Introduction
Stroke is one of the most prevalent diseases in the
world, with high mortality and morbidity rates, and ~80% of all
stroke events are cerebral arterial thrombosis- or embolism-induced
ischemia (1–3). The most effective and basic treatment
is the restoration of the blood supply by recanalization of the
occluded arteries (4). However,
this thrombolytic therapy is often accompanied by additional
injury, which is referred to as cerebral ischemia-reperfusion
injury (CIRI). Under such injury, brain damage has been
demonstrated to be aggravated, with intracellular calcium overload,
energy metabolism dysfunction and apoptosis functioning as the
primary processes involved (5).
The endoplasmic reticulum (ER) is an important
organelle, which serves a major role in maintaining the balance of
cellular Ca2+ and modifying proteins following
translation (6,7). Cerebral ischemia-reperfusion has been
demonstrated to cause ER stress (ERS), which leads to the false
folding and accumulation of proteins in the ER, triggering the
unfolded protein response (UPR). Endoplasmic reticulum chaperone
BiP (GRP78) is a central regulator for ERS as it is able to control
the activation of transmembrane ERS sensors,
serine/threonine-protein kinase/endoribonuclease IRE1, eukaryotic
translation initiation factor 2-alpha kinase 3 and cyclic
AMP-dependent transcription factor ATF-6 alpha, through a
binding-release mechanism. If the stress persists or becomes more
severe, cell apoptosis will be triggered by the UPR via the
activation of caspase-12 and DNA damage-inducible transcript 3
protein (CHOP) (8,9). According to previous studies, ERS is
one of the essential signaling mechanisms in the process of
neuronal injury caused by cerebral ischemia (10,11).
ERS inhibition may protect against neuronal injury (12).
Radix Scrophulariae, known as ‘Xuanshen’, obtained
from the dried root of Scrophularia ningpoensis Hemsl, is
widely used for treating ischemic cerebrovascular and
cardiovascular diseases (13,14).
Pharmacological research and clinical practice have demonstrated
that Radix Scrophulariae may delay the blood clotting process
(15), ameliorate cerebral
ischemia injury (16) and that it
exhibits anti-neurotoxic activities (17). Iridoid glycosides of Radix
Scrophulariae (IGRS) are a group of the major bioactive components
of Radix Scrophulariae, including harpagoside and harpagide. A
previous study presented evidence that acute cerebral ischemia may
be prevented by harpagide, which is known as one of the bioactive
components of IGRS as it exhibits anti-apoptotic effects (18).
However, the total range of pharmacological effects
of IGRS remain unknown. It is unclear whether IGRS protects against
CIRI, and to the best of our knowledge, the therapeutic effect of
IGRS on in vivo CIRI has not been investigated yet.
Therefore, the present study aimed to evaluate the effects of IGRS
on CIRI and to investigate the underlying mechanisms caused by
ERS.
Materials and methods
Experimental drugs
The IGRS components were provided by Chinese
Medicinal Resources Laboratory of Zhejiang Chinese Medical
University. A total of 53.19% of the IGRS was iridoid glycosides.
Edaravone was purchased from Jiangsu Simcere Pharmaceutical Co.
Ltd.
Laboratory animals
A total of 96 healthy male Sprague Dawley rats (6–8
weeks old, 200±20 g) were provided by the Experimental Animal
Center of Zhejiang Chinese Medical University [lot no. SCXK
(Shanghai) 2013–0016]. The animals were housed in the room under a
controlled temperature (20-24°C) for 7 days prior to use, with a 12
h light/dark cycle. All experiments were performed according to the
National Institutes of Health Guide for the Care and Use of
Laboratory Animals and approved by the Animal Care Committee of
Zhejiang Chinese Medical University. The procedures were
implemented in accordance with the National Centre for the
Replacement, Refinement and Reduction of Animals in Research ARRIVE
guidelines (www.nc3rs.org.uk/arrive-guidelines) (19) and the AVMA euthanasia guidelines
2013 (20). All efforts were made
to minimize the number of animals used and the suffering.
Preparation of CIRI rat model
The CIRI rat model was prepared according to an
intraluminal suture method, as previously described (21). Briefly, the rats were anesthetized
by an intraperitoneal (I.P.) injection of 350 mg/kg 10% chloral
hydrate. No signs of peritonitis were observed following the
administration of the 10% chloral hydrate. Following a midline neck
incision, the right common carotid artery and external carotid
arteries were isolated. A 0.28-mm nylon filament (Beijing Cinontech
Co., Ltd.) was inserted through the external carotid arteries into
the right internal carotid artery to block the right middle
cerebral artery with an insertion length of 18–20 mm (22). Reperfusion was initiated 90 min
after the onset of ischemia by gently removing the filament.
Sham-operated rats underwent the same surgery, with the exception
that the filament was inserted and withdrawn immediately. The rats
were kept at 37±0.5°C with a heating lamp during the procedure.
Following recovery from the anesthesia, the rats were placed back
into their cages with ad libitum access to food and
water.
Experimental design
All animals were randomly divided into six groups:
Sham-operation with saline treatment (Sham); CIRI with saline
treatment (Model); model establishment and 3 mg/kg edaravone
administration (edaravone-treated); model establishment and 30, 60
or 120 mg/kg IGRS administration. Prior to surgery, each group was
given the corresponding drugs once a day for 7 days, with the
exception of the edaravone-treated group, which was given
physiological saline (10 ml/kg). A total of 1 h prior to the
procedure, all the rats in the Sham, Model and the different IGRS
groups received gastric perfusions of their respective drugs, while
the rats in the edaravone-treated group were given an I.P.
injection of edaravone. The administration was performed again 6 h
after the operation. A total of 10 ml/kg physiological saline was
administered to the Sham and Model groups (Fig. 1).
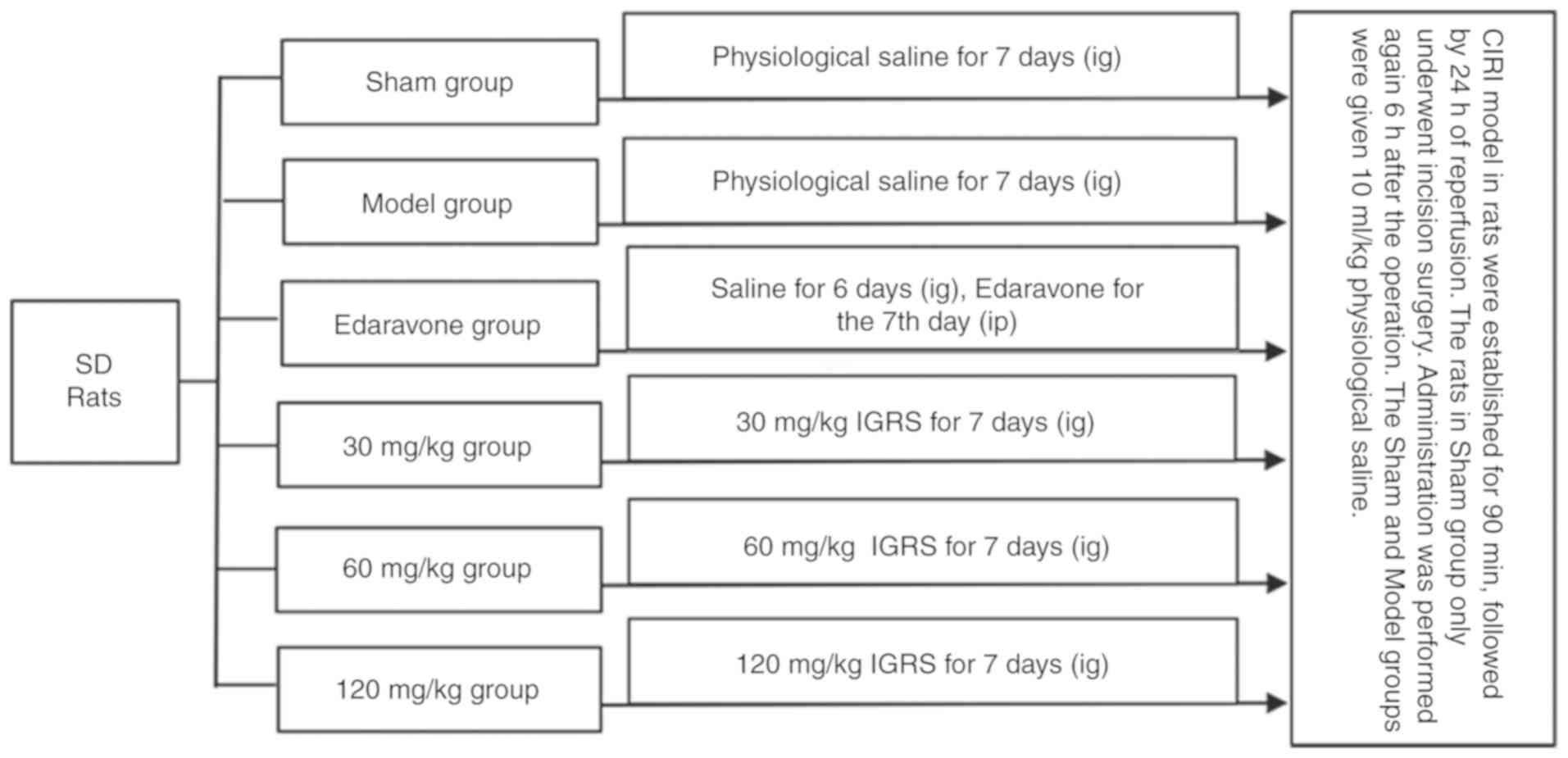 | Figure 1.Experimental design. A CIRI model in
rats was established by occluding the right middle cerebral artery
for 90 min and followed by 24 h of reperfusion. Prior to surgery,
each group was given the corresponding drugs once a day for 7 days,
with the exception of the edaravone-treated group, which was given
physiological saline (10 ml/kg). A total of 1 h prior to surgery,
rats in the Sham, Model and the IGRS groups had their drugs
administered IG, while the rats in the edaravone group were given
edaravone IP. The drugs were re-administration again 6 h after the
surgical procedure. CIRI, cerebral ischemia-reperfusion injury;
IGRS, iridoid glycosides from Radix Scrophulariae; IG,
intragastric; IP, intraperitoneal; SD, Sprague Dawley. |
The duration of the experiment was 8 days, including
7 days of IGRS pretreatment, 90 min of cerebral ischemia and 24 h
of reperfusion. When cerebral ischemia reperfusion lasted for 24 h,
CIRI rats exhibited hemiplegic symptoms to different degrees on one
side of limbs, which affected their normal eating. The weight of
CIRI rats dropped by 20% on average, accompanied by symptoms of
rapid breathing. Therefore, it was determined that the rats should
be euthanized after 24 h of cerebral ischemia reperfusion. All the
animals used were anesthetized with 10% chloral hydrate (350 mg/kg
body weight; I.P.) and decapitated rapidly after 24 h CIRI. No
treatment-associated mortalities were observed. A combination of
criteria were used to confirm death, including: Lack of pulse,
breathing, corneal reflex and response to firm toe pinch; inability
to hear respiratory sounds and heartbeat by use of a stethoscope;
graying of the mucous membranes; and rigor mortis. Following
confirmation of death, the brain tissues were removed immediately
for subsequent study.
Neurological scores
The rats underwent a neurological severity score
test as previously described (23)
at 24 h of reperfusion, including a set of exercise, sensation,
reflection and balance tests. Neurological function was graded on a
scale of 0–18 (Table I).
 | Table I.Modified neurological severity score
grading system. |
Table I.
Modified neurological severity score
grading system.
| Points | Degree of
injury |
|---|
| 0 | Normal |
| 1–6 | Mild injury |
| 7–12 | Moderate
injury |
| 13–18 | Serious damage |
| 18 | Most severe
neurological deficits |
Measurement of ischemic infarction
volume
At 24 h post-reperfusion, rats were anesthetized
with 10% chloral hydrate (350 mg/kg body weight; I.P.) prior to
sacrifice. Their brains were removed, divided into 6 parts of 2 mm
coronal slices, and dyed with 2% 2,3,5-triphenyltetrazolium
chloride (TTC) in PBS at 37°C for 15 min, which was then replaced
with 4% paraformaldehyde for 10 min (2). The white areas of the brains were
labeled as infarct tissue and the red areas indicated normal
tissue. Images of the TTC-stained sections were captured and
analyzed with Image-Pro Plus 6.0 (Media Cybernetics, Inc.). The
infarct volume was calculated as a percentage as described
previously to avoid inaccurate secondary measurements of edema
(24,25).
Measurement of brain water
content
Rats were sacrificed and the brain tissues were
immediately weighed to obtain the wet weight (WW). The tissues were
then dried at 60°C for 24 h and weighed again to obtain the dry
weight (DW). The water content was calculated according to the
following formula: (WW-DW)/WW ×100 as described previously
(26,27).
Histology analysis
Brain tissues were harvested and fixed in 4%
paraformaldehyde for 24 h at 4°C. An ethanol gradient was used to
dehydrate the samples according to the following concentration: 50%
ethanol (2 h), 60% ethanol (2 h), 75% ethanol (2 h), 85% ethanol (3
h), 95% ethanol (2 h) and 100% ethanol (2 h). Then the tissues were
cleared, paraffin-embedded, sectioned and stuck to glass slides at
4°C. Following conventional de-waxing and washing, the sections
were stained with hematoxylin and eosin Staining kit (Phygene Life
Sciences Co., Ltd). Briefly, the sections were incubated with
hematoxylin (~0.5%) for 5 min and with eosin (~0.5%) for 2 min at
room temperature. Subsequently, the histological outcomes were
evaluated under a light microscope at magnification, ×400.
Denatured cells exhibited a shrinking nucleus, while live cells
retained normal cellular morphology. The numbers of denatured cells
in the cortex were obtained from observing 4 non-overlapping
microscopic fields of view. The degree of injury was indicated
according to the scores of denatured cell index (DCI) by comparing
the number of denatured cells to the number of total cells as
described previously (28).
Detection of apoptotic cell death
using a terminal deoxynucleotidyl transferase mediated dUTP nick
end labeling (TUNEL) assay
The degree of apoptosis in the brain cortex of
paraffin-embedded coronal brain sections of animals from all the
groups was analyzed by TUNEL assay using an In situ Cell
Death Detection kit (Roche Diagnostics). Briefly, the tissue
sections were fixed with 4% paraformaldehyde for 24 h at 4°C.
Following conventional washing, the slides were incubated with
Protein K (20 µg/ml) for 20 min at 37°C. Then the slides were
rinsed twice with 0.1 mol/l phosphate buffered solution (PBS; pH
7.4). The TUNEL reaction mixture (50 µl; 45 µl Label Solution with
5 µl Enzyme Solution) was added to the sample. The section was
incubated for 60 min at 37°C in a humidified chamber in the dark.
After that, the slide was rinsed 4 times with PBS. Sections were
mounted with DAPI (0.5–10 µg/ml) and placed under a fluorescence
microscope (magnification, ×400). A total of 4 sections were used
as specimens and ten fields were randomly selected per section, and
statistically analyzed using Image-Pro Plus 6.0 software(Media
Cybernetics, Inc.). The level of apoptosis was expressed as a ratio
of TUNEL-positive neurons to DAPI-labeled entire neurons as
described previously (29,30).
Transmission electron microscopy
(TEM)
Brain tissues were harvested and the cerebral
hippocampus was dissected and cut into 1×1×1 mm-sized sections and
immediately placed in 2.5% glutaraldehyde in 0.1 mol/l PBS (pH 7.4)
at 4°C overnight. The sections were rinsed three times with 0.1
mol/l PBS and immersed in 1% osmium tetroxide in 0.1 mol/l PBS for
2 h at 4°C. The tissue block was dehydrated in graded ethanol
solutions and embedded in epoxy resin. Polymerization was performed
at 70°C overnight and the samples were sectioned into a thickness
of 70 nm. Following staining with 50% ethanol saturated solution of
uranyl acetate for 1 h and lead citrate solution for 15 min at room
temperature, the sections were observed under a TEM (H-7650 TEM;
Hitachi, Ltd.) (31). The
observations of cell structures in sections were made visually.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Rats were sacrificed at 24 h post-reperfusion. To
analyze the expression levels of GRP78, CHOP and caspase-12, the
hippocampus tissue was separated from the injured side of the
brain. The total RNA was extracted using TRIzol® reagent
(Thermo Fisher Scientific, Inc.), cDNA was produced using a
PrimeScript™ RT reagent kit with gDNA Eraser (Takara Biotechnology
Co., Ltd.). RT-qPCR was conducted on an Applied Biosystems 7500 and
7500 FAST Real-Time PCR detection system (Applied Biosystems;
Thermo Fisher Scientific, Inc.) using SYBR Green (Beijing ComWin
Biotech, Co., Ltd.) for fluorescent quantification. All reactions
were repeated 3 times. Data normalization was completed using GAPDH
as an endogenous control, and the normalized values were assessed
using the 2−ΔΔCq formula to compute the fold difference
between the control and experimental groups (32). The sequences of the primers used in
this experiment are presented in Table II.
| Table II.Primer sequences used in the
quantitative polymerase chain reaction assay. |
Table II.
Primer sequences used in the
quantitative polymerase chain reaction assay.
| Gene | Forward | Reverse |
|---|
| GRP78 |
5′-TGTCTTCTCAGCATCAAGCAAGG-3′ |
5′-CCAACACTTCCTGGACAGGCTT-3′ |
| CHOP |
5′-GGAGGTCCTGTCCTCAGATGAA-3′ |
5′-GCTCCTCTGTCAGCCAAGCTAG-3′ |
| Caspase-12 |
5′-CAGATGAGGAACGTGTGTTGAGC-3′ |
5′-GGAACCAGTCTTGCCTACCTTC-3′ |
| GAPDH |
5′-ACAGCAACAGGGTGGTGAC-3′ |
5′-TTTGAGGGTGCAGCGAACTT-3′ |
Western blot analysis
The hippocampus tissue was separated from the
injured side of the brain and frozen quickly in liquid nitrogen,
then transferred to a −80°C freezer for storage. The tissue were
homogenized in 300 µl RIPA lysis buffer containing PMSF (cat. no.
ST506; Beyotime institute of Biotechnology) and then incubated at
0°C for 30 min, followed by centrifugation at 12,000 × g at 4°C for
5 min. The supernatant was collected and denatured by boiling for 5
min. Protein concentration was quantified by the
micro-bicinchoninic acid (BCA) kit (cat. no. CW0014; Beijing ComWin
Biotech, Co., Ltd.). The lysates were loaded onto 10% SDS-PAGE for
the separation of protein GRP78 and caspase-12, 15% SDS-PAGE for
the separation of protein CHOP (30 µg of protein was loaded per
lane). The separated protein bands were transferred onto
polyvinylidene fluoride membranes (EMD Millipore) at 300 mA for 1.5
h. The membrane was blocked with blocking buffer containing 5%
fat-free milk for 2 h at room temperature and incubated with the
following primary antibodies at 4°C overnight: Rabbit anti-GRP78
(1:1,000, cat. no. ab108613), rabbit anti-CHOP (1:1,000, cat. no.
ab11419), rabbit anti-caspase-12 (1:1,000, cat. no. ab62484) and
mouse anti-β-actin (1:1,000, cat. no. ab8226; all from Abcam). The
membranes were washed three times with Tris-buffered saline (TBS)
containing 0.1% Tween-20 (TBST; pH 7.4) and then incubated in
horseradish peroxidase-conjugated secondary antibody (goat
anti-rabbit, 1:2,000, cat. no. C50113, LI-COR Biosciences; goat
anti-mouse, 1:15,000, cat. no. C50331, LI-COR Biosciences) for 2 h
at room temperature in the dark and then washed three times with
TBST. The membranes were developed using the Odyssey Fluorescence
Scanning Imaging System (LI-BOR Biosciences). To minimize
experimental variation, each protein expression experiment was
processed in parallel (33). The
protein results were analyzed with Image-Pro Plus 6.0 analysis
software (Media Cybernetics, Inc. USA). The ratio of the gray value
of the target protein to that of the internal reference protein was
taken as the relative gray value (34).
Statistical analysis
The experimental data were analyzed by SPSS v.17.0
(SPSS, Inc.) and GraphPad Prism v.5.0 software (GraphPad Software,
Inc.). The results were expressed as mean ± standard deviation and
analyzed by one-way analysis of variance followed by a Dunnett's
post-hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
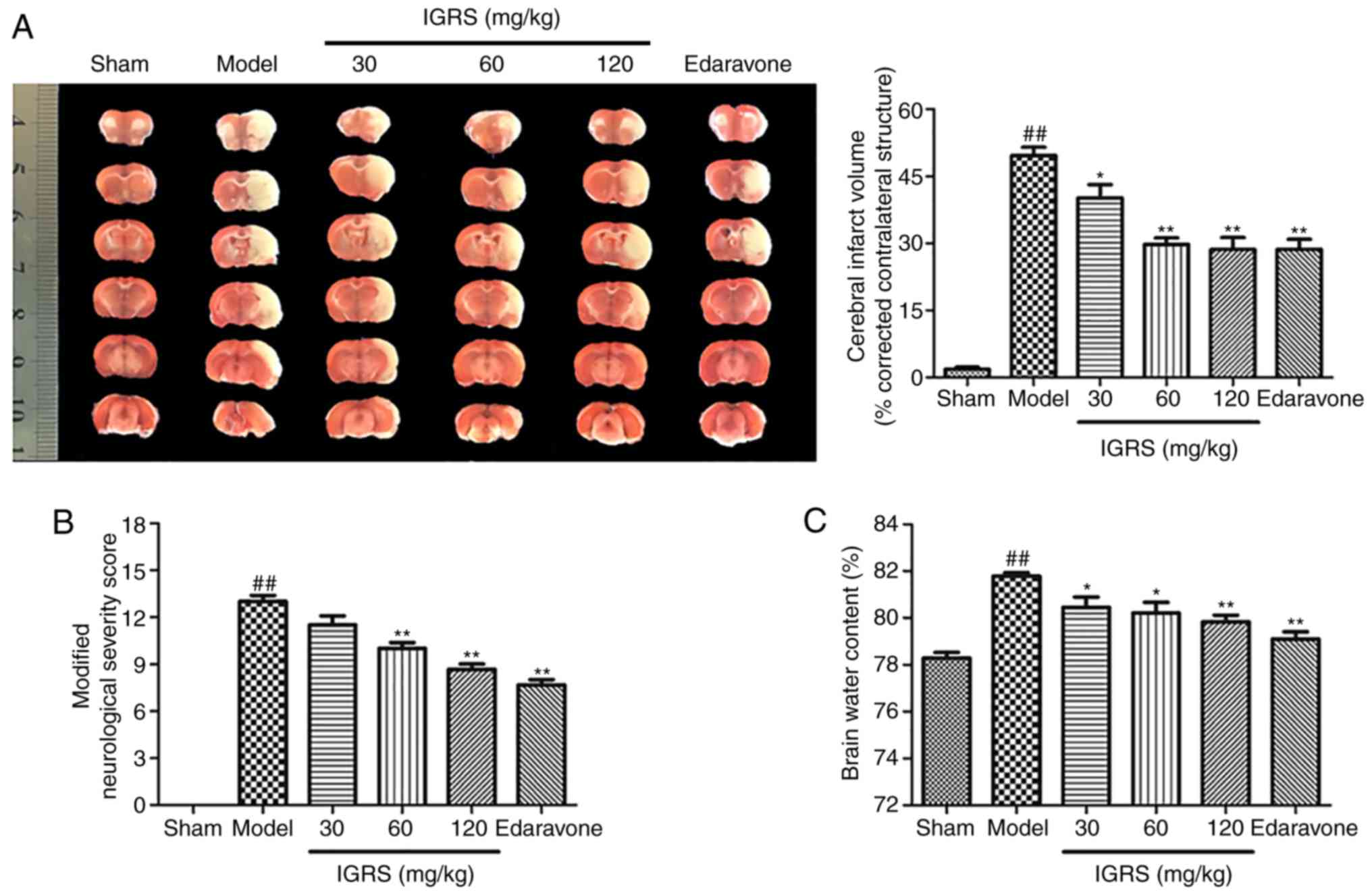
IGRS confers a protective effect
against CIRI
The present study first evaluated whether
pretreatment with IGRS conferred a protective effect in CIRI. After
24 h of reperfusion, infarct volumes were evaluated by using TTC
staining. The 30, 60 and 120 mg/kg IGRS-treated groups exhibited
significantly decreased infarct volumes compared with the Model
group (Fig. 2A). The protective
effect of IGRS was confirmed by comparing the volumes of cerebral
infarction. The cerebral infarct volumes of the 30, 60 and 120
mg/kg IGRS-treated groups were 40.2, 29.75 and 28.61%,
respectively, while that the Model group was 49.69%. Neurological
scores were examined at 24 h after reperfusion and scored on an
18-point scale (Fig. 2B). The
neurological scores decreased from 13 points in the Model group to
8.67 in the 120 mg/kg IGRS-treated group, indicating that the
degree of injury was significantly decreased.
Brain water content was subsequently evaluated using
the wet-dry method after reperfusion of 24 h compared with the
Model group, the brain water content in the 30, 60 and 120 mg/kg
IGRS-treated groups were decreased significantly (Fig. 2C). Altogether, the results
demonstrated that IGRS decreased CIRI-induced neurological deficits
and attenuated CIRI infarct volumes and brain edema, suggesting its
therapeutic potential for ischemic brain injury.
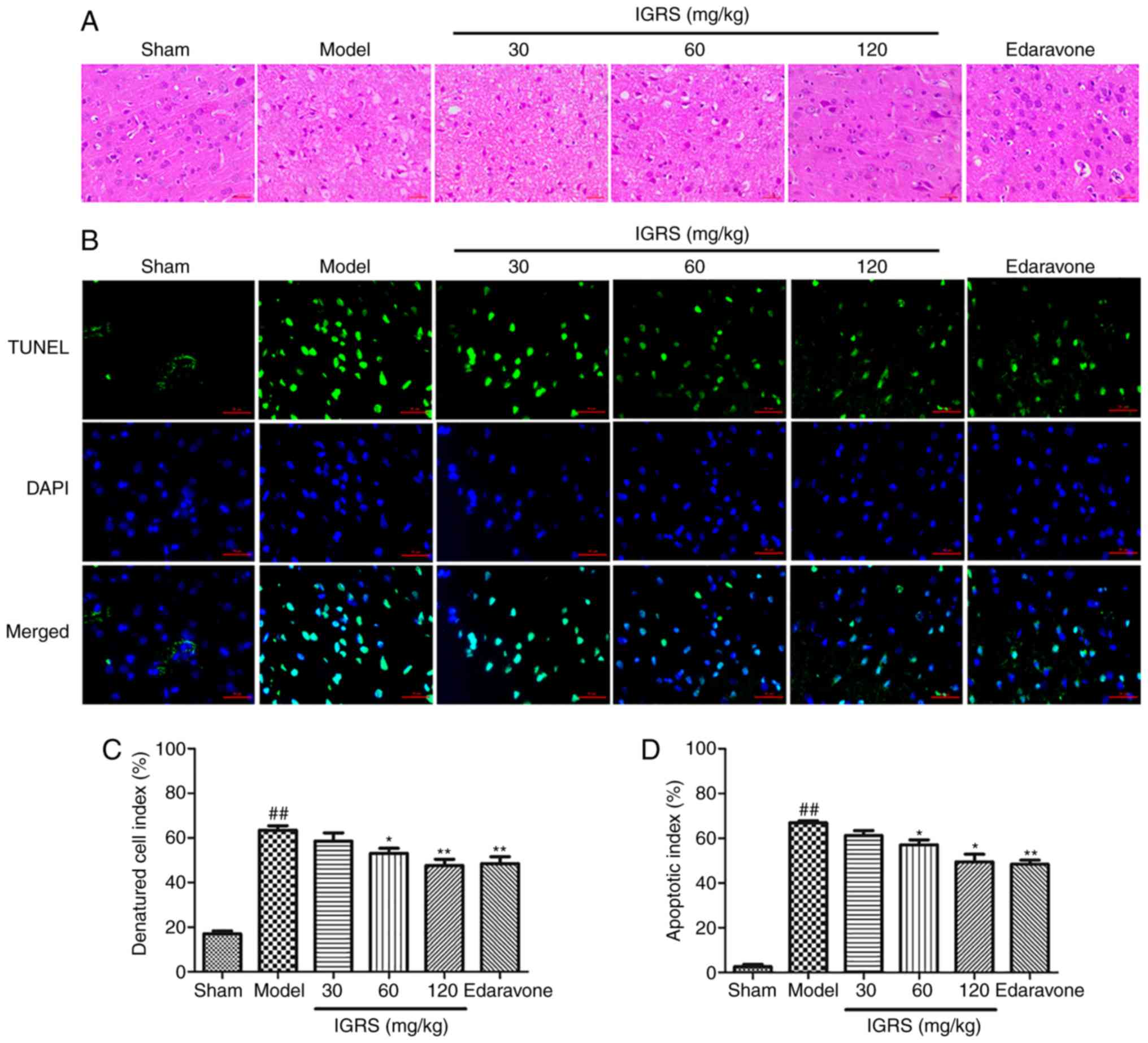
IGRS ameliorates CIRI-induced neuronal
damage
In order to investigate the neuroprotective effect
of IGRS on CIRI-induced neuronal damage, the cortical tissues of
the rats were stained, and the morphological changes were evaluated
under a microscope (Fig. 3A and
C). In the Sham group, the cerebral cortex was normal in
morphology and structure; it had a large number of nerve cells with
abundant cytoplasm, and large and round nuclei. Compared with the
normal neurons in the Sham group, severe cellular edema, condensed
nuclei, nuclear loss and a significant DCI rises were observed in
the Model group (P<0.01). A protective effect of IGRS was
observed in the 30, 60 and 120 mg/kg IGRS-treated groups; the
tissues were less edematous and the neurons possessed clearer
nuclei (Fig. 3A and C).
Furthermore, IGRS treatment normalized the glial cells in CIRI rats
and markedly decreased the DCI scores.
TUNEL staining was used to detect nerve cell
apoptosis in the cerebral cortex at 24 h after reperfusion
(Fig. 3B and D). Apoptotic cells
exhibited a green fluorescence signal, which was regarded as
positive TUNEL staining. Fewer TUNEL-positive cells were observed
in the Sham group. Conversely, the number of apoptotic cells in the
Model group increased significantly within the cortical ischemic
region (P<0.01). The apoptosis rate of cortical nerve cells was
2.57% in Sham group and 66.89% in Model group, while in the 30, 60
and 120 mg/kg IGRS-treated groups the apoptosis rates were 61.25,
57.16 and 49.48%, respectively. Therefore, in contrast to the Model
group, the apoptosis rate of nerve cells in the IGRS-treated
groups, especially at 120 mg/kg, was decreased significantly
(P<0.05).
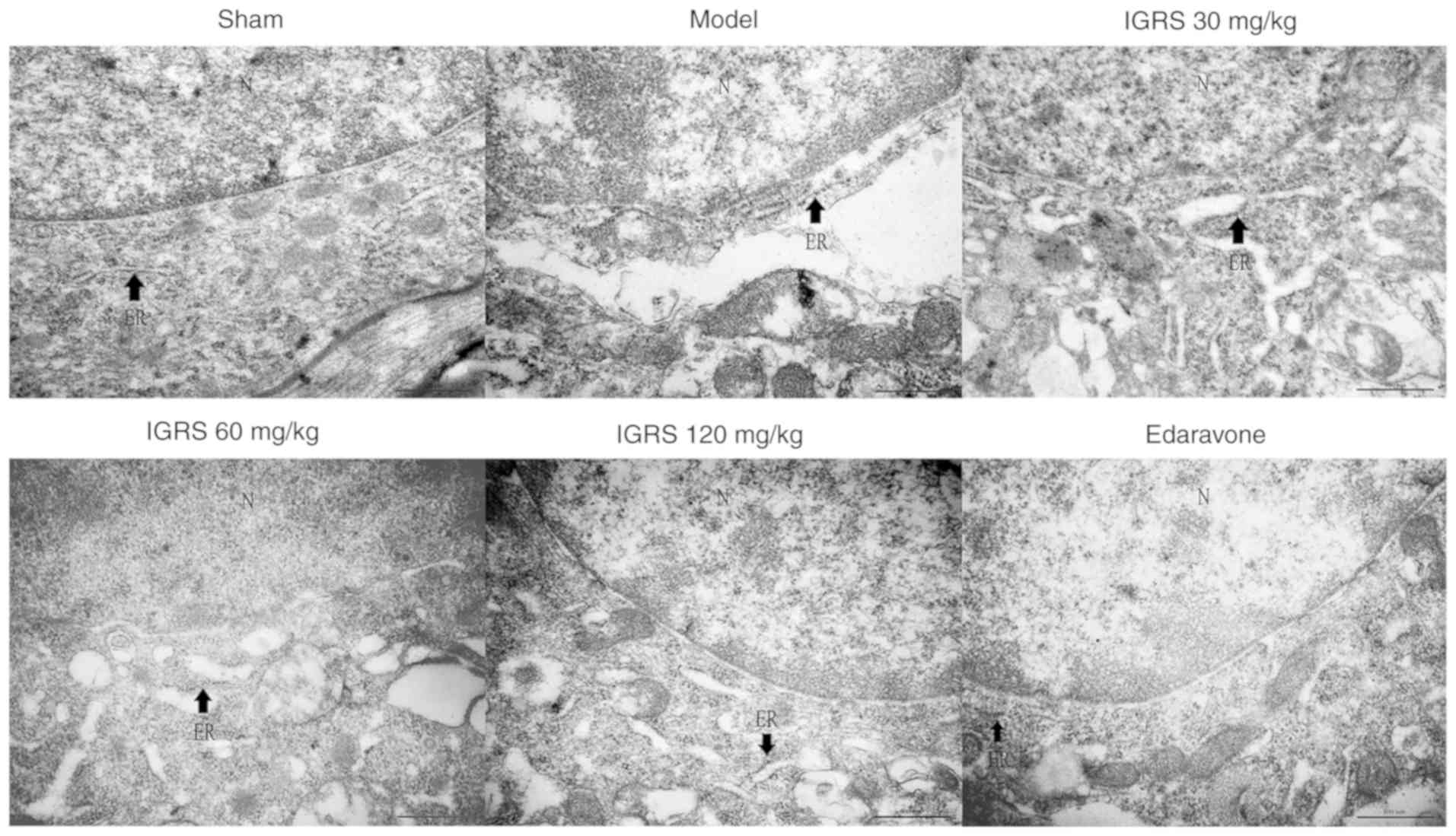
IGRS improves ER morphological
changes
To provide further insight into the protective
effect of IGRS on ER morphological changes in neurons induced by
CIRI, the hippocampus tissue was examined for ultrastructural
changes by TEM. The neurons in the Sham group exhibited integrated
structures, there were large numbers of rough (R)ER with normal
morphology and the mitochondria around the nuclei were normal.
Compared with the Sham group, the neurons in the Model group were
significantly swollen, the RER was dilated and swollen, and the
ribosomes had disassociated from the RER. These observations are
characteristic changes of ER morphology in states of ERS. In
contrast to the Model group, the ER morphology in the neurons of
the 30, 60 and 120 mg/kg IGRS-treated groups and the
edaravone-treated group exhibited different degrees of recovery.
The degree of swelling and the loss of ribosomes were notably
alleviated, and the number of ribosomes in the cytoplasmic matrix
was notably decreased (Fig.
4).
IGRS ameliorates the expression of
apoptosis factors mediated by ERS
The aforementioned results indicated that IGRS
protected against CIRI by suppressing ERS. To additionally confirm
the regulation mechanisms of IGRS, the expression levels of ERS
indicators, GRP78, CHOP and caspase-12, were determined in the
Model and IGRS-treated groups using RT-qPCR and western blot
analyses. The RT-qPCR data indicated that the mRNA levels of GRP78,
CHOP and caspase-12 were markedly upregulated in the Model group
compared with the Sham group, whereas IGRS treatment significantly
downregulated the mRNA levels of GRP78, CHOP and caspase-12
(P<0.05; Fig. 5A-C). The
relative expression levels of GRP78 mRNA in hippocampus of rats in
the 120 mg/kg IGRS group was 1.64, which was decreased by ~36.4%
compared with the Model group (P<0.05). The relative expression
of CHOP mRNA in the 120 mg/kg IGRS group was 1.56, which was
decreased by ~43.3% compared with the Model group (P<0.01). The
relative expression of all mRNA transcripts in the 30 mg/kg IGRS
group was slightly increased compared with that in the 120 mg/kg
IGRS group, but much lower compared with that in the Model group.
The relative expression levels of caspase-12 mRNA in the 60 and 120
mg/kg groups were similar to each other.
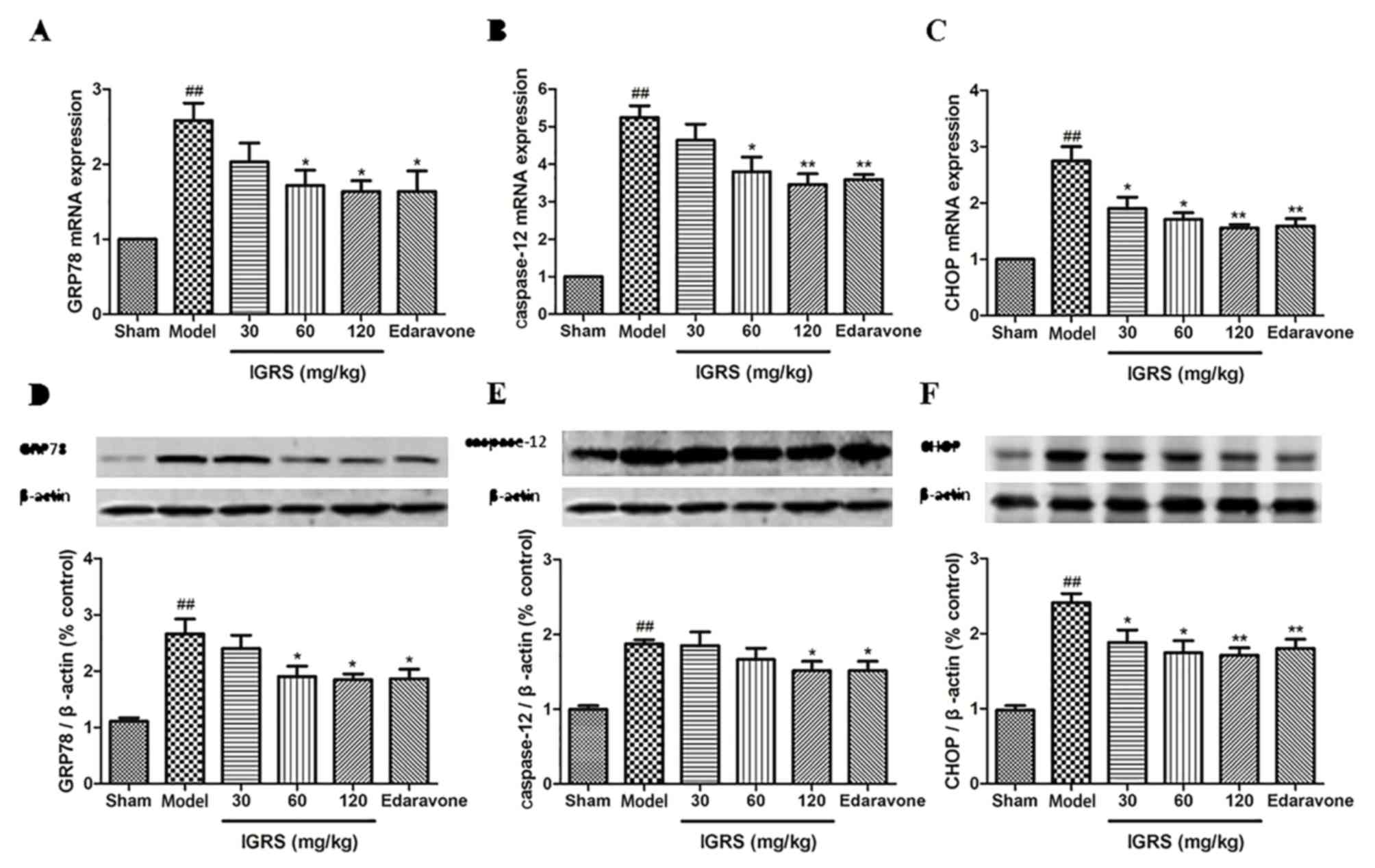 | Figure 5.Effect of IGRS on the expression of
GRP78, caspase-12 and CHOP mRNA and protein in cerebral
ischemia-reperfusion injured rats. The brain tissues of the
experimental rats were removed, and the hippocampus tissues were
separated from the injured side of the brain at 24 h
post-reperfusion. The total RNA was extracted, cDNA was produced,
and RT-qPCR was conducted. The expression of GAPDH was used as a
loading control. The expression of β-actin was used as a loading
control for western blot analysis. (A) GRP78, (B) caspase-12 and
(C) CHOP mRNA levels were determined by RT-qPCR (n=5 per group).
The expression levels of (D) GRP78, (E) caspase-12 and (F) CHOP
protein were determined by western blot analysis (n=5 per group).
Values are presented as mean ± standard deviation of each group.
##P<0.01 vs. Sham group. *P<0.05 and **P<0.01
vs. Model group. RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; IGRS, iridoid glycosides from Radix
Scrophulariae; GRP78, endoplasmic reticulum chaperone BiP; CHOP,
DNA damage-inducible transcript 3 protein. |
The western blot analysis data demonstrated similar
protein expression results for GRP78, CHOP and caspase-12, which
were significantly increased in the Model group when compared with
that of the Sham group. The ERS-induced protein expression levels
was significantly downregulated following IGRS treatment
(P<0.05) and had similar trends to the mRNA results (Fig. 5D-F).
Discussion
Acute ischemic stroke remains a leading cause of
mortality and long-term disability in the word. Ischemic stroke is
the result of a transient or permanent decrease of cerebral blood
flow caused by the blocking of a cerebral artery; in animal models,
this is achieved by simulating a local thrombus with an embolus.
During the prompt recovery of blood flow to the ischemic tissue,
reperfusion injury occurs and aggravates the initial injury, known
as secondary neuronal damage (35,36).
In cases of strokes in humans, cerebral vessel occlusion is seldom
permanent; the majority of cases of human ischemic stroke result in
spontaneous or thrombolytic therapy-induced reperfusion. Currently,
the thread embolism method is commonly used in surgical procedures
to establish a CIRI model without the need for a craniotomy. Using
this method, different states of human transient and permanent
focal cerebral ischemia may be simulated, and ischemia and
reperfusion times may be accurately controlled (37–39).
Intravenous tissue plasminogen activator is an approved medication
demonstrated to improve functional outcomes when stroke occurs
(40). The aim of the present
study was to explore the effect of IGRS on the injury caused by
cerebral ischemia-reperfusion. Various mechanisms are involved in
the pathological process of CIRI; there is considerable evidence
indicating that apoptosis serves an essential role in its
progression (41–42). In the present study, a CIRI model
was successfully established in rats, leading to brain damage,
which was consistent with previous results (43–45).
Previous studies had used TUNEL staining to evaluate the apoptosis
of neurons in a CIRI model (46).
Therefore, TUNEL staining was used to detect neuronal apoptosis in
CIRI rats in the present study.
Radix Scrophulariae is widely used in Traditional
Chinese Medicine for a broad range of diseases as it has a high
concentration of iridoids. The iridoids possess a wide range of
pharmacological properties, including anti-angiogenesis,
neuroprotection and cardiovascular protection (47). The iridoids from Radix
Scrophulariae have been identified as the active class of compounds
with the neuroprotective effect (48). There are a number of previous
studies demonstrating that early administration of extracts from
natural plants may improve cerebral ischemia reperfusion injury and
neurological function (49–51).
Therefore, prior to surgery, the IGRS-treated groups were given the
corresponding drugs once a day for 7 days. In addition, edaravone,
a potent free radical scavenger, was used to verify the protective
effect of IGRS in the cerebral ischemia-reperfusion injury
model.
It was observed that neurological deficits were
effectively improved, the cerebral infarct volume was decreased,
brain edema was alleviated and neuronal cells were protected
subsequent to treatment with IGRS following CIRI. The results
indicated dose-dependent neuroprotective effects, suggesting the
neuroprotective roles of IGRS. Consistent with the previous studies
(52,53), the apoptosis rate was high in the
Model group according to the results of the TUNEL assay. The
decrease in the apoptosis rate in the IGRS-treated groups suggested
that IGRS has the potential to inhibit neuronal apoptosis and
apoptosis pathways.
ERS is the primary intracellular signal transduction
pathway of cell apoptosis, serving a critical role in ischemic
neuronal cell apoptosis, as described previously (9). ER chaperones including GRP78, CHOP
and caspase-12 are highly sensitive to CIRI and are representative
of ERS (54). Previous data has
indicated that GRP78 and caspase-12 serve major roles in cerebral
ischemia, and that cell apoptosis was induced by the overexpression
of CHOP via the inhibition of Bcl-2; therefore, it was suggested
that CIRI could be alleviated by modulating CHOP (55,56).
In the present study, it was identified that the marked increase in
the expression levels of GRP78, CHOP and caspase-12 was caused by
CIRI, but was markedly downregulated following IGRS treatment.
Therefore, IGRS may be a potential neuroprotective medicine by
inhibiting the expression of GRP78, CHOP and caspase-12 induced by
ERS.
Due to the severe clinical consequences of stroke,
prevention and acute management are of paramount importance in
clinical treatment. In addition, secondary stroke prevention is
concerned with averting recurrent strokes following an initial
stroke or transient ischemic attack (40). The majority of patients survive a
first-time ischemic stroke event but are at high risk for recurrent
stroke and concomitant cardio- and peripheral vascular diseases.
Therefore, preventive measures are also an indispensable part in
the treatment of patients who have suffered a stroke, especially
for the prevention of recurrent stroke in individuals with a
history of ischemic stroke. Therefore, in the present study, a week
of pre-administration was performed prior to construction of the
CIRI model, to relieve tissue damage caused by stroke. Permanent
necrosis of nerve cells can occur easily following cerebral
ischemia reperfusion; therefore, an effective rescue time of 6 h
has been accepted to improve the penumbra in the ischemic area.
Consolidating treatment on the basis of prevention can decrease
injury to a great extent, which is consistent with the goal of the
clinical prevention and treatment of stroke. In drug-based therapy,
there are usually ≥2 types of drugs used in combination, to
function synergistically. In the present study, IGRS were
administered early in the establishment of the model and
post-surgery. The results of the present study also provided
evidence that IGRS attenuated CIRI and the protective effects were
related to the inhibition of ERS, demonstrating the feasibility of
this drug delivery method. In summary, the present study provided a
theoretical basis for the development and application of IGRS in
the prevention and treatment of stroke.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81573643).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZH and XZ conceived, designed and carried out the
experiments. YC and LZ performed the experiments and were major
contributors in writing the manuscript. XG and HG assisted in parts
of the experiment and participated in the data analysis. RC
participated in the interpretation of data statistics and revised
the manuscript. FQ participated in the establishment of cerebral
ischemia-reperfusion model in rats and the acquisition of data
during the experiments. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The rats were provided by the Experimental Animal
Center of Zhejiang Chinese Medical University. All experiments were
performed in accordance with the National Institutes of Health
Guide for the Care and Use of Laboratory Animals, and approved by
the Animal Care Committee of Zhejiang Chinese Medical
University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IGRS
|
iridoid glycosides from Radix
Scrophulariae
|
|
CIRI
|
cerebral ischemia-reperfusion
injury
|
|
ERS
|
endoplasmic reticulum stress
|
References
|
1
|
Sun K, Fan J and Han J: Ameliorating
effects of traditional Chinese medicine preparation, Chinese
materia medica and active compounds on ischemia/reperfusion-induced
cerebral microcirculatory disturbances and neuron damage. Acta
Pharm Sin B. 5:8–24. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen HS, Qi SH and Shen JG:
One-compound-multi-target: Combination prospect of natural
compounds with thrombolytic therapy in acute ischemic stroke. Curr
Neuropharmacol. 15:134–156. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cheng CY and Lee YC: Anti-inflammatory
effects of traditional chinese medicines against ischemic injury in
in vivo models of cerebral ischemia. Evid Based Complement Alternat
Med. 2016:57394342016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen XM, Chen HS, Xu MJ and Shen JG:
Targeting reactive nitrogen species: A promising therapeutic
strategy for cerebral ischemia-reperfusion injury. Acta Pharmacol
Sin. 34:67–77. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hu Y, Deng H, Xu S and Zhang J: MicroRNAs
regulate mitochondrial function in cerebral ischemia-reperfusion
injury. Int J Mol Sci. 16:24895–24917. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang HY, Wang ZG, Lu XH, Kong XX and Xiao
J: Endoplasmic reticulum stress: Relevance and therapeutics in
central nervous system diseases. Mol Neurobiol. 51:1343–1352. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li JQ, Yu JT, Jiang T and Tan L:
Endoplasmic reticulum dysfunction in Alzheimer's disease. Mol
Neurobiol. 51:383–395. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee AS: The ER chaperone and signaling
regulator GRP78/BIP as a monitor of endoplasmic reticulum stress.
Methods. 35:373–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gong L, Tang Y, An R, Lin M, Chen L and Du
J: RTN1-C mediates cerebral ischemia/reperfusion injury via ER
stress and mitochondria-associated apoptosis pathways. Cell Death
Dis. 8:e30802017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cao G, Zhou H, Jiang N, Han Y, Hu Y, Zhang
Y, Qi J, Kou J and Yu B: Yi Qi Fu Mai powder injection ameliorates
cerebral ischemia by inhibiting endoplasmic reticulum
stress-mediated neuronal apoptosis. Oxid Med Cell Longev.
2016:54932792016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin YW, Chen TY, Hung C, Tai SH, Huang SY,
Chang CC, Hung HY and Lee EJ: Melatonin protects brain against
ischemia/reperfusion injury by attenuating endoplasmic reticulum
stress. Int J Mol Med. 42:182–192. 2018.PubMed/NCBI
|
|
12
|
Chaudhari N, Talwar P, Parimisetty A,
Lefebvre d'Hellencourt C and Ravanan P: A molecular web:
Endoplasmic reticulum stress, inflammation, and oxidative stress.
Front Cell Neurosci. 8:2132014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li X, Fang D, Cong X, Cao G, Cai H and Cai
B: Application of Fourier transform near-infrared spectroscopy
combined with high-performance liquid chromatography in rapid and
simultaneous determination of essential components in crude Radix
Scrophulariae. AAPS PharmSciTech. 13:1428–1435. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang CC, Gu WL, Wu XM, Li YM, Chen CX and
Huang XY: Active components from Radix Scrophulariae inhibits the
ventricular remodeling induced by hypertension in rats.
Springerplus. 5:3582016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li YM, Zeng HW, He X, Jiang YY, Jiang SH
and Zhu DY: Iridoid and phenylpropanoid glycosides of
Scrophularia ningpoensis inhibit the formation of LTB (4)
and platelet aggregation Ti Erh Chun i Ta Hsueh Pao. Acad J Sec Mil
Med Univ. 2:301–303. 1999.
|
|
16
|
Huang Q, Gong QY, Yao MH, Yu R and Shi NC:
Protective effect of Scrophularia ningpoensis extracts on
cerebral ischemia injury in rats. Chin J New Drugs Clini Remedies.
23:323–327. 2004.
|
|
17
|
Kim A, Im M and Jin YM: SRVF, a novel
herbal formula including scrophulariae radix and fructus viticis,
disrupts focal adhesion and causes detachment-induced apoptosis in
malignant cancer cells. Sci Rep. 7:127562017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ying Xl, Zhong XM, Xu MD, Chen MJ, Xiao
WX, Li GZ, Wang H and Huang Z: Neuro-protective effect of harpagide
on acute cerebral ischemic injury in mice and its mechanism
involving mitochondria. Chin Pharm J. 50:1026–1031. 2015.
|
|
19
|
Kilkenny C, Browne WJ, Cuthill IC, Emerson
M and Altman DG: Improving bioscience research reporting: The
ARRIVE guidelines for reporting animal research. PLoS Biol.
8:e10004122010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Leary S, Underwood W, Anthony R, Cartner
S, Corey D, Grandin T, Greenacre C, Gwaltney-Brant S, McCrackin MA,
Meyer R, et al: AVMA guidelines for the euthanasia of animals: 2013
edition. Am Vet Med Assoc. 2013.
|
|
21
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lv MR, Li B, Wang MG, Meng FG, Yu JJ, Guo
F and Li Y: Activation of the PI3K-Akt pathway promotes
neuroprotection of the δ-opioid receptor agonist against cerebral
ischemia-reperfusion injury in rat models. Biomed Pharmacother.
93:230–237. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhai ZY and Feng J: Left-right asymmetry
influenced the infarct volume and neurological dysfunction
following focal middle cerebral artery occlusion in rats. Brain
Behav. 8:e011662018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu F, Schafer DP and McCullough LD: TTC,
fluoro-Jade B and NeuN staining confirm evolving phases of
infarction induced by middle cerebral artery occlusion. J Neurosci
Methods. 179:1–8. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Swanson RA, Morton MT, Tsao-Wu G, Savalos
RA, Davidson C and Sharp FR: A semiautomated method for measuring
brain infarct volume. J Cereb Blood Flow Metab. 10:290–293. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ya BL, Li HF, Wang HY, Wu F, Xin Q, Cheng
HJ, Li WJ, Lin N, Ba ZH, Zhang RJ, et al: 5-HMF attenuates striatum
oxidative damage via Nrf2/ARE signaling pathway following transient
global cerebral ischemia. Cell Stress Chaperones. 22:55–65. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tyagi N, Qipshidze N, Munjal C, Vacek JC,
Metreveli N, Givvimani S and Tyagi SC: Tetrahydrocurcumin
ameliorates homocysteinylated cytochrome-c mediated autophagy in
hyperhomocysteinemia mice after cerebral ischemia. J Mol Neurosci.
47:128–138. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu P, Yang X, Hei C, Meli Y, Niu J, Sun T
and Li PA: Rapamycin reduced ischemic brain damage in diabetic
animals is Associated with suppressions of mTOR and ERK1/2
signaling. Int J Biol Sci. 12:1032–1040. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li W, Yang Y, Hu Z, Ling S and Fang M:
Neuroprotective effects of DAHP and Triptolide in focal cerebral
ischemia via apoptosis inhibition and PI3K/Akt/mTOR pathway
activation. Front Neuroanat. 9:482015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jiang CJ, Wang ZJ, Zhao YJ, Zhang ZY, Tao
JJ and Ma JY: Erythropoietin reduces apoptosis of brain tissue
cells in rats after cerebral ischemia/reperfusion injury: A
characteristic analysis using magnetic resonance imaging. Neural
Regen Res. 11:1450–1455. 2016.PubMed/NCBI
|
|
31
|
Janyou A, Wicha P, Jittiwat J, Suksamrarn
A, Tocharus C and Tocharus J: Dihydrocapsaicin attenuates blood
brain barrier and cerebral damage in focal cerebral
ischemia/reperfusion via oxidative stress and inflammatory. Sci
Rep. 7:105562017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2 (Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Villamil-Ortiz JG and Cardona-Gomez GP:
Comparative analysis of autophagy and tauopathy related markers in
cerebral ischemia and Alzheimer's disease animal models. Fron Aging
Neurosci. 7:842015.
|
|
34
|
Li XB, Ding MX, Ding CL, Li LL, Feng JZ
and Yu XJ: Toll-Like receptor 4 promotes the phosphorylation of
CRMP2 via the activation of Rho-kinase in MCAO rats. Mol Med Rep.
18:342–348. 2018.PubMed/NCBI
|
|
35
|
Caltagirone C, Cisari C, Schievano C, Di
Paola R, Cordaro M, Bruschetta G, Esposito E and Cuzzocrea S;
Stroke Study Group, : Co-ultramicronized
palmitoylethanolamide/luteolin in the treatment of cerebral
ischemia: From rodent to man. Transl Stroke Res. 7:54–69. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ferlito M, Wang Q, Fulton WB, Colombani
PM, Marchionni L, Fox-Talbot K, Paolocci N and Steenbergen C:
Correction: Hydrogen sulfide increases survival during sepsis:
Protective effect of CHOP inhibition. J Immunol. 192:1806–1814.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Miao M, Yan X, Guo L and Shao S: Effects
of the Rabdosia rubescens total flavonoids on focal cerebral
ischemia reperfusion model in rats. Saudi Pharm J. 25:607–614.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jung SY, Kim KM, Cho S, Lim S, Lim C and
Kim YK: Effects of pretreatment with methanol extract of Peucedani
Radix on transient ischemic brain injury in mice. Chin Med.
12:30–40. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang C, Liu M, Pan Y, Bai B and Chen J:
Global gene expression profile of cerebral ischemia-reperfusion
injury in rat MCAO model. Oncotarget. 8:74607–74622. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Caprio FZ and Sorond FA: Cerebrovascular
disease: Primary and secondary stroke prevention. Med Clin North
Am. 103:295–308. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xiao B, Chai Y, Lv S, Ye M, Wu M, Xie L,
Fan Y and Zhu X: Endothelial cell-derived exosomes protect SH-SY5Y
nerve cells against ischemia/reperfusion injury. Int J Mol Med.
40:1201–1209. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhao Q, Wang X, Chen A, Cheng X, Zhang G,
Sun J, Zhao Y, Huang Y and Zhu Y: Rhein protects against cerebral
ischemic-/reperfusion-induced oxidative stress and apoptosis in
rats. Int J Mol Med. 41:2802–2812. 2018.PubMed/NCBI
|
|
43
|
He F, Zhang N, Lv Y, Sun W and Chen H:
Low-dose lipopolysaccharide inhibits neuronal apoptosis induced by
cerebral ischemia/reperfusion injury via the PI3K/Akt/FoxO1
signaling pathway in rat. Mol Med Rep. 19:1443–1452.
2019.PubMed/NCBI
|
|
44
|
Chen S, Sun M, Zhao X, Yang Z, Liu W, Cao
J, Qiao Y, Luo X and Wen A: Neuroprotection of hydroxysafflor
yellow A in experimental cerebral ischemia/reperfusion injury via
metabolic inhibition of phenylalanine and mitochondrial biogenesis.
Mol Med Rep. 19:3009–3020. 2019.PubMed/NCBI
|
|
45
|
Hou Y, Wang Y, He Q, Li L, Xie H, Zhao Y
and Zhao J: Nrf2 inhibits NLRP3 inflammasome activation through
regulating Trx1/TXNIP complex in cerebral ischemia reperfusion
injury. Behav Brain Res. 336:32–39. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Su D, Ma J, Zhang Z, Tian Y and Shen B:
Protective effects of UCF-101 on cerebral ischemia-reperfusion
(CIR) is depended on the MAPK/p38/ERK signaling pathway. Cell Mol
Neurobiol. 36:907–914. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shen X, Eichhorn T, Greten HJ and Efferth
T: Effects of Scrophularia ningpoensis hemsl. On inhibition
of proliferation, apoptosis induction and NF-κB signaling of
immortalized and cancer cell lines. Pharmaceuticals (Basel).
5:189–208. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liang W, Lam WP, Tang HC, Leung PC and Yew
DT: Current evidence of Chinese herbal constituents with effects on
NMDA receptor blockade. Pharmaceuticals (Basel). 6:1039–1054. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang Q, An RD, Tian XC, Yang M, Li MH,
Lou J, Xu L and Dong Z: β-Caryophyllene pretreatment alleviates
focal cerebral ischemia-reperfusion injury by activating PI3K/Akt
signaling pathway. Neurochem Res. 42:1459–1469. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Gong G, Xiang L, Yuan L, Hu L, Wu W, Cai
L, Yin L and Dong H: Protective effect of glycyrrhizin, a direct
HMGB1 inhibitor, on focal cerebral ischemia/reperfusion-induced
inflammation, oxidative stress, and apoptosis in rats. PLoS One.
9:e894502014. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang Y, Ren QY, Zhang X, Lu HL and Chen J:
Neuroprotective mechanisms of calycosin against focal cerebral
ischemia and reperfusion injury in rats. Cell Physiol Biochem.
45:537–546. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liu B, Li F, Shi J, Yang D, Deng Y and
Gong Q: Gastrodin ameliorates subacute phase cerebral
ischemia-reperfusion injury by inhibiting inflammation and
apoptosis in rats. Mol Med Rep. 14:4144–4152. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li X, Guo H, Zhao L, Wang B, Liu H, Yue L,
Bai H, Jiang H, Gao L, Feng D and Qu Y: Adiponectin attenuates
NADPH oxidase-mediated oxidative stress and neuronal damage induced
by cerebral ischemia-reperfusion injury. Biochim Biophys Acta Mol
Basis Dis. 1863:3265–3276. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Song W, Guo F, Zhong H, Liu L, Yang R,
Wang Q and Xiong L: Therapeutic window of globular adiponectin
against cerebral ischemia in diabetic mice: The role of dynamic
alteration of adiponectin/adiponectin receptor expression. Sci Rep.
5:173102015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhao Y, Fang Y, Zhao H, Li J and Luo Y:
Chrysophanol inhibits endoplasmic reticulum stress in cerebral
ischemia and reperfusion mice. Eur J Pharmacol. 818:1–9. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Liu X, Zhao S, Liu F, Kang J, Xiao A, Li
F, Zhang C, Yan F, Zhao H, Luo M, et al: Remote ischemic
postconditioning alleviates cerebral ischemic injury by attenuating
endoplasmic reticulum stress-mediated apoptosis. Transl Stroke Res.
5:692–700. 2014. View Article : Google Scholar : PubMed/NCBI
|