Introduction
Sjögren's syndrome (SS), a common chronic systemic
autoimmune disease with mononuclear infiltration in exocrine
glands, is typically characterized by dry mouth and eyes (1–3).
Patients suffer from lack of effective treatment for the complex
pathogenesis of SS (4). High
levels of pro-inflammatory cytokines, such as interleukin (IL)-12,
IL-6, interferon (IFN)α and IFNγ, tumor necrosis factor (TNF)-α,
serum autoantibodies, including antinuclear antibodies, antibodies
against Ro/SSA and La/SSB, rheumatoid factor (RF) and serious
infiltration of T and B cells are considered as the key factors
leading to exocrine gland dysfunction (5–7).
Targeting of the disordered cytokine network has shown promise for
SS treatment (8).
Interleukin (IL)-12, consisting of two subunits,
namely IL-12p40 and IL-12p35, has a powerful impact on the T cell
responses in inflammation (9).
IL-12 has been recognized to be produced by inflammatory myeloid
cells and is involved in various autoimmune inflammatory conditions
by linking innate and adaptive immunity (10). Studies have shown that IL-12 is
increased in the salivary glands of SS patients and MRL-lpr
mice during the course of murine SS (11–13),
and IL-12-transgenic mice develop SS-like symptoms (14). Currently, biologics that block
IL-12 and its family member IL-23, considered to be effective
therapeutic targets, have been used in clinical testing for a
variety of autoimmune diseases including SS (8,15).
Yet, the effects and mechanisms of IL-12 on the immunity system in
different disorders are far from clear.
Myeloid-derived suppressor cells (MDSCs) were
initially described as a heterogeneous population of immature
myeloid cells with immune-regulating activity (16,17)
and consist of two subsets: Granulocytic MDSCs (G-MDSCs) and
monocytic MDSCs (M-MDSCs) (18).
MDSCs were firstly identified in tumor infiltration and were found
to increase and support tumor growth and development (19,20).
Recent studies have demonstrated that MDSCs also play an important
role in pathological conditions including autoimmune diseases.
Vlachou and colleagues demonstrated that MDSCs showed impaired
expansion and function in NZB/W F1 lupus-prone mice
(21). Our previous studies
demonstrated that MDSCs exacerbate SS by inhibiting Th2 cells
(22) and have a crucial role in
systemic lupus erythematosus (SLE) by regulating the balance of
Th17/Treg cells (23), while the
mechanisms of MDSC accumulation in SS are largely unknown. In
addition, MDSCs were found to aggravate rheumatoid arthritis (RA)
by promoting Th17 cell differentiation (24) and were found to show aberrant
function in experimental inflammatory colitis (25). However, the regulation of MDSCs in
SS warrants further investigation.
Previous studies have demonstrated that IL-12 plays
a pro-inflammatory role in the pathogenesis of SS. IL-12 was
reported to promote the recruitment of MDSCs and impair their
suppressive function (26–28). Therefore, the present study aimed
to ascertain whether IL-12 aggravates SS by modulating the
expansion and function of MDSCs.
In the present study, increased plasma IL-12 was
demonstrated in NOD mice which is one of the most accurate models
in deciphering the pathologic mechanisms of SS (29,30).
In vivo, exogenous IL-12 aggravated SS-like disease by
promoting the expansion of bone marrow (BM) and splenic MDSCs,
while anti-IL-12 showed protective function for SS-like disease by
inhibiting the development of BM and splenic MDSCs in NOD mice. In
addition, IL-12 deficiency decreased the generation of MDSCs. These
results indicated that IL-12 aggravates SS by promoting the
expansion of MDSCs.
Materials and methods
Mice
In total, five female NOD mice (weight, 15–17 g;
age, 3 weeks), 35 female NOD mice (weight, 19–22 g; age, 7 weeks),
five female ICR mice (weight, 22–25 g; age, 3 weeks), five female
C57BL/6 (B6) mice (weight, 22–25 g; age, 3 weeks) and 25 female
Il-12−/− B6 mice (weight, 22–25 g; age, 3 weeks)
were purchased from the Model Animal Research Center of Nanjing
University, and were housed in the animal center of the Affiliated
Drum Tower Hospital of Nanjing University Medical School under
pathogen-free conditions at 22±2°C with a relative humidity of
55±15% under a 12-h light/dark cycle. All animals had free access
to water and food. Il-12−/- B6 mice also known as
IL-12p40 knockout (KO) B6 mice were obtained from The Jackson
Laboratory. All animal experiments conformed to the Regulation of
Animal Care Management of the Ministry of Public Health, China and
were approved by the Ethics Committee of the Medical School of
Nanjing University (Nanjing, China). Mice in each group were
anesthetized by intraperitoneal injection of 5% pentobarbital
sodium (50 mg/kg). Under anesthesia, mice were exsanguinated and
the submandibular glands and spleen were immediately collected.
Non-obese diabetic (NOD) mouse as an appropriate
model of Sjögren's syndrome (SS) has biochemical and immunological
similarities with human SS (31).
Previous studies including ours suggested that 4-week old female
NOD mice showed no lymphocyte infiltration in lacrimal glands and
submandibular glands. However, the infiltrating cells appeared in
the lacrimal glands and submandibular glands in 6-week old mice
(32). Therefore, we selected
4-week-old NOD mice as control mice without SS-like symptoms and
8-week-old NOD mice with SS-like symptoms.
Salivary flow rate
The salivary flow rate of mice was measured as
previously described (33).
Briefly, the mice were anesthetized with 5% pentobarbital sodium
(50 mg/kg) and stimulated with pilocarpine (0.1 mg/kg)
intraperitoneally (i.p.). The saliva was collected for a 15-min
period from the oral cavity by micropipette, and the volume of
saliva was determined gravimetrically.
Histological analysis
For histological analysis, submandibular glands
(SGs) were fixed with 4% paraformaldehyde at 4°C for 24 h, embedded
in paraffin and cut into 3-µm-thick sections. Tissue sections were
stained with hematoxylin and eosin (H&E). Each staining step
was performed at room temperature. Images were captured using a
light microscope (Olympus FSX100; Olympus Corporation;
magnification, ×10).
Flow cytometric analysis
Spleen single-cell suspensions were prepared after
lysing red blood cells. The appropriate number of cells was
pre-incubated with antibodies (eBioscience) in optimal
concentration. For analysis of MDSCs, cells were pre-incubated with
surface marker antibodies, including anti-mouse CD11b-APC and
anti-mouse Gr1-PE and then analyzed on a FACS Calibur flow
cytometer (BD Biosciences, Mountain View, CA, USA).
For analysis of the CD3+IFNγ+
T cells, the cells were incubated with 20 ng/ml phorbol myristate
acetate (PMA) plus 1 µg/ml ionomycin with 5 µg/ml of brefeldin A
(Enzo Life Sciences, Inc., East Farmingdale, NY, USA) at 37°C for 4
h. First, surface CD3 with anti-mouse CD3-APC was stained.
Subsequently, cells were fixed with Cytofix/Cytoperm solution (BD
Pharmingen), and then incubated with anti-mouse IFN-γ-PerCP-Cyanine
5.5 (eBioscience; clone no. XMG1.2; cat. no. 16-7311-81) and
analyzed on a FACSCalibur flow cytometer (BD Biosciences).
MDSCs suppress T cell
proliferation
According to the manufacturer's instructions
(Invitrogen; Thermo Fisher Scientific, Inc.), cells from the spleen
of 12-week old female NOD mice were labeled with CFSE. CFSE-labeled
splenocytes were co-cultured without or with MDSCs at ratios of
2:1, 4:1 and 8:1 in 96-well round-bottom plates and were stimulated
with the anti-CD3/CD28 antibody for 3 days. CFSE-labeled
CD3+ T cells were analyzed using a FACSCalibur flow
cytometer.
IL-12 and anti-IL-12 treatment of the
mice
In the IL-12 treatment experiments, female
11-week-old NOD mice were injected with 40 µg/kg IL-12 (BioLegend)
once a day for a total of 7 times or with the same volume of
PBS.
In the anti-IL-12 treatment experiments, female
11-week-old NOD mice were injected with 4 mg/kg anti-IL-12
(BioLegend) once intraperitoneally (i.p.) and with commensurable
Rat IgG2a isotype.
ELISA assay
IL-12 consists of two subunits, IL-12p40 and
IL-12p35, to the formation of the biologically active p70 compound.
In our study, plasma IL-12p70 was assessed by a standard mouse
sandwich ELISA kit (R&D Systems) according to the
manufacturer's instructions.
Statistical analysis
The results of 3 independent experiments are
presented as mean ± SEM. The two experimental groups were analyzed
using Student's t-test. One-way ANOVA followed by the Least
Significant Difference post hoc test were used to account for
multiple comparisons. A P-value <0.05 was considered to indicate
statistical significance.
Results
Increased IL-12 in the Sjögren's
syndrome (SS) mice
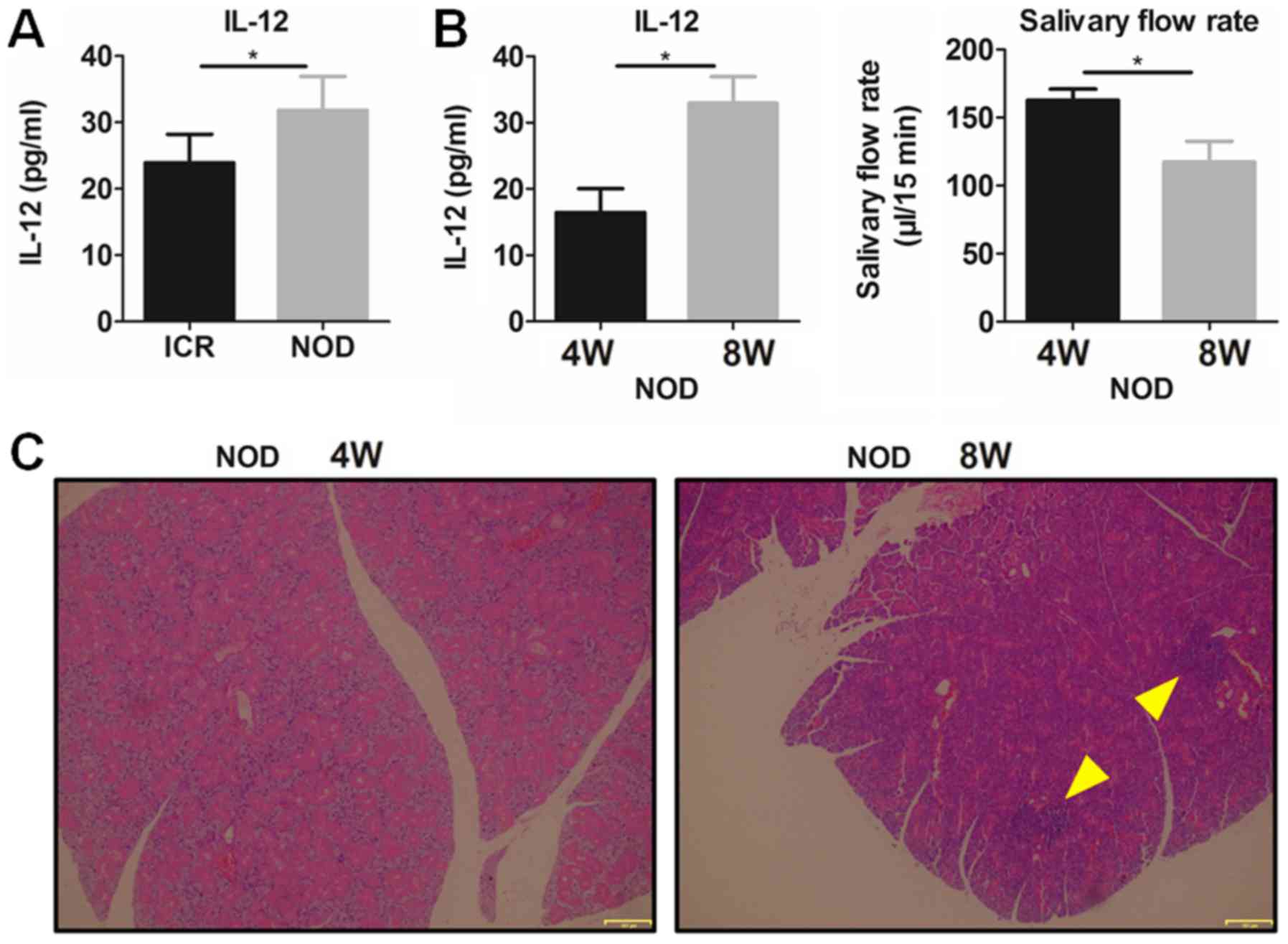
To confirm the role of IL-12 in SS, plasma IL-12 was
detected in the NOD mice and ICR control mice. The results showed
that plasma IL-12 was significantly increased in the NOD mice with
SS-like symptoms comparing with that noted in the ICR mice
(Fig. 1A). It was found that
8-week-old NOD mice had a significantly higher plasma IL-12 and a
significantly lower salivary flow rate (Fig. 1B) and more serious lymphocytic
infiltration in submandibular glands (SG) (Fig. 1C) than these parameters in the
4-week NOD mice, which indicated that SS-like symptoms existed in
the 8-week-old NOD mice but not in the 4-week NOD mice. These
results suggest that increased IL-12 participates in the
pathogenesis of SS-like disease.
IL-12 increases MDSCs and exacerbates
SS in NOD mice
Previous studies have reported that MDSCs are
significantly involved in autoimmune diseases by regulating various
immune responses (34), and IL-12
could modulate the number and function of MDSCs (28). Therefore, we subsequently
determined the relationship between IL-12 and MDSCs in the SS mice.
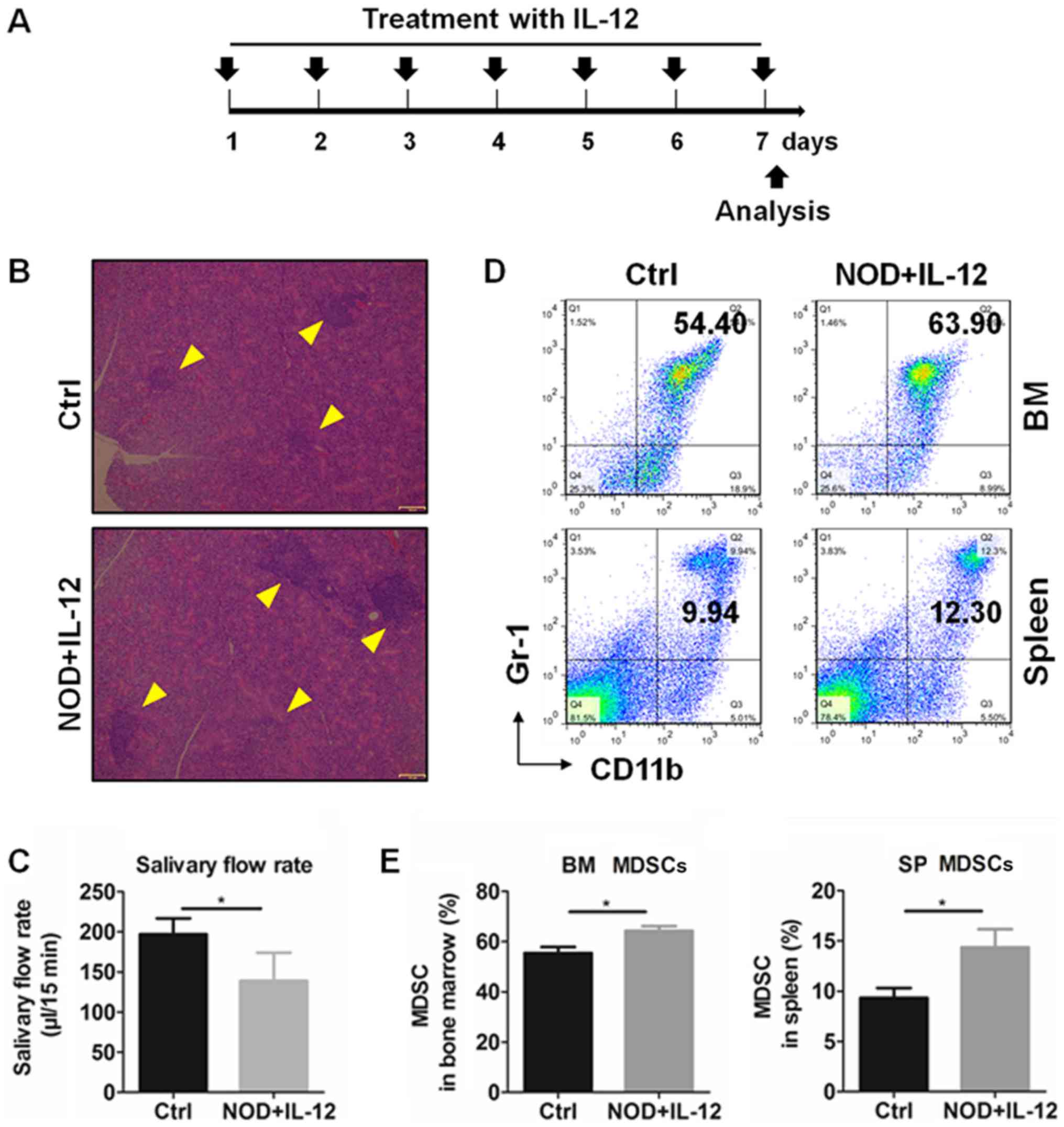
Exogenous IL-12 was injected into 11-week-old NOD mice according to
a treatment schedule (Fig. 2A) and
mice were sacrificed at day 7. According to results of the
histologic and salivary flow rate from mice, IL-12 aggravated the
inflammation in the SGs (Fig. 2B)
and reduced the secretion function of the exocrine glands (Fig. 2C). The percentages of MDSCs in the
bone marrow (BM) and spleen (SP) were detected by flow cytometry
(Fig. 2D) and the results showed
that treatment with IL-12 significantly increased the percentage of
the BM and SP MDSCs (Fig. 2E).
These findings suggest that IL-12 promotes the generation of MDSCs
and exacerbates SS.
Anti-IL-12 decreases the generation of
MDSCs and improves SS in NOD mice
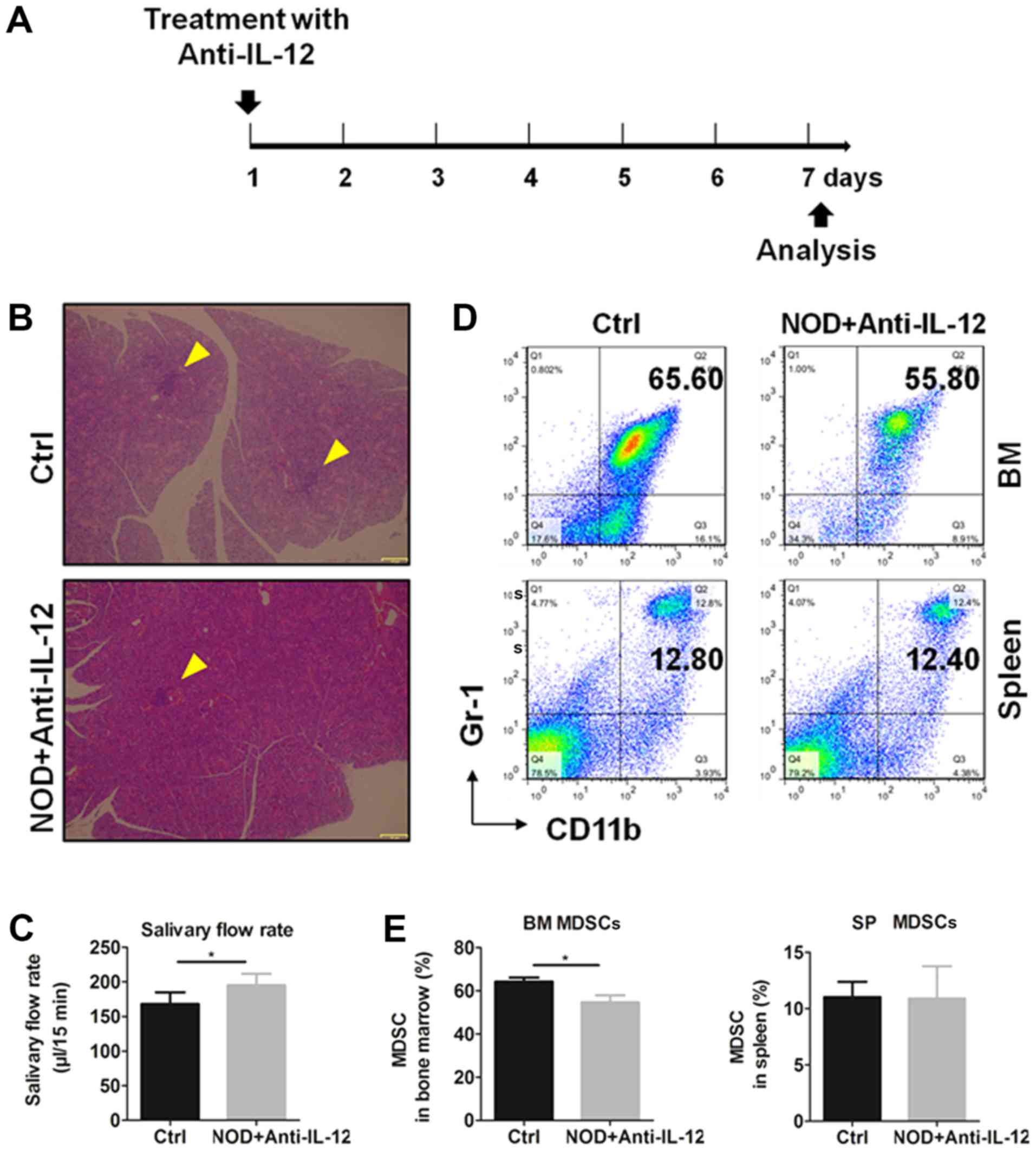
To verify the effects of IL-12 on the generation of
MDSCs, NOD mice were treated i.p. with or without anti-IL-12
antibody, respectively, according to the treatment schedule
(Fig. 3A) and mice were sacrificed
at day 7. Histologic results and salivary flow rate from mice
showed that anti-IL-12 alleviated the inflammation in SGs (Fig. 3B) and partially restored the
secretion function of exocrine glands (Fig. 3C). The percentages of MDSCs in bone
marrow and spleen were detected by flow cytometry (Fig. 3D) and the results showed that
anti-IL-12 significantly increased the percentages of BM and SP
MDSCs (Fig. 3E). These results
showed that anti-IL-12 decreased MDSCs and improved SS-like
syndrome.
IL-12 deficiency impairs the
generation of MDSCs
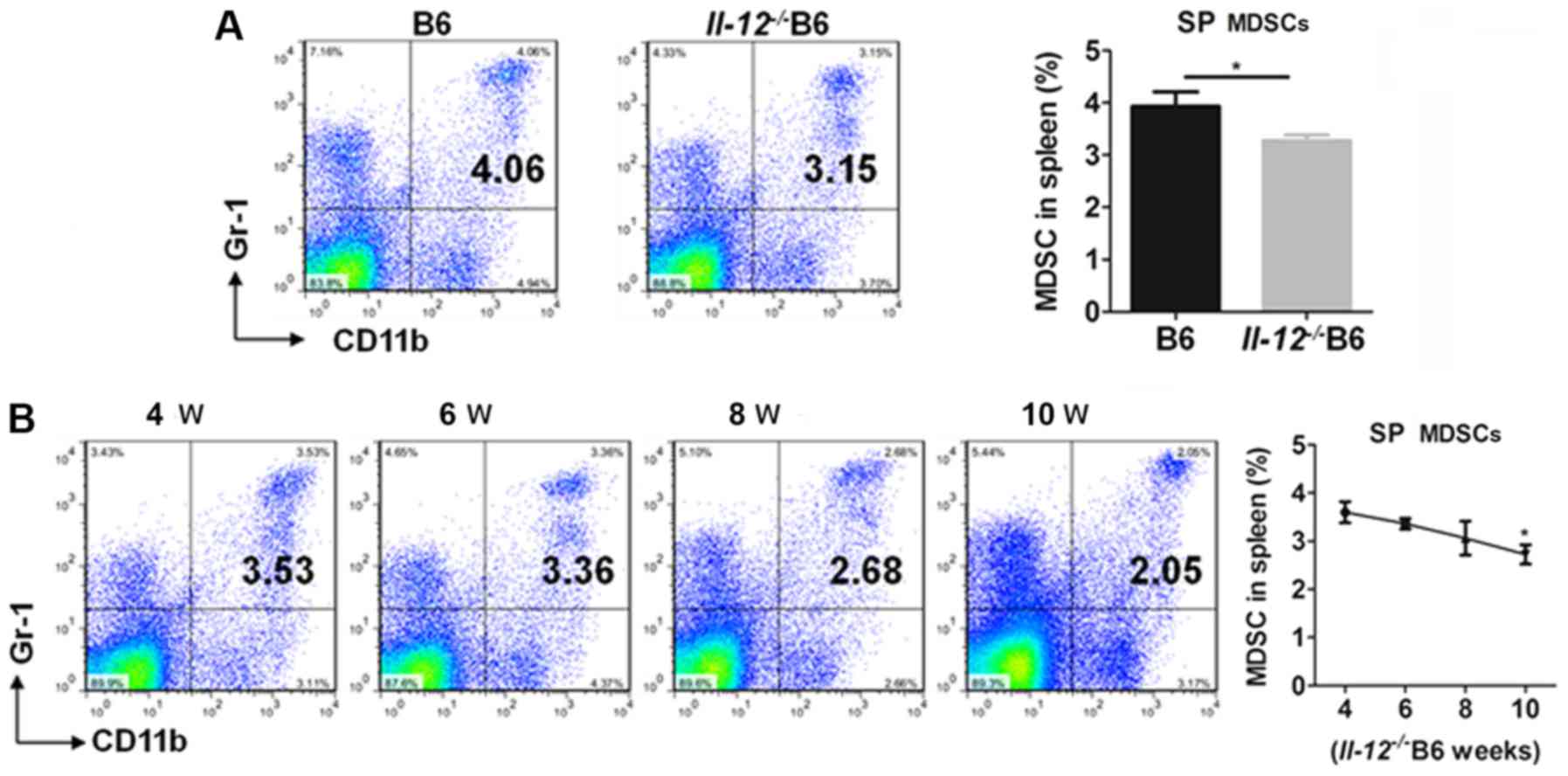
To determine the regulatory effects of IL-12 on
MDSCs, the percentages of MDSCs were detected in IL-12-deficient
C57BL/6 (Il-12−/− B6) mice. Compared with the B6
mice, Il-12−/- B6 mice had a significantly lower
percentage of SP MDSCs (Fig. 4A).
In addition, the percentage of SP MDSCs decreased with age from 4
to 10 weeks in the Il-12−/ B6 mice (Fig. 4B). These findings suggest that
IL-12 induces the generation of MDSCs.
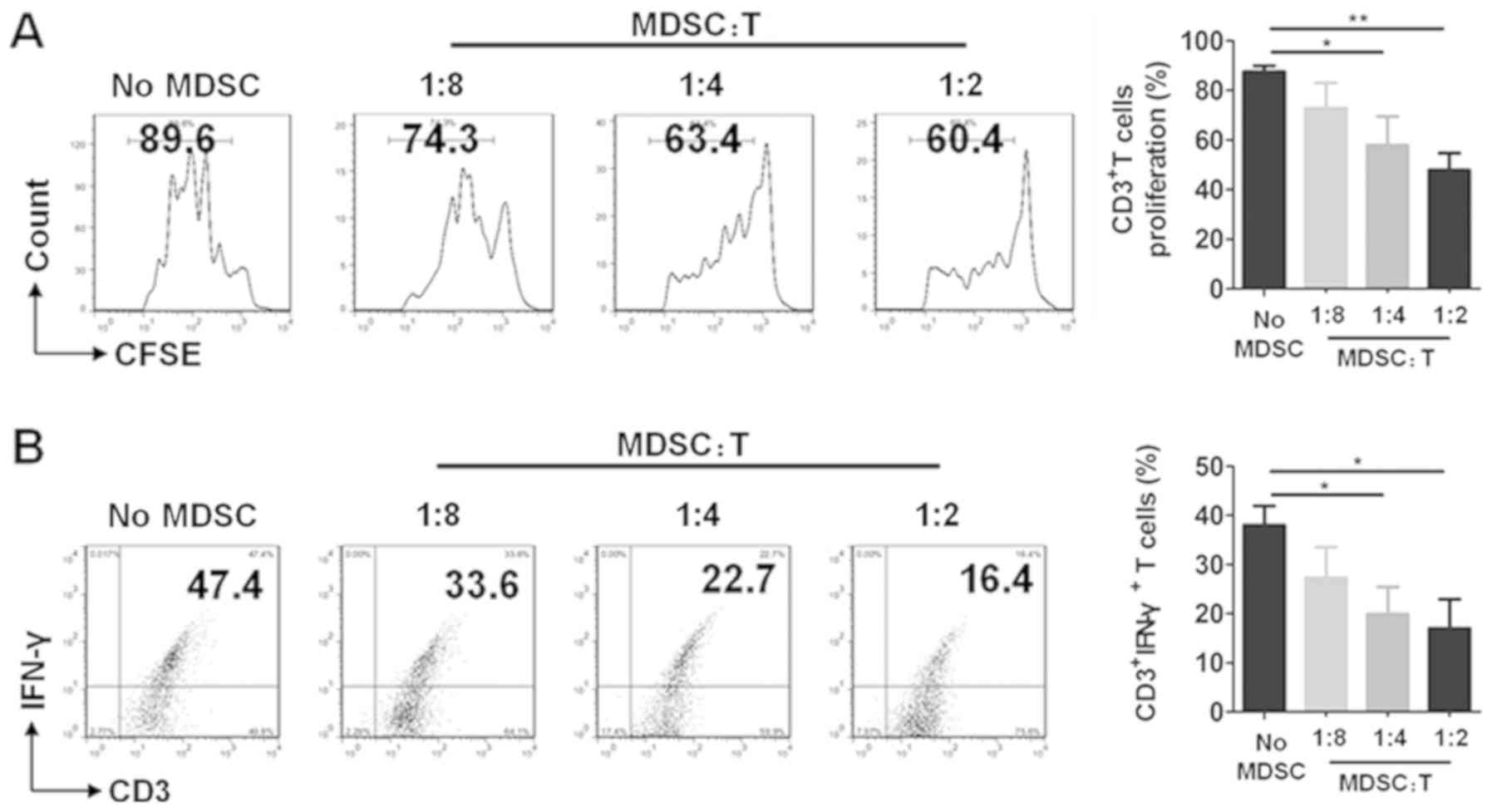
MDSCs display immunosuppressive
activity in SS mice
The immunosuppressive function is an important
characteristic of MDSCs. To confirm that
CD11b+Gr-1+ cells that were determined are
MDSCs in this study, T cells were co-cultured without or with MDSCs
at the different ratios of 8:1, 4:1 and 2:1, respectively. The
results showed that MDSCs had significant suppressive ability on T
cell proliferation (Fig. 5A) and
decreased the percentages of CD3+IFNγ+ T
cells in ratios of 4:1 and 2:1 (Fig.
5B). These results demonstrated that
CD11b+Gr1+ MDSCs exhibited immunosuppressive
activity in our study.
Discussion
In the present study, it was demonstrated that IL-12
and MDSCs are involved in the disease progression of Sjögren's
syndrome (SS). Plasma IL-12 was elevated in NOD mice, and IL-12
enhanced the expansion of MDSCs in the NOD mice. Anti-IL-12
decreased the percentages of MDSCs in the NOD mice, and IL-12
deficiency impaired the generation of MDSCs gradually with age. All
of these results suggest a pathogenic role of IL-12 in SS by
promoting the expansion of MDSCs.
IL-12 is known as an inflammatory cytokine involved
in various autoimmune diseases (35–38).
According to major advances in the last few years, dysregulation of
plasma cytokines has been demonstrated to play a significant role
in contributing to the dysfunction of exocrine glands in SS, and
IL-12 has been considered as one of the main pathogenic factors as
well as IL-17 and IL-23 (8,39).
However, IL-12 has been mainly studied in terms of lymphocyte
responses and the exact mechanisms of IL-12 in the regulation of
myeloid cells in SS are not fully understood.
MDSCs, possessing a prominent capacity to suppress T
cell activation, have been regarded as a potent inducer for tumor
immune escape. The immunosuppressive function is an important
characteristic of MDSCs. T cells were co-cultured with MDSCs and
the results showed that MDSCs had significant suppressive ability
on T cell proliferation and decreased the percentages of
CD3+IFNγ+ T cells. The expression of IFNγ
indicates the activation level of T cells (23,24).
Thus, changes in the CD3+IFNγ+ T cells in T
cells co-cultured with CD11b+Gr-1+ cells
represented the suppressive effects of the
CD11b+Gr-1+ cells, which demonstrated that
the purified CD11b+Gr-1+ cells were MDSCs in
this study. In addition, functional molecules Arg1, iNOS in splenic
G-MDSCs and M-MDSCs were determined by qPCR in our previous study
(23). The abnormal expansion of
MDSCs has also been studied in inflammatory conditions including
SLE mice and lupus patients (23),
collagen-induced arthritis mice and RA patients (24,40),
inflammatory bowel diseases (41)
and experimental autoimmune encephalitis (EAE) (42,43).
The percentages and function of MDSCs, G-MDSCs and M-MDSCs have
been demonstrated to play a significant role in immune disorders.
In the present study, it was demonstrated that, compared to
12-week-old ICR mice, MDSCs were increased significantly in the
bone marrow and spleen of the age-matched NOD mice (Fig. S1). Our previous research
demonstrated that accumulative MDSCs exacerbate SS by inhibiting
the Th2 cell response (22).
However, the cause of MDSCs and these subset accumulations in SS
are largely unknown.
In the present study, our results demonstrated that
IL-12 aggravated SS-like disease, while anti-IL-12 attenuated
SS-like disease in NOD mice. Moreover, increased IL-12 and a higher
percentage of MDSCs were associated with SS pathogenesis. Increased
percentages of MDSCs in IL-12-treated NOD mice, decreased
percentages of MDSCs in anti-IL-12-treated NOD mice and gradually
reduced percentages of MDSCs in Il-12−/- B6 mice
with age suggested that IL-12 and the regulation of MDSCs may be
important in SS pathogenesis. Anti-IL-12 treatment resulted in the
amelioration of SS-like symptoms in the NOD mice accompanied with
an approximately 10% decreased MDSCs in the bone marrow (BM) but
not in the spleen (SP) in the present study. The reason for this
phenomenon may due to the fact that anti-IL-12 mainly ameliorates
SS-like disease in NOD mice by inhibiting the generation of BM
MDSCs and improving the suppressive function of MDSCs, but not by
inhibiting the expansion of SP MDSCs. IL-12 is a critical cytokine
that bridges innate and adaptive immunity and can increase an
antitumor immune response by inducing the production of IFNγ
(44,45). MDSCs were firstly identified as
being involved in tumor infiltration and increased and supported
tumor growth and development (19,20).
Studies found that IL-12 promotes the recruitment of MDSCs and
impairs their suppressive function in tumors and infection
(26–28). In our previous study, the number of
MDSCs were found to be increased in SS and MDSC transfer
exacerbated the SS-like symptoms in NOD mice (22). In the present study, it was found
that IL-12 upregulated BM and SP MDSCs and aggravated SS in NOD
mice. The mechanisms of MDSCs involved in SS remain largely
unknown. Our findings indicated that the number of MDSCs was
increased following IL-12 treatment, while the suppressive function
did not increase accordingly. Therefore, our results suggest that
IL-12 impairs the suppressive function of MDSCs in NOD mice. This
is in line with the previous study that IL-12 can promote the
recruitment of MDSCs and impair their suppressive function. Our
findings indicated that MDSCs under SS conditions are functionally
similar to those associated with tumors and infections in response
to IL-12.
MDSCs consist of two subsets: Granulocytic MDSCs
(G-MDSCs) and monocytic MDSCs (M-MDSCs) (18). The percentages and function of
MDSCs, G-MDSCs and M-MDSCs are important in the pathogenesis of
immune disorders. The changes in G-MDSCs and M-MDSCs were
determined in the present study. It was found that IL-12 and
anti-IL-12 significantly altered the percentages of total MDSCs and
the percentages of G-MDSCs and M-MDSCs slightly in the BM and SP
(Fig. S2). SP G-MDSCs were
significantly decreased in 10-week-old M-MDSCs and were
significantly decreased in the 8-week and 10-week-old
Il-12−/− B6 mice (Fig. S3). Previous studies indicated that
macrophages, dendritic cells (DCs) and regulatory T cells
participate in the pathogenesis of autoimmune diseases, including
SS. The role of regulatory T cells (Treg), as one of the important
immunosuppressive cells, in SS pathogenesis is still controversial
due to its complex pathogeny (46,47).
In the present study, IL-12 and anti-IL-12-treatment showed almost
no effects on changes in Treg cells (Fig. S4A). Exogenous IL-12 slightly
increased the percentage of spleen macrophages, while
anti-IL-12-treatment significantly decreased the percentage of
macrophages in the NOD mice (Fig.
S4B). The contribution of DCs in SS remains to be evaluated.
This is partly due to the low frequency of DCs in the circulation
and inflamed tissues. DCs were not detected in response to IL-12 in
this study. However, previous studies indicated that DCs migrated
from the peripheral blood into the exocrine glands to initiate and
perpetuate autoimmune response (48,49).
In summary, these findings indicted that the changes were not
specific to MDSCs in response to IL-12 in SS.
The development and expansion of MDSCs are regulated
by multiple factors which is a complex and gradual phenomenon.
Among the related signaling groups, the inflammatory cytokines and
damage-associated molecular patterns are important (17). Previous studies demonstrated that
IL-12 treatment impaired the suppressive function of MDSCs by
upregulating surface markers and decreasing nitric oxide synthase
and IFNγ mRNA in the tumor microenvironment (28). Furthermore, transfer of
IL-12-producing cells significantly decreased the number of MDSCs
in tumors but not in the spleen of C57BL/6 mice and promoted
G-MDSCs in tumors to secrete IFNγ to induce tumor regression
(27). Moreover, one study found
that pro-inflammatory signals induce MDSC recruitment and both p40
and p35 (two subunits of IL-12) knockout mice showed decreased MDSC
recruitment and increased monocyte and neutrophil influx (26). In the present study increased IL-12
was found to be an important pro-inflammatory cytokine, promoted
MDSC development to accelerate SS progression. However, the exact
mechanisms of IL-12 involved in the promotion of MDSCs and the
inflammatory role of MDSCs in SS warrant further investigation.
In conclusion, collectively, it was revealed that
IL-12 deteriorates SS-like disease by enhancing MDSC expansion.
These results provide new insight into the pathogenetic mechanisms
and elucidate a potential novel therapy for SS.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This research was supported by the National Natural
Science Foundation of China (NSFC) (grant nos. 81571583 and
81770061 to GY; 31872732 to YH).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YH and GY initiated, designed and supervised the
project. JQ and DL carried out the experiments. XZ, YP and TW
contributed to the animal experiments such as breeding procedures
and tissue collection. GS and HD performed the flow cytometric
analysis and conducted the data analyses. All authors were involved
in writing the paper and approved the submitted version.
Ethics approval and consent to
participate
All animal experiments conformed to the Regulation
of Animal Care Management of the Ministry of Public Health, P.R.
China and were approved by the Ethics Committee of the Medical
School of Nanjing University (Nanjing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
References
|
1
|
Soliotis FC and Moutsopoulos HM: Sjogren's
syndrome. Autoimmunity. 37:305–307. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhou Y, Jin L, Kong F, Zhang H, Fang X,
Chen Z, Wang G and Li X and Li X: Clinical and immunological
consequences of total glucosides of paeony treatment in Sjögren's
syndrome: A randomized controlled pilot trial. Int Immunopharmacol.
39:314–319. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mavragani CP and Moutsopoulos HM:
Sjögren's syndrome. Annu Rev Pathol. 9:273–285. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu J, Wang D, Liu D, Fan Z, Zhang H, Liu
O, Ding G, Gao R, Zhang C, Ding Y, et al: Allogeneic mesenchymal
stem cell treatment alleviates experimental and clinical Sjögren
syndrome. Blood. 120:3142–3151. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tzioufas AG, Kapsogeorgou EK and
Moutsopoulos HM: Pathogenesis of Sjögren's syndrome: What we know
and what we should learn. J Autoimmun. 39:4–8. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moriyama M, Hayashida JN, Toyoshima T,
Ohyama Y, Shinozaki S, Tanaka A, Maehara T and Nakamura S:
Cytokine/chemokine profiles contribute to understanding the
pathogenesis and diagnosis of primary Sjögren's syndrome. Clin Exp
Immunol. 169:17–26. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mavragani CP: Mechanisms and new
strategies for primary Sjögren's syndrome. Annu Rev Med.
68:331–343. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Roescher N, Tak PP and Illei GG: Cytokines
in Sjogren's syndrome: Potential therapeutic targets. Ann Rheum
Dis. 69:945–948. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Vignali DA and Kuchroo VK: IL-12 family
cytokines: Immunological playmakers. Nat Immunol. 13:722–728. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kastelein RA, Hunter CA and Cua DJ:
Discovery and biology of IL-23 and IL-27: related but functionally
distinct regulators of inflammation. Annu Rev Immunol. 25:221–242.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bikker A, van Woerkom JM, Kruize AA,
Wenting-van Wijk M, de Jager W, Bijlsma JW, Lafeber FP and van Roon
JA: Increased expression of interleukin-7 in labial salivary glands
of patients with primary Sjögren's syndrome correlates with
increased inflammation. Arthritis Rheum. 62:969–977. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Manoussakis MN, Boiu S, Korkolopoulou P,
Kapsogeorgou EK, Kavantzas N, Ziakas P, Patsouris E and
Moutsopoulos HM: Rates of infiltration by macrophages and dendritic
cells and expression of interleukin-18 and interleukin-12 in the
chronic inflammatory lesions of Sjögren's syndrome: Correlation
with certain features of immune hyperactivity and factors
associated with high risk of lymphoma development. Arthritis Rheum.
56:3977–3988. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hayashi Y, Haneji N and Hamano H: Cytokine
gene expression and autoantibody production in Sjögren's syndrome
of MRL/lpr mice. Autoimmunity. 23:269–277. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vosters JL, Landek-Salgado MA, Yin H,
Swaim WD, Kimura H, Tak PP, Caturegli P and Chiorini JA:
Interleukin-12 induces salivary gland dysfunction in transgenic
mice, providing a new model of Sjögren's syndrome. Arthritis Rheum.
60:3633–3641. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Teng MW, Bowman EP, McElwee JJ, Smyth MJ,
Casanova JL, Cooper AM and Cua DJ: IL-12 and IL-23 cytokines: From
discovery to targeted therapies for immune-mediated inflammatory
diseases. Nat Med. 21:719–729. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gabrilovich DI and Nagaraj S:
Myeloid-derived suppressor cells as regulators of the immune
system. Nat Rev Immunol. 9:162–174. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Veglia F, Perego M and Gabrilovich D:
Myeloid-derived suppressor cells coming of age. Nat Immunol.
19:108–119. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Parker KH, Beury DW and Ostrand-Rosenberg
S: Myeloid-derived suppressor cells: Critical cells driving immune
suppression in the tumor microenvironment. Adv Cancer Res.
128:95–139. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Talmadge JE and Gabrilovich DI: History of
myeloid-derived suppressor cells. Nat Rev Cancer. 13:739–752. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Younos I, Donkor M, Hoke T, Dafferner A,
Samson H, Westphal S and Talmadge J: Tumor- and organ-dependent
infiltration by myeloid-derived suppressor cells. Int
Immunopharmacol. 11:816–826. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Vlachou K, Mintzas K, Glymenaki M, Ioannou
M, Papadaki G, Bertsias GK, Sidiropoulos P, Boumpas DT and Verginis
P: Elimination of granulocytic myeloid-derived suppressor cells in
lupus-prone mice linked to reactive oxygen species-dependent
extracellular trap formation. Arthritis Rheumatol. 68:449–461.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Qi J, Li D, Shi G, Zhang X, Pan Y, Dou H,
Yao G and Hou Y: Myeloid-derived suppressor cells exacerbate
Sjögren's syndrome by inhibiting Th2 immune responses. Mol Immunol.
101:251–258. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ji J, Xu J, Zhao S, Liu F, Qi J, Song Y,
Ren J, Wang T, Dou H and Hou Y: Myeloid-derived suppressor cells
contribute to systemic lupus erythaematosus by regulating
differentiation of Th17 cells and Tregs. Clin Sci (Lond).
130:1453–1467. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Guo C, Hu F, Yi H, Feng Z, Li C, Shi L, Li
Y, Liu H, Yu X, Wang H, et al: Myeloid-derived suppressor cells
have a proinflammatory role in the pathogenesis of autoimmune
arthritis. Ann Rheum Dis. 75:278–285. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kontaki E, Boumpas DT, Tzardi M, Mouzas
IA, Papadakis KA and Verginis P: Aberrant function of
myeloid-derived suppressor cells (MDSCs) in experimental colitis
and in inflammatory bowel disease (IBD) immune responses.
Autoimmunity. 50:170–181. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Heim CE, Vidlak D, Scherr TD, Hartman CW,
Garvin KL and Kielian T: IL-12 promotes myeloid-derived suppressor
cell recruitment and bacterial persistence during Staphylococcus
aureus orthopedic implant infection. J Immunol. 194:3861–3872.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kerkar SP, Goldszmid RS, Muranski P,
Chinnasamy D, Yu Z, Reger RN, Leonardi AJ, Morgan RA, Wang E,
Marincola FM, et al: IL-12 triggers a programmatic change in
dysfunctional myeloid-derived cells within mouse tumors. J Clin
Invest. 121:4746–4757. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Steding CE, Wu ST, Zhang Y, Jeng MH, Elzey
BD and Kao C: The role of interleukin-12 on modulating
myeloid-derived suppressor cells, increasing overall survival and
reducing metastasis. Immunology. 133:221–238. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yoon KC, De Paiva CS, Qi H, Chen Z, Farley
WJ, Li DQ, Stern ME and Pflugfelder SC: Desiccating environmental
stress exacerbates autoimmune lacrimal keratoconjunctivitis in
non-obese diabetic mice. J Autoimmun. 30:212–221. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shah M, Edman MC, Janga SR, Shi P,
Dhandhukia J, Liu S, Louie SG, Rodgers K, Mackay JA and
Hamm-Alvarez SF: A rapamycin-binding protein polymer nanoparticle
shows potent therapeutic activity in suppressing autoimmune
dacryoadenitis in a mouse model of Sjögren's syndrome. J Control
Release. 171:269–279. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cha S, Peck AB and Humphreys-Beher MG:
Progress in understanding autoimmune exocrinopathy using the
non-obese diabetic mouse: An update. Crit Rev Oral Biol Med.
13:5–16. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hunger RE, Carnaud C, Vogt I and Mueller
C: Male gonadal environment paradoxically promotes dacryoadenitis
in nonobese diabetic mice. J Clin Invest. 101:1300–1309. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin X, Rui K, Deng J, Tian J, Wang X, Wang
S, Ko KH, Jiao Z, Chan VS, Lau CS, et al: Th17 cells play a
critical role in the development of experimental Sjögren's
syndrome. Ann Rheum Dis. 74:1302–1310. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Crook KR and Liu P: Role of
myeloid-derived suppressor cells in autoimmune disease. World J
Immunol. 4:26–33. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Trinchieri G: Interleukin-12 and the
regulation of innate resistance and adaptive immunity. Nat Rev
Immunol. 3:133–146. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hagberg N, Joelsson M, Leonard D, Reid S,
Eloranta ML, Mo J, Nilsson MK, Syvänen AC, Bryceson YT and Rönnblom
L: The STAT4 SLE risk allele rs7574865[T] is associated with
increased IL-12-induced IFN-γ production in T cells from patients
with SLE. Ann Rheum Dis. 77:1070–1077. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thompson C, Davies R and Choy E: Anti
cytokine therapy in chronic inflammatory arthritis. Cytokine.
86:92–99. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rostami A and Ciric B: Role of Th17 cells
in the pathogenesis of CNS inflammatory demyelination. J Neurol
Sci. 333:76–87. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nocturne G and Mariette X: Advances in
understanding the pathogenesis of primary Sjögren's syndrome. Nat
Rev Rheumatol. 9:544–556. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang H, Wang S, Huang Y, Wang H, Zhao J,
Gaskin F, Yang N and Fu SM: Myeloid-derived suppressor cells are
proinflammatory and regulate collagen-induced arthritis through
manipulating Th17 cell differentiation. Clin Immunol. 157:175–186.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Haile LA, von Wasielewski R,
Gamrekelashvili J, Krüger C, Bachmann O, Westendorf AM, Buer J,
Liblau R, Manns MP, Korangy F and Greten TF: Myeloid-derived
suppressor cells in inflammatory bowel disease: A new
immunoregulatory pathway. Gastroenterology. 135:871–881, 881.e1-e5.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
King IL, Dickendesher TL and Segal BM:
Circulating Ly-6C+ myeloid precursors migrate to the CNS and play a
pathogenic role during autoimmune demyelinating disease. Blood.
113:3190–3197. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yi H, Guo C, Yu X, Zuo D and Wang XY:
Mouse CD11b+Gr-1+ myeloid cells can promote Th17 cell
differentiation and experimental autoimmune encephalomyelitis. J
Immunol. 189:4295–4304. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Waldner MJ and Neurath MF: Gene therapy
using IL 12 family members in infection, auto immunity, and cancer.
Curr Gene Ther. 9:239–247. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zundler S and Neurath MF: Interleukin-12:
Functional activities and implications for disease. Cytokine Growth
Factor Rev. 26:559–568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Christodoulou MI, Kapsogeorgou EK,
Moutsopoulos NM and Moutsopoulos HM: Foxp3+ T-regulatory cells in
Sjogren's syndrome: Correlation with the grade of the autoimmune
lesion and certain adverse prognostic factors. Am J Pathol.
173:1389–1396. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gottenberg JE, Lavie F, Abbed K, Gasnault
J, Le Nevot E, Delfraissy JF, Taoufik Y and Mariette X: CD4
CD25high regulatory T cells are not impaired in patients with
primary Sjögren's syndrome. J Autoimmun. 24:235–242. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ozaki Y, Ito T, Son Y, Amuro H, Shimamoto
K, Sugimoto H, Katashiba Y, Ogata M, Miyamoto R, Murakami N, et al:
Decrease of blood dendritic cells and increase of
tissue-infiltrating dendritic cells are involved in the induction
of Sjögren's syndrome but not in the maintenance. Clin Exp Immunol.
159:315–326. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
van Blokland SC, van Helden-Meeuwsen CG,
Wierenga-Wolf AF, Drexhage HA, Hooijkaas H, van de Merwe JP and
Versnel MA: Two different types of sialoadenitis in the NOD- and
MRL/lpr mouse models for Sjögren's syndrome: A differential role
for dendritic cells in the initiation of sialoadenitis? Lab Invest.
80:575–585. 2000. View Article : Google Scholar : PubMed/NCBI
|