Introduction
The aging of skin progresses by two contributing
causes: time and photodamage. Chronological skin aging progresses
by time and is observed as damage-prone skin with a dry and thin
epidermis (1). Photoaging, which
is stimulated by ultraviolet (UV) irradiation, is characterized by
damage to skin DNA, degraded collagen build-up and excessive
oxidative stress (2,3). The advancement of aging due to UV
irradiation occurs in specific skin cells called keratinocytes,
fibroblasts and neutrophils. Among the different types of UV rays
that can reach the skin surface, UVB exposure is considered to be
the most harmful type of UV irradiation that acts predominantly on
the epidermis layer and keratinocytes (4,5). UVB
irradiation activates the nuclear factor-κB (NF-κB) pathway
resulting in the elevated activity and expression of matrix
metalloproteinases and cytokines (6). Activated cytokines induce the
degradation of the extracellular matrix. Continued UVB exposure
causes accumulation of the degraded extracellular matrix and
manifests itself as wrinkles and brittle skin (7).
UVB irradiation is also known to generate reactive
oxygen species (ROS) and is linked to inflammatory responses in
skin such as sunburn, erythema and edema (8,9). The
main share of skin damage that occurs by prolonged UVB irradiation
is associated with ROS and ROS-linked inflammation. The harmful
effects of ROS in skin are generally negated by antioxidant defense
mechanism of the body consisting of antioxidant enzymes and free
radical scavenging molecules. However, excessive ROS as a result of
continuous UVB exposure lead to reaction with fatty acids and
initiation of lipid peroxidation, which at the end causes cellular
damage (10,11). On the other hand, UVB-stimulated
overexpression of COX-2 and activation of NF-κB play a crucial role
in the progression of the inflammatory response in the skin
together with the activation of several cytokines including tumor
necrosis factor (TNF)-α, interleukin-6 (IL-6) and IL-1β (12–14).
UVB-induced overexpression of inflammatory cytokines and their
roles in the deteriorating effects on the skin has been reported in
detail (15).
Natural antioxidants, mainly bioactive substances
from plants are important compounds against the photoaging effect
of solar irradiation (16). These
bioactive agents from natural sources are potent antioxidants,
which also can alleviate inflammatory responses (17), help skin to recover from
photodamage (18,19) and also prevent further solar
radiation damages (20). Reports
indicate promising health beneficial effects of dietary
phytochemicals in protecting skin damage and nourishing the skin
following photodamage (21).
Caffeoylquinic acid (CQA) is one of the naturally occurring
abundant phytochemicals found in a variety of sources including but
not limited to coffee beans, sweet potato, propolis and other
plants. In the literature, CQA derivatives have been reported as
bioactive agents that may be regarded as promising antioxidants
(22), antibacterial (23) and antitumor (24) agents. Recently, various CQA-related
derivatives have been credited as inhibitors of UV-linked oxidative
stress and skin damage (25–27).
In addition, studies have shown that they alleviate skin disorders
secondary to their free radical scavenging effect (25). In this context, the present study
aimed to analyze the potential of 3,5-dicaffeoyl-epi-quinic acid
(DCEQA) against UVB-induced oxidative stress and inflammatory
response in the HaCaT immortal keratinocyte cell line.
Materials and methods
Plant materials and isolation
The sample (3 kg) of A. gmelinii was air
dried and cut into small pieces prior to maceration. Ground samples
were extracted twice with methylene chloride
(CH2Cl2) for 24 h at room temperature. The
extract was concentrated and dried by evaporation in vacuo,
using a vacuum rotary evaporator (RV 10 Series, IKA, Staufen,
Germany), yielding a sticky crude extract. Remaining ground samples
were then subjected to extraction again, twice with methanol
(MeOH), using the same procedure as above. The combined crude
extracts from the CH2Cl2 and MeOH extraction
were partitioned between CH2Cl2 and water
(H2O). The dichloromethane layer was evaporated under
reduced pressure and the residue was repartitioned between
n-hexane and 85% aqueous MeOH. The H2O layer was
also further partitioned between n-BuOH and H2O.
A portion of the n-BuOH fraction (10.74 g) was further
subjected to separation by fractionation between EtOAc and
H2O. The EtOAc layer was concentrated using a rotary
evaporator to obtain an extract of 3.10 g, a portion (74.4 mg) of
which was subjected to isolation via preparative thin layer
chromatography on silica gel using EtOAc/MeOH/H2O
(30:5:4) as a solvent and yielded the compound (18 mg) as a white
powder. The molecular formula of the compound was defined as
C25H24O12 and its purity and
structure were confirmed with HPLC and 1H- and
13C-NMR spectra results (Table I), which were compared with
previous reports of similar compounds (28) for further identification of the
compound. The compound was identified as 3,5-dicaffeoyl-epi-quinic
acid (DCEQA) and was dissolved in methanol and diluted with
Dulbecco's modified Eagle's Medium (DMEM) for further
experiments.
 | Table I.1H and 13C NMR spectral data for
DCEQA. |
Table I.
1H and 13C NMR spectral data for
DCEQA.
| Position | δH | δC |
|---|
| 1 |
| 76.3s |
| 2 | 2.11 (2H, m) | 40.6t |
| 3 | 5.55 (1H, dt,
J=10.0, 5.8 Hz) | 72.4d |
| 4 | 3.91 (1H, dd,
J=9.9, 3.4 Hz) | 73.0d |
| 5 | 5.39 (1H, m) | 74.4d |
| 6 | 2.04 (1H, m), 2.28
(1H, dd, J=15.2, 3.4 Hz) | 37.5t |
| 1 ′ |
| 127.8s, 128.0s |
| 2 ′ | 7.06, 7.08 (each
1H, d, J=2.0 Hz) | 115.2d |
| 3 ′ |
| 146.8s, 146.9s |
| 4 ′ |
| 149.2s, 149.4s |
| 5 ′ | 6.78 (2H, d, J=8.2
Hz) | 116.4d |
| 6 ′ | 6.96, 6.97 (each
1H, dd, J=8.2, 2.0 Hz) | 122.9d |
| 7 ′ | 7.59, 7.62 (each
1H, d, J=15.8 Hz) | 146.6d, 146.6d |
| 8 ′ | 6.31, 6.43 (each
1H, d, J=15.8 Hz) | 115.4d, 115.9d |
| 9 ′ |
| 169.0s, 169.4s |
| COOH |
| 181.3s |
Cell culture and UVB irradiation
Human HaCaT immortal keratinocyte cell line was
purchased from Cell Line Service (Eppelheim, Germany). Cells were
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 10% fetal bovine serum (FBS), 100 µg/ml
penicillin-streptomycin antibiotics and 2 mM glutamine (Gibco-BRL;
Thermo Fisher Scientific, Inc.) and incubated in a humidified
atmosphere of 5% CO2 at 37°C. Cells were trypsinized and
subcultured after reaching ~90% confluency for further
experiments.
Cells were irradiated by UVB using a Bio-Sun UV
Irradiation System (Vilber Lourmat, Marine, France) fitted with a
312-nm UVB source designed for microplates. HaCaT cells grown in
microplates were irradiated at 15 mJ/cm2 UVB dose, which
is the approximate dose of UVB irradiation (24 h) thought to induce
oxidative stress-mediated photoaging in HaCaT Cells without
significant mortality or malformation (29,30).
The dose was validated in preliminary experiments on HaCaT cells
following 24 h exposure (data not shown). Cells were irradiated in
phosphate-buffered saline (PBS) without the plastic lid. When the
irradiation received matched the desired programmed energy, the UVB
irradiation stopped automatically, and subsequently the cells were
incubated with DMEM without FBS until analysis.
Cell viability assay
The viability of HaCaT cells was analyzed by using
ability of viable cells to convert MTT to an insoluble formazan
product that can be quantified with a colorimetric method. Cells
were cultured in 96-well plates and incubated for 24 h prior to
washing with PBS and treatment with different concentrations (1, 5
and 10 µM) of DCEQA introduced with a serum-free fresh medium.
After incubation for 24 h, the supernatant was removed and 100 µl
of 1 mg/ml MTT in PBS was added to the culture wells. The MTT
solution was then removed after 4 h of incubation and 50 µl DMSO
was introduced to each well to stop the reaction and quantify the
converted MTT. Optical density of the wells was measured at a
wavelength of 540 nm using a GENios® microplate reader
(Tecan Group, Ltd., Mannedorf, Switzerland). MTT test was conducted
in triplicate wells for each condition and repeated at least
thrice. Viability of the cells was plotted as a relative percentage
against the untreated control cell group.
Determination of intracellular ROS
generation
Intracellular generation of ROS was determined using
an oxidizing radical species-sensitive dye 2′,7′-dichlorofluorescin
diacetate (DCFH-DA). HaCaT cells that were grown in fluorescence
microtiter 96-well plates and incubated for 24 h were loaded with
20 µM DCFH-DA in PBS and incubated for 20 min in the dark at room
temperature. Cells were then treated with different concentrations
of DCEQA and incubated for 1 h. After washing the cells with PBS
three times, 500 µM H2O2 dissolved in PBS was
added to the cells. Using a GENios® microplate reader
(Tecan Group, Ltd.) the plate fluorescence intensity was read every
30 min for 3 h at an excitation wavelength of 485 nm and emission
wavelength of 528 nm to detect the 2′,7′-dichlorofluorescein (DCF)
which was formed via oxidation of DCFH in the cells by ROS.
Dose-dependent and time-dependent changes in DCF fluorescence
intensity were plotted and compared with untreated control and not
irradiated blank cells.
Reverse transcription-semi
quantitative polymerase chain reaction (RT-sqPCR) analysis
Total RNA was isolated from UVB-irradiated and
unexposed HaCaT keratinocytes treated with/without DCEQA using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). For synthesis of cDNA, RNA (2 µg) and oligo(dT) were mixed
in RNase-free water. This mixture was denatured at 70°C for 5 min
and cooled immediately. RNA reverse transcription was carried out
in a master mix containing 1X RT buffer, 1 mM dNTPs, 500 ng
oligo(dT), 140 units M-MLV reserve transcriptase and 40 units RNase
inhibitor at 42°C for 60 min and at 72°C for 5 min using an
automatic T100 Thermo Cycler (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). The target cDNA was amplified using the following sense
and antisense primers: Forward 5′-GGA-GCC-AGC-TCC-CTC-TAT-TT-3′ and
reverse 5′-GGC-TAC-ATG-GGA-ACA-GCC-TA-3′ for TNF-α; forward
5′-AGA-AGG-AAA-TGG-CTG-CAG-AA-3′ and reverse
5′-GCT-CGG-CTT-CCA-GTA-TTG-AG-3′ for COX-2; forward
5′-AGT-TGC-CTT-CTT-GGGACT-GA-3′ and reverse
5′-CAG-AAT-TGC-CAT-TGCACA-AC-3′ for IL-6; forward
5′-CTG-TCC-TGC-GTG-TTG-AAA-GA-3′ and reverse
5′-TTC-TGC-TTG-AGA-GGT-GCT-GA-3′ for IL-1β; forward
5′-AGG-GCA-TCA-TCA-ATT-TCG-AG-3′ and reverse
5′-TGC-CTC-TCT-TCA-TCC-TTT-GG-3′ for SOD-1; forward
5′-CAC-GCA-TAT-ACC-CGC-TAC-CT-3′ and reverse
5′-AAG-GCG-GTC-TTA-GCC-TCT-TC-3′ for HO-1; forward
5′-CCA-CAG-CTG-AGA-GGG-AAA-TC-3′ and reverse
5′-AAG-GAA-GGC-TGG-AAA-AGA-GC-3′ for β-actin. For the sqPCR
amplification, the thermocycling conditions consisted of 30 cycles
of 95°C for 45 sec, 60°C for 1 min and 72°C for 45 sec. The final
PCR products were separated by agarose gel (1.5%) electrophoresis
for 30 min at 100 V. Gels were then stained with 1 mg/ml ethidium
bromide and visualized by UV light using Davinch-Chemi imager™
(CAS-400SM, Seoul, Korea). Band densities were analyzed using
MultiGauge software (v3.0; Fujifilm, Tokyo, Japan).
Immunoblotting
Western blotting was performed according to standard
procedures. Briefly, cells were lysed in RIPA lysis buffer
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 4°C for 30 min.
Protein amount was measured using a bicinchoninic protein assay kit
(cat. no. 23225; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Equal amounts (25 µg) of protein samples
were separated by 12% SDS-polyacrylamide gel electrophoresis,
transferred onto a polyvinylidene fluoride membrane (Amersham
Pharmacia Biotech; GE Healthcare Life Sciences, Little Chalfont,
UK), blocked with 5% skimmed milk for 1 h at room temperature and
hybridized with the primary antibodies (diluted 1:1,000) overnight
at 4°C. Anti-COX-2 (cat. no. ab15191; Abcam, Cambridge, UK),
anti-IL-6 (cat. no. ab6672; Abcam), anti-TNF-α (cat. no. ab9739;
Abcam) and anti-superoxide dismutase (SOD)-1 (cat. no. sc-11407;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) antibodies were
polyclonal rabbit antibodies, while anti-IL-1β (cat. no. 12242;
Cell Signaling Technology, Inc., Danvers, MA, USA), anti-heme
oxygenase (HO)-1 (cat. no. sc-136960; Santa Cruz Biotechnology,
Inc.), anti-Nrf-2 (cat. no. sc-365949; Santa Cruz Biotechnology,
Inc.) and anti-β-actin (cat. no. sc-47778; Santa Cruz
Biotechnology, Inc.) antibodies were polyclonal mouse antibodies.
After incubation with horseradish-peroxidase-conjugated anti-mouse
(cat. no. 7076; Cell Signaling Technology, Inc.) or anti-rabbit
(cat. no. 7074; Cell Signaling Technology, Inc.) secondary
antibodies (diluted 1:1,000) at room temperature for 1 h,
immunoreactive proteins were detected using a chemiluminescence ECL
assay kit (Amersham Pharmacia Biosciences; GE Healthcare Life
Sciences) according to the manufacturer's instructions. Western
blot bands were visualized using a Davinch-Chemi imager™
(CAS-400SM, Seoul, Korea). Band densities were analyzed using
MultiGauge software (v3.0; Fujifilm, Tokyo, Japan).
Statistical analysis
Data are presented as the mean ± standard deviation
of three repeated experiments. Differences between the means of the
individual groups were analyzed using the analysis of variance
(ANOVA) procedure of the Statistical Analysis System, SAS v9.1 (SAS
Institute, Cary, NC, USA) followed by Duncan's multiple range
tests. The correlation between treatment dose and cell viability
was determined by Pearson correlation analysis, and the correlation
coefficient was expressed as the r-value. P<0.05 was considered
to indicate a statistically significant difference.
Results
Cytotoxicity of DCEQA and UVB exposure
in HaCaT cells
Prior to the in vitro analysis of the
protective effect of DCEQA against UVB-induced oxidative stress,
its biocompatibility was tested by
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT)
assay. Following a 48 h sample treatment at different
concentrations (1, 5 and 10 µM), DCEQA did not show any significant
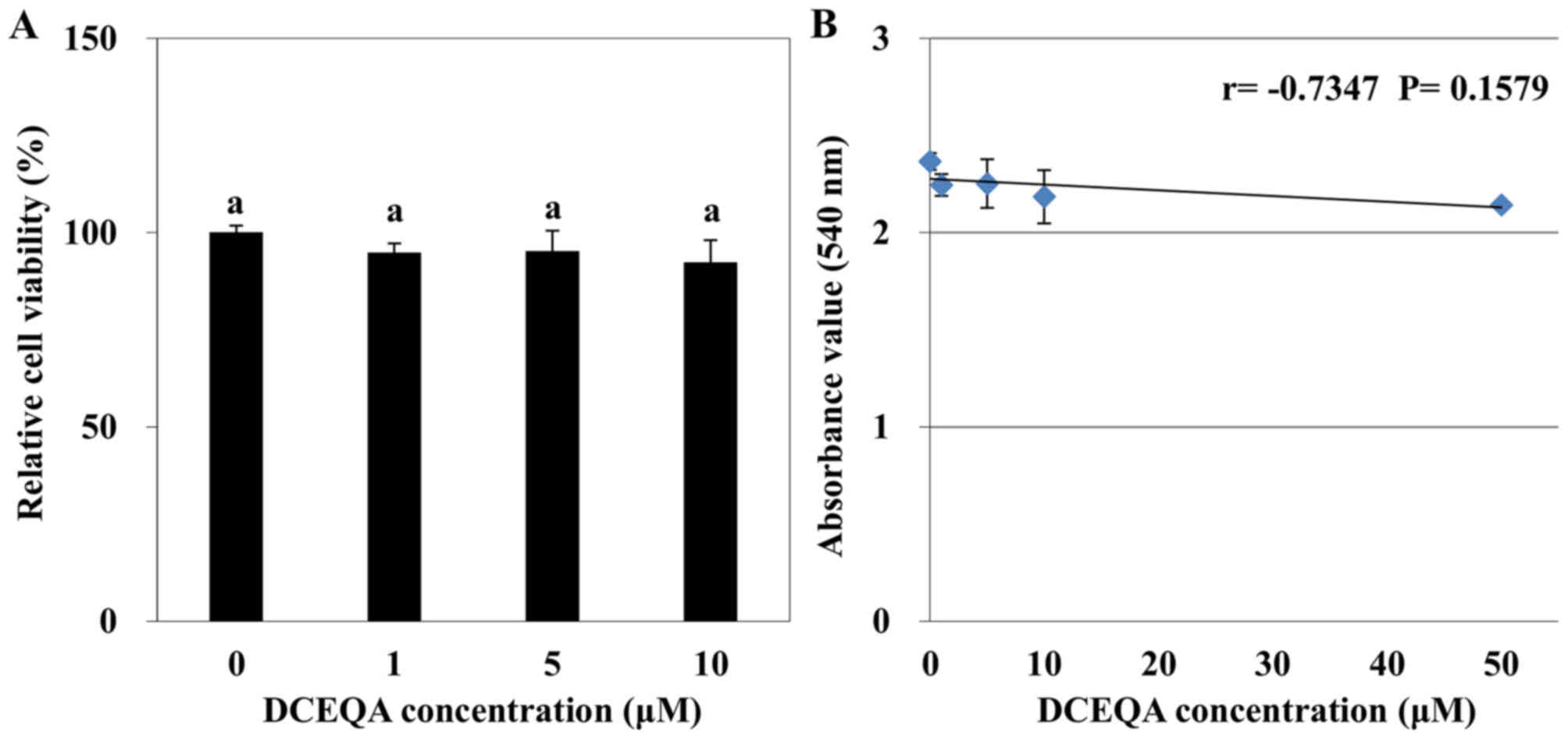
cytotoxicity in HaCaT cells compared to the control group (Fig. 1A). A negative correlation was found
between the DCEQA dosage and viability of the HaCaT keratinocytes
with a LC50 value of 372.53 µM. Although Pearson
correlation analysis revealed that there was a moderate negative
correlation (r=−0.7347), among the tested concentrations (0 to 10
µM) the decrease in cell viability was not statistically
significant (P=0.1579; Fig. 1B).
The first statistically significant decline in cell viability was
observed for concentrations >10 µM. Hence, subsequent assays
were carried out using concentrations of DCEQA at 1, 5 and 10 µM.
The sufficient energy level of UVB irradiation was determined
through the cell viability of HaCaT cells obtained from the MTT
assay following exposure to different energy levels of UVB from 10
to 1,000 mJ/cm2 for different time periods. After 24 h
of UVB exposure at 15 mJ/cm2 a rapid decline in cell
viability was observed (data not shown). Therefore, this energy
level was chosen as the appropriate level for further
experiments.
Effect of DCEQA on UVB-induced
intracellular ROS generation in HaCaT cells
Formation of intracellular ROS was evaluated using
the colorimetric analysis of ROS generation-sensitive dye
2′,7′-dichlorodihydrofluorescin diacetate (DCFH-DA). Evaluation of
the potential scavenging ability of DCEQA on intracellular ROS
generation was carried out by measuring the intensity of DCFH-DA
conversion to highly fluorescent 2′,7′-dichlorofluorescein (DCF) in
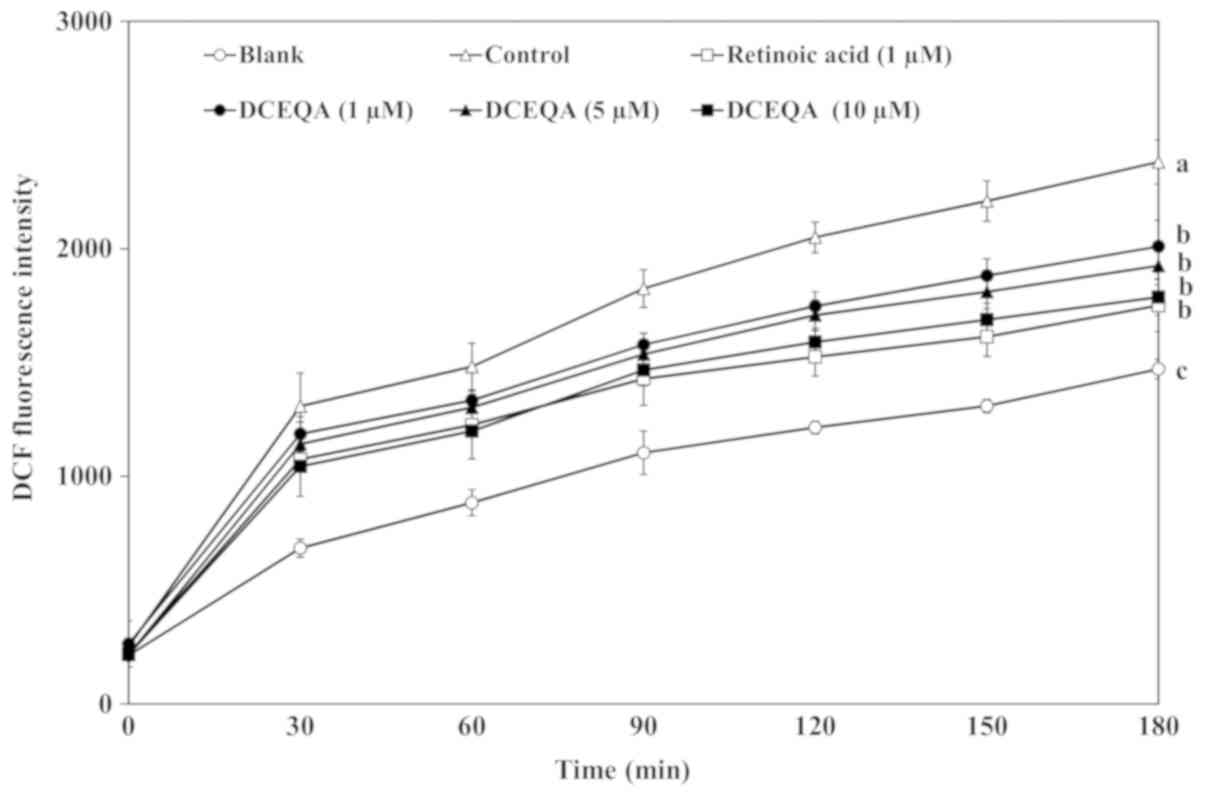
the presence of ROS. UVB irradiation caused a sharp time-dependent
increase in the DCF intensity indicating elevated oxidative stress.
Treatment with DCEQA notably decreased the formation of DCF in a
dose-dependent manner comparable to that of retinoic acid (1 µM),
which was used as a positive control, at the concentration of 10 µM
(Fig. 2). Results showed that
DCEQA was able to reduce the generation of ROS either through its
own ROS scavenging ability or by enhancing the intracellular
scavenging mechanisms. Nevertheless, DCEQA significantly reduced
the intracellular oxidative stress in HaCaT cells.
Effect of DCEQA on UVB-induced
activation of the inflammatory response
As an attempt to determine the effects of DCEQA on
UVB-induced expression of proinflammatory cytokines, both mRNA and
protein expression levels of TNF-α, COX-2, IL-6 and IL-1β were
investigated by RT-sqPCR and western blotting, respectively. The
exposure of HaCaT cells to UVB irradiation resulted in elevated
expression levels of all analyzed cytokines. UVB-induced mRNA
(Fig. 3A) and protein (Fig. 3B) expression levels of TNF-α,
COX-2, IL-6 and IL-1β were lower in the DCEQA-treated keratinocytes
than these levels in the UVB-irradiated control cells. All
cytokines were significantly (P<0.05) reduced in terms of both
mRNA and protein levels as compared to the untreated irradiated
control cells according to quantification of RT-sqPCR (Fig. 4A) and western blot (Fig. 4C) bands. The present results showed
that DCEQA can attenuate the UVB-linked activation of the
proinflammatory response.
 | Figure 3.Effect of DCEQA on the UVB-induced
expression of proinflammatory cytokines (TNF-α, COX-2, IL-6 and
IL-1β), antioxidant enzymes (SOD-1 and HO-1) and Nrf2. (A) Effect
of DCEQA on the mRNA levels of the cytokines, enzymes and Nrf2 was
analyzed by RT-PCR. (B) Effect of DCEQA on the protein levels of
the cytokines, enzymes and Nrf2 was analyzed by western blotting.
DCEQA, 3,5-dicaffeoyl-epi-quinic acid; TNF-α, tumor necrosis
factor-α; IL, interleukin; SOD-1, superoxide dismutase-1; HO-1,
heme oxygenase-1; Nrf2, nuclear factor-like 2. |
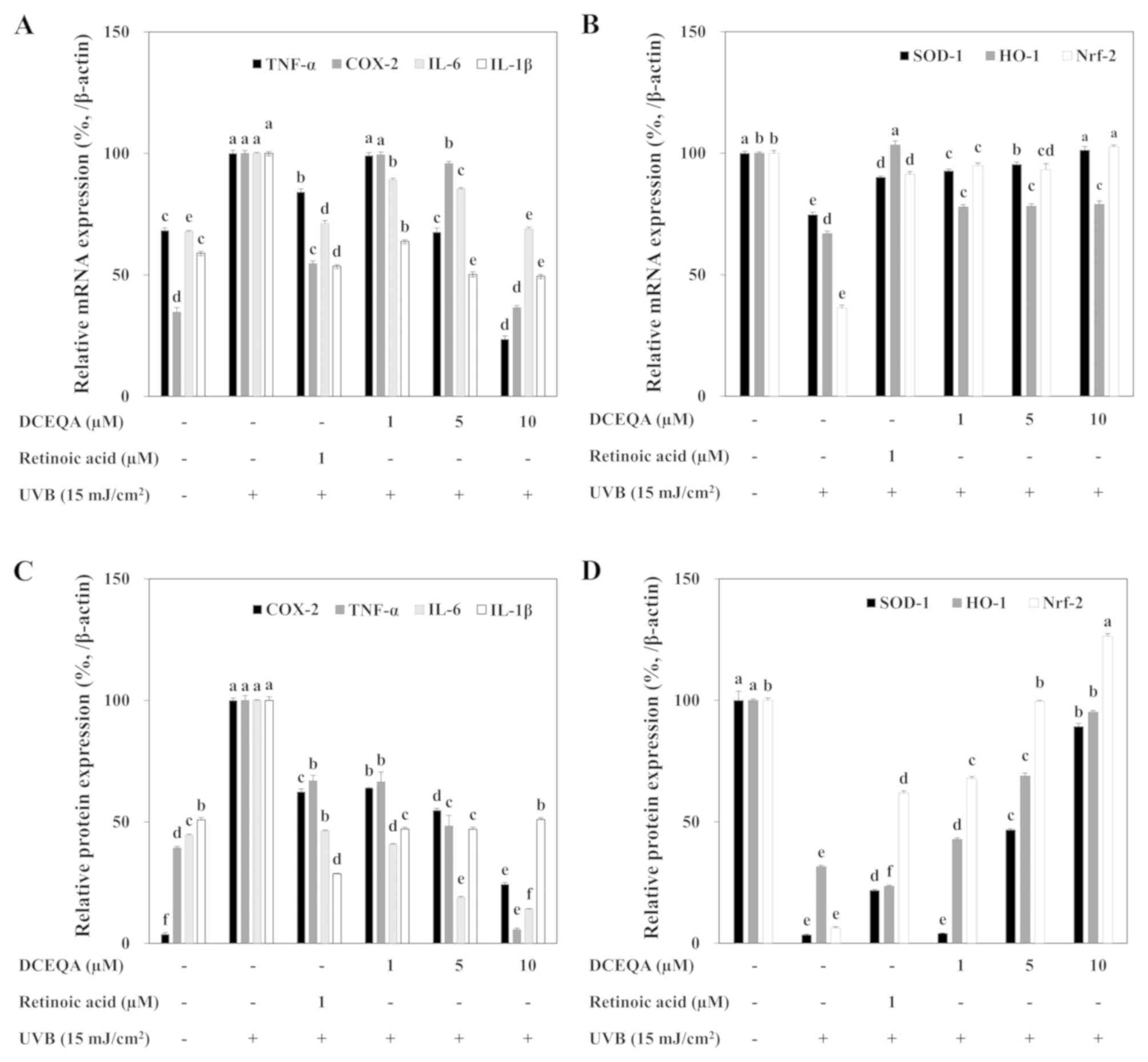 | Figure 4.Quantification of the effect of DCEQA
on the UVB-induced expression of proinflammatory cytokines (TNF-α,
COX-2, IL-6 and IL-1β), antioxidant enzymes (SOD-1 and HO-1) and
Nrf2. The mRNA (A and C) and protein (B and D) levels were
quantified by the density of the bands and normalized against the
housekeeping gene and protein β-actin. Quantification of the
expression levels was carried out through densitometric calculation
and the data were normalized against housekeeping β-actin mRNA and
protein. Effect of DCEQA treatment was plotted as the relative
percentage to UVB-exposed untreated control cells (A and C) or
untreated unirradiated blank cells (C and D). Values are expressed
as the means ± SD (n=3). a-fDifferent letters above the
bars indicate statistically significant differences (P<0.05),
while identical letters indicate no significant differences. DCEQA,
3,5-dicaffeoyl-epi-quinic acid; TNF-α, tumor necrosis factor-α; IL,
interleukin; SOD-1, superoxide dismutase-1; HO-1, heme oxygenase-1;
Nrf2, nuclear factor-like 2. |
Effect of DCEQA on UVB-induced
antioxidant enzyme expression through the nuclear factor-erythroid
2-related factor-2 (Nrf2) pathway
To investigate the effect of DCEQA on UVB-induced
antioxidant enzyme expression, RT-sqPCR and western blotting were
utilized to observe the mRNA and protein levels respectively for
SOD-1 and HO-1. Following UVB exposure, HaCaT cells were shown to
express diminished mRNA and protein levels of SOD-1 and HO-1. Cells
treated with DCEQA exhibited significantly increased expression of
SOD-1 and HO-1 in a dose-dependent manner (Fig. 3). Expression levels of both
antioxidant enzymes were dose-dependently regulated by DCEQA to the
levels prior to UVB irradiation according to the quantification of
the bands (Fig. 4B and D).
In order to investigate the mechanism of action
behind antioxidant enzyme upregulation, the effect of DCEQA on Nrf2
mRNA and protein levels were investigated with RT-sqPCR and western
blotting, respectively (Fig. 3A and
B, respectively). The expression of Nrf2 was significantly
inhibited after UVB exposure. The presence of DCEQA resulted in
activation of the Nrf2 pathway depicted as elevated mRNA (Fig. 4B) and protein (Fig. 4D) expression. This result suggests
that DCEQA upregulated the antioxidant response of HaCaT cells
through Nrf2-dependent activation of SOD-1 and HO-1 production.
Discussion
Oxidative stress plays a critical role in the
progression of photoaging (31).
Exposure to ultraviolet (UV) rays causes elevated oxidative stress
in the cells forming the skin layers. UV-induced generation of
intracellular ROS leads to upregulation of lipid peroxidation and
collagen degradation, which are known causes of DNA damage and
aging of the skin (7). Therefore,
relieving the oxidative stress of skin cells is one of the main
strategies for the prevention of skin photoaging. Use of natural
antioxidants with their biocompatibility and additional health
benefits is a favorable approach in light of recent studies
(32–34). Accordingly, several studies
reported the antioxidant properties of CQA derivatives from
different sources. In the present study, DCEQA significantly
inhibited the UVB-induced intracellular ROS generation and
proinflammatory response (Fig.
2).
UVB exposure is shown to lead to activation of
mitogen activated protein kinase (MAPK) pathways due to increased
ROS levels (13). This harmful
increase in ROS levels activates the inflammatory response due to
possible tissue damage. Coupled with elevated ROS, activation of
the MAPK pathways leads to the upregulation of MMP production and
degradation of skin collagen, which gives the strength and
resiliency to the skin. Exposure to UVB stimulates the production
of proinflammatory cytokines as a component of cellular damage such
as TNF-α, COX-2, IL-6 and IL-1β, which accelerates the photoaging
process. The inflammatory response associated with UVB irradiation
and mediated by these cytokines gradually increases ROS and other
cytokine production augmenting the harmful effects of UV exposure
(7). As showed by the suppressive
effect of DCEQA on ROS levels, it was suggested that DCEQA also
attenuates the inflammation of skin through these cytokines. As
shown in Fig. 4A and B, DCEQA
successfully downregulated the production of proinflammatory TNF-α,
COX-2, IL-6 and IL-1β cytokines. This demonstrates that DCEQA
prevents photoaging by inhibiting inflammation and therefore
reducing skin damage.
Unscavenged free radicals in aging keratinocytes
have been shown to gradually reduce antioxidant enzyme production,
which further amplifies the photoaging effects of UVB irradiation
(35). Several studies have
reported that antioxidants also relieve the suppression of
antioxidant enzyme production pathways (36). It is hence probable that DCEQA can
affect intracellular antioxidant enzyme expression as DCEQA was
shown to possess antioxidant properties. The results showed that
UVB exposure caused significant suppression in the mRNA and protein
expression of SOD-1 and HO-1 antioxidant enzymes which was
upregulated following DCEQA treatment. During the regulation of
antioxidant enzyme production and in turn reducing the oxidative
stress and photoaging process, the Nrf2 pathway plays a pivotal
role (37). Production of oxidant
metabolizing enzymes can neutralize the ROS damage in
keratinocytes. Different CQA derivatives have been reported as
antioxidants that can also regulate the Nrf2/Keap1 pathway while
acting against oxidative stress-mediated cellular damage (38). In the present study, in addition to
reducing intracellular ROS generation, DCEQA increased the
expression of Nrf2 mRNA and protein.
DCEQA was tested in comparison to retinoic acid (1
µM) as a positive control. Retinoic acid is evidently a strong
antioxidant with skin protective effects against UV exposure. At
the same concentration of 1 µM, retinoic acid was more effective
than DCEQA. Yet, the results were notable, and the additional
health benefits of DCEQA as indicated by several previous studies
(39–41) suggest that the efficacy of DCEQA is
comparable to retinoic acid and similar protective agents.
According to the results, the effective dose of DCEQA to be
utilized was suggested to be between approximately 10 µM and up to
but below 50 µM taking into account the possible cytotoxicity. As a
derivative of caffeoylquinic acid, it was suggested that DCEQA
exhibits its photoaging activity due to caffeoyl groups bound to
quinic acids, which are important for CQA-based antioxidant
activity. In addition, the cytoprotective activity of DCEQA can be
credited to the caffeoyl moieties and the position of its
cyclohexane skeleton when compared to similar CQA derivatives
(42).
Upregulation of the Nrf2 pathways, which was
observed to be inhibited by UVB irradiation was suggested to be the
mechanism of action for DCEQA. One of the activation pathways of
Nrf2 is the TNF signaling cascade including the pro-inflammatory
response mediated by TNF-α. Thus, it was suggested that upon DCEQA
treatment, by attenuating the deteriorated Nrf2 pathway,
antioxidant enzyme production was increased, and intracellular ROS
generation was suppressed resulting in diminished inflammation.
This resulted in a reduction in the UVB-linked damage in
keratinocytes and halting of photoaging.
In conclusion, the present study demonstrated the
protective effect of DCEQA against UVB irradiation-mediated damage
in HaCaT keratinocytes, suggestively via ROS scavenging and
increased antioxidant enzyme production through the Nrf2 pathway.
Furthermore, DCEQA was able to downregulate proinflammatory
cytokine production. Taken together, the present results
demonstrated that DCEQA is a potential natural product to be
utilized as a lead compound in the cosmetics field due to its
anti-photoaging properties. However, future studies regarding the
underlying mechanisms and potential scavenging effects on other
free radicals will further enable the use of DCEQA in respective
fields as a potent bioactive substance for skin protection.
Acknowledgements
Not applicable.
Funding
The present study was supported by the BB21+ Project
in 2018.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YS and CSK conceived the research idea, designed the
experiments and supplied the necessary materials. JHO performed the
experiments and collected the data. JIL conducted the isolation and
chemical elucidation analysis. FK interpreted the data and drafted
the manuscript. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Fisher GJ, Kang S, Varani J, Bata-Csorgo
Z, Wan Y, Datta S and Voorhees JJ: Mechanisms of photoaging and
chronological skin aging. Arch Dermatol. 138:1462–1470. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rittié L and Fisher GJ: UV-light-induced
signal cascades and skin aging. Ageing Res Rev. 1:705–720. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fisher GJ: The pathophysiology of
photoaging of the skin. Cutis 75 (2 Suppl). S5–S9. 2005.
|
|
4
|
Ichihashi M, Ueda M, Budiyanto A, Bito T,
Oka M, Fukunaga M, Tsuru K and Horikawa T: UV-induced skin damage.
Toxicology. 189:21–39. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Gruijl FR: Photocarcinogenesis: UVA vs.
UVB radiation. Skin Pharmacol Appl Skin Physiol. 15:316–320. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Simon MM, Aragane Y, Schwarz A, Luger TA
and Schwarz T: UVB light induces nuclear factor kappa B (NF kappa
B) activity independently from chromosomal DNA damage in cell-free
cytosolic extracts. J Invest Dermatol. 102:422–427. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pillai S, Oresajo C and Hayward J:
Ultraviolet radiation and skin aging: roles of reactive oxygen
species, inflammation and protease activation, and strategies for
prevention of inflammation-induced matrix degradation-a review. Int
J Cosmet Sci. 27:17–34. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Herrling T, Jung K and Fuchs J:
Measurements of UV-generated free radicals/reactive oxygen species
(ROS) in skin. Spectrochim Acta A Mol Biomol Spectrosc. 63:840–845.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yoshizumi M, Nakamura T, Kato M, Ishioka
T, Kozawa K, Wakamatsu K and Kimura H: Release of
cytokines/chemokines and cell death in UVB-irradiated human
keratinocytes, HaCaT. Cell Biol Int. 32:1405–1411. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang WS and Stockwell BR: Ferroptosis:
Death by lipid peroxidation. Trends Cell Biol. 26:165–176. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Briganti S and Picardo M: Antioxidant
activity, lipid peroxidation and skin diseases. What's new. J Eur
Acad Dermatol Venereol. 17:663–669. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Buckman SY, Gresham A, Hale P, Hruza G,
Anast J, Masferrer J and Pentland AP: COX-2 expression is induced
by UVB exposure in human skin: Implications for the development of
skin cancer. Carcinogenesis. 19:723–729. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim AL, Labasi JM, Zhu Y, Tang X, McClure
K, Gabel CA, Athar M and Bickers DR: Role of p38 MAPK in
UVB-induced inflammatory responses in the skin of SKH-1 hairless
mice. J Invest Dermatol. 124:1318–1325. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Suschek CV, Mahotka C, Schnorr O and
Kolb-Bachofen V: UVB radiation-mediated expression of inducible
nitric oxide synthase activity and the augmenting role of
co-induced TNF-alpha in human skin endothelial cells. J Invest
Dermatol. 123:950–957. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bashir MM, Sharma MR and Werth VP: UVB and
proinflammatory cytokines synergistically activate TNF-alpha
production in keratinocytes through enhanced gene transcription. J
Invest Dermatol. 129:994–1001. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bosch R, Philips N, Suárez-Pérez JA,
Juarranz A, Devmurari A, Chalensouk-Khaosaat J and González S:
Mechanisms of photoaging and cutaneous photocarcinogenesis, and
photoprotective strategies with phytochemicals. Antioxidants
(Basel). 4:248–268. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin TK, Zhong L and Santiago JL:
Anti-inflammatory and skin barrier repair effects of topical
application of some plant oils. Int J Mol Sci. 19(pii): E702017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu PY, Huang CC, Chu Y, Huang YH, Lin P,
Liu YH, Wen KC, Lin CY, Hsu MC and Chiang HM: Alleviation of
ultraviolet B-induced photodamage by coffea arabica extract in
human skin fibroblasts and hairless mouse skin. Int J Mol Sci.
18(pii): E7822017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
De Luca C, Mikhal'chik EV, Suprun MV,
Papacharalambous M, Truhanov AI and Korkina LG: Skin antiageing and
systemic redox effects of supplementation with marine collagen
peptides and plant-derived antioxidants: A single-blind
case-control clinical study. Oxid Med Cell Longev.
2016:43894102016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Działo M, Mierziak J, Korzun U, Preisner
M, Szopa J and Kulma A: The potential of plant phenolics in
prevention and therapy of skin disorders. Int J Mol Sci.
17:1602016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Katiyar SK: Dietary proanthocyanidins
inhibit UV radiation-induced skin tumor development through
functional activation of the immune system. Mol Nutr Food Res.
60:1374–1382. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang XW, Bai JP, Zhang Q, Hu XL, Tian X,
Zhu J, Liu J, Meng WH and Zhao QC: Caffeoylquinic acid derivatives
protect SH-SY5Y neuroblastoma cells from hydrogen peroxide-induced
injury through modulating oxidative status. Cell Mol Neurobiol.
37:499–509. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tian Y, Puganen A, Alakomi HL, Uusitupa A,
Saarela M and Yang B: Antioxidative and antibacterial activities of
aqueous ethanol extracts of berries, leaves, and branches of berry
plants. Food Res Int. 106:291–303. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Roleira FM, Tavares-da-Silva EJ, Varela
CL, Costa SC, Silva T, Garrido J and Borges F: Plant derived and
dietary phenolic antioxidants: Anticancer properties. Food Chem.
183:235–258. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liang N and Kitts DD: Role of chlorogenic
acids in controlling oxidative and inflammatory stress conditions.
Nutrients. 8(pii): E162015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kwak CS, Yang J, Shin CY and Chung JH:
Topical or oral treatment of peach flower extract attenuates
UV-induced epidermal thickening, matrix metalloproteinase-13
expression and pro-inflammatory cytokine production in hairless
mice skin. Nutr Res Pract. 12:29–40. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Choi HS, Park ED, Park Y and Suh HJ: Spent
coffee ground extract suppresses ultraviolet B-induced photoaging
in hairless mice. J Photochem Photobiol B. 153:164–172. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim HJ and Lee YS: Identification of new
dicaffeoylquinic acids from Chrysanthemum morifolium and their
antioxidant activities. Planta Med. 71:871–876. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Miller SA, Coelho SG, Miller SW, Yamaguchi
Y, Hearing VJ and Beer JZ: Evidence for a new paradigm for
ultraviolet exposure: A universal schedule that is skin phototype
independent. Photodermatol Photoimmunol Photomed. 28:187–195. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Oh JH, Seo Y and Kong CS: Anti-photoaging
effects of solvent-partitioned fractions from Portulaca
oleracea L. on UVB-stressed human keratinocytes. J Food
Biochem. 43:25–Feb;2019.doi.org/10.1111/jfbc.12814. View Article : Google Scholar
|
|
31
|
Wenk J, Brenneisen P, Meewes C, Wlaschek
M, Peters T, Blaudschun R, Ma W, Kuhr L, Schneider L and
Scharffetter-Kochanek K: UV-induced oxidative stress and
photoaging. In: Oxidants and antioxidants in cutaneous biology.
Curr Probl Dermatol. 29:83–94. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Saewan N and Jimtaisong A: Natural
products as photoprotection. J Cosmet Dermatol. 14:47–63. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pallela R, Na-Young Y and Kim SK:
Anti-photoaging and photoprotective compounds derived from marine
organisms. Mar Drugs. 8:1189–1202. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mukherjee PK, Maity N, Nema NK and Sarkar
BK: Bioactive compounds from natural resources against skin aging.
Phytomedicine. 19:64–73. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rabe JH, Mamelak AJ, McElgunn PJ, Morison
WL and Sauder DN: Photoaging: Mechanisms and repair. J Am Acad
Dermatol. 55:1–19. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schmidt HH, Stocker R, Vollbracht C,
Paulsen G, Riley D, Daiber A and Cuadrado A: Antioxidants in
translational medicine. Antioxid Redox Signal. 23:1130–1143. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rojo de la Vega M, Krajisnik A, Zhang DD
and Wondrak GT: Targeting NRF2 for improved skin barrier function
and photoprotection: Focus on the achiote-derived apocarotenoid
bixin. Nutrients. 9(pii): E13712017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Saito Y, Tsuruma K, Ichihara K, Shimazawa
M and Hara H: Brazilian green propolis water extract up-regulates
the early expression level of HO-1 and accelerates Nrf2 after UVA
irradiation. BMC Complement Altern Med. 15:4212015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Oh JH, Lee JI, Karadeniz F, Seo Y and Kong
CS: 3,5-dicaffeoyl-epi-quinic acid isolated from edible halophyte
Atriplex gmelinii inhibits adipogenesis via AMPK/MAPK
pathway in 3T3-L1 adipocytes. Evid Based Complement Alternat Med.
2018:85725712018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ryu KJ, Yoou MS, Seo Y, Yoon KW, Kim HM
and Jeong HJ: Therapeutic effects of Artemisia scoparia Waldst. et
Kitaib in a murine model of atopic dermatitis. Clin Exp Dermatol.
43:798–805. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee JY, Song DG, Lee EH, Jung SH, Nho CW,
Cha KH and Pan CH: Inhibitory effects of
3,5-O-dicaffeoyl-epi-quinic acid from Gymnaster koraiensis
on AKR1B10. J Korean Soc Appl Biol Chem. 52:731–734. 2009.
View Article : Google Scholar
|
|
42
|
Miyamae Y, Kurisu M, Han J, Isoda H and
Shigemori H: Structure-activity relationship of caffeoylquinic
acids on the accelerating activity on ATP production. Chem Pharm
Bull (Tokyo). 59:502–507. 2011. View Article : Google Scholar : PubMed/NCBI
|