Introduction
Secondary injury serves a key function in the
outcome of patients with spinal cord injury (SCI) (1). It is important to reduce the
apoptosis or necrosis of neurons, and maintain the links between
neurons/glial cells and axons in SCI (2,3). The
mechanism of secondary injury is complex (4). Numerous studies have identified that
microvascular perfusion changes, free radical production and lipid
peroxidation, necrosis and apoptotic cell death and the
dysregulation of ionic homeostasis are able to promote secondary
injury following SCI (5–8). Previous studies have attempted to
identify a desirable target which is able to interrupt
multi-mechanisms underlying secondary injury.
Nuclear factor erythroid 2p45-related factor 2
(Nrf2) is a member of the Cap ‘n’ Collar basic-leucine-zipper
family of transcription factors (9). Under numerous stimuli, Nrf2
translocates from the cytoplasm to the nucleus (10) and sequentially binds to antioxidant
response element (ARE) (11). ARE
is a promoter element commonly identified in protective genes and
its products are involved in reducing oxidative stress,
inflammatory damage and reducing the accumulation of toxic
metabolites (12). Nrf2
transactivates the expression of a number of cytoprotective enzymes
by binding to ARE motifs, including heme oxygenase-1 (HO-1) and
NAD(P)H-quinone oxidoreductase-1 (NQO1). These products regulated
by the Nrf2 gene, in order to protect the cell from oxidative or
xenobiotic damage (13–16).
It has been noted that ~7% of the normal number of
axons below the injury level are required to mediate meaningful
distal neurologic function (17,18).
In order to maintain the necessary neurologic functions, 1.4–12% of
the total number of axons across the spinal cord injury site are
required (19–21). Therefore, even small increases in
neuroprotection may affect functionally relevant neurologic
recovery and thus is important for SCI patients (22). However, neurons have low
antioxidant abilities and are highly sensitive to oxidative stress,
therefore, increased levels of reactive oxygen species (ROS) easily
induces neuron damage (23,24).
The Nrf2-ARE pathway in central nervous system (CNS) injury serves
a protective function (25). It
has been proven that Nrf2 serves pivotal functions in the cell,
which may defend against the oxidative stress of traumatic brain
injury (TBI) in rats or mice, and decrease the severity of
neurological deficit. On the contrary, Nrf2 knockout increased the
severity of TBI, even with the use of an Nrf2 inducer (26,27).
Furthermore, the disruption of Nrf2 may upregulate the activity of
nuclear factor-κβ and proinflammatory cytokines following TBI or
SCI in mice (28,29). The aim of the present study was to
investigate whether Nrf2 gene transfer overexpression can protect
neurons/glial cells, and the association between neurons/glial
cells and axons during SCI.
Materials and methods
Experimental overview
Gene transfer has been widely used for experimental
research (30). However, to the
best of our knowledge, Nrf2 gene transfer to TBI and SCI have not
been reported. In the present study, Nrf2 recombinant adenovirus
vectors were constructed that were then transfected into PC12 cells
and locally injected into SCI in rats. The protein levels of Nrf2
in the nucleus and the Nrf2-regulated gene expression of HO-1 and
NQO1 were detected using western blot analysis in PC12 cells
following 48 h of transfection. Furthermore, the expression of Nrf2
was localized by using immunofluorescence and the expression of
Nrf2, HO-1 and NQO1 were detected using immunohistochemistry in the
grey matter of the spinal cord in rats. Post-injury motor behavior
was assessed via the Basso, Beattie and Bresnahan (BBB) locomotor
scale method.
Cell line
PC12 cell line (a neuron model) was provided by
Department of Neurology of the First Affiliated Hospital of
Chongqing Medical University (Chongqing, China) and cultured in
RPMI 1640 medium (HyClone; GE Healthcare Life Sciences, Logan, UT,
USA) with 10% calf serum (HyClone; GE Healthcare Life Sciences), 5%
horse serum (HyClone; GE Healthcare Life Sciences), 100 U/ml
penicillin and 100 µg/ml streptomycin. Cells were cultured in a
humidified atmosphere incubator at 5% CO2 and 37°C, and
the medium was changed every other day.
Constructing recombinant adenoviral
vectors
The adenovirus shuttle plasmids pAV-MCMV-green
fluorescent protein (GFP)-Nrf2 and pAV-MCMV-GFP were purchased from
Microbix Biosystems (Mississauga, ON, Canada). Recombinant
adenoviral vectors were generated by using the Admax Cre-lox system
(Microbix Biosystems, Inc.). The adenovirus was propagated in 293
cells (Health Science Research Resources Bank Osaka, Japan). and
purified by CsCl2 density gradient centrifugation
(40,000 × g, 2 h, 4°C). Virus titers were determined using plaque
assays. For PC12 cell transfection, 2 ml PC12 cells
(1×105/ml) supernatant was incubated at 37°C in 6-well
plates overnight. Then they were infected for 20 min at 37°C with
viral supernatant containing vectors at a multiplicity of infection
of 100 in the presence of 8 µg/ml polybrene. PC12 cells were
divided into three groups: PC12-Control (no virus infection),
PC12-Ad-Nrf2 group (Ad-Nrf2 infection) and PC12-Ad-GFP group
(Ad-GFP infection).
Animal preparation
A total of 100 adult (5 weeks) male and female
Sprague-Dawley rats of Specific-pathogen free (SPF) (180–250 g;
Chongqing Medical University, Chongqing, China) were housed under a
room temperature and humidity (24°C and 50%) on a 12 h light-dark
cycle with ad libitum access to food and water. All
experimental procedures were performed in accordance with the
National Institutes of Health (NIH) Guide for the Care and Use of
Laboratory Animals (NIH publication no. 80-23, revised 1996), and
the number of animals used and their suffering were minimized.
Ethical approval was provided by the Yongchuan Hospital of
Chongqing Medical University (Chongqing, China). Using a random
number table, rats were divided into: i) a Sham-operated group
(n=25), ii) a SCI group (n=25), iii) an Ad-Nrf2 group (n=25), and
iv) an Ad-GFP group (n=25). On the 1st, 3rd, 7th, 14th and 28th day
subsequent to surgery, 5 surviving rats were selected from each
group for further experimental study.
The extradural compression of the modified Allen's
method was used to produce the SCI animal model (31). Briefly; rats were anesthetized
intraperitoneally with chloral hydrate (300 mg/kg) and underwent a
laminectomy to expose the dorsal portion of spinal cord from T8 to
T10 levels. Moderate or severe contusion injury was performed with
a weight-drop device by dropping a 10 g rod (3 mm in diameter, 5 cm
in height with injury pulse 10×5 gcf). Within 30 min following the
injury, 1 µl adenoviral vector, diluted to 5×1010 pfu/ml
prior to use, was injected into the 2 mm spinal cord stump from the
wound site vertically into each stump at a depth of 0.8 mm using a
Hamilton micro-injector. The injection rate was slow in order to
minimize damage to the spinal cord. Each animal was injected with a
total of 5×108 pfu viruses. A constant body temperature
was maintained with an overhead heating lamp during the experiment.
In the Sham-operated group, the rats underwent the same laminectomy
procedure but no trauma was produced. The GFP expression of PC12
cells were detected at 48 h after virus transfection with
fluorescence microscopy.
Western blot analysis
The proteins were extracted using a commercial kit
according to the manufacturer's protocol (NBP2-37853, Novus
Biologicals), and the protein expression of Nrf2, HO-1 and NQO1 in
a neuron model (PC12 cells) were analyzed using western blot
analysis. Briefly, 50 µg of protein extracts determined by Pierce
BCA Protein Assay Kit (23227, Thermo Fisher Scientific, Inc.) were
separated on 15% SDS-PAGE and then were electrotransferred to a
polyvinylidene difluoride filter (PVDF, EMD Millipore, Billerica,
MA, USA). The wet electroblotting (using Mini Trans-Blot Module,
1703935, Bio-Rad Laboratories, Inc.) was performed at constant
voltage (20 V) for 1 h at 4°C with CAPS based transfer buffer (10
mM CAPS, pH 11, 10% methanol) Then the membranes were blocked with
5% nonfat milk for 1 h at room temperature and incubated with
primary antibodies anti-Nrf2 (ab89443, 1:500; Abcam), anti-HO-1
(CL5275, 1:500; Abcam), anti-NQO1 antibodies (ab28947, 1:500;
Abcam) and anti β-actin (mAbcam 8226, 1:2,000; Abcam) overnight at
4°C, followed by anti-goat immunoglobulin G horseradish
peroxidase-conjugated secondary antibodies (sc-2354, 1:5,000; Santa
Cruz Biotechnology, Inc.) for 2 h at room temperature. Following
rinsing with a buffer, the protein bands were visualized using an
enhanced chemiluminescence kit (Beyotime Institute of
Biotechnology) according to the manufacturer's protocol. Film
signals were digitally quantified by internal control β-actin using
Quantity 4.6.2 software (Bio-Rad Laboratories, Inc.).
BBB locomotor scale
The BBB locomotor scale method was used to assess
the hind limb functional improvement of treated animals with spinal
cord contusion (32). At 1, 3, 7,
14 and 28 days subsequent to surgery, behavioral analysis was
performed based on movement of the hind limb, weight support,
forelimb-hind limb coordination and trunk stability.
GFP detection
All animals were perfused transcardially with 4%
paraformaldehyde. The cords, ~2 cm in length with the contused site
at the center of the sample, were segmented. The parasagittal
ice-frozen spinal cord sections (~500 µm away from the sagittal
plane, with a thickness of 10 µm) were prepared to detect GFP
expression with fluorescent microscopy.
Immunohistochemistry
Five samples from each group on the 3rd
postoperative day were fixed with 4% paraformaldehyde for 48 h at
room temperature, embedded in paraffin and cross-sections
(thickness, 10 µm) were prepared. After deparaffinization and
dehydration, sections were boiled in Tris-EDTA buffer (10 mM Tris
Base, 1 mM EDTA Solution, 0.05% Tween-20, pH 9.0) for antigen
unmasking, followed by extensive washing with PBS. Sections were
subsequently incubated with 3% H2O2 for 10
min, and then rinsed with PBS for three times. Sections were
blocked with 2% goat serum albumin in PBS for 20 min and incubated
with mouse anti-Nrf2, anti-HO-1 and anti-NQO1 antibodies (all at a
dilution of 1:100) overnight at 4°C. Sections incubated without a
primary antibody were used as negative controls. After washing 3
times with PBS, sections were incubated with Goat anti-Mouse IgG
Secondary Antibody [HRP (Horseradish Peroxidase), HAF007, 1;1,000,
Novus Biologicals] for 10 min at 37°C. After washing with PBS,
peroxidase was stained with Mouse specific HRP/DAB (ABC) Detection
IHC kit (ab64259; Abcam) and viewed under a light microscope
(magnification, ×400,; Olympus BX50; Olympus Corporation). The
positive cells were counted in five different fields of view in the
gray matter in five random sections of each rat. All images were
captured using Sim PCI 6.0 (Compix Media, Inc.).
Statistical analysis
Data were analyzed using SPSS 17.0 software (SPSS,
Inc., Chicago, IL, USA) and were presented as the mean ± standard
error of the mean. Statistical differences were measured using a
one-way analysis of variance and Bonferroni's test to compare the
differences between groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
PC12 cell data
GFP expression in PC12 cells was observed 48 h
subsequent to adenovirus transfection. It is likely that Nrf2 is
able to affect PC12 proliferation and axonal growth, so the
morphology is different between Fig.
1A-C and between Fig. 1D-F
(33). The expression of GFP
following Ad-Nrf2 transfection was mainly located in the nucleus,
and its expression in the cytoplasm was observed following Ad-GFP
transfection (Fig. 1A-F).
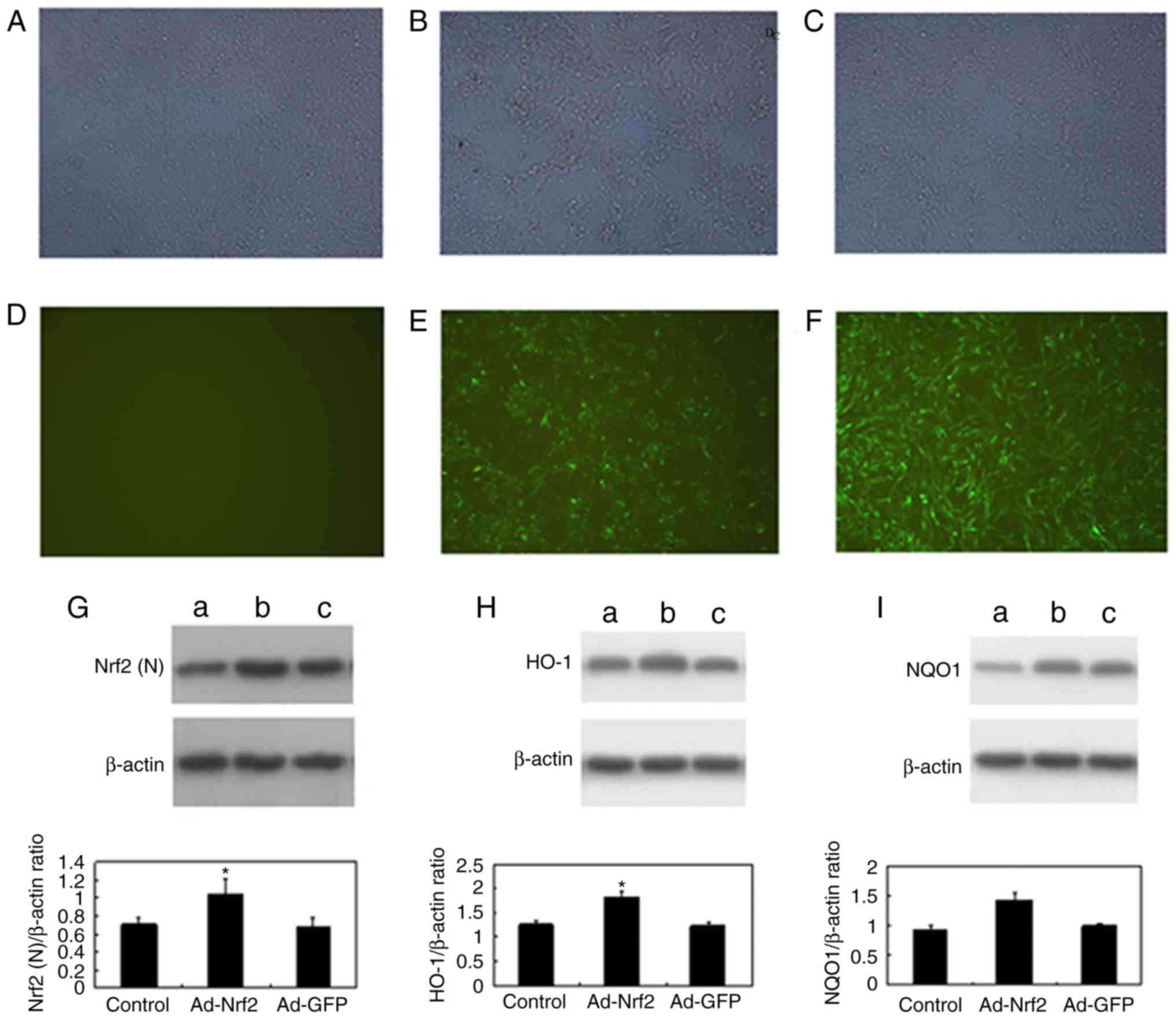 | Figure 1.Expression of GFP in cytoplasm 48 h
after recombinant adenovirus infection. Expression of GFP in the
(A) PC12-Control, (B) PC12-Ad-Nrf2 and (C) PC12-Ad-GFP groups
captured by light microscopy. Expression of GFP in the (D)
PC12-Control, (E) PC12-Ad-Nrf2 and (F) PC12-Ad-GFP groups captured
by fluorescent microscopy. Magnification, ×200. Subsequent to
Ad-Nrf2 transfection, western blot analysis was used to examine the
levels of (G) nuclear Nrf2, (H) HO-1 and (I) NQO1 in the
PC12-Control, PC12-Ad-Nrf2 and PC12-Ad-GFP groups. Bars represented
the ratio of the β-actin value (mean ± standard error of the mean).
*P<0.01 vs. control group. GFP, green fluorescent protein; Ad,
adenovirus; Nrf2, nuclear factor erythroid 2p45-related factor 2;
HO-1, heme oxygenase-1; NQO1, NAD(P)H: quinone
oxidoreductase-1. |
Western blot analysis was performed to evaluate the
protein levels of nuclear Nrf2 (57 kDa), HO-1 (32 kDa) and NQO1 (30
kDa) in whole cells. Subsequent to Ad-Nrf2 transfection, nuclear
Nrf2, HO-1 and NQO1 were significantly increased compared with the
control (P<0.01; Fig. 1G-I).
There was statistically significant changes in the PC12-Ad-Nrf2
group [Nrf2 (1.146±0.095), HO-1 (1.816±0.095) and NQO1
(1.421±0.138)] compared with the PC12-Control [Nrf2 (0.717±0.055),
HO-1 (1.264±0.081) and NQO1 (0.921±0.088)] and PC12-Ad-GFP group
[Nrf2 (0.714±0.111), HO-1 (1.238±0.053) and NQO1 (0.987±0.045);
(P<0.01)].
BBB Locomotor scale results
The animal behavioral analysis was performed at 1,
3, 7, 14 and 28 days following the operation. In the Sham-operated
group, BBB scores between day 1 and day 28 did not decrease
significantly. On the 3rd day following the operation, BBB scores
in the Ad-Nrf2 group (0.167±0.408) exhibited a significantly
decreased comparing with the SCI group (1±0.894; P<0.05). On the
7th day following the operation, the BBB scores in the Ad-Nrf2
group (2.333±0.516) were significantly increased compared with the
Ad-GFP group (0.9±0.21; P<0.05). On the 14th and 28th day, there
was a statistically significant increase in the Ad-Nrf2 group (day
14, 9.833±1.17; day 28, 14±2.608) compared with the SCI group (day
14, 8±1.265; day 28, 11.167±1.901) and the Ad-GFP group (day 14,
7.167±1.17; day 28, 10.333±1.633; P<0.05; Fig. 2).
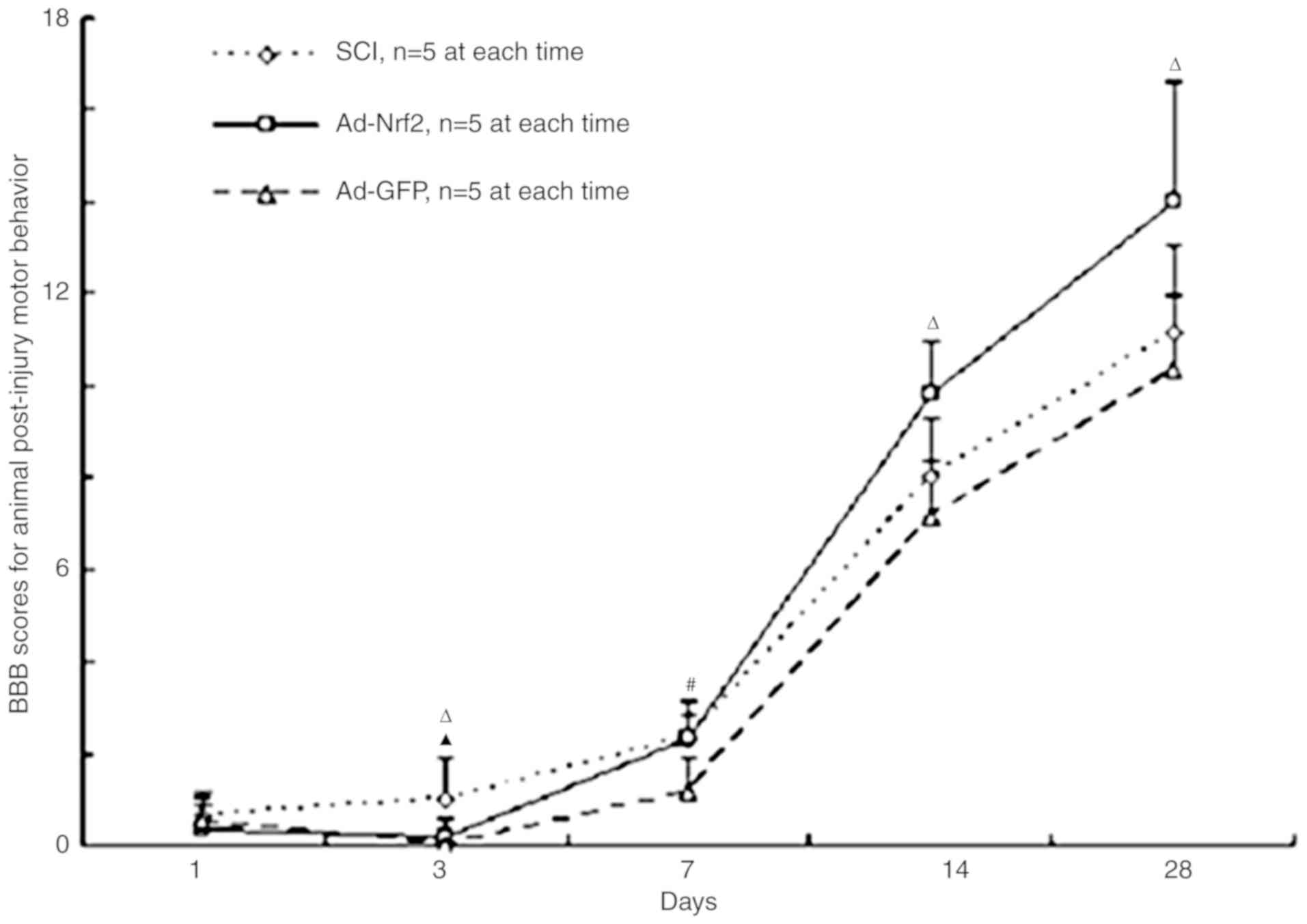 | Figure 2.BBB scores of post-injury motor
behavior for the SCI group, the SCI rats injected with Ad-Nrf2
(Ad-Nrf2 group) and the SCI rats injected with Ad-GFP (Ad-GFP
group). n=5, mean ± standard error of the mean. *P<0.05, SCI
group vs. the Ad-Nrf2 and Ad-GFP groups; #P<0.05, SCI
and Ad-Nrf2 groups vs. the Ad-GFP group; ΔP<0.05,
Ad-Nrf2 group vs. the SCI and Ad-GFP groups. SCI, spinal cord
injury; Nrf2, nuclear factor erythroid 2p45-related factor 2; BBB,
the Basso, Beattie and Bresnahan locomotor scale; GFP, green
fluorescent protein; Ad, adenovirus. |
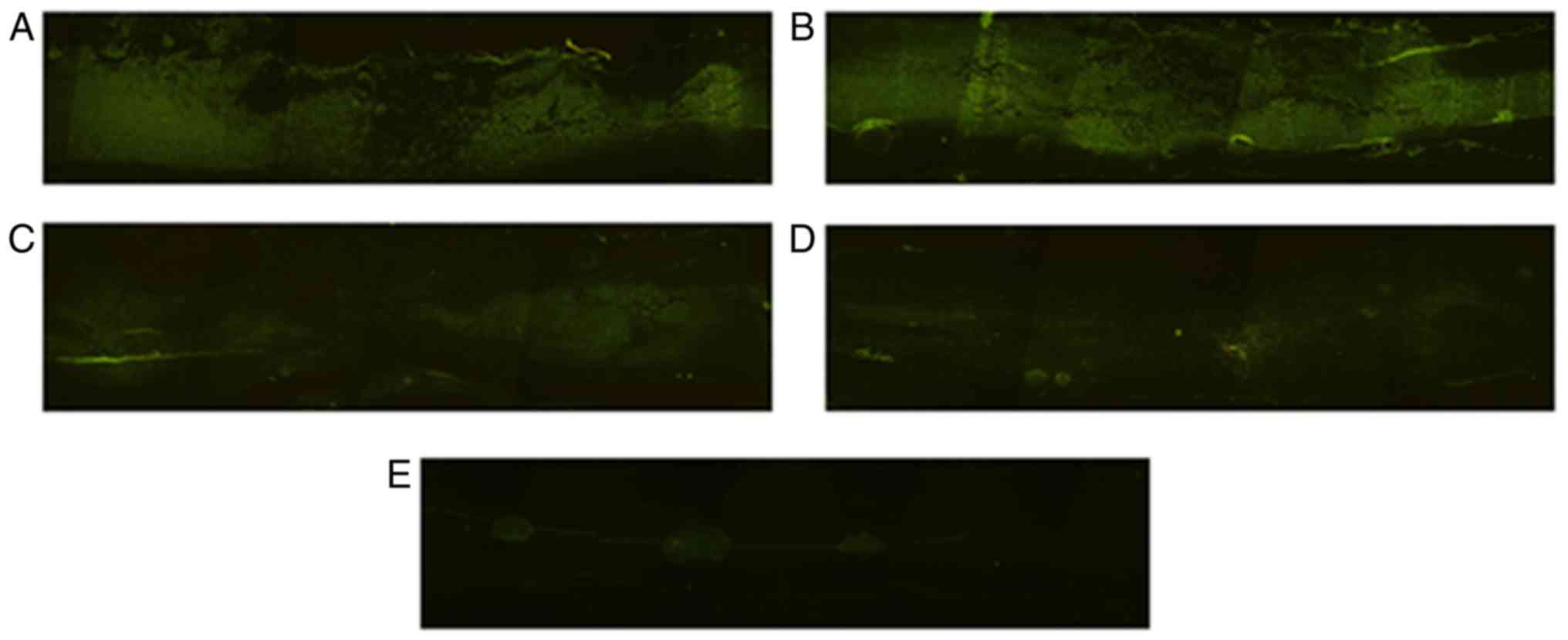
Gene transfer efficacy
To identify the efficacy of the gene transfer of
Ad-Nrf2 and Ad-GFP, the ice-frozen parasagittal sections of the
spinal cords, ~2 cm in length with the contused site at the center
of the sample, were detected using a fluorescence microscope. Green
fluorescence was detected in the spinal cords, primarily near the
injected sections, on the 1st day following SCI. The prevalence of
the fluorescence expression was observed on the 3rd day following
SCI and the contused parts also exhibited GFP expression. From that
point onwards, the fluorescence expression decreased gradually
(Fig. 3A-E).
Immunohistochemical study
The expression levels of Nrf2, HO-1 and NQO1 in grey
matter were localized and analyzed using an immunohistochemical
experiment on the 3rd day following the operation. Very few cells
were positive for Nrf2, HO-1 and NQO1 in the Sham-operated group.
Nrf2, HO-1 and NQO1 immunoreactivity was present in neurons and
glial cells following SCI. In the Ad-Nrf2 group, the positive
neurons and glial cells for Nrf2, HO-1 and NQO1 were significantly
increased compared with the control and SCI groups (P<0.01;
Fig. 4). It should be noted that
due to the poor homogeneity of spinal cord tissue, the tissue
specimens from different rats ought to be treated carefully at the
time of fixing and the section orientated to reduce the
morphological effects.
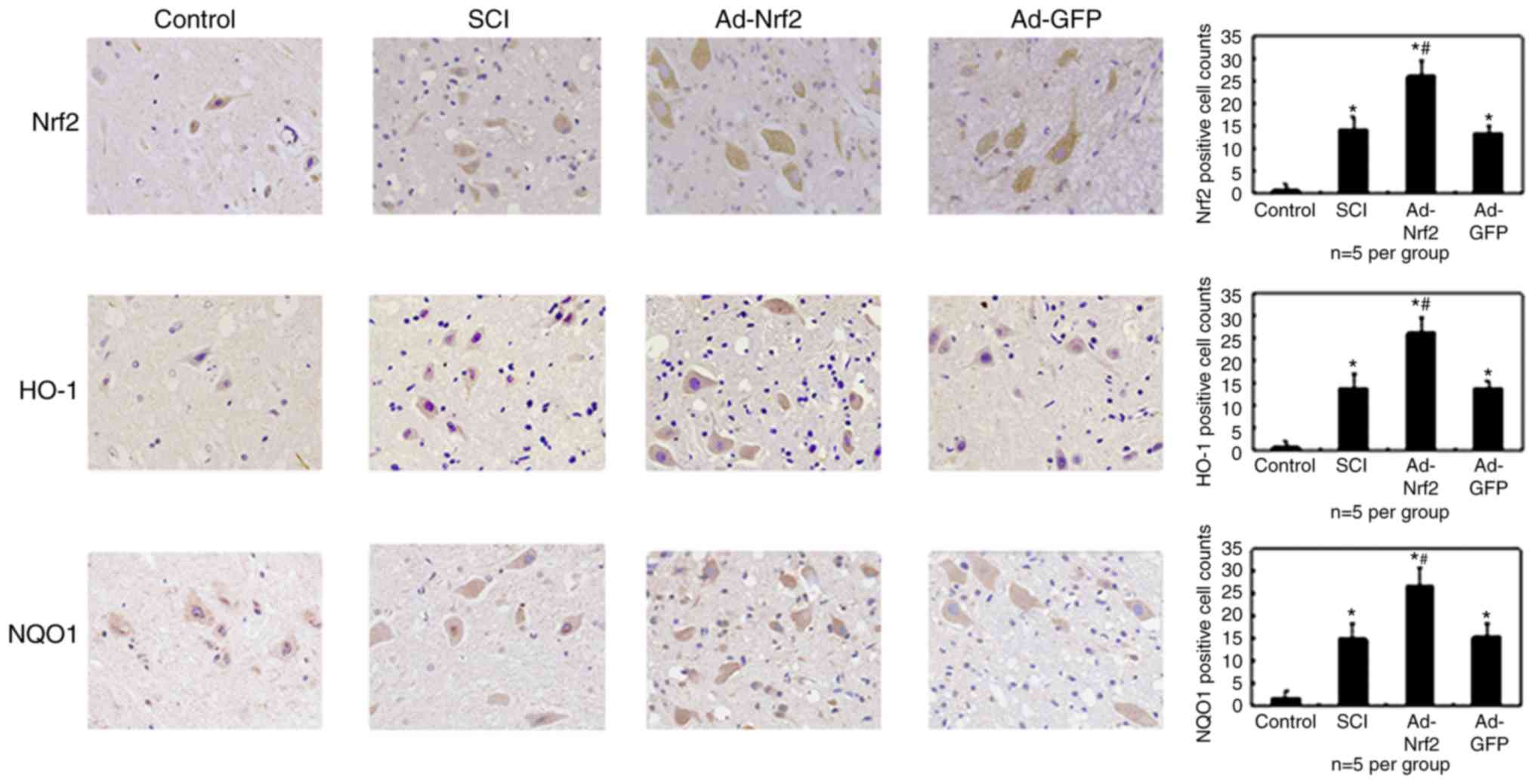 | Figure 4.Immunohistochemical staining for
Nrf2, HO-1 and NQO1 in the grey matter of the rats on the 3rd day
following the operation (magnification, ×400). Significantly
increased Nrf2, HO-1 and NQO1 immunoreactivities were present in
neurons/glial cells. In the Ad-Nrf2 group, the number of positive
cells increased significantly vs. the SCI and Ad-DFP groups. Bars
(mean ± standard error of the mean, n=5 in each group) presented
the number of the positive glial cells following SCI. *P<0.01
vs. the control group; #P<0.01 vs. the SCI and Ad-GFP
groups. GFP, green fluorescent protein; Ad, adenovirus; Nrf2,
nuclear factor erythroid 2p45-related factor 2; HO-1, heme
oxygenase-1; NQO1, NAD(P)H: quinone oxidoreductase-1; SCI, spinal
cord injury. |
Discussion
The leucine zipper transcription factor Nrf2 is a
major component of ARE-driven gene expression (33). Oligonucleotide microarray analysis
has indicated that Nrf2 is necessary in combating electrophiles and
ROS (23,34–37).
It serves a key function in protecting cells from oxidative stress.
Nrf2 may protect the liver from acetaminophen-induced injury
(38) and the lung from butylated
hydroxytoluene-induced toxicity (39). Cho et al (40) demonstrated that disruption of Nrf2
significantly enhanced pulmonary sensitivity and responsivity to
hyperoxic challenge. Compared with Nrf2+/+ astrocytes,
one previous study confirmed that Nrf2−/− primary
astrocytes are more susceptible to oxidative stress and
inflammation (37).
Nrf2+/+ astrocytes pretreated with t-butylhydroquinone
induce Nrf2 nuclear translocation, resulting in the coordinated
upregulation of ARE-driven genes and attenuation of
H2O2− and platelet-activating
factor-induced cell death (37).
PC12 cells (adrenal pheochromocytoma) was originally
isolated from tumors in the rat adrenal medulla in 1976 (41). They resemble the phenotype of
sympathetic ganglion neurons upon differentiation with nerve growth
factor (NGF) and may be subcultured indefinitely. The PC12 cell
line is traceable to a pheochromocytoma from the rat adrenal
medulla (42–45). It has been used as the classical
neuronal cell model due to its ability to acquire the features of
sympathetic neurons (46,47). PC12 cells have been used to
investigate the cellular mechanisms by which prion protein
fragments cause neuronal dysfunction (48), the nerve injury-induced neuropathic
pain model (49), the nitric
oxide-induced neurotoxicity model (50) and NGF inducing the differentiation
of PC12 cells by functioning through the tropomyosin receptor
kinase A receptor (51).
In the present study, the Ad-Nrf2 gene was
successfully transferred into PC12 cells and the spinal cord (by
local injection). Furthermore, the protein levels of nuclear Nrf2
and its regulated gene products, HO-1 and NQO1, were significantly
increased in PC12 cells compared with the control, P<0.01), and
the number of positive cells for Nrf2, HO-1 and NQO1 were promoted
in the neurons/glial cells of the spinal cord grey matter. The
function of the hind limb in SCI rats was significantly improved
following Ad-Nrf2 gene transfer compared with the SCI group
(P<0.05). According to these results, it was postulated that the
gene transfer of Nrf2 was able to alleviate SCI and promote the
functional recovery of the injured spinal cord. In view of the fact
that neurons are more susceptible than glial cells to oxidative
stress in Ad-Nrf2 group (23,52),
it may be considered that the Nrf2-ARE pathway may serve a
protective function in the pathological process of SCI.
Nrf2 mediates a group of cytoprotective enzymes. It
is believed to be the key regulator in CNS diseases by inducing the
expression of a group of antioxidant and detoxification enzymes
(26,53). In the present study, it was
revealed that SCI-induction significantly increased the number of
positive cells for Nrf2, HO-1 and NQO1 in grey matter neurons
compared with the control (P<0.01), suggesting that the Nrf2-ARE
pathway was activated. Such phenomena have additionally been
observed in TBI (54).
Increasingly, evidence suggests that a group of
cytoprotective enzymes mediated by the Nrf2-ARE pathway serve a
pivotal role in antioxidant, anti-inflammation and detoxification
functions, including HO-1 and NQO1 (55,56).
HO-1 produces biliverdin and reduces bilirubin to reduce ROS
production. The expression of ferritin (HO-1 dependent) may prevent
the conversion of H2O2 to hydroxyl radicals
by the Fenton reaction (57). NQO1
catalyzes the double-electron reduction and detoxification of
quinones and their derivatives, therefore protecting cells from the
harmful effects of quinones and their associated compounds
(58). In SCI, it is considered
important to reduce the neurons apoptosis or necrosis and maintain
the associations between neurons and axons. However, neurons have a
low antioxidant capacity and are highly sensitive to oxidative
stress (59). It is reported that
Nrf2-mediated neuroprotection is conferred primarily by glia
(23). SCI induces an increase in
oxidative stress and simultaneously causes glial dysfunction
(60). It is therefore important
to activate the remaining functional glia and neuronal
self-protection. The present study hypothesized that the
gene-transfer of Nrf2 may promote the Nrf2-ARE pathway to reduce
neuron necrosis or apoptosis, particularly by self-protection. The
results revealed that Ad-Nrf2 increased nuclear Nrf2, HO-1 and NQO1
expression in PC12 cells and neurons in the contusion site of SCI,
and hind limb functional recovery was also observed. Certain phase
II enzyme inducers (activating Nrf2), even fibroblast growth
factor-1, have demonstrated neuroprotective effects on motor neuron
survival in traumatic SCI (61–63).
Genetic ablation of the transcription repressor Bach1, a
transcriptional repressor of the HO-1 gene, may substantially
increase HO-1 expression and cytoprotection against SCI (64). The present study did not
successfully isolate the spinal cord neurons of adult rats. PC12 is
a tumor cell, which is still different from the neurons themselves.
This is the main limitation of the present experiment.
Therefore in conclusion, Nrf2 gene transfer may be a
direct method of protecting neurons/glial cells in SCI. Although
local injection may injure nerve cells, this method works and will
still benefit clinical treatment. Nrf2-adenovirus-mediated in
vivo gene transfer may promote functional recovery following
spinal cord contusion.
Acknowledgments
Not applicable.
Funding
The present study was supported by the Scientific
and Technological Research Program of Chongqing Municipal Education
Commission (grant no. KJ130315) and the Recruited Talent Supporting
Program of Yongchuan Hospital Affiliated Chongqing Medical
University (grant no. YJYJ20120004).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FCZ was responsible for the study design, literature
research, experiments, manuscript preparation, editing and review.
DMJ was responsible for the conception of the study and the
guarantor of integrity of the entire study. MHZ was responsible for
the definition of intellectual content and acquisition of data. BZ
was responsible for data acquisition. CH was responsible for data
analysis. JY was responsible for statistical analysis. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
All experimental procedures were performed in
accordance with the National Institutes of Health (NIH) Guide for
the Care and Use of Laboratory Animals (NIH publication no. 80-23,
revised 1996), and the number of animals used and their suffering
were minimized. Ethical approval was provided by the Yongchuan
Hospital of Chongqing Medical University (Chongqing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Oyinbo CA: Secondary injury mechanisms in
traumatic spinal cord injury: A nugget of this multiply cascade.
Acta Neurobiol Exp (Wars). 71:281–299. 2011.PubMed/NCBI
|
|
2
|
Gwak YS, Hulsebosch CE and Leem JW:
Neuronal-glial interactions maintain chronic neuropathic pain after
spinal cord injury. Neural Plast. 2017:24806892017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alizadeh A, Dyck SM and Karimi-Abdolrezaee
S: Traumatic spinal cord injury: An overview of pathophysiology,
models and acute injury mechanisms. Front Neurol. 10:2822019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bylicky MA, Mueller GP and Day RM:
Mechanisms of endogenous neuroprotective effects of astrocytes in
brain injury. Oxid Med Cell Longev. 2018:65010312018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kwon BK, Tetzlaff W, Grauer JN, Beiner J
and Vaccaro AR: Pathophysiology and pharmacologic treatment of
acute spinal cord injury. Spine J. 4:451–464. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kwon BK, Oxland TR and Tetzlaff W: Animal
models used in spinal cord regeneration research. Spine (Phila Pa
1976). 27:1504–1510. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lukácová N, Halát G, Chavko M and Marsala
J: Ischemia-reperfusion injury in the spinal cord of rabbits
strongly enhances lipid peroxidation and modifies phospholipid
profiles. Neurochem Res. 21:869–873. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Emery E, Aldana P, Bunge MB, Puckett W,
Srinivasan A, Keane RW, Bethea J and Levi AD: Apoptosis after
traumatic human spinal cord injury. J Neurosurg. 89:911–920. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Andrews NC, Erdjument-Bromage H, Davidson
MB, Tempst P and Orkin SH: Erythroid transcription factor NF-E2 is
a haematopoietic-specific basic-leucine zipper protein. Nature.
362:722–728. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Motohashi H, Katsuoka F, Engel JD and
Yamamoto M: Small Maf proteins serve as transcriptional cofactors
for keratinocyte differentiation in the Keap1-Nrf2 regulatory
pathway. Proc Natl Acad Sci USA. 101:6379–6384. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jain AK, Bloom DA and Jaiswal AK: Nuclear
import and export signals in control of Nrf2. J Biol Chem.
280:29158–29168. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Itoh K, Chiba T, Takahashi S, Ishii T,
Igarashi K, Katoh Y, Oyake T, Hayashi N, Satoh K, Hatayama I, et
al: An Nrf2/small Maf heterodimer mediates the induction of phase
II detoxifying enzyme genes through antioxidant response elements.
Biochem Biophys Res Commun. 236:313–322. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jazwa A and Cuadrado A: Targeting heme
oxygenase-1 for neuroprotection and neuroinflammation in
neurodegenerative diseases. Curr Drug Targets. 11:1517–1531. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Niture SK, Kaspar JW, Shen J and Jaiswal
AK: Nrf2 signaling and cell survival. Toxicol Appl Pharmacol.
244:37–42. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tanito M, Agbaga MP and Anderson RE:
Upregulation of thioredoxin system via Nrf2-antioxidant responsive
element pathway in adaptive-retinal neuroprotection in vivo and in
vitro. Free Radic Biol Med. 42:1838–1850. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Du H, Ma L, Chen G and Li S: The effects
of oxyresveratrol abrogates inflammation and oxidative stress in
rat model of spinal cord injury. Mol Med Rep. 17:4067–4073.
2018.PubMed/NCBI
|
|
17
|
Kaelan C, Jacobsen P, Morling P and
Kakulas BA: A quantitative study of motoneurons and cortico-spinal
fibers related to function in human spinal cord injury (SCI).
Paraplegia. 27:1531989.
|
|
18
|
Kakulas BA: The applied neuropathology of
human spinal cord injury. Spinal Cord. 37:79–88. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Blight AR: Cellular morphology of chronic
spinal cord injury in the cat: Analysis of myelinated axons by
line-sampling. Neuroscience. 10:521–543. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Eidelberg E, Straehley D, Erspamer R and
Watkins CJ: Relationship between residual hindlimb-assisted
locomotion and surviving axons after incomplete spinal cord
injuries. Exp Neurol. 56:312–322. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fehlings MG and Tator CH: The
relationships among the severity of spinal cord injury, residual
neurological function, axon counts, and counts of retrogradely
labeled neurons after experimental spinal cord injury. Exp Neurol.
132:220–228. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Delamarter RB and Coyle J: Acute
management of spinal cord injury. J Am Acad Orthop Surg. 7:166–175.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shih AY, Johnson DA, Wong G, Kraft AD,
Jiang L, Erb H, Johnson JA and Murphy TH: Coordinate regulation of
glutathione biosynthesis and release by Nrf2-expressing glia
potently protects neurons from oxidative stress. J Neurosci.
23:3394–3406. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu W, Chi L, Xu R, Ke Y, Luo C, Cai J, Qiu
M, Gozal D and Liu R: Increased production of reactive oxygen
species contributes to motor neuron death in a compression mouse
model of spinal cord injury. Spinal Cord. 43:204–213. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Adibhatla RM and Hatcher JF: Lipid
oxidation and peroxidation in CNS health and disease: From
molecular mechanisms to therapeutic opportunities. Antioxid Redox
Signal. 12:125–169. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hong Y, Yan W, Chen S, Sun CR and Zhang
JM: The role of Nrf2 signaling in the regulation of antioxidants
and detoxifying enzymes after traumatic brain injury in rats and
mice. Acta Pharmacol Sin. 31:1421–1430. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jin W, Wang H, Yan W, Zhu L, Hu Z, Ding Y
and Tang K: Role of Nrf2 in protection against traumatic brain
injury in mice. J Neurotrauma. 26:131–139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jin W, Wang H, Yan W, Xu L, Wang X, Zhao
X, Yang X, Chen G and Ji Y: Disruption of Nrf2 enhances
upregulation of nuclear factor-kappaB activity, proinflammatory
cytokines, and intercellular adhesion molecule-1 in the brain after
traumatic brain injury. Mediators Inflamm. 2008:7251742008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mao L, Wang H, Qiao L and Wang X:
Disruption of Nrf2 enhances the upregulation of nuclear
factor-kappaB activity, tumor necrosis factor-α, and matrix
metalloproteinase-9 after spinal cord injury in mice. Mediators
Inflamm. 2010:2383212010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kay MA: State-of-the-art gene-based
therapies: The road ahead. Nat Rev Genet. 12:316–328. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tasdemiroglu E and Tibbs PA: Long-term
follow-up results of thoracolumbar fractures after posterior
instrumentation. Spine (Phila Pa 1976). 20:1704–1708. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
testing in rats. J Neurotrauma. 12:1–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tonelli C, Chio IIC and Tuveson DA:
Transcriptional regulation by Nrf2. Antioxid Redox Signal.
29:1727–1745. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Moi P, Chan K, Asunis I, Cao A and Kan YW:
Isolation of NF-E2-related factor 2 (Nrf2), a NF-E2-like basic
leucine zipper transcriptional activator that binds to the tandem
NF-E2/AP1 repeat of the beta-globin locus control region. Proc Natl
Acad Sci USA. 91:9926–9930. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fan Z, Wirth AK, Chen D, Wruck CJ, Rauh M,
Buchfelder M and Savaskan N: Nrf2-Keap1 pathway promotes cell
proliferation and diminishes ferroptosis. Oncogenesis. 6:e3712017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee JM, Calkins MJ, Chan K, Kan YW and
Johnson JA: Identification of the NF-E2-related factor-2-dependent
genes conferring protection against oxidative stress in primary
cortical astrocytes using oligonucleotide microarray analysis. J
Biol Chem. 278:12029–12038. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Thimmulappa RK, Mai KH, Srisuma S, Kensler
TW, Yamamoto M and Biswal S: Identification of Nrf2-regulated genes
induced by the chemopreventive agent sulforaphane by
oligonucleotide microarray. Cancer Res. 62:5196–5203.
2002.PubMed/NCBI
|
|
38
|
Chan K, Han XD and Kan YW: An important
function of Nrf2 in combating oxidative stress: Detoxification of
acetaminophen. Proc Natl Acad Sci USA. 98:4611–4616. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chan K and Kan YW: Nrf2 is essential for
protection against acute pulmonary injury in mice. Proc Natl Acad
Sci USA. 96:12731–12736. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cho HY, Jedlicka AE, Reddy SP, Kensler TW,
Yamamoto M, Zhang LY and Kleeberger SR: Role of NRF2 in protection
against hyperoxic lung injury in mice. Am J Respir Cell Mol Biol.
26:175–182. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Greene LA and Tischler AS: Establishment
of a noradrenergic clonal line of rat adrenal pheochromocytoma
cells which respond to nerve growth factor. Proc Natl Acad Sci USA.
73:2424–2428. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Uceda G, Artalejo AR, López MG, Abad F,
Neher E and García AG: Ca(2+)-activated K+ channels modulate
muscarinic secretion in cat chromaffin cells. J Physiol.
454:213–230. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhou Z and Neher E: Calcium permeability
of nicotinic acetylcholine receptor channels in bovine adrenal
chromaffin cells. Pflugers Arch. 425:511–517. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Nooney JM, Peters JA and Lambert JJ: A
patch clamp study of the nicotinic acetylcholine receptor of bovine
adrenomedullary chromaffin cells in culture. J Physiol.
455:503–527. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Horrigan FT and Bookman RJ: Releasable
pools and the kinetics of exocytosis in adrenal chromaffin cells.
Neuron. 13:1119–1129. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Westerink RH and Ewing AG: The PC12 cell
as model for neurosecretion. Acta Physiol (Oxf). 192:273–285. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hu R, Cao Q, Sun Z, Chen J, Zheng Q and
Xiao F: A novel method of neural differentiation of PC12 cells by
using Opti-MEM as a basic induction medium. Int J Mol Med.
41:195–201. 2018.PubMed/NCBI
|
|
48
|
Taylor SC, Green KN, Smith IF and Peers C:
Prion protein fragment 106–126 potentiates catecholamine secretion
from PC-12 cells. Am J Physiol Cell Physiol. 281:C1850–C1857. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shao J, Cao J, Wang J, Ren X, Su S, Li M,
Li Z, Zhao Q and Zang W: MicroRNA-30b regulates expression of the
sodium channel Nav1.7 in nerve injury-induced neuropathic pain in
the rat. Mol Pain. 12:2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zheng W, Chong CM, Wang H, Zhou X, Zhang
L, Wang R, Meng Q, Lazarovici P and Fang J: Artemisinin conferred
ERK mediated neuroprotection to PC12 cells and cortical neurons
exposed to sodium nitroprusside-induced oxidative insult. Free
Radic Biol Med. 97:158–167. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liu L, Sun T, Xin F, Cui W, Guo J and Hu
J: Nerve growth factor protects against alcohol-induced
neurotoxicity in PC12 cells via I3K/Akt/mTOR pathway. Alcohol
Alcohol. 52:12–18. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Gilgun-Sherki Y, Melamed E and Offen D:
Oxidative stress induced-neurodegenerative diseases: The need for
antioxidants that penetrate the blood brain barrier.
Neuropharmacology. 40:959–975. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
van Muiswinkel FL and Kuiperij HB: The
Nrf2-ARE signalling pathway: Promising drug target to combat
oxidative stress in neurodegenerative disorders. Curr Drug Targets
CNS Neurol Disord. 4:267–281. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yan W, Wang HD, Hu ZG, Wang QF and Yin HX:
Activation of Nrf2-ARE pathway in brain after traumatic brain
injury. Neurosci Lett. 431:150–154. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Reisman SA, Buckley DB, Tanaka Y and
Klaassen CD: CDDO-Im protects from acetaminophen hepatotoxicity
through induction of Nrf2-dependent genes. Toxicol Appl Pharmacol.
236:109–114. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang Y, Guan L, Wang X, Wen T, Xing J and
Zhao J: Protection of chlorophyllin against oxidative damage by
inducing HO-1 and NQO1 expression mediated by PI3K/Akt and Nrf2.
Free Radic Res. 42:362–371. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Jansen T and Daiber A: Direct antioxidant
properties of bilirubin and biliverdin. Is there a role for
biliverdin reductase? Front Pharmacol. 3:302012.
|
|
58
|
Gaikwad A, Long DJ II, Stringer JL and
Jaiswal AK: In vivo role of NAD(P)H: Quinone oxidoreductase 1
(NQO1) in the regulation of intracellular redox state and
accumulation of abdominal adipose tissue. J Biol Chem.
276:22559–22564. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen X, Guo C and Kong J: Oxidative stress
in neurodegenerative diseases. Neural Regen Res. 7:376–385.
2012.PubMed/NCBI
|
|
60
|
Santos-Nogueira E, López-Serrano C,
Hernández J, Lago N, Astudillo AM, Balsinde J, Estivill-Torrús G,
de Fonseca FR, Chun J and López-Vales R: Activation of
lysophosphatidic acid receptor type 1 contributes to
pathophysiology of spinal cord injury. J Neurosci. 35:10224–10235.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Liu XY, Li CY, Bu H, Li Z, Li B, Sun MM,
Guo YS, Zhang L, Ren WB, Fan ZL, et al: The neuroprotective
potential of phase II enzyme inducer on motor neuron survival in
traumatic spinal cord injury in vitro. Cell Mol Neurobiol.
28:769–779. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Sun MM, Bu H, Li B, Yu JX, Guo YS and Li
CY: Neuroprotective potential of phase II enzyme inducer diallyl
trisulfide. Neurol Res. 31:23–27. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Vargas MR, Pehar M, Cassina P,
Martínez-Palma L, Thompson JA, Beckman JS and Barbeito L:
Fibroblast growth factor-1 induces heme oxygenase-1 via nuclear
factor erythroid 2-related factor 2 (Nrf2) in spinal cord
astrocytes: Consequences for motor neuron survival. J Biol Chem.
280:25571–25579. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kanno H, Ozawa H, Dohi Y, Sekiguchi A,
Igarashi K and Itoi E: Genetic ablation of transcription repressor
Bach1 reduces neural tissue damage and improves locomotor function
after spinal cord injury in mice. J Neurotrauma. 26:31–39. 2009.
View Article : Google Scholar : PubMed/NCBI
|