Introduction
Ovarian cancer remains one of the most aggressive
and highly recurrent malignant diseases worldwide with a poor
prognosis (1,2). The vague clinical symptoms in the
early stages of ovarian cancer have a major role in delaying
intervention and treatment (3).
Several diagnostic biomarkers have been identified for ovarian
cancer. Serum human epididymis protein 4 (HE4) and transthyretin
(TTR) were identified as novel biomarkers for ovarian cancer
(4). Compared with the golden
standard marker mucin-16 (CA125), HE4 is suitable for patients with
advanced ovarian cancer, while TTR is a better serum biomarker for
patients with early stage ovarian cancer (4). A panel of four serum biomarkers
(CA125, HE4, E-cadherin and interleukin-6) has been shown to have a
sensitivity of 95–100% for patients with early stage ovarian cancer
(5). However, unlike the
progression in the discovery of diagnostic biomarkers for ovarian
cancer, biomarkers associated with more efficient therapeutic
targets remain elusive.
Sialylation is one of the essential molecular
post-translational modifications, which has important roles in
metabolism, immunity, development and cancer biology (6–9). In
ovarian cancer, sialylation of glycoproteins is a common
modification in ovarian cancer proximal fluids (10). Aberrantly sialylated N-linked
glycopeptides may serve as serum biomarkers for patients with
ovarian cancer (11–13). The fully sialylated α-chain of
complement 4-binding protein has been identified as a diagnostic
and prognostic marker for ovarian cancer (14,15).
β-galactoside-α2,3-sialyltransferase I (ST3Gal1) was found to be
expressed at a higher level in the advanced stage of epithelial
ovarian cancer, and was demonstrated to facilitate epidermal growth
factor receptor (EGFR) signaling and the migration and peritoneal
dissemination of ovarian cancer cells (16). The α2,6 N-linked sialylation of the
β1 integrins was reported to promote cell adhesion and invasion of
ovarian cancer cells (17). In
addition, the volume of ascites indicated the occurrence of
transmesothelial invasion, which correlated with a poorer prognosis
for patients with ovarian cancer (18,19).
It has recently been reported that there is a positive correlation
between the volume of ascites and the amount of serum sialylated
structures in patients with epithelial ovarian cancer (8). Therefore, sialyltransferase-catalyzed
sialylations are a prevalent and aggravating factor in ovarian
cancer initiation and progression.
The treatment of ovarian cancer includes surgery,
chemotherapy and immunotherapy (20). Due to the vague clinic symptoms in
the early stages of ovarian cancer, most patients with ovarian
cancer are identified when the malignancy becomes advanced, which
reduces the availability of surgical intervention (21,22).
The first line chemotherapies used for ovarian cancer are
predominantly platinum-based and taxane-based drugs (22,23).
Surgery in combination with chemotherapy or chemotherapy alone
remain the standard care for advanced ovarian cancer, although most
patients with advanced ovarian cancer develop chemoresistance,
metastasis and bowel obstructions, which are the most frequent
causes of mortality (22). The
molecular mechanisms of chemoresistance are complex, including the
maintenance of cancer stem cells, the aberrant activation of
multi-drug resistant pathways, the aberrant activation of ABC
transporters and oncogenic mutation (24–27).
HE4 not only serves as a diagnostic marker for ovarian cancer, but
is also a predictor of platinum sensitivity in ovarian cancer
(28). α2,6 N-linked sialylated
EGFR confers acquired resistance to gefitinib in ovarian cancer
(29). ST6GAL1 confers cisplatin
resistance in ovarian tumor cells (30). Our previous study showed that
ST3Gal3 correlated with cisplatin resistance in ovarian tumor cells
(31). In the present study, the
relationship between ST3Gal3 expression and paclitaxel resistance
in ovarian tumor cells was investigated in order to provide a
better understanding of the effect of paclitaxel treatment alone or
in combination with ST3Gal3.
Materials and methods
Cell culture
The human ovarian cell line SKOV3 was purchased from
the American Type Culture Collection and the HO8910PM cell line was
purchased from The Cell Bank of Type Culture Collection of The
Chinese Academy of Sciences. SKOV3 cells were cultured in RPMI-1640
medium (Gibco; Thermo Fisher Scientific, Inc) containing 10% FBS
(Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Gibco; Thermo Fisher Scientific, Inc.).
HO8910PM cells were cultured in DMEM high-glucose medium (Gibco;
Thermo Fisher Scientific, Inc.) containing 10% FBS and 1%
penicillin-streptomycin. All cell lines were cultured at 37°C in a
humidified incubator with 5% CO2.
RNA isolation and reverse
transcription-quantitative (RT-q) PCR
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions. Total RNA was reverse transcribed
using TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix
(Beijing Transgen Biotech Co., Ltd.). Complementary DNA was
amplified using UltraSYBR Mixture (High ROX; CWBio) and the
following primers: ST3Gal3 forward, 5′-AAAACGACACTGCGCATCAC-3′ and
reverse, 5′-TCGAGTGGCCACAGATTTCC-3′; and GAPDH forward,
5′-AGCCTCAAGATCATCAGC-3′ and reverse 5′-GAGTCCTTCCACGATACC-3′. The
qPCR cycling conditions were as follows: 95°C for 30 sec, followed
by 40 cycles of 95°C for 5 sec and 60°C for 40 sec. The relative
levels of mRNA expression were normalized to GAPDH and calculated
using the 2−ΔΔCq method (32).
Cell Counting Kit-8 (CCK-8)
assays
SKOV3 and HO8910PM cells were seeded in 96-well
plates at a density of 5×103 cells/well. The next day,
the cells were treated with 0, 5, 10, 20, 40, 80, 160 or 320 ng/ml
paclitaxel (Sigma-Aldrich; Merck KGaA) or the equivalent volume of
DMSO as a negative control. After 48 h of incubation, the cell
viability was determined using the CCK-8 assay (Beyotime Institute
of Biotechnology), according to the manufacturer's instructions.
Cytotoxicity was calculated as follows: Cytotoxicity
(%)=[1-(optical density of tested cells)/(optical density of
control cells)] ×100. The IC50 (half maximal inhibitory
concentration) value was calculated by GraphPad Prism 6 (GraphPad
Software, Inc.).
Small interfering (si)RNA transfection
and paclitaxel treatment
In total, three ST3Gal3 siRNA sequences were
designed and synthesized by Guangzhou RiboBio Co., Ltd. The siRNA
sequences for siCTRL was: 5′-TTCTCCGAACGTGTCACGT-3′. The siRNA
sequences for ST3Gal3 were: 1#-sense 5′-CGTGGAAGCTACACTTACT-3′,
2#-sense 5′-CCTGAATCTGGACTCTAAA-3′ and 3#-sense
5′-CCTGGACGCACAATATCCA-3′. These siRNA sequences were tested in a
previous study (31), and the 1#
siRNA sequence was then used for further experiments. Briefly,
cells were seeded into 6-well plates at a density of
2×105 cells/well. The next day, 7.5 µl
Lipofectamine® RNAiMAX reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was diluted in 125 µl Opti-MEM
(Invitrogen; Thermo Fisher Scientific, Inc.) and 30 pmol siRNA was
diluted in 125 µl Opti-MEM, and each was mixed by vortexing for 10
sec. The diluted siRNA was added to the diluted
Lipofectamine® RNAiMAX Reagent and incubated for 10 min
at room temperature. During the incubation, the cells were washed
once with 3 ml of PBS and 2 ml fresh growth medium was added.
Subsequently, the 250 µl transfection mixture was added dropwise
onto the cells in the 6-well plate, incubated for two days and then
exposed to 20 ng/ml paclitaxel for a further 48 h.
Western blot
Cells were collected and lysed on ice using RIPA
lysis buffer (Beyotime Institute of Biotechnology) containing 1%
PMSF (Beyotime Institute of Biotechnology). The protein
concentration was determined using the bicinchoninic acid method.
In total, 50 µg of total protein was separated using 10% SDS-PAGE
and transferred onto PVDF membranes. After 5% bovine serum albumin
(cat. no. ST023; Beyotime Institute of Biotechnology) blocking at
room temperature for 1 h, membranes were incubated overnight at 4°C
with primary antibodies. The primary antibodies used were:
anti-ST3Gal3 (1:500 dilution; cat. no. SC-134040; Santa Cruz
Biotechnology, Inc.), anti-caspase-8 (1:1,000 dilution; cat. no.
4790; Cell Signaling Technologies, Inc.), anti-caspase-3 (1:1,000
dilution; cat. no. 14220; Cell Signaling Technologies, Inc.) and
anti-GAPDH (1:2,000 dilution; cat. no. AF0006; Beyotime Institute
of Biotechnology). Membranes were then incubated with secondary
anti-rabbit or anti-mouse horseradish peroxidase (HRP)-conjugated
secondary antibodies (1:3,000 dilution; cat. nos. A0208 and A0216;
Beyotime Institute of Biotechnology) for 1 h at room temperature.
Results were acquired using the Gel Logic 1500 imaging system
(Kodak). GAPDH was used as a loading control.
Flow cytometry for Maackia amurensis
lectin II (MAL II) staining
Briefly, after the medium was discarded, cells were
washed twice with PBS and digested with trypsin/0.25% EDTA (Gibco;
Thermo Fisher Scientific, Inc.). Fresh growth medium was added to
terminate the digestion and cells were collected by centrifugation
at 1,000 × g for 5 min at room temperature. The cells were
resuspended and washed twice in 1 ml PBS. The cell pellets were
resuspended in 100 µl of HEPES containing 0.5% BSA and 2.5 µg/ml
biotinylated MALII (Vector Laboratories, Ltd.), and incubated at
room temperature for 2 h in the dark. The cells were washed twice
with PBS and incubated with 1 µg/ml streptavidin-phycoerythrin
(Sigma-Aldrich; Merck KGaA) for 1 h at room temperature in the
dark. After washing the stained cells twice with PBS, cell surface
MAL II was quantified using a FACSCanto II flow cytometer (BD
Biosciences), and analyzed by BD CellQuest Software version 3.3 (BD
Biosciences) according to the manufacturer's instructions.
Flow cytometry for apoptosis
analysis
Cells were digested and collected as aforementioned.
Cells were resuspended and briefly washed twice in 1ml PBS. The
final pellets were resuspended in 500 µl of 1X binding buffer, and
5 µl of Annexin-V-FITC (BD Biosciences) was added and incubated at
room temperature for 15 min in the dark. Then 5 µl of propidium
iodide (BD Biosciences) was added to the cells and incubated at
room temperature for 5 min in the dark. Apoptosis was immediately
quantified using the FACSCanto II flow cytometer (BD Biosciences),
and analyzed by BD CellQuest Software version 3.3 (BD
Biosciences).
TUNEL assay
In brief, cells were fixed with 4% paraformaldehyde
at room temperature for 20 min and washed twice with PBS. Cells
were then permeabilized with 0.3% Triton X-100 at room temperature
for 10 min, washed once with PBS and incubated with 0.3%
H2O2 in PBS for a further 20 min in the dark.
After washing twice with PBS, 50 µl TUNEL working solution
(Beyotime Institute of Biotechnology) was added to each sample and
incubated at room temperature for 1 h. After washing twice with
PBS, stop solution was added at room temperature for 10 min. After
washing twice with PBS, streptavidin-HRP working solution was added
to the cells at room temperature for 30 min. After washing twice
with PBS, 3,3′diaminobenzidine solution was dropped onto the
samples and incubated at room temperature for 3 min. After washing
with PBS and mounting with neutral balsam, images were captured
using a Leica optical microscope (Leica Microsystems, Inc.) and
routine light microscopy (magnification, ×100).
Statistical analysis
Experiments were independently performed for a
minimum of three times and results were qualitatively similar.
Representative experiments are shown. Numerical data are presented
as the mean ± SD. Statistical analyses were performed using one-way
ANOVA and the Tukey-Kramer multiple comparisons test using GraphPad
Prism 6 (GraphPad Software, Inc.). P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression and activity of ST3Gal3 in
ovarian cancer cell lines
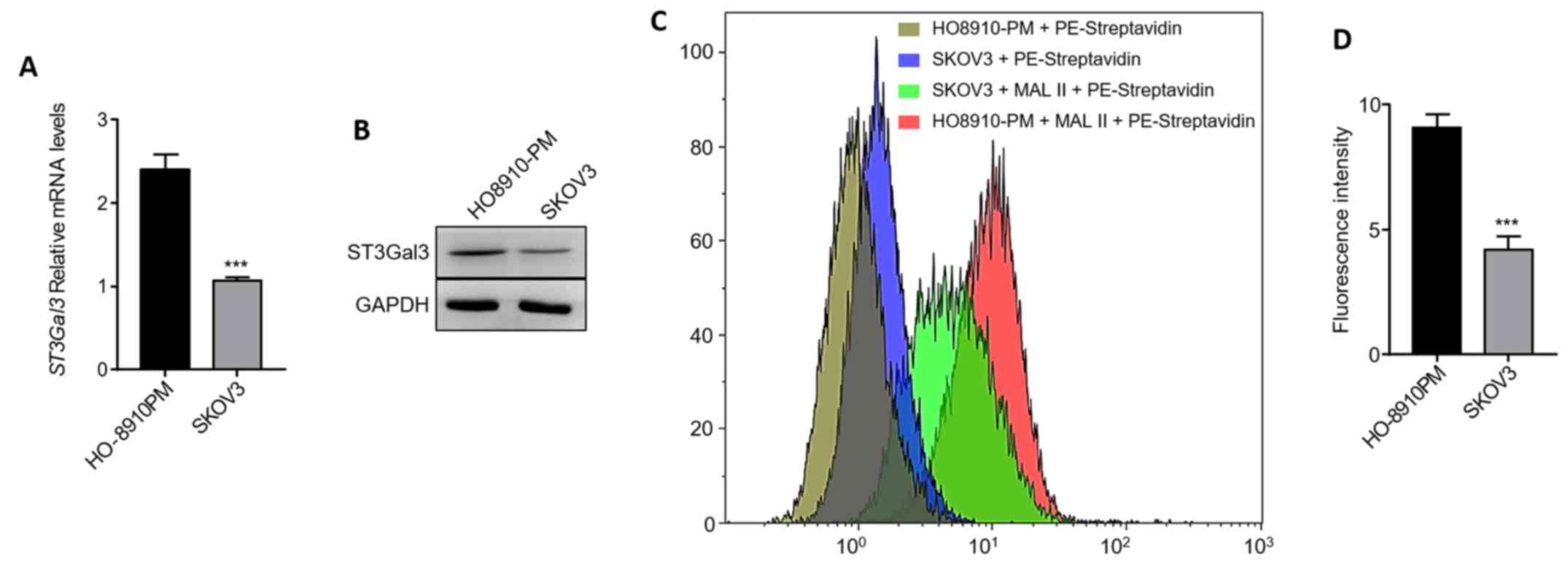
To identify the relationship between ST3GAL3 and MAL
II in ovarian cancer cell lines, the mRNA expression levels of
ST3GAL3 were investigated. The results showed that the levels of
ST3Gal3 mRNA expression in HO8910PM cells was more than double that
in SKOV3 cells (Fig. 1A). The
protein expression levels of ST3Gal3 were also markedly higher in
HO8910PM cells than in SKOV3 cells (Fig. 1B). MAL II binds to sialic acid in
an α-2,3 linkage rather than an α-2,6 linkage, which is specific
for the activity of ST3Gal3 (33).
Biotinylated MAL II was used to investigate the levels of cell
surface MAL II in ovarian cancer cell lines. In line with the
expression levels of ST3Gal3, the α-2,3 sialic acid linked terminal
glycosylated modification was ~2-fold higher in HO8910PM cells than
in SKOV3 cells (Fig. 1C and D).
Collectively, the expression and activity of ST3Gal3 were
significantly higher in HO8910PM cells than in SKOV3 cells.
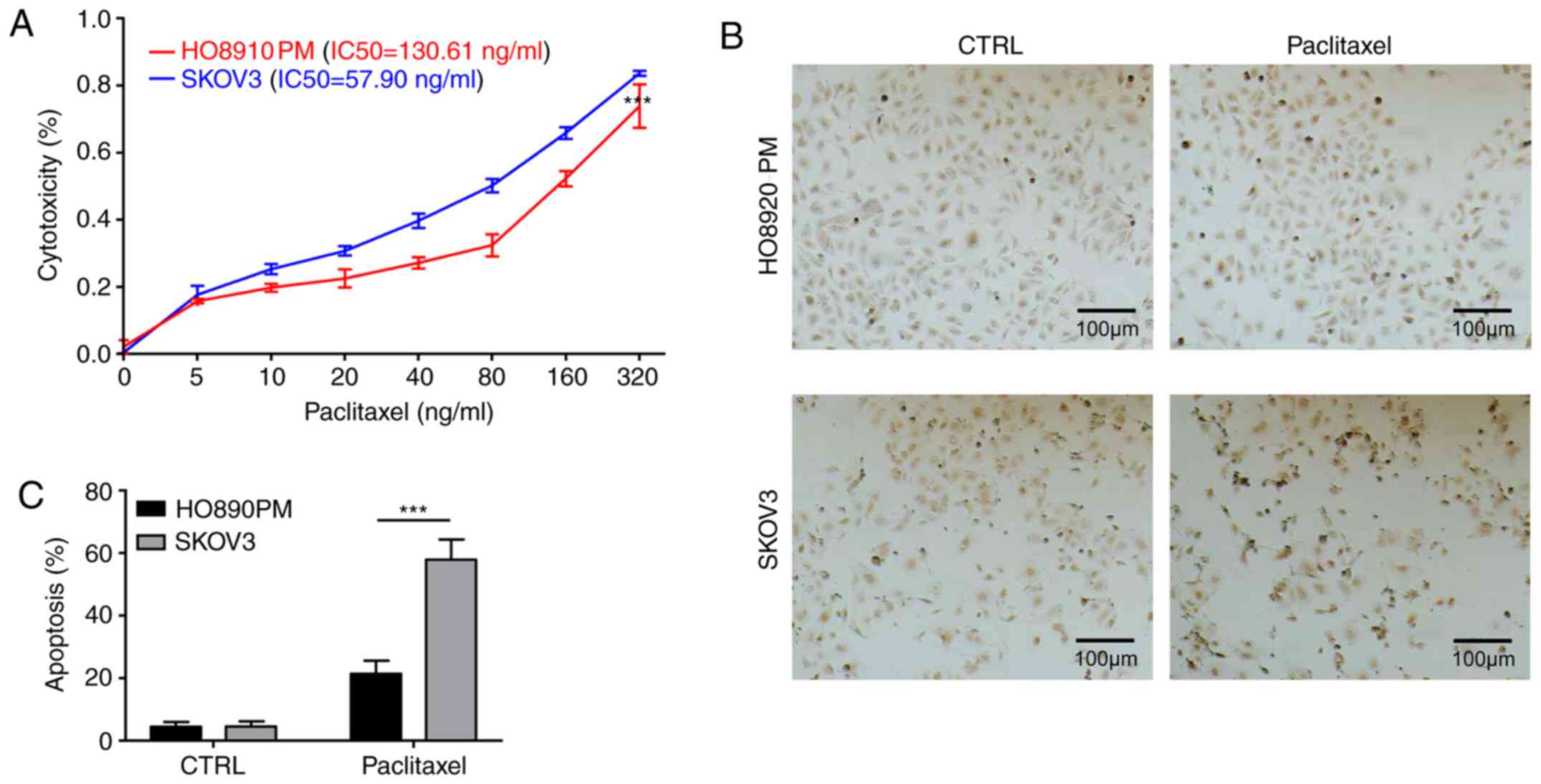
ST3Gal3 affects paclitaxel-resistance
in ovarian cancer cells
The aberrant expression and activity of ST3Gal3 in
ovarian cancer cells prompted an investigation into the
relationship between the expression of ST3Gal3 and
paclitaxel-resistance. Paclitaxel treatments ranging from 0 to 320
ng/ml were used to induce cytotoxicity in SKOV3 and HO8910PM cells.
The IC50 of paclitaxel in SKOV3 cells and HO8910PM cells
were calculated to be 57.90 and 130.61 ng/ml, respectively,
indicating that the paclitaxel-resistance in HO8910PM cells was
>2-fold higher than in SKOV3 cells (Fig. 2A). TUNEL assays were then used to
examine paclitaxel-induced apoptosis in SKOV3 and HO8910PM cells.
The results revealed that the apoptotic ratio was ~2-fold higher in
SKOV3 cells than in HO8910PM cells following 48 h exposure to 50
ng/ml paclitaxel (Fig. 2B and C),
indicating that paclitaxel-resistance was higher in HO8910PM cells
than in SKOV3 cells.
ST3Gal3 knockdown sensitizes ovarian
cancer cells to paclitaxel-induced apoptosis
To further investigate the role of ST3Gal3 in
paclitaxel resistance, ST3Gal3 was silenced. siRNA was used to
knockdown ST3Gal3 expression in SKOV3 and HO8910PM cells, followed
by treatment with 20 ng/ml paclitaxel for 48 h. The level of
apoptosis was then quantified using flow cytometry. In HO8910PM
cells, the knockdown of ST3GAL3 significantly increased the level
of apoptosis, (si-CTRL vs. si-ST3Gal3; Fig. 3A and B). The apoptosis ratio was
~3-fold higher in si-CTRL knockdown cells treated with paclitaxel
than in si-CTRL knockdown cells without paclitaxel treatment. The
apoptosis ratio was ~2-fold higher in the si-ST3Gal3 cells compared
with the si-CTRL cells after treated with paclitaxel (Fig. 3A and B). In SKOV3 cells, the
knockdown of ST3Gal3 also increased the level of apoptosis, but not
significantly (si-CTRL vs. si-ST3Gal3). The apoptosis ratio was
~1.5-fold higher in the si-ST3Gal3 cells plus paclitaxel compared
with the si-CTRL cells plus paclitaxel (Fig. 3C and D). In the absence of
STG3Gal3, HO8910PM cells are no longer significantly more resistant
to paclitaxel than SKOV3 cells. (Fig.
3E). ST3Gal3 knockdown was found to increase
paclitaxel-chemosensitivity in both SKOV3 and HO8910PM cells.
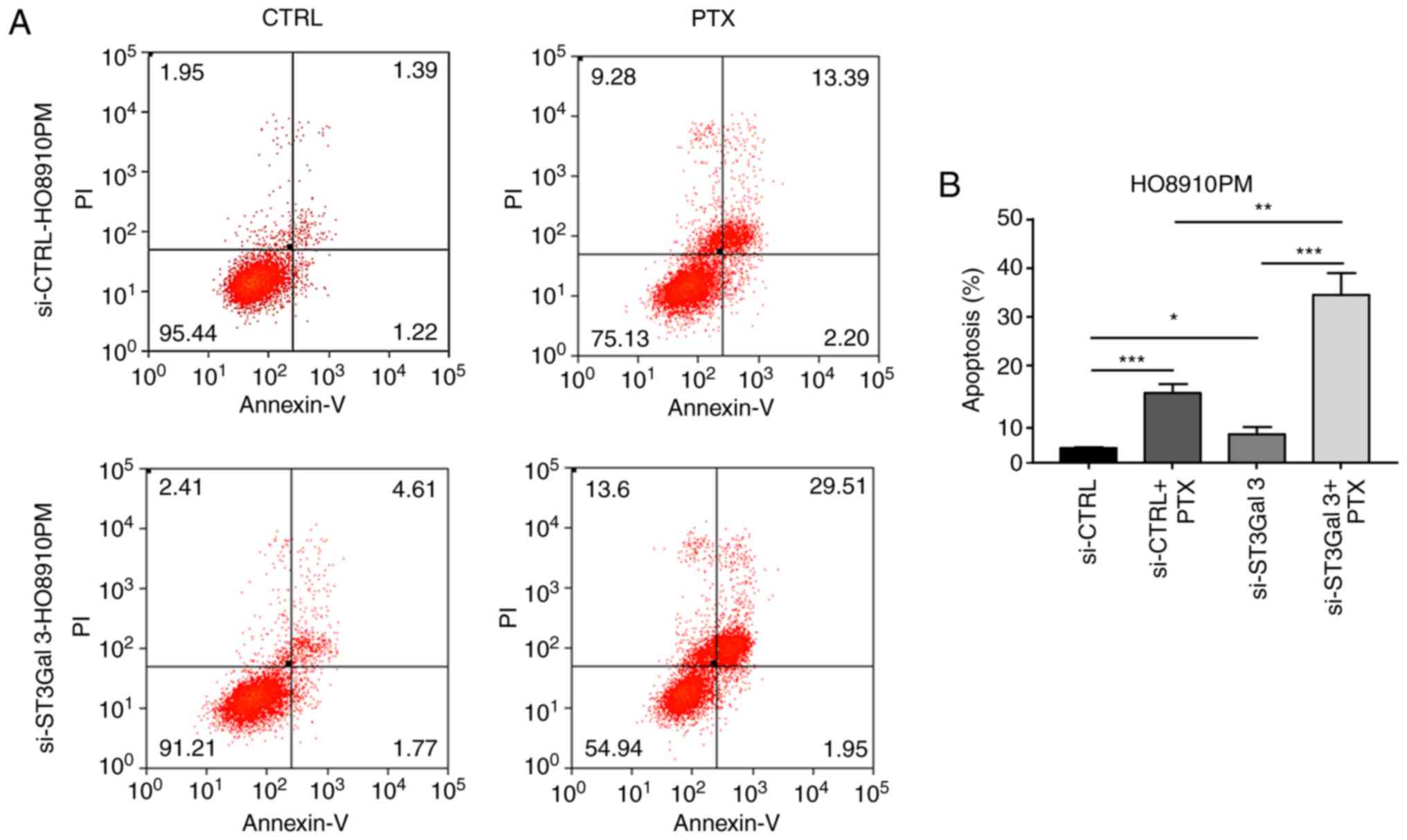 | Figure 3.ST3Gal3 knockdown sensitizes ovarian
cancer cells to PTX-induced apoptosis. (A) After transfection of
si-ST3Gal3 into HO8910PM cells and treatment with or without 20
ng/ml PTX for 48 h, flow cytometry was performed to determine the
rates of apoptosis. (B) Quantification of apoptosis rates in
HO8910PM cells. (C) After transfection of si-ST3GAL3 into SKOV3
cells and treatment with or without 20 ng/ml PTX for next 48 h,
flow cytometry was performed to determine the rates of apoptosis.
(D) Quantification of apoptosis rates in SKOV3 cells. (E)
Comparison of the apoptosis in HO8910PM and SKOV3 cells treated
with PTX. *P<0.05, **P<0.01, ***P<0.001. ST3Gal3,
β-galactoside-α2,3-sialyltransferase III; si, small interfering;
PTX, paclitaxel; CTRL, control; PI, propidium iodide. |
ST3Gal3 knockdown increases
paclitaxel-mediated activation of caspase-8/3 signaling
The mechanism of ST3Gal3-induced chemoresistance was
investigated next. RT-qPCR and western blot results first confirmed
that the si-ST3Gal3 siRNA significantly downregulated STG3Gal3 mRNA
and protein levels in both HO8910PM and SKOV3 cells (Fig. 4). Paclitaxel treatment induced
ST3GAL3 expression at the mRNA and protein level in HO8910PM cells
(Fig. 4A and B). After the
siRNA-induced downregulation of ST3Gal3, paclitaxel treatment
further decreased the levels of ST3Gal3 mRNA in HO8910PM cells,
rather than rescuing STG3Gal3 expression (Fig. 4A). The change in the protein level
was similar (Fig. 4B). In
addition, it was found that paclitaxel increased caspase-8 and
caspase-3 protein expression levels in si-CTRL HO8910PM cells.
Knockdown of ST3Gal3 alone only induced caspase-3 protein levels in
HO8910PM cells compared with si-CTRL, while caspase-8 levels were
not altered (Fig. 4B).
Furthermore, si-ST3Gal3 knockdown plus paclitaxel treatment
markedly elevated caspase-8 and caspase-3 protein levels in
HO8910PM cells compared with si-ST3Gal3 knockdown alone (Fig. 4B).
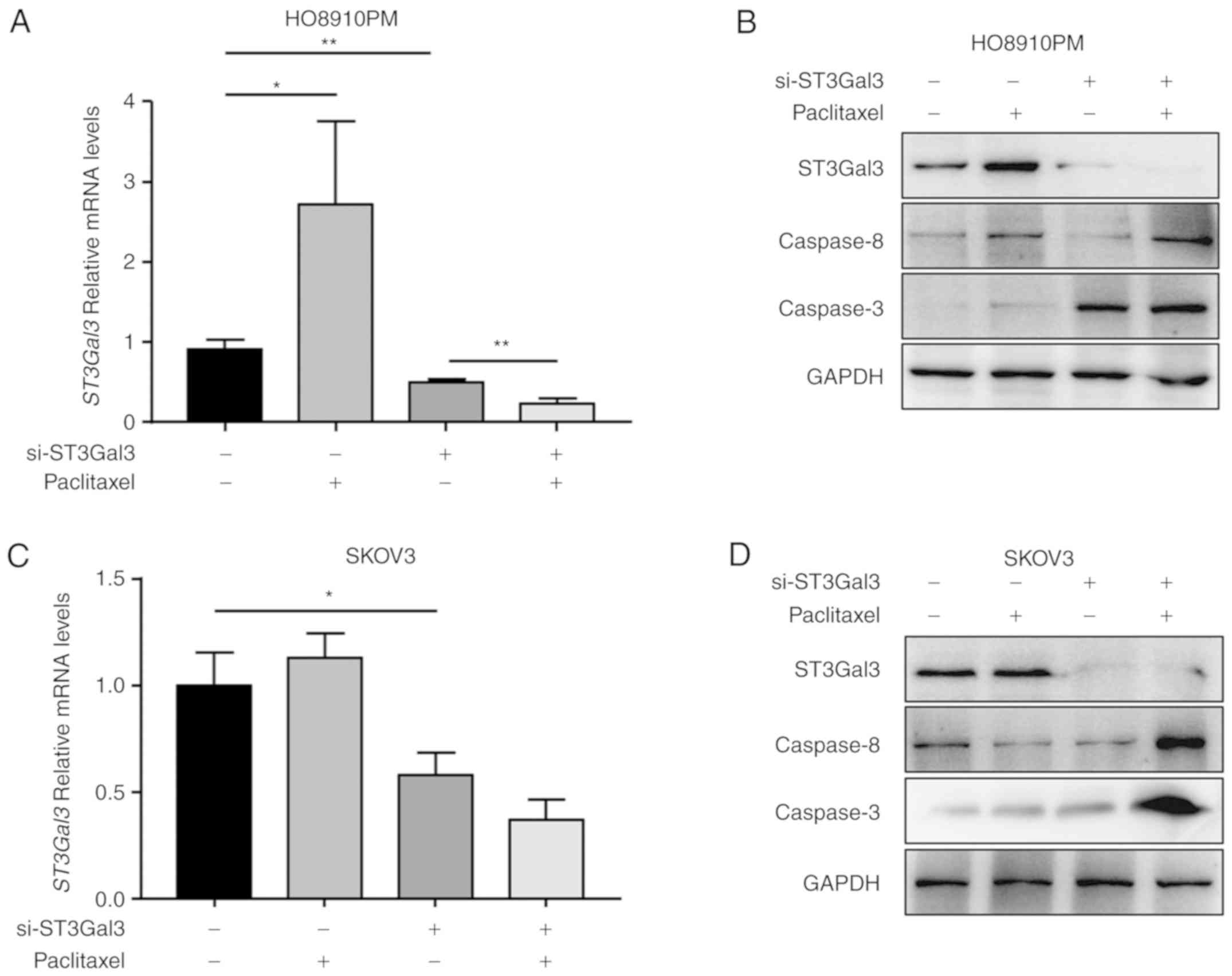 | Figure 4.ST3Gal3 knockdown synergistically
facilitates the paclitaxel-mediated activation of caspase-8/3. (A)
Following the indicated treatments, RT-qPCR was used to determine
the mRNA levels of ST3Gal3 in HO8910PM cells. (B) After the
indicated treatments, western blot analysis was performed to
determine the protein expression levels of ST3Gal3, caspase-3 and
caspase-8 in HO8910-PM cells. GAPDH served as the loading control.
(C) Following the indicated treatments, RT-qPCR was used to
determine the mRNA levels of ST3Gal3 in SKOV3 cells. (D) After the
indicated treatments, western blot analysis was performed to
determine the protein expression levels of ST3Gal3, caspase-3 and
caspase-8 in SKOV3 cells. GAPDH served as the loading control.
*P<0.05 and **P<0.01, with comparisons indicated by lines.
ST3Gal3, β-galactoside-α2,3-sialyltransferase III; RT-qPCR, reverse
transcription-quantitative PCR; si, small interfering. |
By contrast, paclitaxel treatment did not alter the
mRNA and protein expression levels of ST3Gal3 in si-CTRL SKOV3
cells or si-ST3Gal3 SKOV3 cells (Fig.
4C and D). It was also found that paclitaxel induced caspase-3
protein expression, but not caspase-8 expression, in si-CTRL SKOV3
cells. ST3Gal3 knockdown alone induced caspase-3 expression, but
seemed to decrease caspase-8 expression, in SKOV3 cells, compared
with the si-CTRL. Finally, ST3Gal3 knockdown plus paclitaxel
treatment markedly increased both caspase-8 and caspase-3 protein
level in SKOV3 cells, compared with si-ST3Gal3 knockdown alone
(Fig. 4D). These results indicated
that ST3Gal3 knockdown synergistically facilitated
paclitaxel-mediated activation of caspase-8/3 signaling.
Discussion
Paclitaxel, a taxane-based drug, has been approved
for use in a number of solid tumors for decades. However, serious
side effects and chemoresistance are major obstacles that limit its
clinical application, even though this drug has a remarkable
response rate and increases the survival rate (34,35).
Therefore, focusing on reversing chemoresistance to paclitaxel
remains an important and urgent focus in cancer treatment. The
present results demonstrated that the expression levels of ST3Gal3
were associated with paclitaxel-resistance in ovarian cancer cell
lines. Of note, paclitaxel treatment increased the mRNA and protein
levels of ST3Gal3, while ST3Gal3 knockdown reduced the level of
paclitaxel-induced apoptosis in ovarian cancer cells. Thus, the
results of the present study suggested that ST3Gal3 may be a
potential target for improving paclitaxel-based chemotherapy.
The advance in glycoinformatics and glycoproteomics
has provided the needed tools to probe and understand glycosylation
in development and cancer biology (8,31). A
number of sialyltransferases and sialylated glycoproteins have been
identified as diagnostic or prognostic markers for ovarian cancer
(8,36). The ascites volume has been shown to
be correlated with the degree of sialylation in epithelial ovarian
cancer, providing a new insight into tumor progression and
recurrence (8). Our previous study
showed that the expression of sialyltransferase mRNA differs
significantly among different ovarian cancer cell lines (31). In the present study, it was found
that the expression of ST3Gal3 was higher in HO8910PM cells than in
SKOV3 cells, indicating that more (α-2,3)-linked sialylation events
may be catalyzed on the cell surface of HO8910PM cells. Aberrant
sialylations have been associated with a number of processes,
including adhesion, tumor progression and metastasis (7,37,38).
In addition, ST3Gal3 and ST6Gal1 play roles in
cisplatin-resistance, as previously reported (30,31,39).
However, the combination of cisplatin and paclitaxel serves as a
standard chemotherapy regimen for patients with advanced ovarian
cancer (40,41). Therefore, in the present study the
relationship between ST3Gal3 and paclitaxel-resistance in ovarian
cancer cell lines was investigated. The results of the present
study showed that a higher expression level of ST3Gal3 conferred
paclitaxel resistance to HO8910PM cells. Following siRNA depletion
of ST3Gal3 in HO8910PM cells, a higher level of apoptosis was
induced by paclitaxel compared with si-CTRL cells. There was no
significant difference in paclitaxel-induced apoptosis between
si-ST3Gal3 SKOV3 cells and si-ST3Gal3 HO8910PM cells. These results
suggested that ST3Gal3 plays an important role in chemoresistance
to paclitaxel in ovarian cancer cells.
The mechanism by which ST3Gal3 regulates
paclitaxel-resistance was also explored. At a mechanistic level,
paclitaxel stabilizes microtubule polymers to inhibit mitotic
spindle assembly, blocking cell division and triggering apoptosis
(34,35). The mechanism for
paclitaxel-resistance has been reported to be associated with
microtubule dynamics, tubulin isotype expression and the mutation,
or modifications, of tubulin-/microtubule-regulatory proteins
(34). The results of the present
study suggested an alternative mechanism for paclitaxel-resistance
based on the following findings: i) Paclitaxel treatment
upregulated ST3Gal3 expression at both the mRNA and protein level
in HO8910PM cells, but not in SKOV3 cells; ii) either paclitaxel
treatment alone or ST3Gal3 knockdown induced caspase-3 expression
in ovarian cancer cell lines; iii) paclitaxel alone induced
caspase-8 expression in HO8910PM cells, but not in SKOV3 cells; iv)
ST3Gal3 knockdown did not affect caspase-8 expression in HO8910PM
cells, and even reduced caspase-8 expression in SKOV3 cells; and v)
the combination of paclitaxel treatment and ST3Gal3 knockdown
markedly increased caspase-8 and caspase-3 expression in ovarian
cancer cell lines. These results indicated that ST3Gal3 knockdown
activated caspase-3, rather than caspase-8, signaling, however,
paclitaxel-induced caspase signaling depended on the cell type
involved. Therefore, ST3Gal3 knockdown may directly and
synergistically facilitate the paclitaxel-mediated activation of
caspase-3 signaling, while the activation of caspase-8 may be
dependent on the activation of caspase-3.
In conclusion, the results of the present study
suggested an alternative mechanism for paclitaxel-associated
chemoresistance in ovarian cancer cells. It was also indicated that
aberrant ST3Gal3 expression may serve as a diagnostic and
prognostic marker, and a potential chemotherapeutic target, for
ovarian cancer. Future studies should be directed towards the
development of sialyltransferase inhibitors; for example,
high-affinity lectin may be used to block sialylated sites, or gene
modification may be employed to downregulate the expression and
function of sialyltransferases, with the aim to ameliorate
resistance to paclitaxel-based chemotherapies.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Science Foundation of China (grant nos. 81071751 and 30770487), the
Opening Project of Zhejiang Provincial Top Key Discipline of
Pharmaceutical Sciences (grant no. 2016009) and the Guangzhou
Science and Technology Project (grant no. 201704030059).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW and SL conceived and designed the study, and
revised the manuscripts. XZ, XY, MC, SZ and JL contributed to in
vitro studies. XZ analyzed the data. XZ drafted and submitted
the manuscript. All authors have read the final manuscript and
approved the submission.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Prat J; FIGO Committee on Gynecologic
Oncology, : Abridged republication of FIGO's staging classification
for cancer of the ovary, fallopian tube, and peritoneum. Cancer.
121:3452–3454. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Rossing MA, Wicklund KG, Cushing-Haugen KL
and Weiss NS: Predictive value of symptoms for early detection of
ovarian cancer. J Natl Cancer Inst. 102:222–229. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zheng X, Chen S, Li L, Liu X, Liu X, Dai
S, Zhang P, Lu H, Lin Z, Yu Y and Li G: Evaluation of HE4 and TTR
for diagnosis of ovarian cancer: Comparison with CA-125. J Gynecol
Obstet Hum Reprod. 47:227–230. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Han C, Bellone S, Siegel ER, Altwerger G,
Menderes G, Bonazzoli E, Egawa-Takata T, Pettinella F, Bianchi A,
Riccio F, et al: A novel multiple biomarker panel for the early
detection of high-grade serous ovarian carcinoma. Gynecol Oncol.
149:585–591. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ferreira JA, Magalhães A, Gomes J, Peixoto
A, Gaiteiro C, Fernandes E, Santos LL and Reis CA: Protein
glycosylation in gastric and colorectal cancers: Toward cancer
detection and targeted therapeutics. Cancer Lett. 387:32–45. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dall'Olio F, Malagolini N, Trinchera M and
Chiricolo M: Sialosignaling: Sialyltransferases as engines of
self-fueling loops in cancer progression. Biochim Biophys Acta.
1840:2752–2764. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Biskup K, Braicu EI, Sehouli J, Tauber R
and Blanchard V: The ascites N-glycome of epithelial ovarian cancer
patients. J Proteomics. 157:33–39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rodrigues JG, Balmaña M, Macedo JA, Poças
J, Fernandes Â, de-Freitas-Junior JCM, Pinho SS, Gomes J, Magalhães
A, Gomes C, et al: Glycosylation in cancer: Selected roles in
tumour progression, immune modulation and metastasis. Cell Immunol.
333:46–57. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kuzmanov U, Musrap N, Kosanam H, Smith CR,
Batruch I, Dimitromanolakis A and Diamandis EP: Glycoproteomic
identification of potential glycoprotein biomarkers in ovarian
cancer proximal fluids. Clin Chem Lab Med. 51:1467–1476. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Alley WR Jr, Vasseur JA, Goetz JA, Svoboda
M, Mann BF, Matei DE, Menning N, Hussein A, Mechref Y and Novotny
MV: N-linked glycan structures and their expressions change in the
blood sera of ovarian cancer patients. J Proteome Res.
11:2282–2300. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shetty V, Nickens Z, Shah P, Sinnathamby
G, Semmes OJ and Philip R: Investigation of sialylation aberration
in N-linked glycopeptides by lectin and tandem labeling (LTL)
quantitative proteomics. Anal Chem. 82:9201–9210. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shetty V, Hafner J, Shah P, Nickens Z and
Philip R: Investigation of ovarian cancer associated sialylation
changes in N-linked glycopeptides by quantitative proteomics. Clin
Proteomics. 9:102012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mikami M, Tanabe K, Matsuo K, Miyazaki Y,
Miyazawa M, Hayashi M, Asai S, Ikeda M, Shida M, Hirasawa T, et al:
Fully-sialylated alpha-chain of complement 4-binding protein:
Diagnostic utility for ovarian clear cell carcinoma. Gynecol Oncol.
139:520–528. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Matsuo K, Tanabe K, Ikeda M, Shibata T,
Kajiwara H, Miyazawa M, Miyazawa M, Hayashi M, Shida M, Hirasawa T,
et al: Fully sialylated alpha-chain of complement 4-binding protein
(A2160): A novel prognostic marker for epithelial ovarian
carcinoma. Arch Gynecol Obstet. 297:749–756. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wen KC, Sung PL, Hsieh SL, Chou YT, Lee
OK, Wu CW and Wang PH: α2,3-sialyltransferase type I regulates
migration and peritoneal dissemination of ovarian cancer cells.
Oncotarget. 8:29013–2902. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Christie DR, Shaikh FM, Lucas JA IV, Lucas
JA III and Bellis SL: ST6Gal-I expression in ovarian cancer cells
promotes an invasive phenotype by altering integrin glycosylation
and function. J Ovarian Res. 1:32008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mikuła-Pietrasik J, Uruski P, Szubert S,
Szpurek D, Sajdak S, Tykarski A and Książek K: Malignant ascites
determine the transmesothelial invasion of ovarian cancer cells.
Int J Biochem Cell Biol. 92:6–13. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Szender JB, Emmons T, Belliotti S, Dickson
D, Khan A, Morrell K, Khan AN, Singel KL, Mayor PC, Moysich KB, et
al: Impact of ascites volume on clinical outcomes in ovarian
cancer: A cohort study. Gynecol Oncol. 146:491–497. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Orr B and Edwards RP: Diagnosis and
treatment of ovarian cancer. Hematol Oncol Clin North Am.
32:943–964. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zeng J, Huang H, Shan Y, Li Y, Jin Y and
Pan L: The effect of CA125 nadir level on survival of
advanced-stage epithelial ovarian carcinoma after interval
debulking surgery. J Cancer. 8:3410–3415. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jayson GC, Kohn EC, Kitchener HC and
Ledermann JA: Ovarian cancer. Lancet. 384:1376–1388. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Webber K and Friedlander M: Chemotherapy
for epithelial ovarian, fallopian tube and primary peritoneal
cancer. Best Pract Res Clin Obstet Gynaecol. 41:126–138. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu N, Mei L, Fan X, Tang C, Ji X, Hu X,
Shi W, Qian Y, Hussain M, Wu J, et al: Phosphodiesterase 5/protein
kinase G signal governs stemness of prostate cancer stem cells
through Hippo pathway. Cancer Lett. 378:38–50. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Maji S, Panda S, Samal SK, Shriwas O, Rath
R, Pellecchia M, Emdad L, Das SK, Fisher PB and Dash R: Chapter
three-Bcl-2 antiapoptotic family proteins and chemoresistance in
cancer. Adv Cancer Res. 137:37–75. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ween MP, Armstrong MA, Oehler MK and
Ricciardelli C: The role of ABC transporters in ovarian cancer
progression and chemoresistance. Crit Rev Oncol Hematol.
96:220–256. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kuchenbaecker KB, Hopper JL, Barnes DR,
Phillips KA, Mooij TM, Roos-Blom MJ, Jervis S, van Leeuwen FE,
Milne RL, Andrieu N, et al: Risks of breast, ovarian, and
contralateral breast cancer for BRCA1 and BRCA2 mutation carriers.
JAMA. 317:2402–2416. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chudecka-Głaz A, Cymbaluk-Płoska A,
Wężowska M and Menkiszak J: Could HE4 level measurements during
first-line chemotherapy predict response to treatment among ovarian
cancer patients? PLoS One. 13:e01942702018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Britain CM, Holdbrooks AT, Anderson JC,
Willey CD and Bellis SL: Sialylation of EGFR by the ST6Gal-I
sialyltransferase promotes EGFR activation and resistance to
gefitinib-mediated cell death. J Ovarian Res. 11:122018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schultz MJ, Swindall AF, Wright JW, Sztul
ES, Landen CN and Bellis SL: ST6Gal-I sialyltransferase confers
cisplatin resistance in ovarian tumor cells. J Ovarian Res.
6:252013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang X, Zhang Y, Lin H, Liu Y, Tan Y, Lin
J, Gao F and Lin S: Alpha2,3-sialyltransferase III knockdown
sensitized ovarian cancer cells to cisplatin-induced apoptosis.
Biochem Biophys Res Commun. 482:758–763. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Geisler C and Jarvis DL: Effective
glycoanalysis with Maackia amurensis lectins requires a clear
understanding of their binding specificities. Glycobiology.
21:988–993. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jordan MA and Wilson L: Microtubules as a
target for anticancer drugs. Nat Rev Cancer. 4:253–265. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Orr GA, Verdier-Pinard P, McDaid H and
Horwitz SB: Mechanisms of Taxol resistance related to microtubules.
Oncogene. 22:7280–7295. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bennun SV, Hizal DB, Heffner K, Can O,
Zhang H and Betenbaugh MJ: Systems glycobiology: Integrating
glycogenomics, glycoproteomics, glycomics, and other ‘omics data
sets to characterize cellular glycosylation processes. J Mol Biol.
428:3337–3352. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu J, Xie X, Nie S, Buckanovich RJ and
Lubman DM: Altered expression of sialylated glycoproteins in
ovarian cancer sera using lectin-based ELISA assay and quantitative
glycoproteomics analysis. J Proteome Res. 12:3342–3352. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cheng WK and Oon CE: How glycosylation
aids tumor angiogenesis: An updated review. Biomed Pharmacother.
103:1246–1252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang X, Pan C, Zhou L, Cai Z, Zhao S and
Yu D: Knockdown of ST6Gal-I increases cisplatin sensitivity in
cervical cancer cells. BMC Cancer. 16:9492016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Goldberg JM, Piver MS, Hempling RE and
Recio FO: Paclitaxel and cisplatin combination chemotherapy in
recurrent epithelial ovarian cancer. Gynecol Oncol. 63:312–317.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
du Bois A, Lück HJ, Meier W, Möbus V,
Costa S, Richter B, Warm M, Bauknecht T, Schröder W, Olbricht S, et
al: Carboplatin/paclitaxel versus cisplatin/paclitaxel as
first-line chemotherapy in advanced ovarian cancer: An interim
analysis of a randomized phase III trial of the Arbeitsgemeinschaft
Gynäkologische Onkologie Ovarian Cancer Study Group. Semin Oncol.
24 (5 Suppl 15):S15-44–S15-52. 1997.
|