Introduction
Osteoporosis (OP) can be diagnosed indirectly via
non-invasive measurements of bone mineral density (BMD) (1). In recent years, men have been shown
to be affected by this ‘silent epidemic’. The prevalence of OP was
found to be between 6 and 22% in men (2). The lifetime risk of osteoporotic
fractures in men is approximately one-third of that in women
(3). OP-related fractures are
associated with higher mortality rates in men than in women
(4). Thus, male OP should draw
more attention from clinicians worldwide.
The relationship between hypercholesterolemia and
osteoporosis in men has been preliminarily discovered (5). To the best of our knowledge, the
widespread and profound effects of hypercholesterolaemia on male
bone and their underlying mechanisms have seldom been reported thus
far. As the prevalence of hypercholesterolaemia in male adults is
increasing, and is even higher than that in females (6), the effects of hypercholesterolaemia
on male bone should attract sufficient attention. Therefore, we
aimed to investigate the effects of hypercholesterolaemia on male
bone and the underlying mechanisms.
Materials and methods
Human subjects and design
Between July 2015 and October 2015, 216 men (aged
≥18 years) were recruited by the Jinan Health Organization (Jinan,
China). All subjects completed a self-reported questionnaire
including items pertaining to their demographic characteristics,
lifestyle habits, dietary habits, medical histories and current
medications. Height and body weight were measured to calculate body
mass index (BMI) using the following formula: BMI=body weight
(kg)/height (m2). Fasting blood samples were drawn for
clinical laboratory tests.
The exclusion criteria included participants with
chronic diseases, hepatic, renal, thyroid, other endocrine
diseases, diabetes and hypertriglyceridemia and participants taking
medications such as glucocorticoids, testosterone, thyroid hormone,
bisphosphonates, calcitonin, calcium and active vitamin D
analogues. Overall, 216 men were enrolled in the present study,
which was approved by the Ethics Committee of Provincial Hospital
Affiliated with Shandong University (Jinan, China) and all
participants provided written informed consent before participating
in the study.
According to the criteria outlined in China's Adult
Dyslipidemia Prevention Guide (2016 version) (7), the 216 men enrolled in the present
study were divided into a hypercholesterolaemia group (total
cholesterol ≥5.20 mmol/l, n=102) and a control group (total
cholesterol <5.20 mmol/l, n=114).
Clinical laboratory tests and BMD
measurements
Serum total cholesterol (TC), triglyceride (TG),
low-density lipoprotein cholesterol (LDL-C), high-density
lipoprotein cholesterol (HDL-C) and fasting plasma glucose (FPG)
levels were measured by the Beckman Chemistry Analyzer AU5800
System 195 (Beckman Coulter, Inc., Tokyo, Japan) and beta collagen
cross-linking (beta-CTX), type I anterior collagen amino terminal
peptide (PINP), testosterone (To), free thyroxine (FT4), thyroid
stimulating hormone (TSH) and 25-hydroxyvitamin D [25(OH)D]
concentrations were determined by an electrochemiluminescence
immunoassay (Cobas 8000 analyser; Roche Diagnostics, Basel,
Switzerland) at the Clinical Laboratory of Shandong Provincial
Hospital. BMD was measured in the low-median region of the left
forearm by a trained technologist using a dual-energy X-ray
absorptiometry (DXA) fan beam bone densitometer (EXA-3000;
OsteoSys, Co., Ltd., Seoul, Korea).
Animal experimental design
The animal experimental protocol was approved by the
Animal Ethics Committee of Shandong Provincial Hospital. Two animal
models were used in this study. For the first model, 24 6-week-old
male Sprague-Dawley rats weighing approximately 200 g were obtained
from Vital River Laboratory Animal Technology Co, Ltd. (Beijing,
China). Following being acclimated to their housing environments
for 1 week, the rats received one of two diets, a normal control
diet (NCD, 100% standard rodent chow) or a high cholesterol diet
(HCD, 2% cholesterol + 98% standard rodent chow), and were randomly
assigned into three groups: An NCD group (36 weeks), an HCD group
(36 weeks), or an HCD+NCD group (24 weeks HCD+12 weeks NCD). Rats
of the second model included male wild-type (ApoE+/+)
rats and male ApoE-knockout (KO) (ApoE−/−) rats, which
were obtained by breeding heterozygous (ApoE+/−) rats
purchased from Beijing Biocytogen Co., Ltd. (Beijing, China) and
were fed an NCD for 20 weeks. All animals were maintained in
temperature-controlled rooms (22–25°C) under 12-h light/dark cycles
and allowed free access to food and water. All rats were sacrificed
by exsanguination following anesthesia with subcutaneous injection
of 30 mg/kg pentobarbital sodium at the end of the feeding period
after being fasted for 8 h. Body weights were obtained, and fasting
blood samples were withdrawn. The left femurs were collected and
cleaned of soft tissue before being packed in gauze soaked with a
NaCl solution (9 g/l) and stored at −30°C.
Serum parameter analysis
Serum lipid profiles were measured by an Olympus
AU5400 automatic biochemical analyser (Olympus Corp., Mishama,
Japan). Serum tartrate-resistant acid phosphatase (TRAP), beta-CTX,
PINP, bone alkaline phosphatase (ALPL) and bone glaprotein (BGP)
were measured with ELISA kits (Uscn Life Science, Inc., Wuhan,
China).
Micro-CT (mCT) measurements
The trabecular and cortical microarchitectures of
the left femurs were analysed using mCT (SkyScan 1176;
Bruker-microCT, Kontich, Belgium). The slice thickness was 17.93
µm, and the voxel resolution was 22 µm3. Quantitative
analyses of the morphometric parameters were conducted using the
appropriate software package, and 3D images of the distal femur and
femoral diaphysis were reconstructed for visualization. All
analyses were conducted by a trained professional in a blinded
manner who was unaware of the treatments to which the specimens had
been subjected.
Biomechanical property
measurement
We assessed the mechanical properties of the femoral
diaphysis as previously reported (8) using an ElectroForce 3230 Bose System
(Bose, Minnetonka, MN, USA). Based on these parameters reported by
the instrument, we analysed the mechanical properties of the
femoral diaphysis.
Primary culture of osteoblasts
Primary osteoblasts obtained from new-born rats were
cultured as described in a previous study (9). The cells were cultured in 6-well
plates at a density of 1×105 cells/well in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) containing 10% (v/v) fetal bovine serum
(FBS; Biochrom GmbH, Berlin, Germany), 100 U/ml
penicillin-streptomycin solution (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) and 2 mM L-glutamine (Sigma-Aldrich; Merck
KGaA) at 37°C in a humidified 5% CO2 atmosphere. The
medium was refreshed every 48 h. Haematoxylin and eosin (H&E)
staining was used to identify the typical morphological
characteristics of the osteoblasts. Alkaline phosphatase (ALP)
(Genmed, Shanghai, China) staining, Alizarin red (Beijing Solarbio
Science & Technology Co. Ltd. (Beijing, China) staining and Von
Kossa (Genmed) staining were performed for osteoblast phenotypic
and functional identification. After 3 days of incubation at 37°C
in a humidified 5% CO2 atmosphere, the primary
osteoblasts were starved of serum for 2 h and then treated with
cholesterols (Sigma-Aldrich; Merck KGaA; 0, 10 and 20 µg/ml) for
12, 24 and 48 h. We subsequently analysed osteoblast function by
measuring gene expression.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted using TRIzol (Takara
Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer's instructions. cDNA was synthesized from 500 ng of
total RNA using ReverTra Ace reverse transcriptase (Takara
Biotechnology Co., Ltd.) and oligo dT primers (Takara Biotechnology
Co., Ltd.). The following PCR primers were used in the study: ALPL:
Forward (F), 5′-CATCGCCTATCAGCTAATGCACA-3′ and reverse (R),
5′-ATGAGGTCCAGGCCATCCAG-3′; collagen I: F,
5′-GACATGTTCAGCTTTGTGGACCTC-3′ and R,
5′-AGGGACCCTTAGGCCATTGTGTA-3′; BGP: F, 5′-GGACCCTCTCTCTGCTCACTCT-3′
and R, 5′-CTTACTGCCCTCTGCTTGG-3′; β-actin: F,
5′-ACCCAGATCATGTTTGAGAC-3′ and R, 5′-GTCAGGATCTTCATGAGGTAGT-3′.
Extracellular protein ELISA
Secreted BGP and ALPL levels were measured with
ELISA kits (Uscn Life Science, Inc.) according to the
manufacturers' protocols. The protein concentration of the cell
homogenate was determined using a BCA assay (Shen Neng Bo Cai
Corp., Shanghai, China). The levels of BGP and ALPL in the media
were normalized to the total cellular protein content.
Statistical analysis
Statistical analyses were performed using SPSS 22
(IBM Corp., Armonk, NY, USA). All data are expressed as the mean ±
standard deviation for continuous variables. Unpaired Student's
t-test or one-way analysis of variance (ANOVA) and LDS, S-N-K,
Duncan and Dunnet post hoc tests were used to compare BMD, beta-CT
X and PINP levels between hypercholesterolaemia and controls and
all animal and cell measurement data. Multivariate linear
regression analysis was used to evaluate the correlations between
clinical parameters and serum BMD and beta-CTX and PINP levels.
P-values <0.05 were considered to indicate a statistically
significant result.
Results
Correlation between serum cholesterol
and BMD, beta-CTX and PINP
Table I presents
the characteristics of both groups. BMD was significantly lower in
men with hypercholesterolaemia than in control subjects
(P<0.001), and both beta-CTX and PINP levels were increased in
the hypercholesterolaemia group compared with those in the controls
(P<0.05). Moreover, serum TC, LDL-C and HDL-C levels were
significantly higher among men with hypercholesterolaemia
(P<0.001). There were no significant differences in the age, BMI
or FPG, HbAc1, To, FT4, TSH, 25(OH)D and TG levels between the two
groups.
 | Table I.Clinical characteristics of the
subjects with hypercholesterolemia and control subjects. |
Table I.
Clinical characteristics of the
subjects with hypercholesterolemia and control subjects.
| Characteristics
(mean ± SD) | Controls
(n=112) |
Hypercholesterolemia (n=104) | Total (n=216) |
|---|
| Age (years) | 48.140±13.750 | 50.098±13.995 | 49.065±13.868 |
| BMI
(kg/m2) | 25.337±3.810 | 25.256±3.031 | 25.299±3.456 |
| FPG (mmol/l) | 5.421±0.619 | 5.462±0.469 | 5.462±0.562 |
| HbA1 (%) | 5.503±1.139 | 5.433±0.342 | 5.454±0.365 |
| To (ng/ml) | 5.652±1.624 | 5.669±2.266 | 5.497±1.972 |
| FT4 (pmol/l) | 19.256±2.143 | 18.913±2.704 | 19.094±2.425 |
| TSH (µIU/ml) | 2.338±1.691 | 2.465±1.555 | 2.398±1.626 |
| 25(OH)D
(ng/ml) | 28.448±9.183 | 28.257±10.048 | 28.358±9.579 |
| TG (mmol/l) | 1.201±0.328 | 1.27±0.290 | 1.234±0.312 |
| TC (mmol/l) | 4.485±0.496 |
5.737±0.412b | 5.076±0.776 |
| LDL-C (mmol/l | 2.789±0.406 |
3.722±0.351b | 3.230±0.602 |
| HLD-C (mmol/l | 1.175±0.235 |
1.309±0.262b | 1.238±0.257 |
| BMD
(g/cm2) | 0.536±0.066 |
0.505±0.092b | 0.522±0.081 |
| Beta-CTX
(ng/ml) | 0.437±0.164 |
0.5±0.201a | 0.467±0.185 |
| PINP (ng/ml) | 46.974±16.259 |
52.477±19.011a | 49.572±17.785 |
To further study the effects of cholesterol on BMD
and beta-CTX and PINP levels, we performed multiple linear
regression analysis (Table II) in
which beta-CTX and PINP served as dependent variables and other
factors served as independent variables. As shown in Table II, TC was a significant
independent predictor of BMD, beta-CTX and PINP and was negatively
correlated with BMD and positively correlated with beta-CTX and
PINP. LDL-C was positively correlated with beta-CTX levels but was
not significantly correlated with BMD or PINP levels.
| Table II.Multiple regression analysis between
BMD, serum Beta-CTX or PINP and TC, LDL-C, HDL-C. |
Table II.
Multiple regression analysis between
BMD, serum Beta-CTX or PINP and TC, LDL-C, HDL-C.
|
Characteristics | BMD | Beta-CTX | PINP |
|---|
| Age | −0.389b | −0.241b | −0.257b |
| BMI | 0.195b | −0.051 | −0.102 |
| To | 0.153a | −0.086 | 0.017 |
| TG | −0.024 | 0.049 | 0.123 |
| TC | −0.175b | 0.169a | 0.166a |
| LDL-C | −0.073 | 0.172a | −0.068 |
| HLD-C | 0.073 | −0.038 | −0.021 |
Effects of HCD and HCD withdrawal on
bone in male rats
Serum cholesterol levels were higher in HCD rats
than levels in the controls (P<0.01), inducing exogenous
hypercholesterolaemia in these rats, and these levels were
diminished in the HCD+NCD rats compared with those in the HCD rats
(P<0.01), and there was no significant difference in TG or body
weight among the three groups (Fig.
1A).
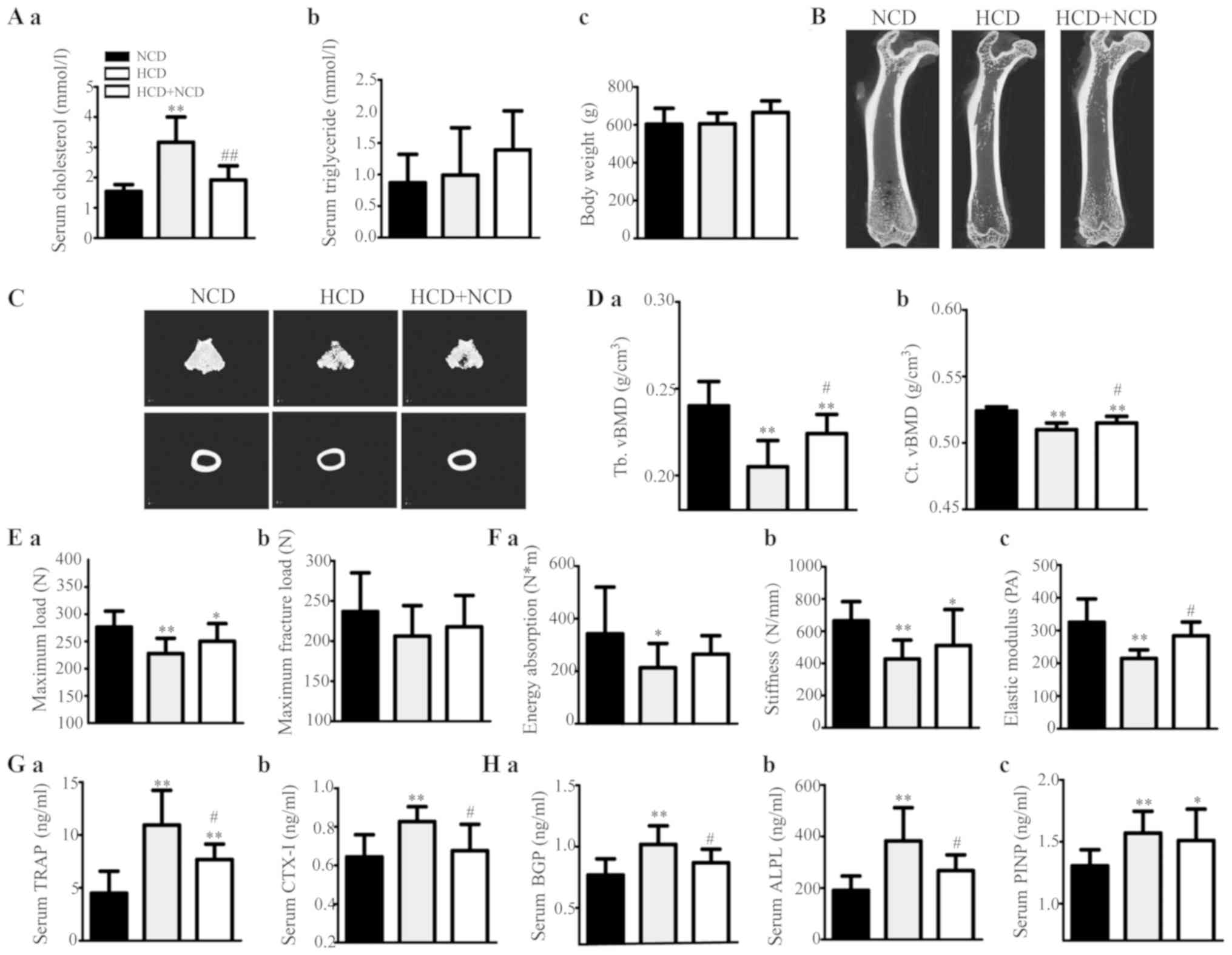 | Figure 1.Effects of exogenous
hypercholesterolaemia induced by HCD administration on male rats.
(A) Serum (a) TC and (b) TG levels, and (c) body weight of the
rats. (B) Representative 2D mCT images showing the trabecular and
cortical microarchitectures of the femurs of each group. (C)
Representative 3D mCT images showing the trabecular
microarchitecture of the distal femur and the cortical
microarchitecture of the midfemoral diaphyses. (D) The trabecular
volumetric BMD (Tb. vBMD) (a) and cortical volumetric BMD (Ct.
vBMD) (b) of the femur, as analysed via mCT. (E and F) The bone
biomechanical parameters of the femoral diaphysis were calculated
from a three-point bending test, and the maximum load (E-a),
maximum fracture load (E-b), energy absorption (F-a), stiffness
(F-b) and elastic modulus (F-c) of the femoral diaphysis were
analysed. (G) The levels of the serum bone resorption markers TRAP
(a) and beta-CTX (b) in rats were measured using ELISA kits. (H)
The levels of serum bone formation markers BGP (a), ALPL (b) and
PINP (c) in rats were measured using ELISA kits. *P<0.05 and
**P<0.01 vs. the corresponding NCD group, #P<0.05
and ##P<0.01 vs. the corresponding HCD group. HCD,
high cholesterol diet; TC, total cholesterol; LDL-C, low-density
lipoprotein cholesterol; HDL-C, high-density lipoprotein
cholesterol; TG, triglyceride; BMD, bone mineral density; BGP, bone
glaprotein; ALPL, alkaline phosphatase; PINP, type I anterior
collagen amino terminal peptide; NCD, normal control diet. |
The bone mCT measurements are shown in Table III. HCD administration decreased
trabecular and cortical vBMD (Fig.
1D) and caused microstructural damage (Fig. 1B and C and Table III); Fig. 1B is two-dimensional and
cross-sectional scanning structure, and Fig. 1C is three-dimensional structure
synthesized by computer analysis, showing the microstructure of
bone in a more detailed manner. vBMD was higher in the HCD+NCD rats
than that in the HCD male rats (Fig.
1D). The levels of mCT 3D microstructural damage were
ameliorated in HCD withdrawal rats compared with those in the HCD
rats (Table III). These results
suggest that HCD passively affected both trabecular and cortical
bone, especially trabecular bone and that dietary modification can
weaken these effects.
| Table III.Micro-CT measurements in male
rats. |
Table III.
Micro-CT measurements in male
rats.
| Parameters
(abbreviations) | NCD | HCD | HCD+NCD | WT |
ApoE−/− |
|---|
| Trebecular
parameter |
|
|
|
|
|
| Percent
bone volume (%) (BV/TV) | 32.615±7.401 |
18.01±3.901b |
25.614±2.834a,d | 42.858±5.627 |
33.137±4.434b |
| Bone
surface density (1/mm) (BS/TV) | 8.533±1.592 |
5.687±1.138b |
6.05±0.907b | 12.359±0.941 |
10.329±0.711b |
|
Trabecular thickness (mm)
(Tb.Th) | 0.142±0.01 |
0.131±0.008b |
0.131±0.003a | 0.125±0.006 |
0.112±0.004b |
|
Trabecular number (1/mm)
(Tb.N) | 2.398±0.492 |
1.45±0.373b |
1.743±0.248b | 3.461±0.334 |
2.916±0.332b |
|
Connectivity density
(1/mm3) (Conn.Dn) | 57.15±15.627 |
24.165±6.569b |
28.796±8.819b | 94.164±8.384 |
110.377±17.264a |
|
Trabecular separation (mm)
(Tb.Sp) | 0.307±0.067 |
0.542±0.116b |
0.452±0.105c | 0.210±0.028 |
0.278±0.051b |
|
Trabecular pattern factor
(1/mm) (Tb.Pf) | 2.66±1.841 |
7.721±2.206b |
5.94±2.038b | −0.837±2.334 |
1.754±1.772a |
|
Structure model index
(SMI) | 1.105±0.25 |
1.782±0.336b |
1.526±0.278b | 0.812±0.246 |
1.267±0.176b |
| Cortical
parameter |
|
|
|
|
|
| Percent
bone volume (%) (BV/TV) | 92.361±0.284 |
91.618±0.277b | 91.888±0.282 | 89.193±0.255 |
88.494±0.29b |
| Bone
surface density (1/mm) (BS/TV) | 6.300±0.68 |
5.871±0.077a | 5.967±0.126 | 7.218±0.086 |
6.999±0.134b |
|
Cortical thickness (mm)
(Ct.Th) | 0.522±0.01 |
0.487±0.06a |
0.518±0.001c | 0.501±0.007 |
0.487±0.008b |
|
Cortical area (mm2)
(Ct.Ar) | 32.409±2.204 |
29.136±1.759b |
31.623±2.325c | 23.074±1.3890 | 24.392±2.358 |
The levels of bone biomechanical parameters were
increased significantly or tended to increase in HCD withdrawal
rats compared with those in HCD rats (Fig. 1E and F). These results suggest that
HCD caused deteriorations in bone strength and that dietary
modification can mitigate these effects.
Our assessment revealed that the levels of serum
bone resorption markers (TRAP and CTX–I) were significantly
increased in HCD rats compared with those in the controls; however,
they decreased after HCD withdrawal (Fig. 1G). Therefore, bone formation
markers (BGP, ALPL and PINP) were identified (Fig. 1H). These results suggest that HCD
administration elevated both bone resorption and bone formation,
and dietary modification can alleviate these effects.
ApoE−/− male rat bone
loss
Serum cholesterol levels were significantly higher
in ApoE−/− male rats than in ApoE+/+ rats
(P<0.01), inducing endogenous hypercholesterolaemia. There were
no differences in TG levels or body weight between the two groups
(Fig. 2A).
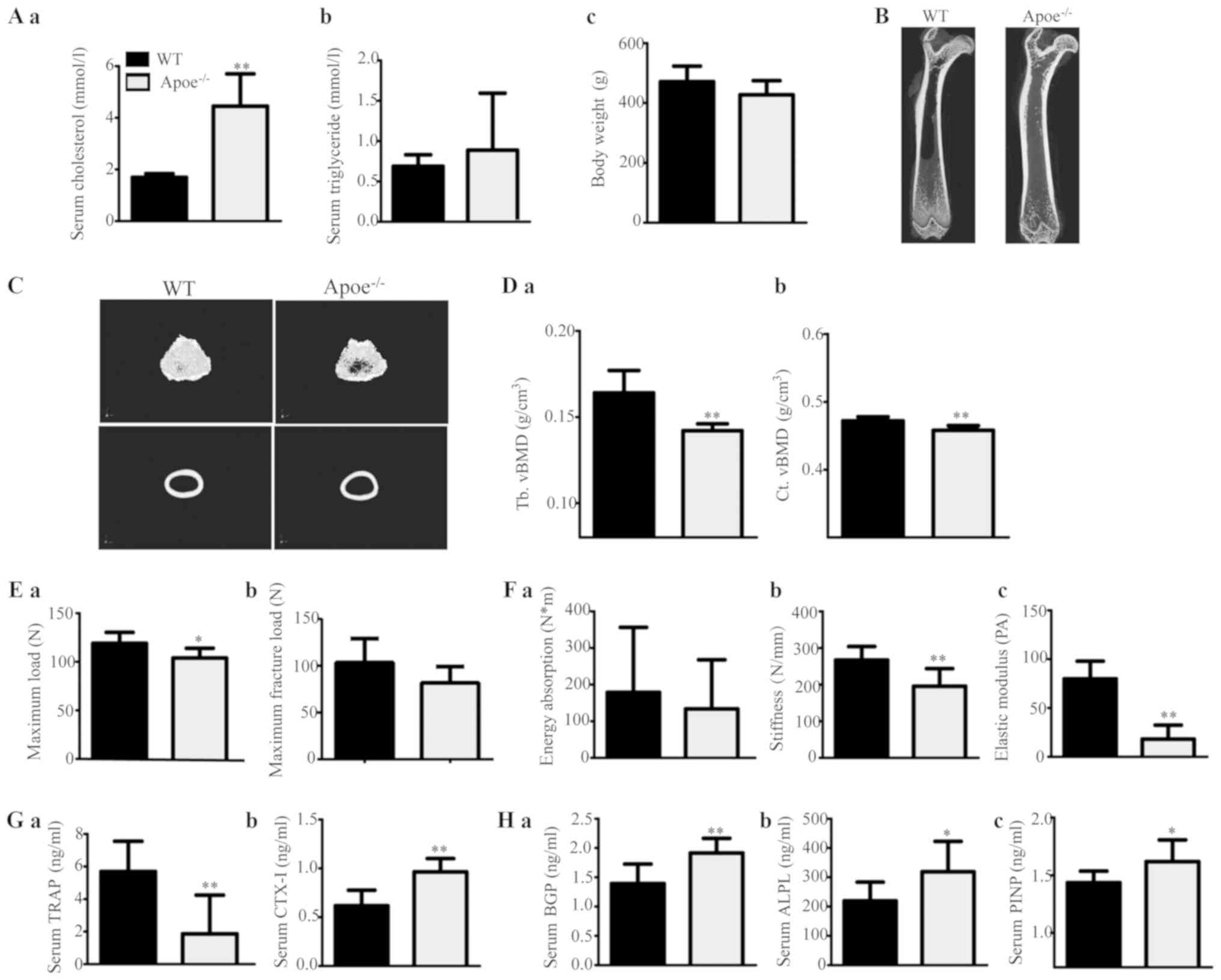 | Figure 2.Effects of endogenous
hypercholesterolaemia induced by ApoE knockout on male rats. (A)
Serum (a) TC and (b) TG levels, and (c) body weight of the rats.
(B) Representative 2D mCT images showing the trabecular and
cortical microarchitectures of the femurs of each group. (C)
Representative 3D mCT images showing the trabecular
microarchitecture of the distal femur and the cortical
microarchitecture of the midfemoral diaphyses. (D) Trabecular
volumetric BMD (Tb. vBMD) (a) and cortical volumetric BMD (Ct.
vBMD) (b) of the femur, as analysed via mCT. (E and F) The bone
biomechanical parameters of the femoral diaphysis were calculated
using a three-point bending test, and the maximum load (E-a),
maximum fracture load (E-b), energy absorption (F-a), stiffness
(F-b) and elastic modulus (F-c) of the femoral diaphysis were
analysed. (G) The levels of the serum bone resorption markers TRAP
(a) and beta-CTX (b) in rats were measured by ELISA kits. (H) The
levels of the serum bone formation markers BGP (a), ALPL (b) and
PINP (c) in rats were measured by ELISA kits. *P<0.05 and
**P<0.01 vs. the corresponding WT group. TC, triglyceride;
LDL-C, low-density lipoprotein cholesterol; HDL-C, high-density
lipoprotein cholesterol; TG, triglyceride; BMD, bone mineral
density; BGP, bone glaprotein; ALPL, alkaline phosphatase; PINP,
type I anterior collagen amino terminal peptide; WT, wild-type;
ApoE−/−, ApoE-knockout (KO). |
The levels of bone mCT measurements are shown in
Table III. ApoE−/−
male rats displayed decreased trabecular and cortical vBMD
(Fig. 2D) and microstructural
damage (Fig. 2B and C) compared
with ApoE+/+ rats. These results suggest that
ApoE−/− rats experienced significant trabecular and
cortical bone loss and damage.
Regarding the biomechanical parameter measurements,
the maximum load, maximum fracture load, stiffness and elastic
modulus of the femoral diaphysis were apparently lower and energy
absorption tended to decrease in ApoE−/− rats compared
with those in ApoE+/+ rats (Fig. 2E and F).
Our assessment revealed that the serum levels of
both bone resorption markers (TRAP and CTX–I) and bone formation
markers (BGP, ALPL and PINP) were increased in ApoE−/−
male rats compared with those in ApoE+/+ rats (Fig. 2G and H). These results suggest that
ApoE−/− rats had elevated bone resorption and bone
formation, improving the bone turnover rate.
Effects of cholesterol on primary
osteoblast function
We examined H&E-stained osteoblasts via
microscopy (Fig. 3A-a). These
osteoblasts displayed cytoplasm that appeared violet in response to
ALP staining (Fig. 3A-b). The
cells were subjected to Alizarin red (Fig. 3A-c) or Von Kossa staining (Fig. 3A-d), revealing the presence of many
round or elliptical opaque red or black nodules, which served as
additional evidence indicating osteoblast mineralization. All of
the above findings suggest that cultured rat calvaria osteoblasts
are ideal experimental osteoblast model cells.
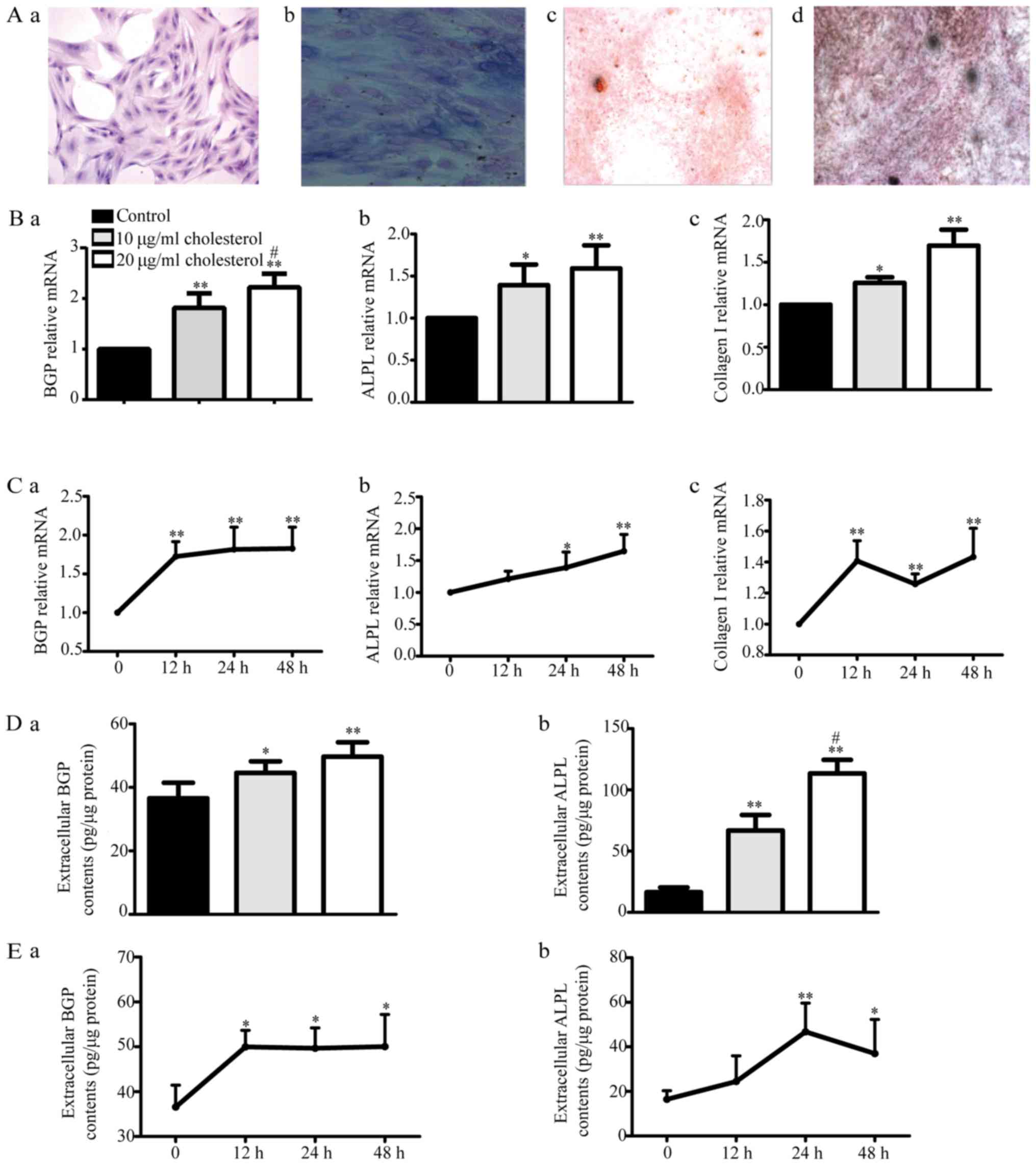 | Figure 3.Effects of cholesterol on primary
osteoblast function. (A) We evaluated primary osteoblast H&E
staining via microscopy. The cells displayed multiple bumps, via
which they appeared to be connected to adjacent cells, as well as
large centrally located or shifted cell nuclei (a; ×100
magnification). The cytoplasm appeared violet after alkaline
phosphatase (ALP) staining (b; ×400 magnification). The osteoblasts
displayed many round or elliptical opaque nodules, which appeared
red after Alizarin red staining (c; ×40 magnification) and black
after Von Kossa staining, after being cultured for 16 days (d; ×40
magnification). (B) BGP (a), ALPL (b) and collagen I (c) mRNA
expression levels were determined by reverse
transcription-quantitative polymerase chain reaction. *P<0.05,
**P<0.01 vs. control, #P<0.05 vs. 10 µg/ml
cholesterol. (C) BGP (a), ALPL (b) and collagen I (c) mRNA
expression levels after 10 µg/ml cholesterol treatment for 12, 24
and 48 h. *P<0.05, **P<0.01 vs. control. (D) Extracellular
BGP (a) and ALPL (b) content was measured by ELISA kits.
*P<0.05, **P<0.01 vs. control, #P<0.05 vs. 10
µg/ml cholesterol. (E) Extracellular BGP (a) and ALPL (b) content
after treatment with 10 µg/ml cholesterol for 12, 24 and 48 h.
*P<0.05, **P<0.01 vs. control. BGP, bone glaprotein; ALPL,
alkaline phosphatase. |
With increasing cholesterol doses, BGP, ALPL and
collagen I mRNA expression as well as BGP and ALPL protein levels
gradually increased compared with those in the control group
(Fig. 3B and D) after treatment
for 24 h, but there were no differences in their levels among the
different times tested (12, 24 and 48 h) after 10 µg/ml cholesterol
treatment (Fig. 3C and E). These
results suggested that cholesterol could increase the mRNA and
protein expression levels of osteoblast functional genes in a
dose-dependent but non-time-dependent manner.
Discussion
Currently, there is a dearth of information in the
literature regarding the effects of hypercholesterolaemia on male
bone metabolism. However, male osteoporosis (OP) should draw more
attention from clinicians worldwide. Elucidating the effects of
hypercholesterolaemia on male bone metabolism may aid in the search
for answers regarding the relationship between
hypercholesterolaemia and male OP and the underlying mechanisms and
will further our understanding of the pathomechanisms underlying
the relationship between the two diseases to reduce the financial
burden imposed by male OP and OP-related hip fractures.
T2 diabetes mellitus (DM), low testosterone (To) and
obesity are often associated with elevated plasma cholesterol
levels (10–12). Diabetes mellitus (both type 1 and
type 2) (13), testosterone
deficiency (14,15), hyperthyroidism and hypothyroidism
(16), body mass index (BMI)
(17), and hypertriglyceridemia
(18,19) reportedly affect bone mineral
density (BMD). In this clinical study, we excluded participants
with thyroid diseases, diabetes and hypertriglyceridemia according
to clinical laboratory test results. There were no differences in
BMI or serum fasting plasma glucose (FPG), HbAc1, To, FT4, thyroid
stimulating hormone (TSH), 25(OH)D and triglyceride (TG) levels
between hypercholesterolaemia and control subjects. Therefore, we
investigated the effects of hypercholesterolaemia on male bone,
excluding the effects of the abovementioned parameters. Multiple
regression analysis suggested that cholesterol was a significant
independent predictor of BMD and negatively correlated with BMD.
Some clinical studies have demonstrated that cholesterol-lowering
statin drugs increase BMD and reduce the risk of OP-associated
fractures (20,21), supporting our results. We also
found that serum cholesterol was a significant independent
predictor of beta-CTX and PINP levels and positively correlated
with both. Beta-CTX fragments are released into the blood after
type I collagen is dissolved by osteoclasts and are recognized as
clinical indices of osteoclast activity and bone resorption. PINP
directly reflects the rate of osteoblast-mediated collagen
synthesis and is recognized as a clinical index of osteoblast
activity and bone formation. Thus, we hypothesized that
hypercholesterolaemia can promote the development of male OP by
promoting both osteoclast-mediated bone resorption and
osteoblast-mediated osteogenesis. Our data were inconsistent with
those of a previous study (22).
Majima et al (22) noted
that higher TC levels were associated with higher bone turnover
marker levels in female patients but not in male patients, perhaps
because their study did not exclude other factors, such as male
gonadal hypofunction, as stated by the authors in the indicated
report. Importantly, the authors did not analyze the association
between cholesterol and BMD. We used multiple regression analysis
to exclude the effects of FPG, HbAc1, To, FT4, TSH, 25(OH)D and TG
on bone, and we believe that our findings are more convincing.
Bone microstructural damage results in low BMD or
OP, and reduced bone strength is correlated with a higher risk of
bone fracture. Thus, to test our clinical discovery and further
explore the effects of hypercholesterolaemia on male bone
microstructure and bone strength, we designed two male animal
models. High cholesterol diet (HCD)-induced exogenous
hypercholesterolaemia and ApoE-knockout (KO)-induced endogenous
hypercholesterolaemia were observed in male rats.
HCD administration significantly increased serum
cholesterol levels but not serum TG levels or body weight. Halade
et al (23) reported that
high fat diet-induced hypertriglyceridemia promotes bone marrow
adiposity with a lower bone mass. Tatsumi et al (24) reported that higher weights have
protective effects on bone metabolism in rodents. Obesity reduced
bone density in rapidly growing male rats (25). Serum TG levels and body weights
were unchanged in the HCD groups compared to those in the controls,
indicating that the effects of serum TG levels and weight were
excluded in this study. High cholesterol decreased BMD and
significantly increased the levels of both bone resorption markers,
such as CTX-1 and TRAP, and bone formation markers, such as bone
glaprotein (BGP), alkaline phosphatase (ALPL), PINP, indicating
that high cholesterol promoted osteoclast-mediated bone resorption
and osteoblast-mediated bone formation, resulting in the increased
risk of osteoporosis in male rodents. These findings were somewhat
inconsistent with those of previous research involving female
rodents (26). You et al
(26) recently reported that HCD
administration did not promote bone resorption but inhibited bone
formation in female rats. We attribute the above discrepancies
mainly to the difference in the genders of the mice used in the two
studies. Other authors aimed to study the effects of an HCD on
female OP. Since differences in bone metabolism due to differences
in endocrine and paracrine factors exist between the sexes
(27–29), the effects of hypercholesterolaemia
on male and female bone metabolism and the underlying mechanisms
may be different. In addition, the authors of one study used an HCD
containing 20% lard, which induces obesity and high TG, leading to
bone formation inhibition (25).
Tintut et al (30) recently
reported that hypercholesterolaemia promotes osteoclastic
differentiation and resorptive activity in vivo, consistent
with our study. These findings supported our clinical hypothesis.
In our animal study, high cholesterol caused trabecular and
cortical microstructural damage and reduced bone mechanical
strength. Therefore, we believe that high cholesterol damages bone
microstructure and decreases bone mechanical strength by
excessively enhancing bone transformation, leading to a higher risk
of OP and a higher risk of fracture in males. In addition, our
findings indicate that the changes observed in the HCD group can be
slightly attenuated by withdrawal of the HCD, suggesting that diet
modification can ameliorate the negative effects of
hypercholesterolaemia on bone.
The ApoE-KO rat is a useful model for studying the
functions of congenital hypercholesterolaemia. Rats lacking ApoE
have congenital hypercholesterolaemia, as demonstrated by their
elevated serum cholesterol levels and unchanged serum TG levels
(31,32). To the best of our knowledge, this
study is the first to investigate the effects of congenital
hypercholesterolaemia on male bone. In our ApoE-KO male rat model,
ApoE−/− rats displayed an enormous increase in their
serum TC levels but no change in their serum TG levels or weights
compared to those in control rats. We found that the effects of
endogenous hypercholesterolaemia on BMD, bone turnover, bone
microstructure and bone strength were similar to those of exogenous
hypercholesterolaemia. We regarded endogenous hypercholesterolaemia
as a positive-control group. Our animal findings suggest that both
endogenous and exogenous hypercholesterolaemia promote the
development of male OP by exceedingly promoting both osteoclast
activity and osteoblast activity.
As the above results show, high cholesterol can
boost both osteoclast and osteoblast functions in vivo. To
research the direct influence of cholesterol on both osteoclast and
osteoblast functions, we designed an in vitro cell
experiment. Since many cell studies have shown that cholesterol
promotes enhanced osteoclast differentiation and osteoclast
activity (33) and the conclusion
of the effects of cholesterol on osteoblasts is contradictory, we
studied only the effects of cholesterol on osteoblast function. We
found that free cholesterol increased the expression levels of
genes involved in osteoblastic function and that cholesterol can
promote osteoblastic function. Li et al (34) documented that cholesterol-treated
mesenchymal stem cells (MSCs) showed increased expression levels of
osteogenic lineage markers, supportive of our conclusion. Liu et
al (35) reported that the
proliferation/viability of primary alveolar osteoblast cells (AOBs)
was significantly decreased by simvastatin (a cholesterol-lowering
drug) treatment at different concentrations, findings consistent
with ours. However, You et al (26) reported that free cholesterol
inhibits the proliferation and differentiation of osteoblasts in
vitro, findings inconsistent with ours. We attribute the above
difference to the fact that the free cholesterol treatment doses
used in that study were different from those used in this study, as
our cholesterol treatment doses were lower. In the present study,
we observed substantial death in the osteoblast population treated
with cholesterol at a concentration higher than 20 µg/ml; thus, we
chose a cholesterol treatment dose below 20 µg/ml in subsequent
experiments to avoid the toxicity induced by cholesterol levels
higher than 20 µg/ml. The authors of the other study used the
MC3T3-E1 osteoblast line, and we used rat calvaria primary
osteoblasts, which we deemed ideal osteoblast model cells. Thus,
our in vitro experimental results conform more to those of
our in vivo animal experiments than to those of the other
study. Thus, we believe that our results are more truthful.
The present study had some limitations. In our in
vitro experiments, we did not investigate the effects of
cholesterol on osteoclasts since many studies have found that
cholesterol promotes osteoclastic differentiation and function, the
results of which were consistent with those of our study. However,
additional studies regarding this issue are needed, as are accurate
evaluations of cell signal transduction in osteoblasts and cell
function and signal transduction in osteoclasts.
In summary, we found that hypercholesterolaemia can
promote the development of male OP by excessively promoting both
osteoclast activity and osteoblast activity, which excessively
enhances the bone transformation rate in men. Both endogenous and
exogenous hypercholesterolaemia in two male animal experimental
models were used to validate the clinical findings. Again, we found
that hypercholesterolaemia damaged the bone microstructure, thus,
increasing the risk of osteopenia or OP and reducing bone strength,
resulting in a higher risk of fracture. Many in vitro
studies have shown that cholesterol enhances osteoclast
differentiation and osteoclast activity (34). In vitro, cholesterol
directly upregulated the functions of osteoblasts. Based on these
results, the detrimental effects of hypercholesterolaemia on bone
health probably represent a higher risk of high-turnover
osteoporosis and future fractures in men. In conclusion, we wish to
emphasize the importance of preventing and treating
hypercholesterolemia to prevent osteoporotic bone loss and of
monitoring bone metabolic markers, preferably in conjunction with
lowering BMD, in men with hypercholesterolemia for the effective
prevention of bone loss and subsequent fractures. In addition, our
findings provide a theoretical basis for the development of
treatments for high cholesterol-induced osteoporosis.
Acknowledgements
We are very thankful to the medical postgraduates of
Shandong Provincial Hospital and the Jinan Health Organization for
their cooperation in sample collection, obtaining consent from
patients and filling out the voluntary questionnaire. We are very
thankful to the Clinical Laboratory Department of Shandong
Provincial Hospital for providing the serum indices for the
clinical tests.
Funding
The present study was supported by grants from the
Key Research and Development Plan Project of Shandong Province
(grant no. 2016 GSF201025), the National Natural Science Foundation
of China (grant no. 81370892) and the Shandong Taishan Scholars
Specially-invited Expert Plan.
Availability of data and materials
The datasets obtained and/or analysed in the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
JX conceived the study and participated in its
design and coordination; YZ conceived and participated in the
design of the study, carried out the animal and cell experiments,
participated in the data analysis and drafting, and wrote the
manuscript; TD and HoZ participated in the design of the study and
carried out some of the cell experiments. HaZ, QG and MZ guided the
epidemiological investigation, the sample collection, the
questionnaire and serum index tests. CY and SS participated in the
design of the study and carried out some animal experiments. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The study protocol using human subjects was approved
by the Ethics Committee of Provincial Hospital Affiliated with
Shandong University (Jinan, China) and all participants provided
written informed consent before participating in the study. The
animal experimental protocol was approved by the Animal Ethics
Committee of Shandong Provincial Hospital.
Patient consent for publication
Not applicable.
Competing interests
Τhe authors declare that they have no competing
interests.
References
|
1
|
Osteoporosis, . Review of the evidence for
prevention, diagnosis and treatment and cost-effectiveness
analysis. Introduction. Osteoporos Int. 8 (Suppl 4):S7–S80. 1998.
View Article : Google Scholar
|
|
2
|
Looker AC, Orwoll ES, Johnston CC Jr,
Lindsay RL, Wahner HW, Dunn WL, Calvo MS, Harris TB and Heyse SP:
Prevalence of low femoral bone density in older U.S. adults from
NHANES III. J Bone Miner Res. 12:1761–1768. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cummings SR and Melton LJ: Epidemiology
and outcomes of osteoporotic fractures. Lancet. 359:1761–1767.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Trivedi DP and Khaw KT: Bone mineral
density at the hip predicts mortality in elderly men. Osteoporos
Int. 12:259–265. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wu LY, Yang TC, Kuo SW, Hsiao CF, Hung YJ,
Hsieh CH, Tseng HC, Hsieh AT, Chen TW, Chang JB, et al: Correlation
between bone mineral density and plasma lipids in Taiwan. Endocr
Res. 29:317–325. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huang Y, Gao L, Xie X and Tan SC:
Epidemiology of dyslipidemia in Chinese adults: Meta-analysis of
prevalence, awareness, treatment, and control. Popul Health Metr.
12:282014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zheng G: Relevant Guidelines for the
management of abnormal lipids in 2016 and updated interpretation of
expert consensus. World Clinical Drugs. 7:441–444. 2017.
|
|
8
|
Turner CH and Burr DB: Basic biomechanical
measurements of bone: A tutorial. Bone. 14:595–608. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li XF, Zhao JM, Su W, et al: Primary
culture and identification of rat osteoblasts. J Clin Rehabil
Tissue Eng Res. 15:990–994. 2011.
|
|
10
|
Mooradian AD: Dyslipidemia in type 2
diabetes mellitus. Nat Clin Pract Endocrinol Metab. 5:150–159.
2009.PubMed/NCBI
|
|
11
|
Koshiyama H, Wada Y and Nakamura Y:
Hypercholesterolemia as a possible risk factor for osteopenia in
type 2 diabetes mellitus. Arch Intern Med. 161:1678. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang N, Zhang H, Zhang X, Zhang B, Wang
F, Wang C, Zhao M, Yu C, Gao L, Zhao J and Guan Q: The relationship
between endogenous testosterone and lipid profile in middle-aged
and elderly Chinese men. Eur J Endocrinol. 170:487–494. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wongdee K and Charoenphandhu N:
Osteoporosis in diabetes mellitus: Possible cellular and molecular
mechanisms. World J Diabetes. 2:41–48. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gaffney CD, Pagano MJ, Kuker AP, Stember
DS and Stahl PJ: Osteoporosis and low bone mineral density in men
with testosterone deficiency syndrome. Sex Med Rev. 3:298–315.
2015. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Saad F, Röhrig G, von Haehling S and
Traish A: Testosterone deficiency and testosterone treatment in
older men. Gerontology. 63:144–156. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsourdi E, Rijntjes E, Köhrle J, Hofbauer
LC and Rauner M: Hyperthyroidism and Hypothyroidism in male mice
and their effects on bone mass, bone turnover, and the Wnt
inhibitors sclerostin and Dickkopf-1. Endocrinology. 156:3517–3527.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reid IR: Relationships between fat and
bone. Osteoporos Int. 19:595–606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Murdolo G, Bartolini D, Tortoioli C,
Piroddi M, Iuliano L and Galli F: Lipokines and oxysterols: Novel
adipose-derived lipid hormones linking adipose dysfunction and
insulin resistance. Free Radic Biol Med. 65:811–820. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Verma S, Rajaratnam JH, Denton J, Hoyland
JA and Byers RJ: Adipocytic proportion of bone marrow is inversely
related to bone formation in osteoporosis. J Clin Pathol.
55:693–698. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Edwards CJ, Hart DJ and Spector TD: Oral
statins and increased bone-mineral density in postmenopausal women.
Lancet. 355:2218–2219. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Meier CR, Schlienger RG, Kraenzlin ME,
Schlegel B and Jick H: HMG-CoA reductase inhibitors and the risk of
fractures. JAMA. 283:3205–3210. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Majima T, Shimatsu A, Komatsu Y, Satoh N,
Fukao A, Ninomiya K, Matsumura T and Nakao K: Increased bone
turnover in patients with hypercholesterolemia. Endocr J.
55:143–151. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Halade GV, Rahman MM, Williams PJ and
Fernandes G: High fat diet-induced animal model of age-associated
obesity and osteoporosis. J Nutr Biochem. 21:1162–1169. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tatsumi S, Ito M, Asaba Y, Tsutsumi K and
Ikeda K: Life-long caloric restriction reveals biphasic and
dimorphic effects on bone metabolism in rodents. Endocrinology.
149:634–641. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen JR, Lazarenko OP, Wu X, Tong Y,
Blackburn ML, Shankar K, Badger TM and Ronis MJ: Obesity reduces
bone density associated with activation of PPARγ and suppression of
Wnt/β-catenin in rapidly growing male rats. PLoS One. 5:e137042010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
You L, Sheng ZY, Tang CL, Chen L, Pan L
and Chen JY: High cholesterol diet increases osteoporosis risk via
inhibiting bone formation in rats. Acta Pharmacol Sin.
32:1498–1504. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yan L, Prentice A, Zhou B, Zhang H, Wang
X, Stirling DM, Laidlaw A, Han Y and Laskey A: Age- and
gender-related differences in bone mineral status and biochemical
markers of bone metabolism in Northern Chinese men and women. Bone.
30:412–415. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Khosla S, Melton LJ III and Riggs BL:
Osteoporosis: Gender differences and similarities. Lupus.
8:393–396. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ebbesen EN, Thomsen JS, Beck-Nielsen H,
Nepper-Rasmussen HJ and Mosekilde L: Age- and gender-related
differences in vertebral bone mass, density, and strength. J Bone
Miner Res. 14:1394–1403. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tintut Y, Morony S and Demer LL:
Hyperlipidemia promotes osteoclastic potential of bone marrow cells
ex vivo. Arterioscler Thromb Vasc Biol. 24:e6–e10. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wei S, Zhang Y, Su L, He K, Wang Q, Zhang
Y, Yang D, Yang Y and Ma S: Apolipoprotein E-deficient rats develop
atherosclerotic plaques in partially ligated carotid arteries.
Atherosclerosis. 243:589–592. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ekuni D, Yoneda T, Endo Y, Kasuyama K,
Irie K, Mizutani S, Azuma T, Tomofuji T and Morita M: Occlusal
disharmony accelerates the initiation of atherosclerosis in apoE
knockout rats. Lipids Health Dis. 13:1442014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Luegmayr E, Glantschnig H, Wesolowski GA,
Gentile MA, Fisher JE, Rodan GA and Reszka AA: Osteoclast
formation, survival and morphology are highly dependent on
exogenous cholesterol/lipoproteins. Cell Death Differ. 11 (Suppl
1):S108–S118. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li H, Guo H and Li H: Cholesterol loading
affects osteoblastic differentiation in mouse mesenchymal stem
cells. Steroids. 78:426–433. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu S, Bertl K, Sun H, Liu ZH, Andrukhov O
and Rausch-Fan X: Effect of simvastatin on the osteogenetic
behavior of alveolar osteoblasts and periodontal ligament cells.
Hum Cell. 25:29–35. 2012. View Article : Google Scholar : PubMed/NCBI
|














