Introduction
Spinal cord injury (SCI) is among the most
devastating conditions to the health and quality of life of
patients and their families (1,2). SCI
generally causes severe and permanent neurological deficits, is
associated with high healthcare costs, and is becoming a major
clinical concern worldwide (3).
The pathological events of SCI include primary and secondary injury
(4–6). Although both types of injury are
responsible for the progressive neuronal death following trauma,
there is currently no effective medical or surgical treatment for
primary injury, and all efforts are focused on alleviating
secondary injury (6,7).
Although the precise pathogenesis of SCI remains
elusive, multiple mechanisms, such as inflammation, oxidative
stress, immune neurotoxicity, apoptosis and autophagy, have been
implicated (8–10). For example, as the currently
standard treatment for acute SCI, methylprednisolone exerts
neuroprotective effects through its potent anti-inflammatory,
anti-apoptotic and antioxidant properties (11). However, the effectiveness of
methylprednisolone regarding functional improvement after SCI
remains a subject of debate (12);
thus, a number of other agents have been trialed for SCI treatment.
Recently, Tohda and Nagata (12)
confirmed the beneficial effect of icariin, a flavonoid derived
from the Chinese herbal plant Epimedium sagittatum Maxim, on
experimental SCI based on locomotor rating scale scoring. The
underlying protective mechanism, however, remains largely unknown.
In order to achieve a better understanding of the beneficial effect
of icariin on SCI, the present study attempted to determine its
precise underlying mechanism on the basis of previous reports.
Attention was specifically focused on the modulatory effect of
icariin on apoptotic, redox and inflammatory molecules.
Materials and methods
Animals and drug treatment
A total of 24 adult male Sprague-Dawley rats (6
weeks old) weighing 180–220 g were purchased from the Laboratory
Animal Center of Jinzhou Medical University, and were housed in a
temperature-controlled room (23±0.5°C) with 35–55% relative
humidity and a 12/12-h light/dark cycle. The rats were given free
access to food and water throughout the experimental period. All
procedures were approved by the Experimental Animal Care and Use
Committee at Jinzhou Medical University.
The animals were randomly divided into three groups
(n=8/group): The sham group, which underwent laminectomy; the SCI
group, in which SCI was induced through the Allen method (13), followed by oral administration of
0.1% DMSO/saline solution 1 h after SCI and then once daily for 7
consecutive days; and the icariin treatment group, with oral
administration of 35 µmol/kg icariin (Sigma-Aldrich; Merck KGaA) 1
h after SCI and then once daily for 7 consecutive days. Icariin was
dissolved in 0.1% DMSO/saline solution. The icariin dose used in
the present study was determined based on a previous study by Tohda
and Nagata (12); in that study,
icariin effectively ameliorated SCI-induced motor dysfunction at a
50 µmol/kg dose in mice, which is equivalent to a 35 µmol/kg dose
in rats as used in the present study, according to standard dose
conversion between mice and rats (the conversion coefficient
between mice and rats is 0.7). As for the duration of medication
therapy, the study by Tohda and Nagata (12) showed that 3 consecutive days'
administration of icariin to the SCI mice produced a marked
protective effect. The duration was increased to 7 days in the
present study in order to ensure that the effectiveness of the
agent could be seen.
Establishment of the SCI model
The SCI model was established as described
previously (14). In brief, rats
were anesthetized intraperitoneally with 40 mg/kg sodium
pentobarbital and the spinal cord was exposed under aseptic
conditions after laminectomy at T9/10. Subsequently, an impactor of
2 mm diameter and weighing 10 g was dropped from a height of 25 mm
onto the surface of T9/10, causing spinal cord congestion. In the
sham group, only laminectomy was performed.
Locomotion recovery assessment
The locomotion recovery of all rats was assessed
using the Basso, Beattie and Bresnahan (BBB) open-field locomotor
rating scale, as described previously (7). Two independent examiners who were
blinded to the grouping assessed the scores of each group. To
ensure there was no baseline discrepancy, BBB scoring was also
performed prior to the surgery. The BBB rating scale was scored
0–21, with higher scores indicating better locomotor function.
Determination of spinal cord water
content
Following treatment with icariin for 7 days, the
water content of the spinal cord was evaluated. All rats were
sacrificed by lethal intraperitoneal injection of 180 mg/kg
sodium pentobarbital, and the spinal cord samples were removed,
placed on ice and then dried for 48 h at 80°C for the determination
of the dry weights. The water content of the spinal cord was
calculated as follows: [(wet weight-dry weight)/wet
weight]x100%.
Western blot analysis
The rats were anesthetized on day 7 after SCI, and
the damaged spinal cord was removed around the injury epicenter (5
mm cephalad and caudally). The tissues were lysed in RIPA buffer
containing 1% PMSF (Beyotime Institute of Biotechnology). The
protein content was determined using a bicinchoninic acid kit
(Beyotime Institute of Biotechnology). Western blotting was
performed as described previously (15). The antibodies used were as follows:
Anti-Bax (rabbit polyclonal antibody; 1:500 dilution; cat. no.
ab53154), anti-Bcl-2 (rabbit polyclonal antibody; 1:1,000 dilution;
cat. no. ab196495), anti-cleaved caspase 3 (rabbit polyclonal
antibody; 1:1,000 dilution; cat. no. ab49822), anti-cleaved caspase
9 (rabbit polyclonal antibody; 1:500 dilution; cat. no. ab25758),
anti-nuclear factor erythroid 2-related factor 2 (Nrf2; rabbit
polyclonal antibody; 1:1,000 dilution; cat. no. ab137550),
anti-NADPH-quinone oxidoreductase-1 (NQO1; rabbit polyclonal
antibody; 1:1,000 dilution; cat. no. ab217302), anti-heme oxygenase
(HO)-1 (rabbit monoclonal antibody; 1:1,500 dilution; cat. no.
ab189491), anti-inducible nitric oxide synthase (iNOS; mouse
monoclonal antibody; 1:2,000 dilution; cat. no. ab49999),
anti-nuclear factor (NF)-κB (rabbit polyclonal antibody; 1:1,000
dilution; cat. no. ab16502) and anti-GAPDH (rabbit monoclonal
antibody; 1:10,000 dilution; cat. no. ab181603). All the primary
antibodies were obtained from Abcam (Cambridge, MA, USA).
Polyclonal horseradish peroxidase-conjugated secondary goat
anti-rabbit antibodies (1:2,500; cat. no. sc-2004) and goat
anti-mouse antibodies (1:3,500; cat. no. sc-2005) were purchased
from Santa Cruz Biotechnology, Inc (Dallas, TX, USA). The relative
optical density of the bands was analyzed using ImageJ 2× software
(National Institutes of Health).
Biochemical index determination
The biochemical indices of spinal cord samples were
determined using commercial kits, following the manufacturers'
instructions. The levels of tumor necrosis factor (TNF)-α (cat. no.
RAB0480), interleukin (IL)-1β (cat. no. RAB0278) and IL-6 (cat. no.
RAB0312) were detected using ELISA kits (Sigma Aldrich; Merck
KGaA). The production of nitric oxide (NO) was determined using a
NO assay kit (cat. no. S0024; Beyotime Institute of Biotechnology).
Caspase-3 (cat. no. E13183) and −9 (cat. no. KHZ0101) activity
levels in the spinal tissue were evaluated using colorimetric
protease activation kits (Invitrogen; Thermo Fisher Scientific,
Inc.). Commercially available kits for the determination of
superoxide dismutase (SOD; cat. no. A001-1) activity, glutathione
(GSH; cat. no. A006-2) content, reactive oxygen species (ROS; cat.
no. E004-1-1) generation, and malondialdehyde (MDA; cat. no.
A003-1) level were purchased from Nanjing Jiancheng Bioengineering
Institute.
Statistical analysis
All data are presented as the mean ± standard error
of the mean from at least five independent experiments and were
analyzed using SPSS v.17.0 (SPSS, Inc.). One-way analysis of
variance with the Student-Newman-Keuls post hoc test was used for
comparisons among multiple groups. P<0.05 was considered to
indicate statistically significant differences.
Results
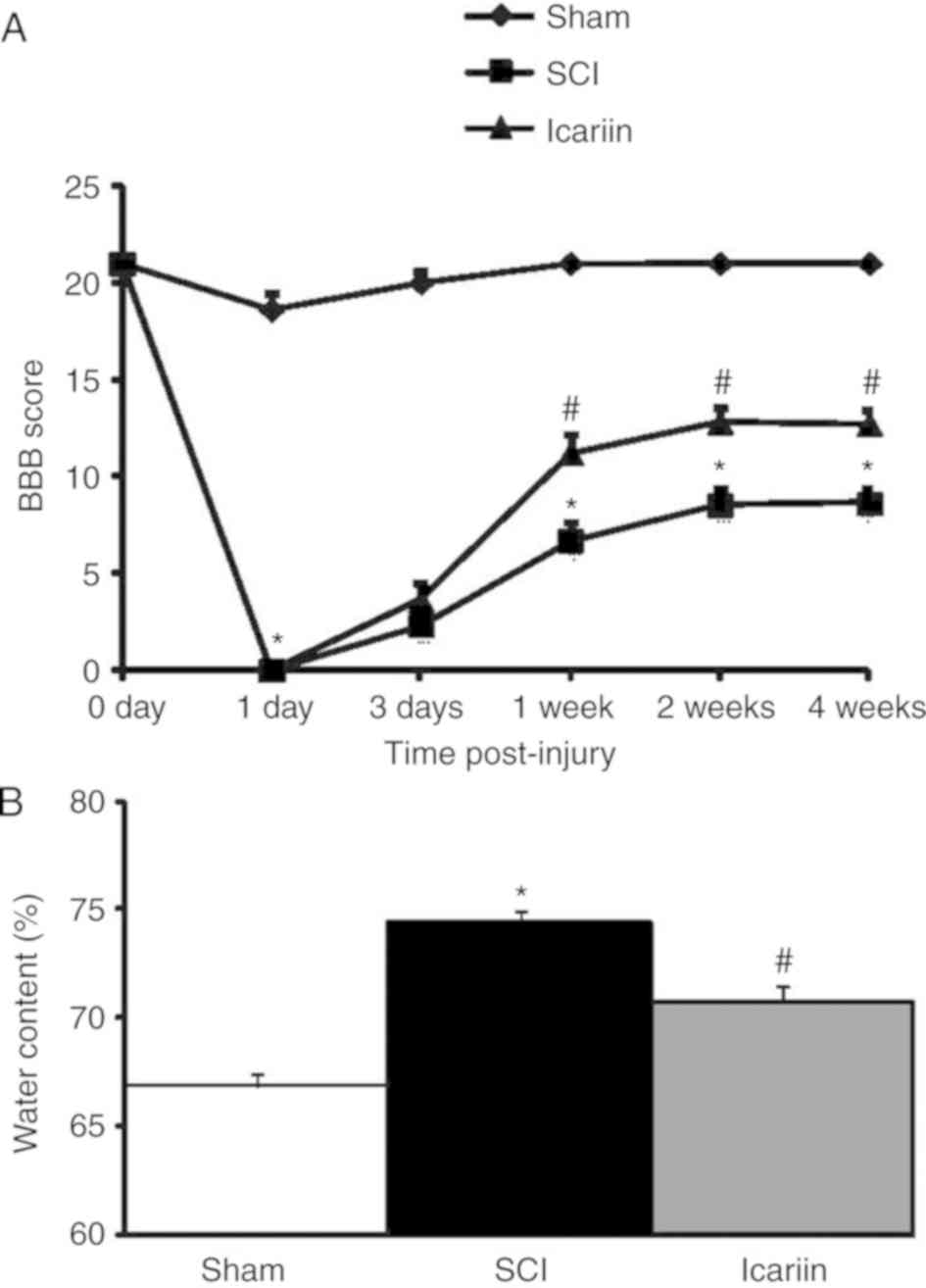
Effect of icariin on locomotor
function recovery and spinal cord water content following SCI
The BBB locomotor rating scale was applied to assess
the locomotor performance of rats at 0, 1, 3, 7, 14 and 28 days
post-injury. As shown in Fig. 1A,
a marked reduction in locomotor function was observed on the first
day after SCI and, over the following days, a gradual recovery was
also observed. Compared with the sham group, a significantly faster
improvement of locomotor function was recorded in icariin-treated
rats at 7, 14 and 28 days after the operation (P<0.05). Rat
spinal cord water content was increased in the SCI group compared
with the sham group, while icariin significantly alleviated the
increase in spinal cord water content (P<0.05; Fig. 1B).
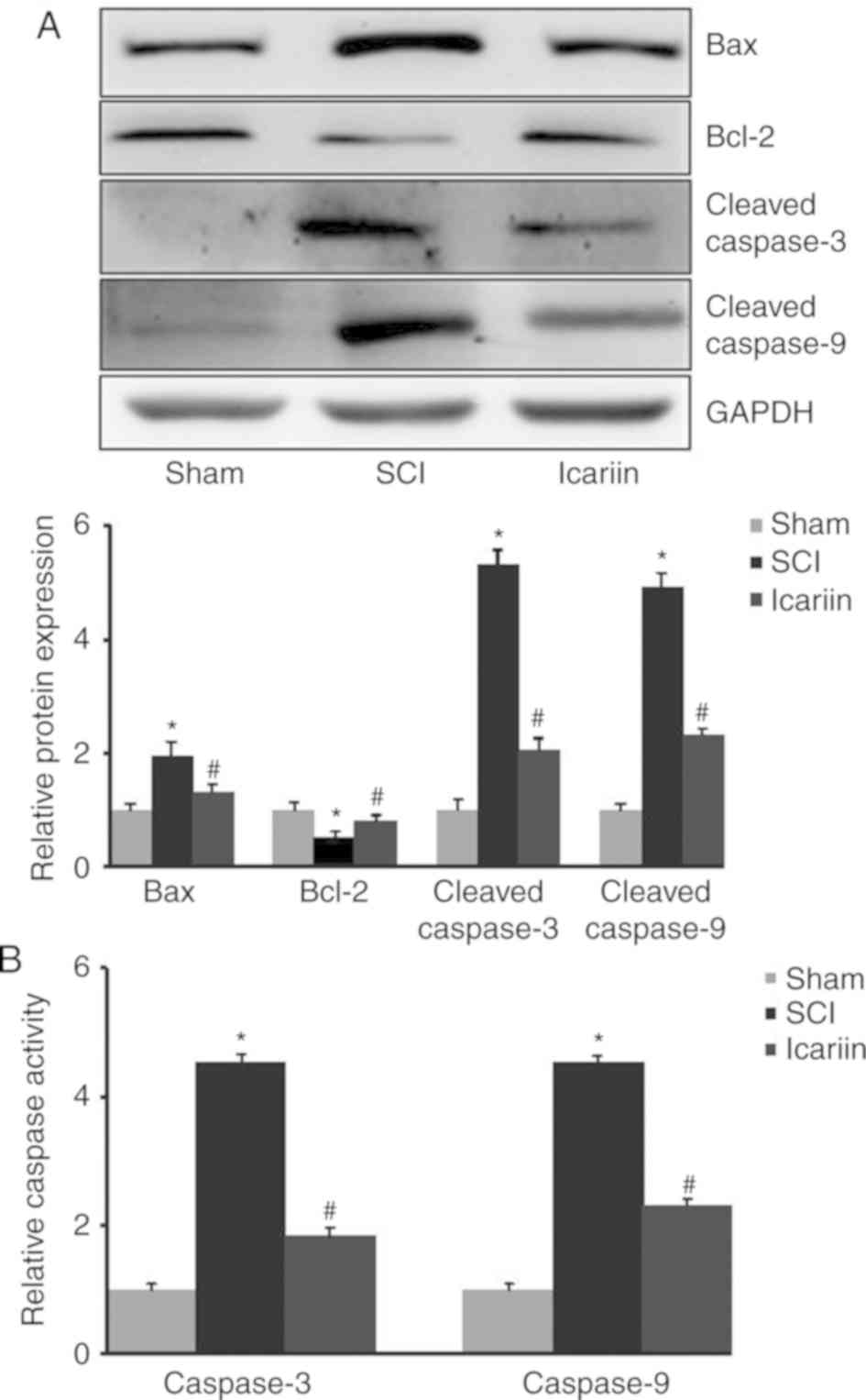
Effect of icariin on the expression
and activity of apoptosis-related proteins in rats with SCI
SCI rats exhibited higher expression levels of
pro-apoptotic proteins, including Bax, cleaved caspase 3 and
cleaved caspase 9. However, the level of the anti-apoptotic factor
Bcl-2 was significantly reduced following SCI. Accordingly, caspase
3 and 9 activities were significantly increased upon SCI. All these
changes were notably reversed by the application of icariin
(P<0.05; Fig. 2).
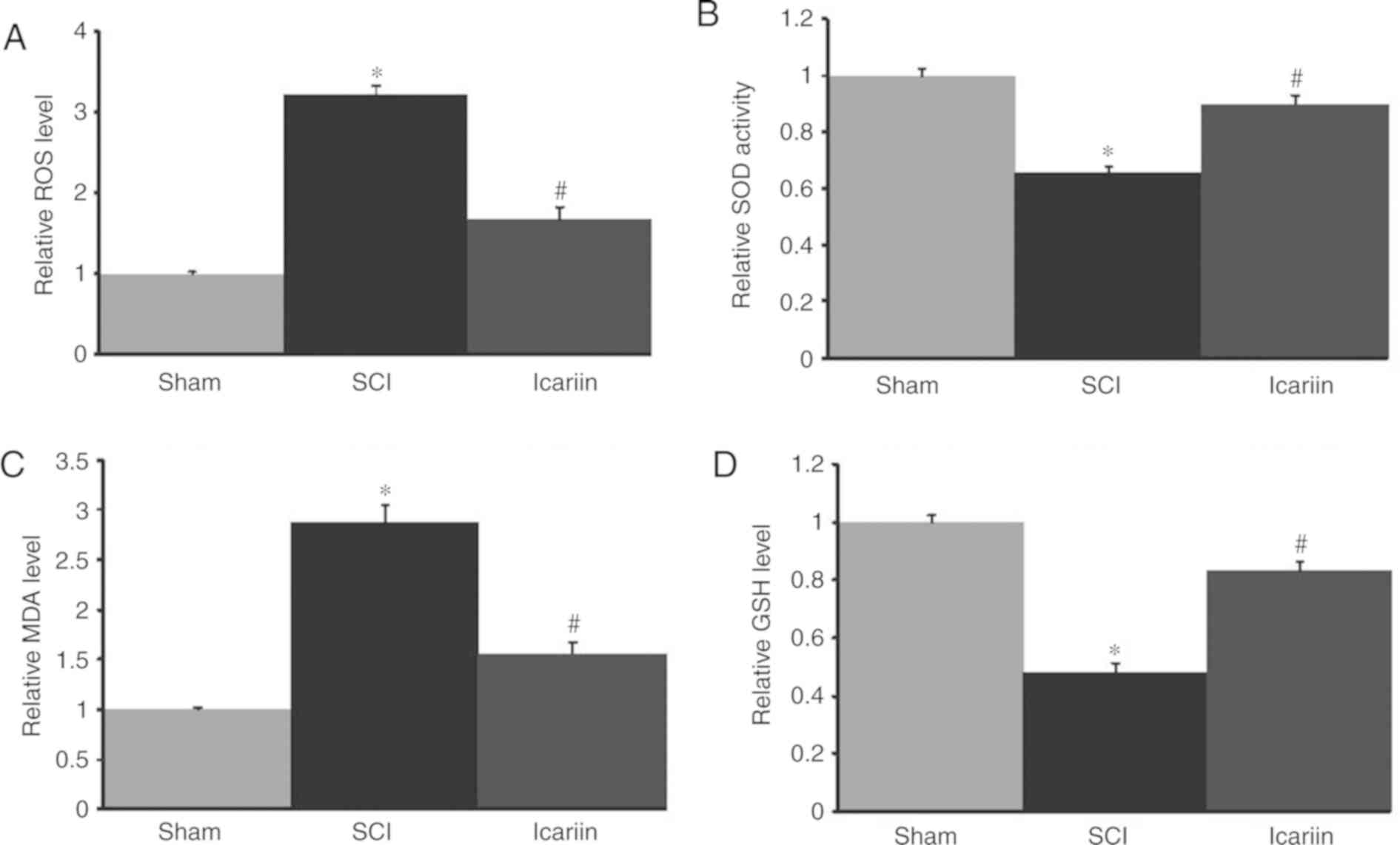
Effect of icariin on oxidative stress
in rats with SCI
SCI rats exhibited higher levels of oxidative stress
when compared with the sham group, as evidenced by an increased ROS
level and MDA content, as well as decreased SOD activity and GSH
levels. All these indices were restored with icariin treatment
(P<0.05; Fig. 3).
Effect of icariin on the expression of
inflammation-associated factors
The present study further investigated whether
icariin exerted protective effects against SCI damage by inhibiting
the expression of inflammatory factors, such as NF-κB and iNOS.
Western blotting revealed a significant (P<0.05) elevation of
NF-κB and iNOS protein expression in the SCI group compared with
the sham group, whereas icariin treatment significantly decreased
the protein expression of NF-κB and iNOS compared with the SCI
group (P<0.05; Fig. 4A). These
results indicated that icariin attenuated inflammatory responses in
the spinal cord of rats subjected to SCI. In addition, SCI
increased the levels of TNF-α, IL-1β, IL-6 and NO in comparison to
the sham group, and these inflammatory cytokines were reduced in
the icariin group compared with the SCI group (P<0.05; Fig. 4B).
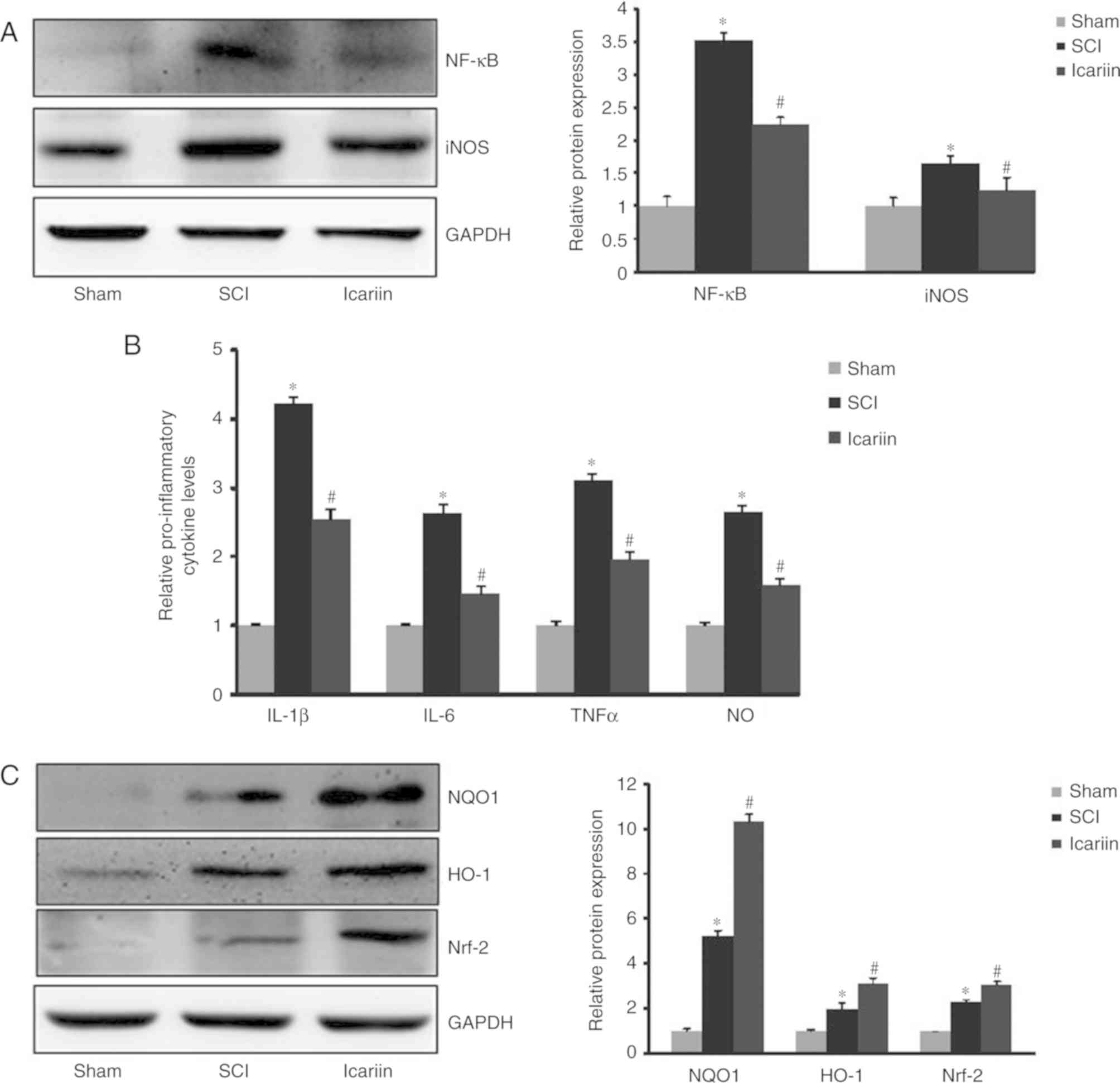 | Figure 4.Icariin exerts anti-inflammatory
effects in the spinal cords of SCI rats. (A) Expression of NF-κB
and iNOS in spinal cord tissues of SCI rats with or without 7-day
application of icariin. (B) Analysis of pro-inflammatory cytokine
levels of IL-1β, IL-6, TNF-α and NO following 7-day icariin
treatment. (C) Analysis of anti-inflammatory proteins NQO1, HO-1
and Nrf2 following 7-day icariin treatment. Data are presented as
mean ± standard error of the mean (n=5 rats/group). *P<0.05 vs.
respective sham group; #P<0.05 vs. respective SCI
group. SCI, spinal cord injury; NF-κB, nuclear factor-κB; iNOS,
inducible nitric oxide synthase; IL, interleukin; TNF, tumor
necrosis factor; NO, nitric oxide; NQO1, NADPH-quinone
oxidoreductase-1; HO, heme oxygenase; Nrf2, nuclear factor
erythroid 2-related factor 2. |
To explore the molecular mechanism underlying the
neuroprotective effects of icariin administration, western blotting
was performed to measure the expression of Nrf2, NQO1 and HO-1,
which are all important anti-inflammatory proteins exerting
neuroprotective effects during SCI. The results of the western
blotting demonstrated that the levels of Nrf2, NQO1 and HO-1 were
all elevated following SCI damage, and icariin treatment further
enhanced their expression (P<0.05; Fig. 4C).
Discussion
Icariin is a flavonoid found in the Chinese
medicinal herb Epimedium sagittatum Maxim. Accumulating
evidence indicates that icariin has potent antioxidant, anti-aging,
anti-tumor, anti-osteoporosis and neuroprotective properties
(16–19). Regarding the neuroprotective
effects of icariin, evidence has shown that icariin may protect
against Alzheimer's disease (20,21),
Parkinson's disease (21,22), multiple sclerosis (21) and SCI (12). In accordance with previously
published results, the present study demonstrated that icariin
improved the locomotor function of SCI rats, accompanied by a
decrease in the spinal cord water content. Furthermore, the present
results also revealed that apoptotic-, redox- and
inflammatory-related signaling mechanisms were involved in the
icariin-mediated anti-SCI effects.
Following SCI, apoptotic death of neural cells is
commonly observed in animal models as well as human tissue, and
plays an important role in functional disability (23). This process is regulated by Bcl-2,
Bax and caspases 3 and 9, indicating the participation of the
intrinsic mitochondrial apoptotic pathway in the pathological
process following SCI (24,25).
In accordance with previous studies, the present study demonstrated
that, following SCI, the expression of Bax, cleaved caspase 3 and
cleaved caspase 9, and the activity of caspases 3 and 9, were all
upregulated, accompanied by a decreased Bcl-2 level. Notably, the
present data demonstrated that icariin treatment significantly
attenuated neural cell apoptosis in SCI rats, as evidenced by the
reversal of these apoptosis-related indices. In fact, considerable
evidence has suggested the protective role of icariin against
mitochondrial apoptotic pathway induction in several pathological
conditions (26–28). Due to the absence of morphological
results, the present study did not elucidate which types of cells
are implicated in the neuroprotective effects of icariin. A recent
study by Li et al (29)
reported that the neuroprotective effects of icariin may be
associated with both neuronal and non-neuronal cells; however, this
hypothesis requires further confirmation.
The imbalance between oxidants and antioxidants
causes oxidative stress, further resulting in cellular damage.
Accumulating evidence has confirmed the critical role of oxidative
stress in secondary SCI (30). GSH
and SOD are important antioxidants, and MDA is an end-product of
lipid peroxidation; these are all widely used as markers of
oxidative stress following SCI (31). In the present study, SCI induced
marked oxidative stress, as evidenced by increased ROS and MDA
levels, with decreased GSH levels and SOD activity, consistent with
previous findings. Recently, an increasing number of studies have
reported the antioxidant properties of icariin (32,33).
Consistently, the present data demonstrated that icariin treatment
markedly alleviated oxidative stress responses in the spinal cord
tissue of SCI rats, which may represent an important
neuroprotective mechanism of this flavonoid.
Inflammation is a crucial factor in secondary damage
following SCI, and anti-inflammatory therapy contributes to
recovery from SCI (34,35). Nrf2 is a factor that regulates the
transcription of cytoprotective genes, including HO-1 and NQO1,
thereby playing a pivotal role in anti-inflammatory response
(36). Previous studies have
confirmed that a number of agents or treatments exert their
anti-neuroinflammatory and antioxidant effects in neuronal and
neurodegenerative conditions, including SCI, through upregulation
of the expression of these signaling molecules (36–39).
It was previously demonstrated that icariin exhibited potent
anti-inflammatory activity (16,40,41).
The present study revealed that the expression of all three
anti-inflammatory molecules was upregulated in SCI rats, and
icariin treatment further promoted their expression. These findings
indicated that there may be a compensatory or adaptive upregulation
of anti-inflammatory molecules in SCI, which may help alleviate
SCI-induced damage, and icariin treatment may further enhance this
protective effect.
NF-κB is a transcription factor that controls the
gene expression of a number of inflammation-related proteins that
contribute to SCI, such as TNF-α, IL-1β and IL-6 (42). It has been documented that icariin
and its metabolite, icariside II, exert their anti-inflammatory
effects via inhibition of the NF-κB signaling pathway (43). In the present study, a significant
elevation of NF-κB p65 subunit expression was observed in SCI rats,
and icariin treatment markedly suppressed its expression,
implicating the involvement of NF-κB pathway inhibition in
icariin-mediated anti-inflammatory action during the attenuation of
SCI in rats.
It was reported that inflammatory reactions may
cause increased expression of iNOS, leading to the excessive NO
production post-SCI (9,44). High levels of NO exert a cytotoxic
effect on the spinal cord. In fact, selective inhibition of iNOS by
aminoguanidine has been found to promote the recovery of
neurological function in rats subjected to SCI (45). Consistent with previous results,
the present findings demonstrated that the protein level of iNOS
and NO production were evidently increased in SCI rats. More
importantly, the administration of icariin markedly reduced these
indices in the traumatic SCI rat model. Thus, it may be concluded
that icariin exerts its protective effects against spinal cord
functional impairment by suppressing iNOS/NO. Although the present
study and a recent report (29)
support the possible involvement of the suppression of iNOS/NO
signaling in icariin-mediated neuroprotection against SCI, the
underlying mechanism remains unclear and requires further
investigation.
Previous studies have shown that icariin possesses
anti-apoptotic, antioxidant and anti-inflammatory bioactivities,
thereby improving the recovery of locomotor function in SCI rats
(12,29,46).
The findings of the present study further supported the soundness
of these arguments by examining additional apoptotic-, redox- and
inflammatory-related mechanisms. The results of the present study
included the attenuating effect of icariin on the activities of
caspases 3 and 9 and levels of NF-κB, IL-6, NO and ROS, as well as
the enhancement of anti-inflammatory factors, such as NQO-1, HO-1
and Nrf-2. It should be noted that, although the present study
using an animal model demonstrated that icariin treatment may be a
promising therapeutic strategy for SCI, the therapeutic
effectiveness of this drug requires further confirmation in
clinical trials.
Acknowledgements
Not applicable.
Funding
The present study was supported by President
Foundation, Aohongboze Foundation of Liaoning Medical University
(grant no. XZJJ20140230), and partially by National Natural Science
Foundation of China (grant no. 81700519).
Availability of data and materials
The datasets generated and analyzed in the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HD conceived and designed the study. GJ performed
the experiments and completed the manuscript draft. GJ, YZ and WL
analyzed the data. All the authors have read and approved the final
version of this manuscript for publication.
Ethics approval and consent to
participate
All procedures were approved by the Experimental
Animal Care and Use Committee at Jinzhou Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Xu M, Li H, Zhao Z, Yang Y, Sun Z, Han H,
Zhang X and Reinhardt JD: Environmental barriers, functioning and
quality of life in 2008 Wenchuan earthquake victims with spinal
cord injury eight years after the disaster: A cross-sectional
study. J Rehabil Med. 50:866–871. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maitan P, Frigerio S, Conti A, Clari M,
Vellone E and Alvaro R: The effect of the burden of caregiving for
people with spinal cord injury (SCI): A cross-sectional study. Ann
Ist Super Sanita. 54:185–193. 2018.PubMed/NCBI
|
|
3
|
Quadri SA, Farooqui M, Ikram A, Zafar A,
Khan MA, Suriya SS, Claus CF, Fiani B, Rahman M, Ramachandran A, et
al: Recent update on basic mechanisms of spinal cord injury.
Neurosurg Rev. July 11–2018.(Epub ahead of print). View Article : Google Scholar
|
|
4
|
Sobrido-Camean D and Barreiro-Iglesias A:
Role of Caspase-8 and fas in cell death after spinal cord injury.
Front Mol Neurosci. 11:1012018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rudman MD, Choi JS, Lee HE, Tan SK, Ayad
NG and Lee JK: Bromodomain and extraterminal domain-containing
protein inhibition attenuates acute inflammation after spinal cord
injury. Exp Neurol. 309:181–192. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Donovan J and Kirshblum S: Clinical trials
in traumatic spinal cord injury. Neurotherapeutics. 15:654–668.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang Y, Gong FL, Zhao GB and Li J:
Chrysin suppressed inflammatory responses and the inducible nitric
oxide synthase pathway after spinal cord injury in rats. Int J Mol
Sci. 15:12270–12279. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kang S, Liu S, Li H, Wang D and Qi X:
Baicalin effects on rats with spinal cord injury by
anti-inflammatory and regulating the serum metabolic disorder. J
Cell Biochem. 119:7767–7779. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhou L, Ouyang L, Lin S, Chen S, Liu Y,
Zhou W and Wang X: Protective role of β-carotene against oxidative
stress and neuroinflammation in a rat model of spinal cord injury.
Int Immunopharmacol. 61:92–99. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu P, Zhang Z, Wang Q, Guo R and Mei W:
Lithium chloride facilitates autophagy following spinal cord injury
via ERK-dependent pathway. Neurotox Res. 32:535–543. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Silva NA, Sousa N, Reis RL and Salgado AJ:
From basics to clinical: A comprehensive review on spinal cord
injury. Prog Neurobiol. 114:25–57. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tohda C and Nagata A: Epimedium koreanum
extract and its constituent icariin improve motor dysfunction in
spinal cord injury. Evid Based Complement Alternat Med.
2012:7312082012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang C, Liu C, Gao K, Zhao H, Zhou Z, Shen
Z, Guo Y, Li Z, Yao T and Mei X: Metformin preconditioning provide
neuroprotection through enhancement of autophagy and suppression of
inflammation and apoptosis after spinal cord injury. Biochem
Biophys Res Commun. 477:534–540. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yacoub A, Hajec MC, Stanger R, Wan W,
Young H and Mathern BE: Neuroprotective effects of perflurocarbon
(oxycyte) after contusive spinal cord injury. J Neurotrauma.
31:256–267. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dai H, Song D, Xu J, Li B, Hertz L and
Peng L: Ammonia-induced Na,K-ATPase/ouabain-mediated EGF receptor
transactivation, MAPK/ERK and PI3K/AKT signaling and ROS formation
cause astrocyte swelling. Neurochem Int. 63:610–625. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang GQ, Li DD, Huang C, Lu DS, Zhang C,
Zhou SY, Liu J and Zhang F: Icariin reduces dopaminergic neuronal
loss and microglia-mediated inflammation in vivo and in vitro.
Front Mol Neurosci. 10:4412018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou J, Wu J, Chen X, Fortenbery N,
Eksioglu E, Kodumudi KN, Pk EB, Dong J, Djeu JY and Wei S: Icariin
and its derivative, ICT, exert anti-inflammatory, anti-tumor
effects, and modulate myeloid derived suppressive cells (MDSCs)
functions. Int Immunopharmacol. 11:890–898. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu Y, Zuo H, Liu X, Xiong J and Pei X:
The antiosteoporosis effect of icariin in ovariectomized rats: A
systematic review and meta-analysis. Cell Mol Biol
(Noisy-le-grand). 63:124–131. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li C, Li Q, Mei Q and Lu T:
Pharmacological effects and pharmacokinetic properties of icariin,
the major bioactive component in Herba Epimedii. Life Sci.
126:57–68. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sheng C, Xu P, Zhou K, Deng D, Zhang C and
Wang Z: Icariin attenuates synaptic and cognitive deficits in an
Aβ1-42-induced rat model of Alzheimer's disease. Biomed Res Int.
2017:74648722017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jin J, Wang H, Hua X, Chen D, Huang C and
Chen Z: An outline for the pharmacological effect of icariin in the
nervous system. Eur J Pharmacol. 842:20–32. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen WF, Wu L, Du ZR, Chen L, Xu AL, Chen
XH, Teng JJ and Wong MS: Neuroprotective properties of icariin in
MPTP-induced mouse model of Parkinson's disease: Involvement of
PI3K/Akt and MEK/ERK signaling pathways. Phytomedicine. 25:93–99.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen XB, Wang ZL, Yang QY, Zhao FY, Qin
XL, Tang XE, Du JL, Chen ZH, Zhang K and Huang FJ: Diosgenin
glucoside protects against spinal cord injury by regulating
autophagy and alleviating apoptosis. Int J Mol Sci. 19(pii):
E22742018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Z, Zhou L, Zheng X, Chen G, Pan R, Li
J and Liu W: Autophagy protects against PI3K/Akt/mTOR-mediated
apoptosis of spinal cord neurons after mechanical injury. Neurosci
Lett. 656:158–164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhao H, Chen S, Gao K, Zhou Z, Wang C,
Shen Z, Guo Y, Li Z, Wan Z, Liu C and Mei X: Resveratrol protects
against spinal cord injury by activating autophagy and inhibiting
apoptosis mediated by the SIRT1/AMPK signaling pathway.
Neuroscience. 348:241–251. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qian ZQ, Wang YW, Li YL, Li YQ, Ling-Zhu
and Yang DL: Icariin prevents hypertension-induced cardiomyocyte
apoptosis through the mitochondrial apoptotic pathway. Biomed
Pharmacother. 88:823–831. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Q, Hao J, Pu J, Zhao L, Lü Z, Hu J,
Yu Q, Wang Y, Xie Y and Li G: Icariin induces apoptosis in mouse
MLTC-10 Leydig tumor cells through activation of the mitochondrial
pathway and down-regulation of the expression of piwil4. Int J
Oncol. 39:973–980. 2011.PubMed/NCBI
|
|
28
|
Li S, Dong P, Wang J, Zhang J, Gu J, Wu X,
Wu W, Fei X, Zhang Z, Wang Y, et al: Icariin, a natural flavonol
glycoside, induces apoptosis in human hepatoma SMMC-7721 cells via
a ROS/JNK-dependent mitochondrial pathway. Cancer Lett.
298:222–230. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li H, Zhang X, Zhu X, Qi X, Lin K and
Cheng L: The effects of icariin on enhancing motor recovery through
attenuating pro-inflammatory factors and oxidative stress via
mitochondrial apoptotic pathway in the mice model of spinal cord
injury. Front Physiol. 9:16172018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kong G, Huang Z, Ji W, Wang X, Liu J, Wu
X, Huang Z, Li R and Zhu Q: The ketone metabolite β-hydroxybutyrate
attenuates oxidative stress in spinal cord injury by suppression of
class I histone deacetylases. JJ Neurotrauma. 34:2645–2655. 2017.
View Article : Google Scholar
|
|
31
|
Cong L and Chen W: Neuroprotective effect
of ginsenoside Rd in spinal cord injury rats. Basic Clin Pharmacol
Toxicol. 119:193–201. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sun S, Liu L, Tian X, Guo Y, Cao Y, Mei Y
and Wang C: Icariin attenuates high glucose-induced apoptosis,
oxidative stress, and inflammation in human umbilical venous
endothelial cells. Planta Med. 85:473–482. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zheng Y, Zhu G, He J, Wang G, Li D and
Zhang F: Icariin targets Nrf2 signaling to inhibit
microglia-mediated neuroinflammation. Int Immunopharmacol.
73:304–311. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu G, Shi D, Zhi Z, Ao R and Yu B:
Melatonin ameliorates spinal cord injury by suppressing the
activation of inflammasomes in rats. J Cell Biochem. 120:5183–5192.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li Z, Yao F, Cheng L, Cheng W, Qi L, Yu S,
Zhang L, Zha X and Jing J: Low frequency pulsed electromagnetic
field promotes the recovery of neurological function after spinal
cord injury in rats. J Orthop Res. 37:449–456. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Park SY, Kim YH and Park G: Cucurbitacins
attenuate microglial activation and protect from neuroinflammatory
injury through Nrf2/ARE activation and STAT/NF-κB inhibition.
Neurosci Lett. 609:129–136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wei W, Shurui C, Zipeng Z, Hongliang D,
Hongyu W, Yuanlong L, Kang Z, Zhaoliang S, Yue G, Chang L and Mei
X: Aspirin suppresses neuronal apoptosis, reduces tissue
inflammation, and restrains astrocyte activation by activating the
Nrf2/HO-1 signaling pathway. Neuroreport. 29:524–531. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xia P, Gao X, Duan L, Zhang W and Sun YF:
Mulberrin (Mul) reduces spinal cord injury (SCI)-induced apoptosis,
inflammation and oxidative stress in rats via miroRNA-337 by
targeting Nrf-2. Biomed Pharmacother. 107:1480–1487. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim Y, Kim J, Ahn M and Shin T: Lithium
ameliorates rat spinal cord injury by suppressing glycogen synthase
kinase-3β and activating heme oxygenase-1. Anat Cell Biol.
50:207–213. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kong L, Liang X, Liu A, Yang X, Luo Q, Lv
Y and Dong J: Icariin inhibits inflammation via immunomodulation of
the cutaneous hypothalamus-pituitary-adrenal axis in vitro. Clin
Exp Dermatol. 44:144–152. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hua W, Zhang Y, Wu X, Kang L, Tu J, Zhao
K, Li S, Wang K, Song Y, Luo R, et al: Icariin attenuates
interleukin-1β-induced inflammatory response in human nucleus
pulposus cells. Curr Pharm Des. 23:6071–6078. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen S, Ye J, Chen X, Shi J, Wu W, Lin W,
Lin W, Li Y, Fu H and Li S: Valproic acid attenuates traumatic
spinal cord injury-induced inflammation via STAT1 and NF-κB pathway
dependent of HDAC3. J Neuroinflammation. 15:1502018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hwang E, Lin P, Ngo HTT, Gao W, Wang YS,
Yu HS and Yi TH: Icariin and icaritin recover UVB-induced
photoaging by stimulating Nrf2/ARE and reducing AP-1 and NF-κB
signaling pathways: A comparative study on UVB-irradiated human
keratinocytes. Photochem Photobiol Sci. 17:1396–1408. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang S and Ren D: Allicin protects
traumatic spinal cord injury through regulating the HSP70/Akt/iNOS
pathway in mice. Mol Med Rep. 14:3086–3092. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li Z, Du J, Sun H, Mang J, He J, Wang J,
Liu H and Xu Z: Effects of the combination of methylprednisolone
with aminoguanidine on functional recovery in rats following spinal
cord injury. Exp Ther Med. 7:1605–1610. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ren XS, Ding W and Yang XY:
Neuroprotective effect of icariin on spinal cord injury in rats.
Zhongguo Gu Shang. 31:1054–1060. 2018.(In Chinese). PubMed/NCBI
|