of apoptosis have been identified in
shikonin-treated glioma cells, suggesting that shikonin may serve
as a potential therapeutic agent for glioma (11,12).
However, little is known about the molecular mechanism of shikonin
in glioma.
Cluster of differentiation (CD), a transmembrane
glycoprotein of the immunoglobulin superfamily, is upregulated in
diverse human tumor cells and has been validated as a therapeutic
target (13). Previous studies
have demonstrated that CD147 is rarely expressed on normal brain
cells but highly expressed on glioma cells, and that the expression
of CD147 is significantly associated with World Health Organization
tumor grade, indicating a potential role of CD147 in the occurrence
and progress of glioma (14). An
in vitro study further demonstrated that silencing of CD147
inhibits proliferation and induces apoptosis in glioma cells
(15). However, whether CD147 is
involved in shikonin-induced glioma cell apoptosis remains to be
elucidated.
The present study hypothesized that CD147 may be an
optional target of shikonin-induced cell apoptosis in glioma cells.
It investigated the influence of shikonin on the proliferation and
apoptosis of glioma cells and examined the potential molecular
mechanisms. The results may be of benefit in developing improved
therapies for glioma.
Materials and methods
Cell culture
Human U251 and U87MG (ATCC® HTB14™,
glioblastoma of unknown origin) cell lines were purchased from the
American Type Culture Collection (Manassas, VA, USA) and cultured
in Dulbecco's Modified Eagle's medium (DMEM; high glucose) medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with 1% penicillin/streptomycin, 2% L-glutamine and
10% fetal calf serum (FCS; HyClone; GE Healthcare Life Sciences,
Logan, UT, USA) at 37°C in an atmosphere humidified with 5%
CO2. Cells in the logarithmic growth phase were
collected for experimentation.
Monitoring cell proliferation using
the xCELLigence system
U251 and U87MG cells were harvested, washed and
resuspended in the DMEM with 10% FCS (HyClone; GE Healthcare Life
Sciences). The impedance values of each well were automatically
monitored using a real-time cell analyzer (RTCA; Roche Applied
Science, Penzberg, Germany) by the xCELLigence system (ACEA
Biosciences, San Diego, CA, USA) and expressed as a cell index (CI)
value. The baseline impedances was recorded using control wells
without cells containing 50 µl DMEM only. The cells were counted to
3×104 cells/ml and 100 µl were seeded into each well of
the E-Plate. Shikonin (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) was dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich;
Merck KGaA), diluted to the required concentrations (0.1, 0.5, 1,
2, 3 and 4 µM) and added into the corresponding wells. The E-plate
was subsequently placed into the xCELLigence system. Scans were run
with sweeps every min for the first 6 h. Subsequent sweeps were
taken every 30 min for 72 h.
Cell Counting Kit-8 (CCK-8) assay
U251 cells were plated on a 96-well plate at a
concentration of 1×105 cells/ml and cultured with
different concentrations of shikonin (0.1, 0.5, 1, 2, 3 and 4 µM)
for 24 h at 37°C. Subsequently, CCK-8 solution (10 µl/well;
Beyotime Institute of Biotechnology, Haimen, China) was added and
the plate was incubated at 37°C for 1 h. The cells were counted by
absorbance measurements at a wavelength of 450 nm.
Cell apoptosis assay
U251 cells were plated at a seeding density of
1×105 cells in a 24-well plate and treated with
different concentrations of shikonin (0.1, 0.5, 1, 2, 3 and 4 µM)
for 24 h at 37°C. The cells were collected and washed twice in cold
PBS. The cells were mixed in 100 µl 1X binding buffer and incubated
with 5 µl Annexin V (BD Pharmingen; BD Biosciences, San Jose, CA,
USA) at room temperature for 15 min in the dark. Subsequently, 5 µl
propidium iodide (PI; BD Pharmingen; BD Biosciences) was added
prior to detection by flow cytometry. The percentages of apoptotic
cells were calculated using FlowJo 7.6.1 (FlowJo LLC, Ashland, OR,
USA).
Knockdown and overexpression of CD147
by RNA interference
To knock down the expression of CD147 in U251 and
U87MG cells, the trans-lentiviral pLKO System (Shanghai GeneChem
Co., Ltd. Shanghai, China) was used to package the lentivirus and
acquire the CD147 knockdown cells (U251-KD and U87MG-KD). The
negative control cells were designated U251-NC and U87MG-NC.
Polybrene (1 µg/ml; Shanghai GeneChem Co., Ltd.) was used to
increase the transfection efficiency. The experiments were
performed as previously described (16), with a multiplicity of infection
(MOI) for U251 and U87MG cells of 10 and 20, respectively. To
upregulate the expression of CD147 in cells, the overexpression
lentivirus was produced by Shanghai GeneChem Co., Ltd. and
transfected into the U251 cells (MOI =5) and U87MG cells (MOI =10)
to acquire the CD147 overexpression cells (U251-OE and U87MG-OE).
According to the manufacturer's protocols, the cells infected with
the lentivirus were selected for puromycin resistance in DMEM
containing 2 µg/ml puromycin (Sigma-Aldrich; Merck KGaA) for 7–10
days. Following selection, the cells were maintained in medium
containing 1 µg/ml puromycin for ≥2 weeks until the final stable
cell clones were harvested and verified by determining the
expression of CD147 via flow cytometry.
Flow cytometric analysis
The expression of CD147 in U251 and U87MG cells was
assessed by flow cytometry. Following transfection for 3 weeks,
cells were fixed with 4% paraformaldehyde for 20 min at room
temperature at a concentration of 1×106 cells/ml. The
cells were blocked with 1% bovine serum albumin (BSA; Beyotime
Institute of Biotechnology) for 1 h at room temperature and
incubated with CD147-FITC (BD Pharmingen; BD Biosciences) for 30
min at room temperature. Following washing with PBS, stained cells
were analyzed on a Fluorescence Activated Cell Sorter flow
cytometer (BD Accuri C6 with CFlow Version 1.0.264.15 software, BD
Biosciences).
Western blot analysis
Cells (1×106) were collected, washed
twice with ice-cold PBS and lysed in 100 µl RIPA buffer (Beyotime
Institute of Biotechnology) supplemented with protease inhibitor
(Sigma-Aldrich; Merck KGaA) for 30 min. A Bicinchoninic Acid
Protein Assay kit (Pierce; Thermo Fisher Scientific, Inc.) was used
to determine the total protein concentration. Protein samples (30
ug/lane) were separated by 10% SDS-PAGE, and then transferred onto
polyvinylidene fluoride membranes (Sigma-Aldrich; Merck KGaA).
Following blocking with 5% milk for 1 h at room temperature, the
membranes were incubated overnight at 4°C with mouse anti-human
CD147 primary antibody (1:1,000; ab666, Abcam, Cambridge, MA, USA).
β-tubulin was used as the loading control (1:2,000; ab44928,
Abcam). The blots were extensively washed with PBS with 5% Tween-20
and incubated with goat anti-mouse horseradish
peroxidase-conjugated secondary antibody (1:3,000; ab205719, Abcam)
for 1 h at room temperature. Following washing, the blots were
visualized with BeyoECL Plus reagent (Beyotime Institute of
Biotechnology) using a ChemiDoc™ detection system (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Protein expression was
quantified using ImageJ v1.50i (National Institutes of Health,
Bethesda, MD, USA).
Measuring intracellular reactive
oxygen species (ROS)
The redox-sensitive dye dichloro-dihydro-fluorescein
diacetate (DCFH-DA; Beyotime Institute of Biotechnology) was used
to evaluate the levels of intracellular ROS. Briefly,
1×106 cells were incubated with 1% Triton X-100
following staining with 2 µmol DCFH-DA in the dark at room
temperature for 20 min. Subsequently, the cells were washed with
PBS and resuspended in RPMI-1640 culture medium (Gibco; Thermo
Fisher Scientific, Inc.) containing different concentrations of
shikonin (0.5, 1 and 2 µM) at 37°C for 2 h. The fluorescence was
measured at an excitation wavelength of 488 nm and an emission
wavelength of 530 nm using a fluorescence spectrometer.
Measuring mitochondrial membrane
potential (MMP)
The fluorochrome dye Rh123 (Beyotime Institute of
Biotechnology) was used to evaluate the alterations in MMP.
Briefly, U251 cells were incubated with different concentrations of
shikonin (0.5, 1 and 2 µM) at 37°C for 24 h. Following incubation,
cells were collected and stained with 5 µg/ml Rh123 at 37°C for 10
min. Cells were harvested and washed twice, and resuspended in 500
µl RPMI-1640 culture medium. The fluorescence intensity of cells
was quantitatively analyzed by flow cytometry (BD Accuri C6 with
CFlow Version 1.0.264.15 software).
Statistical analysis
The results are presented as the mean ± standard
deviation from ≥3 independent experiments. Comparisons between two
groups with respect to ROS generation and MMP (U251NC vs. U251NC+S,
U251KD vs. U251KD+S and U251OE vs. U251OE+S) was performed using a
Student's t-test, and comparisons between multiple groups were
performed using one-way analysis of variance followed by a
Student-Newman-Keuls test. Data analyses were performed using Graph
Pad Prism version 5.0 (GraphPad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Shikonin inhibits U251 cell
proliferation in a dose- and time-dependent manner
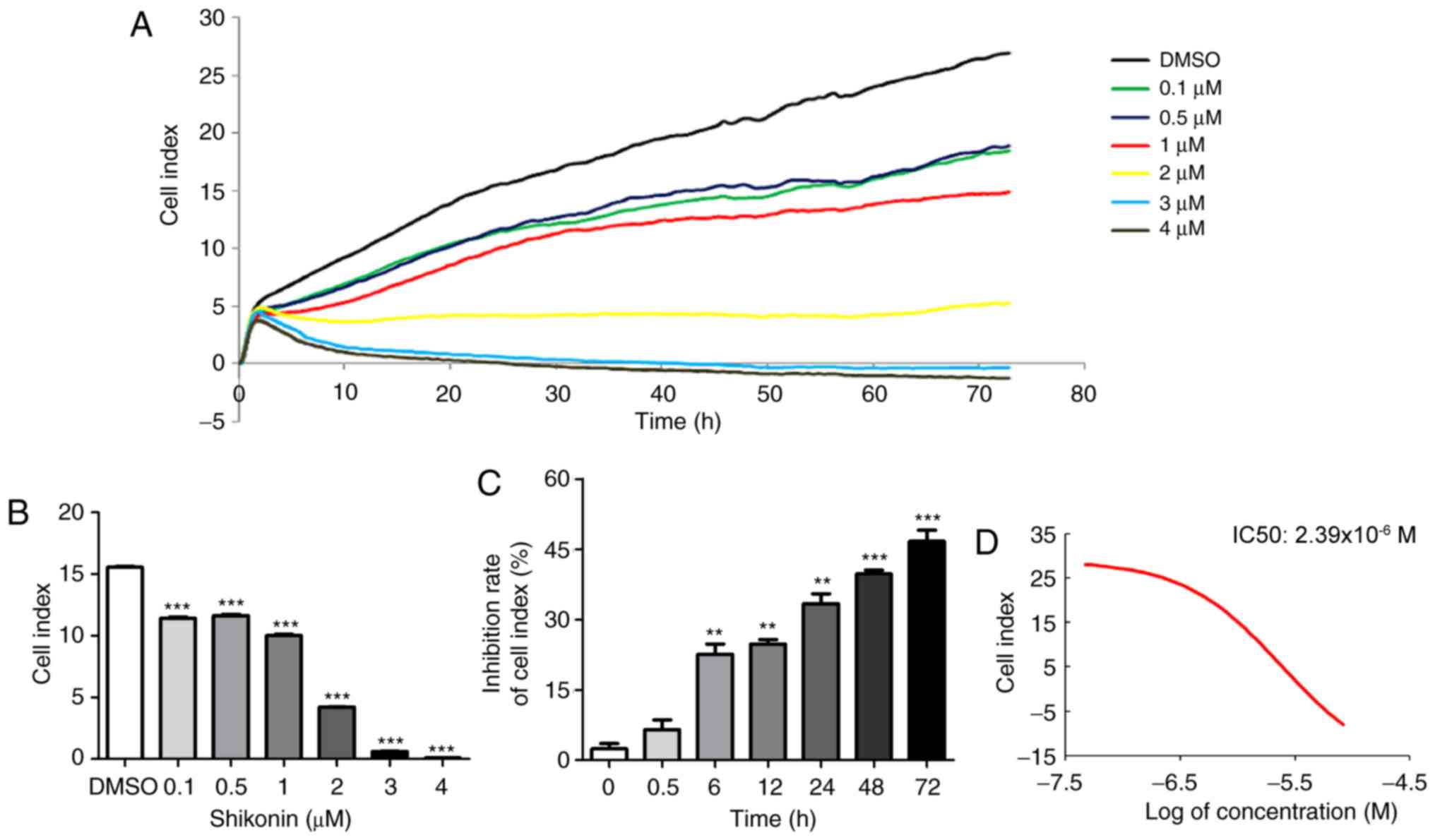
An RTCA was used to assess the inhibitory effect of
shikonin on U251 cell proliferation. The dynamic pattern of cell
index (CI) curves for different shikonin concentrations on U251
cells are presented in Fig. 1A.
The CI was further compared between the different concentration
groups and the DMSO control group at the time point of 24 h, and it
was identified that CI was decreased significantly in a
dose-dependent manner (Fig. 1B).
It was also investigated whether the effect of shikonin on cell
viability was time-dependent. At a concentration of 1 µM, shikonin
exhibited a significant inhibitory effect from 6 h, and the
inhibitory rate gradually increased with time, indicating a
time-dependent effect of shikonin in U251 cells (Fig. 1C). The half-maximal inhibitory
concentration (IC50) value of shikonin for U251 cells
following 24 h of treatment was 2.39×10−6 M (Fig. 1D).
Shikonin induces apoptosis in human
glioma U251 cells
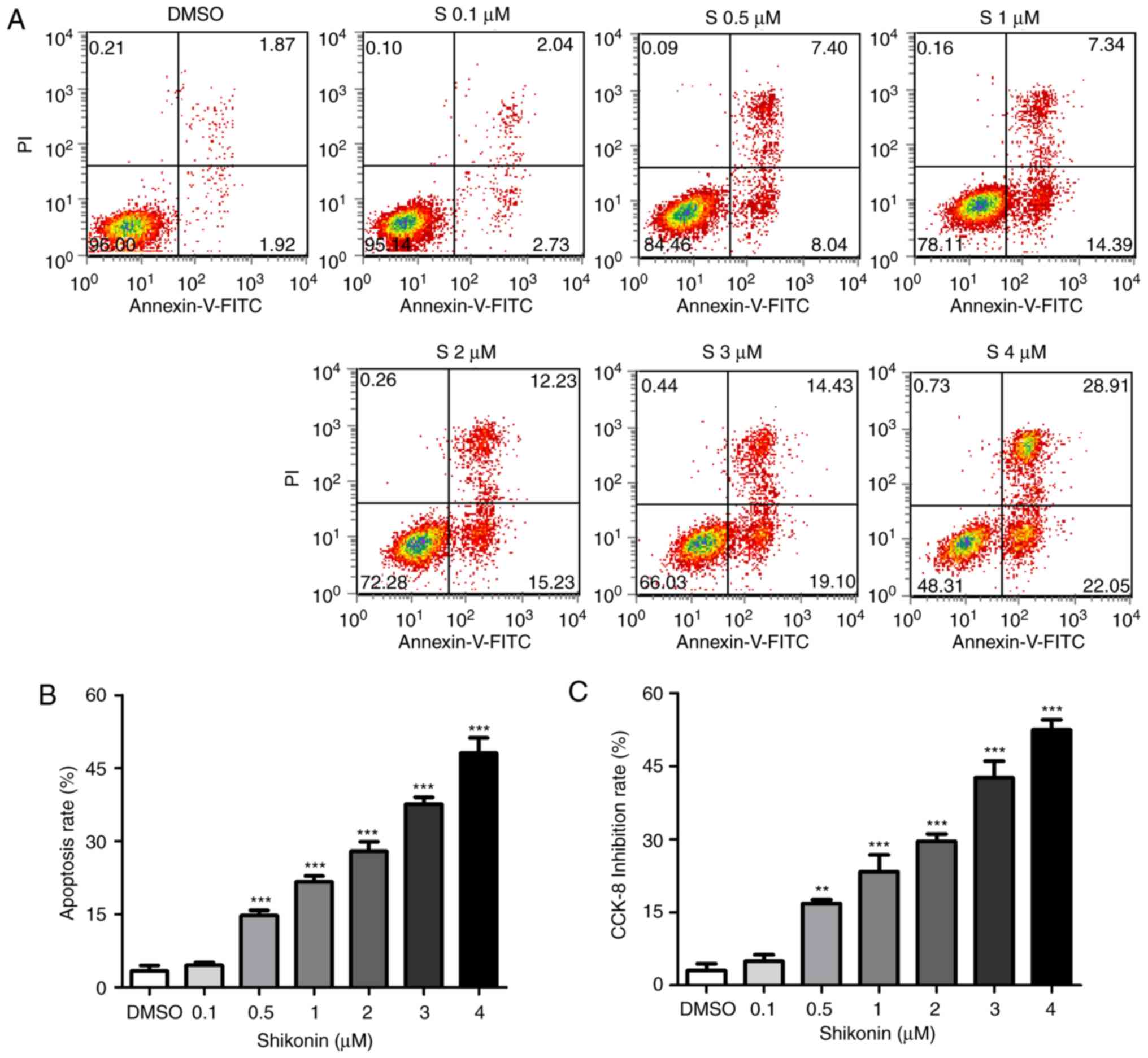
To further investigate whether shikonin inhibited
U251 cell growth through the induction of apoptosis, the percentage
of apoptotic cells was calculated by flow cytometry using the
Annexin V/PI double staining assay following treatment of the cells
with various doses of shikonin. The representative flow cytometry
data are presented in Fig. 2A. It
was demonstrated that treatment with 0.5–4 µM shikonin for 24 h
significantly increased the numbers of apoptotic cells compared
with the control group (Fig. 2B),
and the numbers of apoptotic cells increased in a dose-dependent
manner. Shikonin-induced cell death was also tested by CCK-8. As
presented in Fig. 2C, shikonin
exhibited significant cell growth inhibition at each concentration,
0.5–4 µM, similar to the apoptosis data. These results indicated
that the apoptotic pathway serves a pivotal role in the
anti-proliferative effect of shikonin on U251 cells.
CD147 expression is downregulated in
glioma cells under shikonin treatment
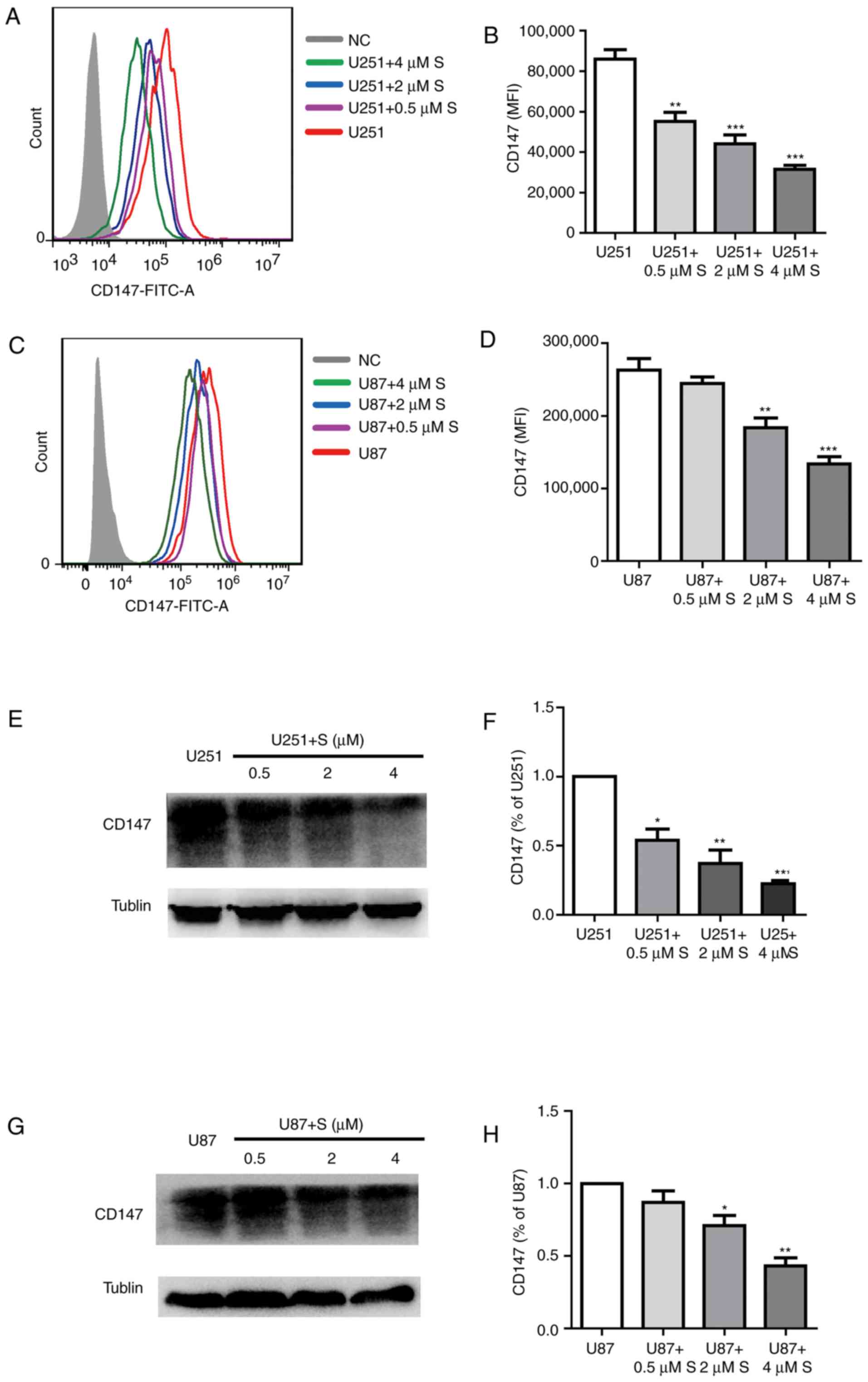
The expression of CD147 identified to be involved in
tumor cell proliferation was examined to study the molecular
mechanism underlying the inhibitory effects of shikonin on U251 and
U87MG cell viability. As presented in Fig. 3A-D, fluorescence-activated cell
sorting analysis for CD147 expression demonstrated a gradual
decrease in U251 and U87MG cells treated with shikonin compared
with the untreated cells. Western blot analysis confirmed that with
the increase in shikonin concentration, the expression of CD147 was
decreased (Fig. 3E-H).
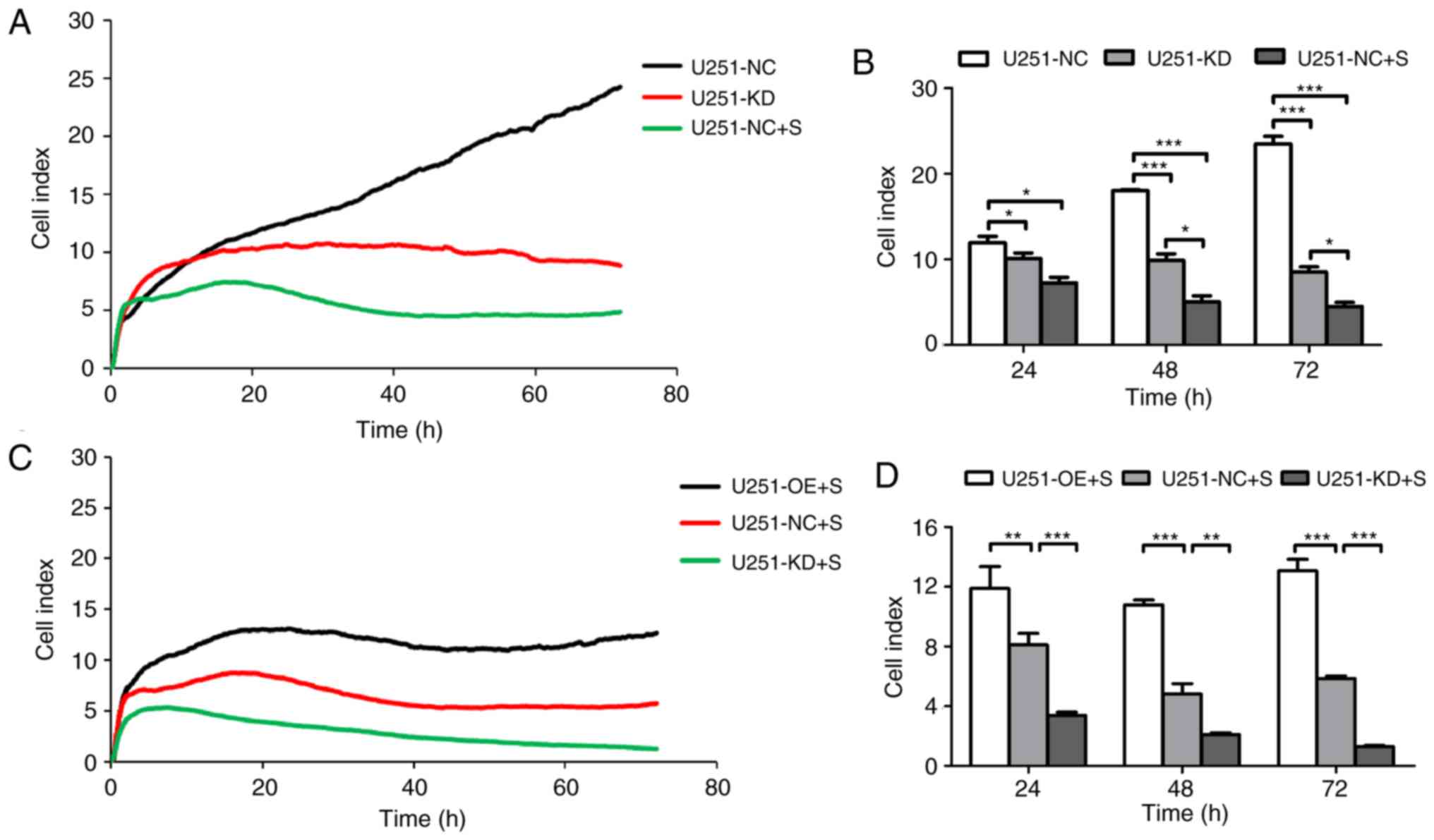
Knocking-down of CD147 inhibits glioma
cell growth, whereas CD147 overexpression enhances cell growth in
glioma cells
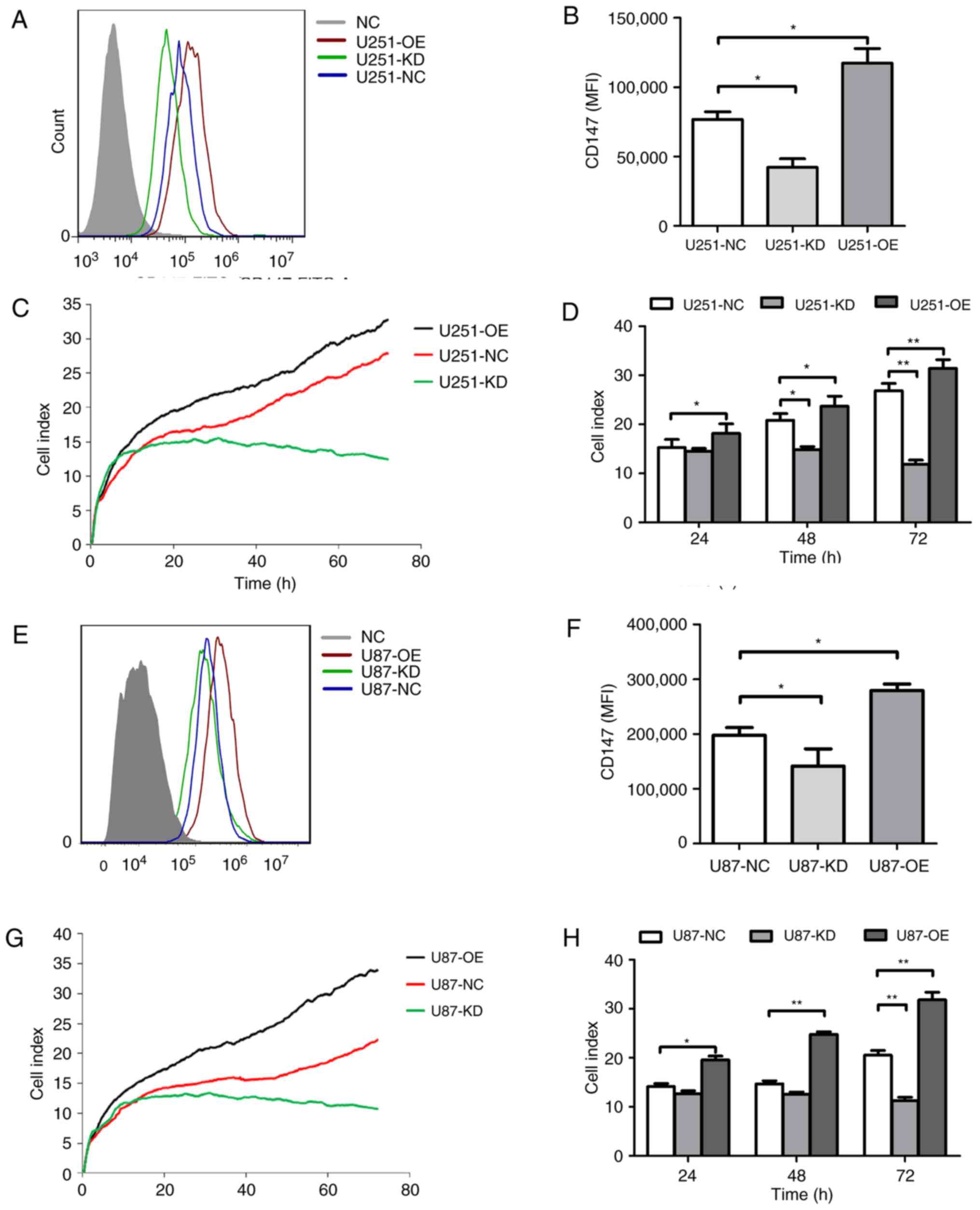
To examine whether altered CD147 expression affects
glioma growth, CD147 was knocked down and overexpressed in U251 and
U87MG cells, respectively, using a lentivirus system. As indicated
in Fig. 4A and B, following
transfection with CD147 KD and CD147 OE viruses, CD147 was
identified to be significantly knocked down in U251-KD cells and
overexpressed in U251-OE cells. The cell viability of each cell
line was evaluated by means of RTCA for 72 h. U251-KD cells had
lower CI values when compared with the CI index values of U251-NC
cells, which indicated a reduced cell viability (Fig. 4C and D). However, increased cell
viability was observed in U251-OE cells, which exhibited relatively
high CI values compared with those of U251-NC cells (Fig. 4C and D). Similar results were
obtained in U87MG-KD and U87MG-OE cells (Fig. 4E-H). All the data suggested a
positive role for CD147 in glioma cell proliferation.
Overexpression of CD147 resists
shikonin-induced U251 cell death
It was further investigated whether CD147 was
involved in shikonin-induced cell death in glioma cells. As
presented in Fig. 5A and B,
downregulation of CD147 exerted a significant inhibitory effect on
U251 cell proliferation, similar to the cells treated with
shikonin. Comparatively strong cell growth inhibition was observed
in shikonin-induced U251 cells with the passage of time. When
treated with shikonin, U251-NC cells demonstrated decreased CI
values compared with those of CD147-overexpressing U251-OE cells
(Fig. 5C and D).
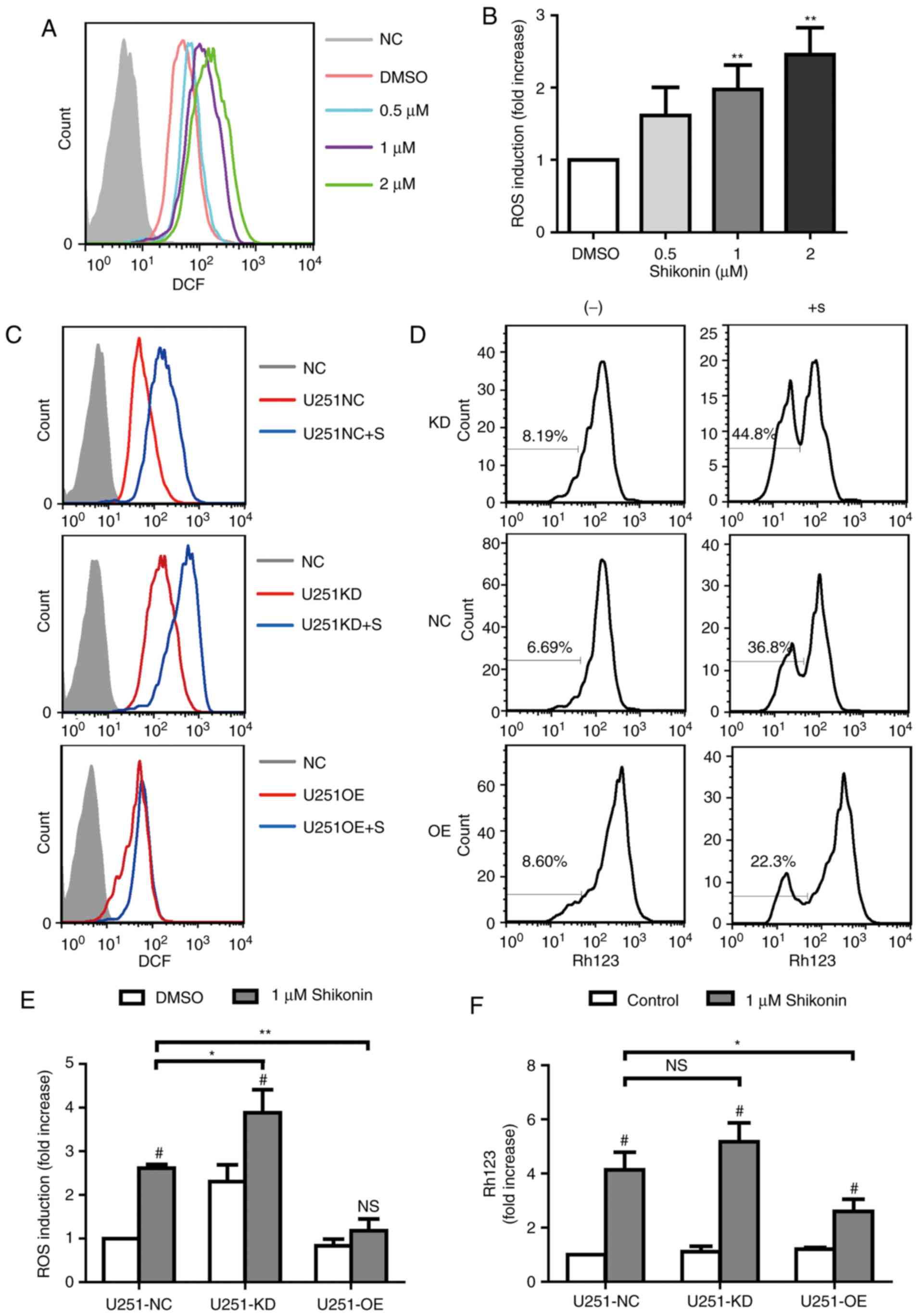
Shikonin induces apoptosis in U251
glioma cells through ROS generation and mitochondrial dysfunction
via CD147 expression
Shikonin has been reported to dysregulate cellular
ROS levels and MMP, which ultimately induces cell apoptosis
(17). Therefore, the
intracellular ROS generation and MMP levels were detected in U251
glioma cells. Treatment with shikonin markedly increased
intracellular ROS generation in a dose-dependent manner (Fig. 6A and B), as measured by
fluorescence staining. Furthermore, it was identified that
downregulation of CD147 in U251 glioma cells exhibited increased
intracellular ROS and MMP levels, which further increased following
the treatment with shikonin (Fig. 6C
and D). However overexpression of CD147 was indicated to defend
against the effect of shikonin on the intracellular ROS and MMP
levels in U251 glioma cells (Fig. 6C
and D). The statistical data are presented in Fig. 6E and F. All the data suggested that
shikonin caused mitochondrial dysfunction, partly via regulation of
CD147 expression.
Discussion
The present study identified the reduced cell
proliferation and decreased expression level of CD147 caused by
shikonin in U251 cells. Shikonin was able to induce apoptosis in
human glioma U251 cells in a dose-dependent manner, and
overexpression of CD147 resisted shikonin-induced U251 apoptosis.
Additionally, an increased expression level of CD147 suppressed the
elevated production of ROS and MMP levels induced by shikonin. The
results of the present study indicated that shikonin-induced
apoptosis in glioma cells was associated with the downregulation of
CD147 and upregulation of oxidative stress.
As an anthraquinone compound derived from the roots
of Lithospermum erythrorhizon, shikonin exhibits a broad
spectrum of anti-tumor effects by inducing apoptosis in cancer of
the gallbladder (18), pancreas
(19), colon (20) and breast (21). The present study also demonstrated
that shikonin inhibited glioma U251 cell proliferation in a dose-
and time-dependent manner. At a concentration of 1 µM, shikonin
exhibited a significant inhibitory effect from 6 h, and the
IC50 value of shikonin for U251 cells following 24 h
treatment was 2.39×10−6 M. Furthermore, treatment with
0.5–4 µM shikonin for 24 h significantly increased the number of
apoptotic cells, and this result was similar to that for cell
growth inhibition. These findings indicated that the apoptotic
pathway serves a pivotal role in the anti-proliferative effect of
shikonin on glioma cells. Lu et al (22) reported that shikonin induced glioma
cell necroptosis in vitro, as PI positive cells following
shikonin (20 µM) treatment did not exhibit apoptotic features, such
as nuclear condensation or fragmentation. This may be due to the
different concentrations of shikonin used in the studies. The
present study identified that a high concentration of shikonin
demonstrated characteristics of necrosis, whereas a low
concentration demonstrated characteristics of apoptosis (data not
shown). In spite of the large number of studies indicating that
shikonin induces apoptosis in various human tumor cell lines
(8,9,11),
the molecular mechanism of the potential anti-glioma effect of
shikonin is not completely understood.
Apoptosis, a type of programmed cell death, is
regulated by multiple genes and protein factors (23); ROS generation has been demonstrated
to serve a critical role in this process (24). As dysfunctional mitochondria are
thought to be one of the principal sources of intracellular ROS
(25), the present study detected
the cellular ROS levels and MMP when treated with shikonin, and
identified the disrupted MMP and increased intracellular ROS
generation in glioma U251 cells. As in hepatoma cells, shikonin
generates large amounts of intracellular ROS during the early stage
of the apoptotic process (26).
These findings demonstrated that shikonin may target mitochondria
and cause mitochondrial dysfunction, further increasing ROS
generation and ultimately inducing apoptosis.
A number of studies have demonstrated that CD147
serves an important role in glioma (15,27).
The results of the present study also proved that CD147 was able to
affect the development of glioma cells; knockdown of CD147
inhibited glioma cell growth, whereas CD147 overexpression enhanced
cell growth in glioma cells. Yin et al (14) demonstrated that downregulation of
CD147 induces apoptosis in glioma. The present study hypothesized
that CD147 may be an optional target of shikonin-induced cell
apoptosis on glioma cells. CD147 expression was first detected when
glioma cells were treated with shikonin, and it was identified that
CD147 expression was downregulated in glioma cells under shikonin
treatment. Furthermore, overexpression of CD147 in glioma cells
resisted shikonin-induced cell death and reduced the effect of
shikonin on the intracellular ROS and MMP levels. In summary, the
present study suggested that shikonin induced apoptosis in glioma
via the regulation of CD147 expression.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant nos. 81601376 and 81471415),
University's Key Disciplines of Molecular Immunology, the Natural
Science Basic Research Plan in Shaanxi Province of China (grant no.
2017JM8086), Scientific Research Program Funded by Shaanxi
Provincial Education Department (grants no. 17JK0666 and 17JK0648),
Xi'an Medical College Fund (grant nos. 2017GJFY32, 2017PT30,
2017PT35 and 2017PT42) and Shaanxi Science and Technology Project
for funding of people studying abroad.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
NG and RM were responsible for the analysis and
interpretation of data of the manuscript; NG, XCGa, DH and ZFH
performed the majority of the experiments; XCGo and FLJ were
responsible for design of the study and drafting of the manuscript.
YN and NCJ contributed to the acquisition, analysis, and
interpretation of data for the study and revised the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Louis DN, Perry A, Reifenberger G, von
Deimling A, Figarella-Branger D, Cavenee WK, Ohgaki H, Wiestler OD,
Kleihues P and Ellison DW: The 2016 World Health Organization
Classification of tumors of the central nervous system: A summary.
Acta Neuropathol. 131:803–820. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Aldape K, Zadeh G, Mansouri S,
Reifenberger G and von Deimling A: Glioblastoma: Pathology,
molecular mechanisms and markers. Acta Neuropathol. 129:829–848.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hottinger AF, Aissa AB, Espeli V, Squiban
D, Dunkel N, Vargaset MI, Hundsberger T, Mach N, Schaller K, Weber
DC, et al: Phase I study of sorafenib combined with radiation
therapy and temozolomide as first-line treatment of high-grade
glioma. Br J Cancer. 110:2655–2661. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bush NA, Chang SM and Berger MS: Current
and future strategies for treatment of glioma. Neurosurg Rev.
40:1–14. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reifenberger G, Wirsching HG,
Knobbe-Thomsen CB and Weller M: Advances in the molecular genetics
of gliomas-implications for classification and therapy. Nat Rev
Clin Oncol. 14:434–452. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Andujar I, Ríos JL, Giner RM and Recio MC:
Pharmacological properties of shikonin-a review of literature since
2002. Planta Med. 79:1685–1697. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang X, Cui JH, Meng QQ, Li SS, Zhou W
and Xiao S: Advance in anti-tumor mechanisms of shikonin, alkannin
and their derivatives. Mini Rev Med Chem. 18:164–172. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo ZL, Li JZ, Ma YY, Qian D, Zhong JY,
Jin MM, Huang P, Che LY, Pan B, Wang Y, et al: Shikonin sensitizes
A549 cells to TRAIL-induced apoptosis through the JNK, STAT3 and
AKT pathways. BMC Cell Biol. 19:292018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Durchschein C, Hufner A, Rinner B,
Stallinger A, Deutsch A, Lohberger B, Bauer R and Kretschmer N:
Synthesis of novel shikonin derivatives and pharmacological effects
of cyclopropylacetyl shikonin on melanoma cells. Molecules.
23:E28202018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang FY, Hu Y, Que ZY, Wang P, Liu YH,
Wang ZH and Xue YX: Shikonin inhibits the migration and invasion of
human glioblastoma cells by targeting phosphorylated β-Catenin and
phosphorylated PI3K/Akt: A potential mechanism for the anti-glioma
efficacy of a traditional Chinese herbal medicine. Int J Mol Sci.
16:23823–23848. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang FL, Wang P, Liu YH, Liu LB, Liu XB,
Li Z and Xue YX: Topoisomerase I inhibitors, shikonin and
topotecan, inhibit growth and induce apoptosis of glioma cells and
glioma stem cells. PLoS One. 8:e818152013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhou Z, Lu B, Wang C, Wang Z, Piao T, Luo
M, Meng F, Chi G, Luo Y and Ge P: RIP1 and RIP3 contribute to
shikonin-induced DNA double-strand breaks in glioma cells via
increase of intracellular reactive oxygen species. Cancer Lett.
390:77–90. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiong L, Edwards CR III and Zhou L: The
biological function and clinical utilization of CD147 in human
diseases: A review of the current scientific literature. Int J Mol
Sci. 15:17411–17441. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yin H, Shao Y and Chen X: The effects of
CD147 on the cell proliferation, apoptosis, invasion, and
angiogenesis in glioma. Neurol Sci. 38:129–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li H, Xi Z, Dai X, Wu W, Li Y, Liu Y and
Zhang H: CD147 and glioma: A meta-analysis. J Neurooncol.
134:145–156. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo N, Zhang K, Lv MH, Miao JL, Chen ZN
and Zhu P: CD147 and CD98 complex-mediated homotypic aggregation
attenuates the CypA-induced chemotactic effect on Jurkat T cells.
Mol Immunol. 63:253–263. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wiench B, Eichhorn T, Paulsen M and
Efferth T: Shikonin directly targets mitochondria and causes
mitochondrial dysfunction in cancer cells. Evid Based Complement
Alternat Med. 2012:7260252012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhai T, Hei Z, Ma Q, Liang H, Xu Y, Zhang
Y, Jin L, Han C and Wang J: Shikonin induces apoptosis and G0/G1
phase arrest of gallbladder cancer cells via the JNK signaling
pathway. Oncol Rep. 38:3473–3480. 2017.PubMed/NCBI
|
|
19
|
Chen C, Xiao W, Huang L, Yu G, Ni J, Yang
L, Wan R and Hu G: Shikonin induces apoptosis and necroptosis in
pancreatic cancer via regulating the expression of RIP1/RIP3 and
synergizes the activity of gemcitabine. Am J Transl Res.
9:5507–5517. 2017.PubMed/NCBI
|
|
20
|
Liang W, Cui J, Zhang K, Xi H, Cai A, Li
J, Gao Y, Hu C, Liu Y, Lu Y, et al: Shikonin induces ROS-based
mitochondria-mediated apoptosis in colon cancer. Oncotarget.
8:109094–109106. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Shahsavari Z, Karami-Tehrani F and Salami
S: Targeting cell necroptosis and apoptosis induced by Shikonin via
receptor interacting protein kinases in estrogen receptor positive
breast cancer cell line, MCF-7. Anticancer Agents Med Chem.
18:245–254. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lu B, Gong X, Wang ZQ, Ding Y, Wang C, Luo
TF, Piao MH, Meng FK, Chi GF, Luo YN and Ge PF: Shikonin induces
glioma cell necroptosis in vitro by ROS overproduction and
promoting RIP1/RIP3 necrosome formation. Acta Pharmacol Sin.
38:1543–1553. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ondroušková E and Vojtěšek B: Programmed
cell death in cancer cells. Klin Onkol. 27 (Suppl 1):S7–S14.
2014.(In Czech). View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Redza-Dutordoir M and Averill-Bates DA:
Activation of apoptosis signalling pathways by reactive oxygen
species. Biochim Biophys Acta. 1863:2977–2992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Balaban RS, Nemoto S and Finkel T:
Mitochondria, oxidants, and aging. Cell. 120:483–495. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gong K and Li W: Shikonin, a Chinese
plant-derived naphthoquinone, induces apoptosis in hepatocellular
carcinoma cells through reactive oxygen species: A potential new
treatment for hepatocellular carcinoma. Free Radic Biol Med.
51:2259–2271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fei F, Li S, Fei Z and Chen ZN: The roles
of CD147 in the progression of gliomas. Expert Rev Anticancer Ther.
15:1351–1359. 2015. View Article : Google Scholar : PubMed/NCBI
|