Introduction
Pre-eclampsia (PE) is a pregnancy-specific
complication diagnosed by the onset of high blood pressure and
proteinuria after 20 weeks of gestation, which affects 3–8% of
pregnancies globally (1,2). Despite developments in the diagnosis
and treatment of PE, the pathogenesis of PE requires further
investigation. The abnormal placenta, characterized by impaired
trophoblastic invasion and placental angiogenesis, can serve key
roles in the occurrence and progression of PE (3,4).
Investigating novel mechanisms for regulating placental function
will improve the diagnosis and treatment of PE.
MicroRNAs (miRNAs/miRs) are endogenous small
non-coding single-stranded RNAs, 22–25 nucleotides in length, which
can modulate target gene expression via translational repression or
degradation (5). The dysregulation
of miRNAs contributes to various human diseases (6). Previous expression profiling studies
have demonstrated dysregulated expression of miRNAs in the placenta
of PE (7–9). In addition, miRNAs may be implicated
in the pathogenesis of PE by regulating the function and
development of the placenta (10,11).
Among the differentially expressed miRNAs,
miR-335-5p is significantly upregulated in PE-affected placentas
(9,12). Previous studies have demonstrated
that miR-335-5p may have a negative correlation with
neovascularization in human retinal microvascular endothelial cells
and in patients with age-related macular degeneration (13,14).
In addition, miR-335-5p can serve as a key tumor suppressor to
inhibit tumor invasion, metastasis and proliferation, and to induce
apoptosis in different human cancer types (15). Dysregulated miR-335-5p may be
associated with the pathogenesis of PE; however, the underlying
mechanism regulating the aberrant expression of miR-335-5p in
PE-affected placentas and the role of miR-335-5p in the
pathogenesis of PE are not fully understood. The present in
vitro study preliminarily investigated the role of miR-335-5p
in JEG-3 cells.
Materials and methods
Cell culture
The JEG-3 choriocarcinoma line was purchased from
Nanjing KeyGen Biotech Co., Ltd. Cells were cultured in minimum
essential medium (MEM; Gibco; Thermo Fisher Scientific, Inc.)
containing 10% heat-inactivated FBS (Biological Industries), 100
U/ml penicillin and 100 µg/ml streptomycin at 37°C in a humidified
atmosphere with 5% CO2. Cells were passaged when
confluence reached ~90%.
Cell transfection
JEG-3 cells were seeded in six-well plates at
5×105 cells/well, and transfected with 50 nM miR-335-5p
mimics, p53 small interfering (si)RNA, specificity protein 1 (Sp1)
siRNA or the corresponding non-specific negative controls (NCs)
using the riboFECT™ CP Transfection kit (Guangzhou RiboBio Co.,
Ltd.), according to the manufacturer's protocol. miR-335-5p mimics,
p53 siRNA, Sp1 siRNA and the corresponding NCs were designed and
synthesized by Shanghai GenePharma Co., Ltd. miR-335-5p mimics,
forward 5′-UCAAGAGCAAUAACGAAAAAUGU-3′, reverse
5′-AUUUUUCGUUAUUGCUCUUGAUU-3′; p53 siRNA, forward
5′-GCAUGAACCGGAGGCCCAUTT-3′, reverse 5′-AUGGGCCUCCGGUUCAUGCTT-3′;
Sp1 siRNA, forward 5′-UGAGAACAGCAACAACUCCTT-3′, reverse
5′-GGAGUUGUUGCUGUUCUCATT-3′; and NCs, forward
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse
5′-ACGUGACACGUUCGGAGAATT-3′. The transfected cells were then
collected for western blot analysis or other functional assays
after 24, 48, 72 or 96 h.
Cell proliferation
JEG-3 cells transfected with NCs or miR-335-5p
mimics were seeded at a density of 5×103 cells/well in
96-well culture plates with 100 µl complete medium under normal
conditions. After culturing for 0, 24, 48, 72 and 96 h, an MTT
assay (Beyotime Institute of Biotechnology) was performed to
measure cell proliferation. In total, 10 µl MTT reagent was added
to each well and the plate was incubated at 37°C for 4 h.
Subsequently, 100 µl dimethyl sulfoxide was added to terminate the
reaction. The optical density (OD) was measured at 490 nm using a
microplate reader (Tecan Infinite M200; Tecan Group, Ltd.). The
experiments were performed in triplicate.
Cell apoptosis
An Annexin V-FITC/PI apoptosis detection kit (cat.
no. BB-4101-1, BestBio) was used to assess the cell apoptotic rate
according to the manufacturer's instructions. After JEG-3 cells
were transfected with NCs or miR-335-5p mimics for 48 h, the cells
were collected using trypsin without EDTA, washed twice with cold
PBS and resuspended with 400 µl binding buffer. The cells were
incubated with 5 µl Annexin V-FITC staining solution for 15 min at
4°C and 10 µl PI staining solution for 5 min at 4°C in the dark.
The apoptotic rate was detected using a flow cytometer (BD
FACSCalibur flow cytometer; BD Biosciences). The flow cytometry
data were analyzed with FlowJo software (v10.0.7r2; FlowJo LLC).
The experiments were performed in triplicate.
Transwell migration assay
The effect of miR-335-5p or Sp1 on the migration of
JEG-3 cells was evaluated using a Transwell migration assay in a
24-well Transwell plate containing polycarbonate filters with 8 µm
pores (Costar; Corning, Inc.). After transfection with miR-335-5p
mimics, Sp1 siRNA or the corresponding NCs for 24 h, JEG-3 cells
were trypsinized and adjusted to 1×106 cells/ml in MEM.
In total, 100 µl resuspended cells was placed in the upper chamber
and 600 µl medium containing 10% FBS was added to the lower
chamber. After incubation at 37°C for 24 h under normal conditions,
the membranes were fixed using 100% methanol at room temperature
for 30 min and stained with 0.1% crystal violet stain solution
(cat. no. G1063, Beijing Solarbio Science & Technology Co.,
Ltd.) at room temperature for 20 min. The numbers of migrated cells
were calculated using a light microscope (magnification, ×400;
Olympus IX51; Olympus Corporation) in five random fields. The
experiments were performed in triplicate.
Wound healing assay
The migratory ability of miR-335-5p was further
investigated using a wound healing assay. JEG-3 cells were
transfected with NCs or miR-335-5p mimics. After cells reached 90%
confluence, a 10-µl sterile pipette tip was used to create a wound
in the monolayer. The cells were grown in FBS-free medium for
another 24 h and evaluated using a light microscope (Olympus IX51;
Olympus Corporation) in phase contrast condition, using
magnification, ×100. The wound areas were measured using ImageJ
software (version 1.46r; National Institutes of Health). The
relative wound closure under each condition was calculated
according to the following formula: Wound area measured at 0 h
minus the wound area at 24 h. The wound repair rate was obtained
using the percentage of the relative wound closure within the wound
area measured at 0 h. The experiments were performed in
triplicate.
Western blot analysis
Total proteins were extracted from JEG-3 cells
transfected with miR-335-5p mimics, p53 siRNA, Sp1 siRNA or the
corresponding NCs using RIPA lysis buffer (cat. no. P0013B;
Beyotime Institute of Biotechnology), according to the
manufacturer's instructions. Protein concentration was measured
using an Enhanced Bicinchoninic Acid Protein assay kit (Beyotime
Institute of Biotechnology). A total of 10 µg each sample was
separated by SDS-PAGE on 12% gels according to the assigned groups
and electrophoretically transferred to PVDF membranes (EMD
Millipore). After the membranes were blocked with TBS-0.1% Tween 20
(TBST) containing 5% non-fat dry milk at room temperature for 1 h,
membranes were incubated with primary antibodies against Sp1
(1:1,000; cat. no. ab124804; Abcam), p53 (1:1,000; cat. no.
10442-1-AP; Wuhan Sanying Biotechnology), E-cadherin (1:1,000; cat.
no. 20874-1-AP; Wuhan Sanying Biotechnology), N-cadherin (1:2,000;
cat. no. ab76011; Abcam), Snail (1:1,000; cat. no. 26183-1-AP;
Wuhan Sanying Biotechnology) and GAPDH (1:2,000; cat. no.
10494-1-AP; Wuhan Sanying Biotechnology) at 4°C overnight.
Subsequently, the membranes were washed three times with TBST for
10 min and exposed to horseradish peroxidase-conjugated affiniPure
goat anti-rabbit IgG (H+L; 1:5,000; cat. no. SA00001-2; Wuhan
Sanying Biotechnology) at room temperature for 1 h. Detections were
performed using an enhanced chemiluminescence kit (Wuhan Sanying
Biotechnology). The bands were semi-quantified with densitometry
using ImageJ software. The target protein levels were presented as
fold change normalized to GAPDH in each sample.
H2O2 treatment
and reactive oxygen species (ROS) assay
JEG-3 cells were plated in 6-well plates at
8×105 cells/well and were treated with 0.1 mM
H2O2 at 37°C for 24 h. N-acetyl-cysteine
(NAC), a specific scavenger of ROS, was purchased from
Sigma-Aldrich; Merck KGaA. To investigate the effect of ROS on the
expression of miR-335-5p, JEG-3 cells were treated with 10 mM NAC
at 37°C for 1 h prior to H2O2 treatment. The
intracellular ROS levels induced by H2O2 with
or without NAC were detected with a ROS assay kit (cat. no.
BB-4705; BestBio) according to the manufacturer's instructions.
After the cells were treated with H2O2 with
or without NAC in 6-well plates as aforementioned, the supernatants
were removed and cells were washed three times with PBS. Cells were
incubated in fresh medium with DCFH-DA at 37°C for 20 min. Cells
were collected, washed two times with PBS and analyzed with a flow
cytometer (BD FACSCalibur flow cytometer; BD Biosciences) with the
excitation wavelength at 488 nm and the emission wavelength at 525
nm. The flow cytometry data were analyzed with FlowJo software. The
experiments were performed in triplicate.
Cell viability assay
Cell viability was evaluated using the CCK-8 assay
(Bestbio), according to the manufacturer's protocols. JEG-3 cells
were cultured in a 96-well plate at 5×103 cells/well
with MEM containing 10% FBS. After incubation with 0.1 mM
H2O2 at 37°C for 24 h, 10 µl CCK-8 solution
was added to each well and incubated at 37°C for 3 h. The OD values
were detected at 450 nm using a microplate reader (Tecan Infinite
M200; Tecan Group, Ltd.). Relative cell viability was calculated
using the following formula:
OD(H2O2)/OD(control) ×100%. The experiments
were performed in triplicate.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted using the miRNeasy mini kit
(cat. no. 217004; Qiagen, Inc.), according to the manufacturer's
instructions. Reverse transcription and RT-qPCR for the expression
of miR-335-5p was performed using a Hairpin-it™ microRNA and U6
snRNA Normalization RT-PCR Quantitation kit (Shanghai GenePharma
Co., Ltd.) on an ABI 7700 PCR Instrument (Applied Biosciences;
Thermo Fisher Scientific, Inc.). The conditions used for reverse
transcription reaction according to the manufacturer's protocols
were: 25°C for 30 min, 42°C for 30 min, 85°C for 5 min and hold at
4°C. The following primers were used: hsa-miR-335-5p, forward
5′-CGTCCTCGTCAAGAGCAATAAC-3′, reverse
5′-TATGCTTGTTCTCGTCTCTGTGTC-3′; and hU6, forward
5′-CAGCACATATACTAAAATTGGAACG-3′ and reverse
5′-ACGAATTTGCGTGTCATCC-3′. The following thermocycling conditions
were used for qPCR according to the manufacturer's protocols:
Initial denaturation at 95°C for 3 min; followed by 40 cycles at
95°C for 12 sec and 60°C for 40 sec. U6 snRNA was used as an
internal reference. The expression levels of miR-335-5p were
quantified using the 2−∆∆Cq method (16) according to the manufacturer's
instructions.
Statistical analysis
Statistical analysis was performed using SPSS
Statistics 21.0 software (IBM Corp.). Data are presented as the
mean ± SD from ≥3 independent experiments. An unpaired Student's
t-test was used to compare two groups, and one-way ANOVA followed
by Bonferroni's multiple comparison test was used to compare ≥3
groups for statistical analyses. P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-335-5p does not affect
proliferation or apoptosis of JEG-3 cells
To analyze the effects of miR-335-5p on JEG-3 cells,
miR-335-5p mimics or corresponding non-specific NCs were
transfected into JEG-3 cells. Transfection efficiency of miR-335-5p
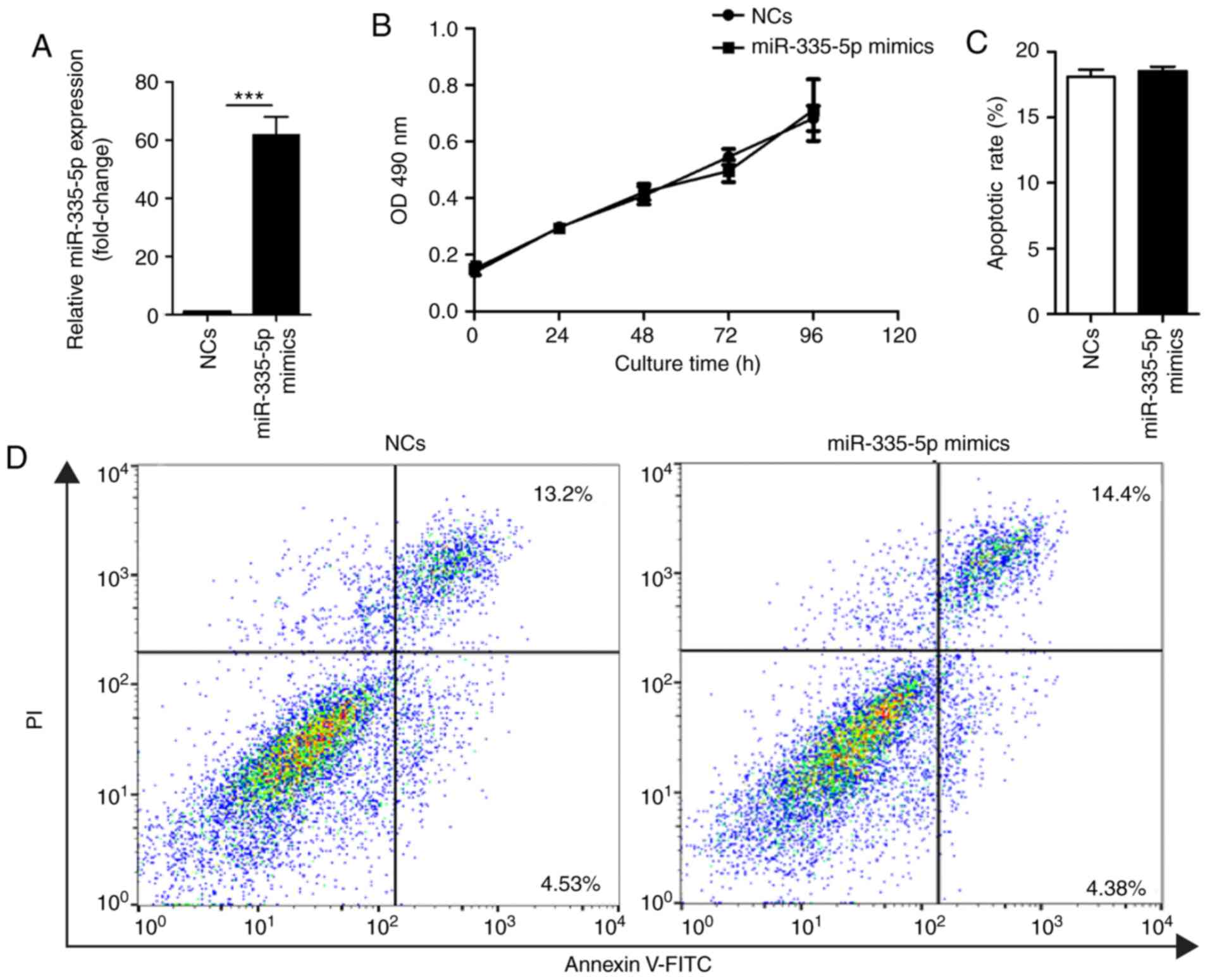
and NCs was confirmed by RT-qPCR (Fig.
1A). MTT assay and Annexin V-FITC/PI assay results suggested
that ectopic overexpression of miR-335-5p did not affect
proliferation (Fig. 1B) or
apoptosis of JEG-3 cells (Fig. 1C and
D).
miR-335-5p significantly suppresses
the migration of JEG-3 cells
The role of miR-335-5p in the migration of JEG-3
cells was investigated using a Transwell cell migration assay and
wound healing assay. Compared with the NCs group, the number of
migrated cells was significantly lower in
miR-335-5p-transfected-JEG-3 cells (43±9 vs. 25±6; P<0.001;
Fig. 2A and B). The wound healing
assay results also showed similar effects (wound repair of the NCs
group, 25.84±4.10%; wound repair of the miR-335-5p mimics group,
11.90±1.48%; P<0.01; Fig. 2C and
D). Epithelial to mesenchymal transition (EMT) is critical for
normal pregnancy and its dysregulation may be associated with
several pregnancy disorders (17,18).
Since EMT is implicated in cell migration, the present study
investigated whether miR-335-5p inhibited the migration of JEG-3
cells via the EMT markers E-cadherin, N-cadherin and Snail. The
expression levels of N-cadherin (P<0.05) and Snail (P<0.01)
were significantly decreased in JEG-3 cells transfected with
miR-335-5p mimics for 48 h compared with those transfected with NCs
(Fig. 2E and F). Conversely,
overexpression of miR-335-5p for 24 h (P<0.05) or 48 h
(P<0.001) significantly increased E-cadherin expression in JEG-3
cells. The expression levels of N-cadherin and Snail exhibited no
significant difference between JEG-3 cells transfected with
miR-335-5p mimics for 24 and 48 h, whereas E-cadherin expression
was significantly increased in the miR-335-5p mimics 48 h group
compared with the 24 h group (P<0.001).
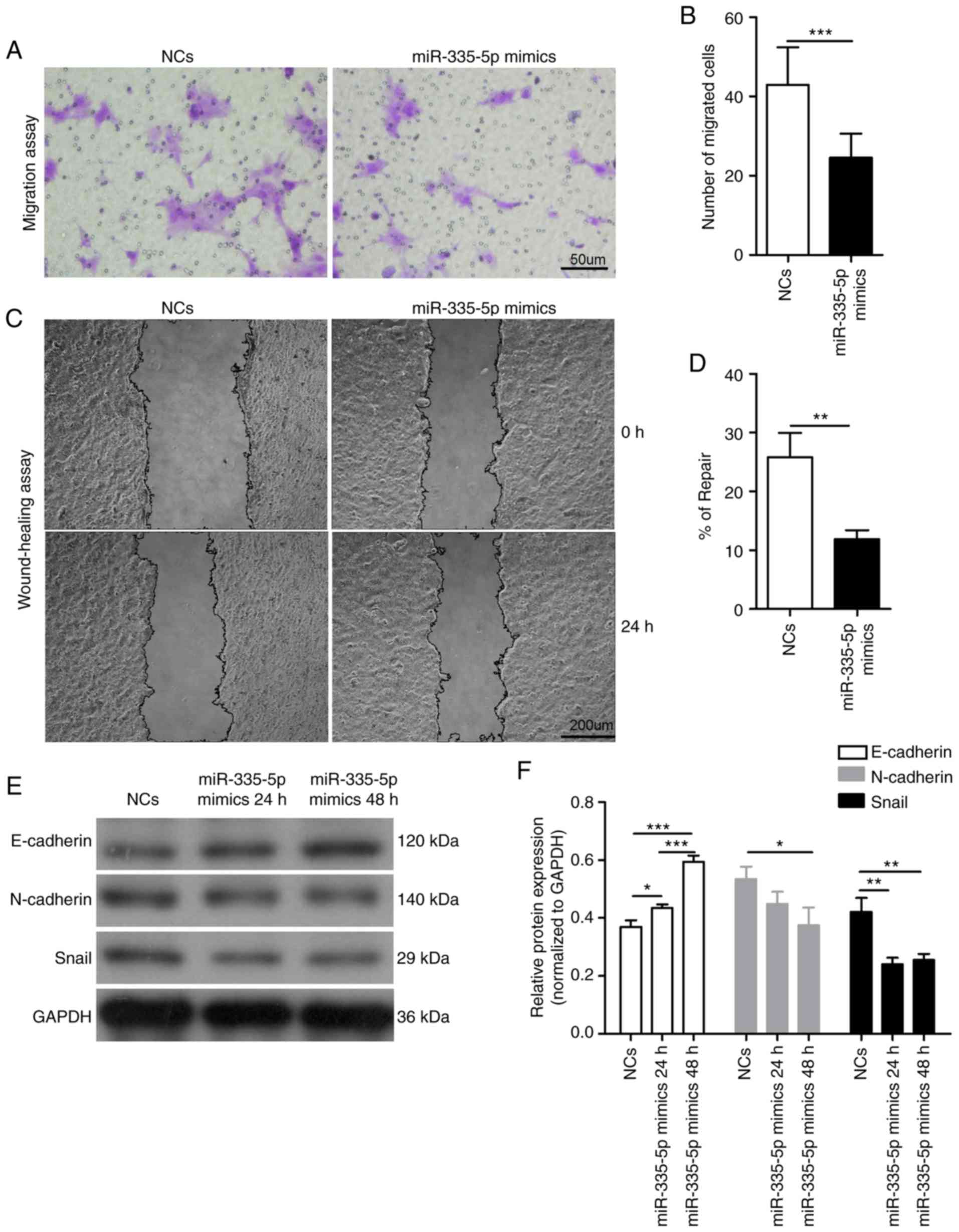 | Figure 2.miR-335-5p significantly suppresses
the migration of JEG-3 cells by modulating epithelial to
mesenchymal transition markers. (A) Representative images of
migrated JEG-3 cells transfected with NCs or miR-335-5p mimics in
the Transwell migration assay. Scale bar, 50 µm. (B) Statistical
analysis of the effect of NCs and miR-335-5p mimics on the
migration of JEG-3 cells. NCs, 43±9 vs. miR-335-5p mimics, 25±6.
n=3. The number of migrated cells was counted using a microscope in
five independent symmetrical visual fields in each independent
experiment. (C) Representative images of the wound healing assay in
JEG-3 cells transfected with NCs or miR-335-5p mimics. Scale bar,
200 µm. (D) Semi-quantification of wound repair. NCs, 25.84±4.10%
vs. miR-335-5p mimics, 11.90±1.48%. n=3. (E) Western blot analysis
of the protein expression levels of E-cadherin, N-cadherin and
Snail in JEG-3 cells transfected with NCs and miR-335-5p mimics.
GAPDH was used as the internal control. (F) Statistical analysis
showing the effect of NCs and miR-335-5p mimics on the protein
expression levels of E-cadherin, N-cadherin and Snail in JEG-3
cells. *P<0.05, **P<0.01, ***P<0.001. miR-335-5p,
microRNA-335-5p; NCs, negative controls. |
Sp1 participates in the inhibitory
role of miR-335-5p on cell migration
Sp1 serves important roles in the progression of EMT
(19,20). A previous study revealed that Sp1
was the target of miR-335-5p in gastric cancer (21). The present study investigated
whether miR-335-5p inhibited the migration of JEG-3 cells via Sp1.
The present western blotting results suggested that overexpression
of miR-335-5p for 24 h (P<0.01) or 48 h (P<0.05)
significantly decreased Sp1 expression in JEG-3 cells (Fig. 3A and B). Compared with the
NCs-transfected cells, JEG-3 cells transfected with a Sp1-specific
siRNA for 24 and 48 h exhibited significantly decreased expression
levels of Sp1, N-cadherin and Snail, whereas the expression levels
of E-cadherin were significantly increased (P<0.05). There was
no difference in the expression levels of E-cadherin, N-cadherin
and Snail between JEG-3 cells transfected with Sp1 siRNA for 24 and
48 h (Fig. 3C and D). In addition,
silencing Sp1 in JEG-3 cells significantly decreased the migratory
ability of cells (NCs, 68±22 vs. Sp1 siRNA, 41±9; P<0.01;
Fig. 3E and F). The present
results suggested that Sp1 may mediate the inhibitory role of
miR-335-5p in cell migration by regulating the expression levels of
EMT markers.
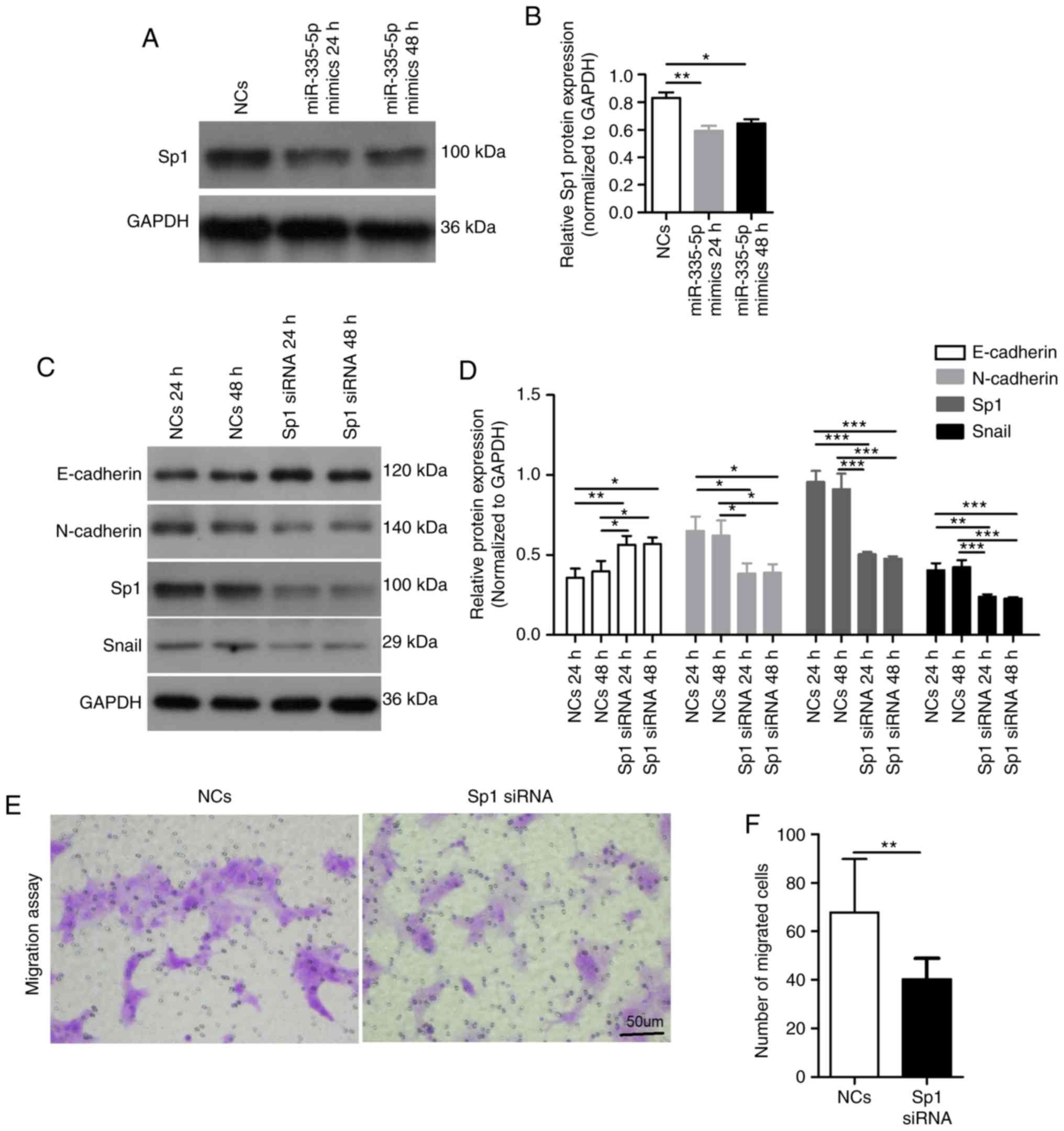 | Figure 3.Sp1 participates in the inhibitory
role of miR-335-5p in cell migration. (A) Western blot analysis of
Sp1 expression in JEG-3 cells transfected with NCs and miR-335-5p
mimics. GAPDH was used as the internal control. (B) Statistical
analysis showing the effect of NCs or miR-335-5p mimics on Sp1
expression in JEG-3 cells. n=3. (C) Western blot analysis of the
protein expression levels of E-cadherin, N-cadherin, Sp1 and Snail
in JEG-3 cells transfected with NCs or Sp1-specific siRNA. (D)
Statistical analysis showing the effect of NCs and Sp1-specific
siRNA on the protein expression levels of E-cadherin, N-cadherin,
Sp1 and Snail in JEG-3 cells. n=3. (E) Representative images of
migrated JEG-3 cells transfected with NCs or Sp1-specific siRNA in
the Transwell migration assay. Scale bar, 50 µm. (F) Statistical
analysis showing the effect of NCs and Sp1-specific siRNA on the
migration of JEG-3 cells; NCs, 68±22 vs. Sp1 siRNA, 41±9. The
number of migrated cells was counted using a microscope in five
independent symmetrical visual fields in each independent
experiment. *P<0.05, **P<0.01, ***P<0.001. miR-335-5p,
microRNA-335-5p; NCs, negative controls; siRNA, small interfering
RNA; Sp1, specificity protein 1. |
Oxidative stress induced by
H2O2 increases miR-335-5p expression
level
Previous studies have shown that oxidative stress
can modulate miRNA expression in cancer via ROS (22,23).
The present study investigated ROS generation and miR-335-5p
expression in JEG-3 cells following H2O2
exposure. The present results suggested that short-term exposure to
0.1 mM H2O2 for 24 h did not affect the
viability of JEG-3 cells (Fig.
4A). Compared with control cells, the intracellular ROS levels
detected using the DCFH-DA assay and miR-335-5p expression levels
identified by RT-qPCR were significantly higher in JEG-3 cells
treated with 0.1 mM H2O2 for 24 h
(P<0.001). Cells pretreated with 10 mM NAC for 1 h prior to
H2O2 treatment had significant decreases in
intracellular ROS levels and miR-335-5p expression compared with
cells exposed to H2O2 only (P<0.001;
Fig. 4B-D).
p53 mediates the ROS-induced increase
of miR-335-5p
miRNA expression can be regulated by ROS via several
important transcription factors, including p53 (22,24).
The present study investigated whether p53 mediated the ROS-induced
increase in expression levels of miR-335-5p in JEG-3 cells.
Compared with in NCs-transfected cells, a p53-specific siRNA
significantly decreased the expression levels of p53 after
transfection for 24 and 48 h (P<0.001; Fig. 5A and B). Silencing p53 in JEG-3
cells significantly reduced miR-335-5p expression and significantly
reversed the increased expression of miR-335-5p induced by 0.1 mM
H2O2 exposure for 24 h (P<0.001; Fig. 5C).
Discussion
Several miRNAs are involved in the regulation of
normal pregnancy and pregnancy complications, such as PE (10,11).
Identifying the mechanism underlying dysregulated miRNA expression
and the roles of miRNA will help understand the etiology of PE
(10,11).
miR-335-5p is downregulated in various types of
human cancer, and is associated with tumorigenesis and cancer
progression (15). Previous
studies have reported that miR-335-5p is upregulated in PE
placentas (9,12). The present study investigated the
role of miR-335-5p in the proliferation of JEG-3 cells. The present
results suggested that miR-335-5p had no obvious effect on the
proliferation or apoptosis of JEG-3 cells. Similarly, in previous
studies, miR-335-5p did not regulate proliferation of gastric
cancer cells and gastrin-releasing peptide receptor-silenced
neuroblastoma cells (21,25). However, in another previous study,
miR-335-5p inhibited the proliferation of granulosa cells via SGK3
in polycystic ovary syndrome (26). The differences in cell source
organizations may lead to the inconsistent evidence of the role of
miR-335-5p.
miR-335-5p serves crucial roles in the migration and
invasion of various types of human cancer by regulating different
target genes (15). miR-335-5p can
directly or indirectly modulate the EMT process, which serves as
the driver of migration and invasion in cancer (27,28).
During placental implantation, the transformation of villous
cytotrophoblasts to extravillous trophoblasts is similar to the
process of EMT (18,29). The dysregulation of this progress
is associated with pregnancy complications (17,18).
Previous studies have reported that E-cadherin expression is
markedly increased in placenta tissues from patients with PE
compared with controls, whereas the expression levels of N-cadherin
and Snail are decreased (30–32).
On the basis of these previous studies, the present study
investigated the effects of miR-335-5p on JEG-3 cell migration and
key EMT markers. The present results suggested that overexpression
of miR-335-5p significantly inhibited JEG-3 cell migration,
suppressed the expression levels of N-cadherin and Snail, and
increased E-cadherin expression. The present results were
consistent with a previous study on the regulation of miR-335-5p on
EMT in colorectal cancer cells (27). The present results suggested that
miR-335-5p may affect the migration of trophoblast cells by
regulating E-cadherin, N-cadherin and Snail.
Sp1 participates in the process of EMT by regulating
the expression of EMT markers (19,20,33).
A previous study identified that in gastric cancer, Sp1 was a
target of miR-335-5p (21).
Another previous study revealed that Sp1 expression was
significantly lower in PE placenta tissues compared with normal
controls (34), which was contrary
to the expression tendency of miR-335-5p in PE placental tissues
(9,12). The present study investigated the
role of Sp1 in the relationship between miR-335-5p and EMT in JEG-3
cells. The present results suggested that miR-335-5p negatively
regulated Sp1 expression levels in JEG-3 cells, and Sp1 knockdown
significantly inhibited the EMT process and migration of JEG-3
cells. The present results suggested that Sp1 may participate in
the inhibitory role of miR-335-5p in cell migration via regulation
of the EMT process.
The dysregulation of oxidative stress is crucial for
the physiopathology of PE (35,36).
Oxidative stress can modulate miRNA expression in cancer and
placental alterations via ROS (22,23,37).
A previous study demonstrated that the miRNA profile containing
miR-335-5p was significantly altered in villous 3A cytotrophoblast
cells exposed to short-term low concentrations of
H2O2 (38).
However, the relationship between ROS and miR-335-5p is not fully
understood. The present results suggested that intracellular ROS
generation and miR-335-5p expression in JEG-3 cells showed a
similar tendency following exposure to only
H2O2 or to NAC pretreatment followed by
H2O2 exposure. Both ROS and miR-335-5p were
increased in JEG-3 cells by H2O2 treatment,
whereas the application of NAC reduced this increase. The present
results suggested there may be a close link between ROS and
miR-335-5p.
The transcription factor p53 is associated with
cellular stress and can be regulated by ROS (39). In addition, p53 has pivotal roles
in the production of stress-induced miRNAs (22). p53 is involved in the upregulation
of miR-200c expression in human umbilical vein endothelial cells
after H2O2 exposure (24). miR-335-5p expression can also be
increased in a p53-dependent manner in several human cancer cells
(40). Notably, the protein
expression, but not mRNA expression of p53 has been reported to be
significantly higher in PE placentas compared with normal controls
(41). The present study
investigated the relationship between ROS, p53 and miR-335-5p. The
present results suggested that p53 knockdown reduced miR-335-5p
expression in JEG-3 cells and decreased the upregulation of
miR-335-5p upon H2O2 exposure. The present
results suggested that p53 may mediate ROS-induced upregulation of
miR-335-5p in JEG-3 cells.
The main limitation of the present study was that
the exact location and expression levels of p53, miR-335-5p, Sp1,
E-cadherin, N-cadherin and Snail in PE and normal placenta tissues
were not investigated. Future studies will need to analyze these
factors. Besides the roles in regulating EMT process and migration,
a previous study reported that the targets of miR-335-5p
participated in the regulation of angiogenesis, as determined using
Gene Ontology enrichment analysis (13). Another study demonstrated that the
vascular endothelial growth factor (VEGF)-A gene was the predicted
target of miR-335-5p by using Mirwalk and Mirtarbase analyses
(14). Since VEGF signaling in
general, and the levels of VEGF-A and sVEGFR1 in particular, have
been implicated in the pathogenesis of PE (4), the potential role of miR-335 in VEGF
signaling requires further investigation.
In summary, the present results suggested that
miR-335-5p expression levels in trophoblast cells may be increased
by ROS in a p53-dependent manner. In addition, miR-335-5p may
inhibit cell migration and EMT via the downregulation of Sp1. Since
the JEG-3 cell line has been widely used as a cell model for the
function of trophoblasts (42,43),
the present in vitro experiments may provide novel evidence
for the etiology of PE and a potential new therapeutic strategy for
patients with PE, particularly for those with dysregulated EMT.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Shandong
Provincial Natural Science Foundation, China (grant nos. ZR2016HM02
and ZR2019BC059).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL carried out experiments and drafted the
manuscript. YYM participated in the study design and data analysis.
QQS, JL, TTQ and YH participated in cell culture, operation of the
microscope and flow cytometry. QJW conceived the study, and
participated in its design, coordination and proofreading. All
authors have participated in the work, read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Steegers EA, von Dadelszen P, Duvekot JJ
and Pijnenborg R: Pre-eclampsia. Lancet. 376:631–644. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Anderson UD, Olsson MG, Kristensen KH,
Akerstrom B and Hansson SR: Review: Biochemical markers to predict
preeclampsia. Placenta. 33 (Suppl):S42–S47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Matsuo K, Kooshesh S, Dinc M, Sun CC,
Kimura T and Baschat AA: Late postpartum eclampsia: Report of two
cases managed by uterine curettage and review of the literature. Am
J Perinatol. 24:257–266. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Powe CE, Levine RJ and Karumanchi SA:
Preeclampsia, a disease of the maternal endothelium: The role of
antiangiogenic factors and implications for later cardiovascular
disease. Circulation. 123:2856–2869. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mayor-Lynn K, Toloubeydokhti T, Cruz AC
and Chegini N: Expression profile of microRNAs and mRNAs in human
placentas from pregnancies complicated by preeclampsia and preterm
labor. Reprod Sci. 18:46–56. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Choi SY, Yun J, Lee OJ, Han HS, Yeo MK,
Lee MA and Suh KS: MicroRNA expression profiles in placenta with
severe preeclampsia using a PNA-based microarray. Placenta.
34:799–804. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu Y, Li P, Hao S, Liu L, Zhao J and Hou
Y: Differential expression of microRNAs in the placentae of Chinese
patients with severe pre-eclampsia. Clin Chem Lab Med. 47:923–929.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen DB and Wang W: Human placental
microRNAs and preeclampsia. Biol Reprod. 88:1302013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hayder H, O'Brien J, Nadeem U and Peng C:
MicroRNAs: Crucial regulators of placental development.
Reproduction. 155:R259–R271. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jiang F, Li J, Wu G, Miao Z, Lu L, Ren G
and Wang X: Upregulation of microRNA335 and microRNA584 contributes
to the pathogenesis of severe preeclampsia through downregulation
of endothelial nitric oxide synthase. Mol Med Rep. 12:5383–5390.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Walz JM, Wecker T, Zhang PP, Cakir B,
Gruening B, Agostini H, Reuer T, Ludwig F, Boneva S, Faerber L, et
al: Impact of angiogenic activation and inhibition on miRNA
profiles of human retinal endothelial cells. Exp Eye Res.
181:98–104. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ertekin S, Yildirim O, Dinc E, Ayaz L,
Fidanci SB and Tamer L: Evaluation of circulating miRNAs in wet
age-related macular degeneration. Mol Vis. 20:1057–1066.
2014.PubMed/NCBI
|
|
15
|
Luo LJ, Wang DD, Wang J, Yang F and Tang
JH: Diverse roles of miR-335 in development and progression of
cancers. Tumour Biol. Oct 8–2016.(Epub ahead of print). View Article : Google Scholar
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ji L, Brkic J, Liu M, Fu G, Peng C and
Wang YL: Placental trophoblast cell differentiation: Physiological
regulation and pathological relevance to preeclampsia. Mol Aspects
Med. 34:981–1023. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kokkinos MI, Murthi P, Wafai R, Thompson
EW and Newgreen DF: Cadherins in the human
placenta-epithelial-mesenchymal transition (EMT) and placental
development. Placenta. 31:747–755. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jungert K, Buck A, von Wichert G, König A,
Buchholz M, Gress TM and Ellenrieder V: Sp1 is required for
transforming growth factor-beta-induced mesenchymal transition and
migration in pancreatic cancer cells. Cancer Res. 67:1563–1570.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peng M, Hu Y, Song W, Duan S, Xu Q, Ding
Y, Geng J and Zhou J: MIER3 suppresses colorectal cancer
progression by down-regulating Sp1, inhibiting
epithelial-mesenchymal transition. Sci Rep. 7:110002017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu Y, Zhao F, Wang Z, Song Y, Luo Y, Zhang
X, Jiang L, Sun Z, Miao Z and Xu H: MicroRNA-335 acts as a
metastasis suppressor in gastric cancer by targeting Bcl-w and
specificity protein 1. Oncogene. 31:1398–1407. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
He J and Jiang BH: Interplay between
reactive oxygen species and MicroRNAs in Cancer. Curr Pharmacol
Rep. 2:82–90. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bao B, Azmi A, Li Y, Ahmad A, Ali S,
Banerjee S, Kong D and Sarkar FH: Targeting CSCs in tumor
microenvironment: The potential role of ROS-associated miRNAs in
tumor aggressiveness. Curr Stem Cell Res Ther. 9:22–35. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Magenta A, Cencioni C, Fasanaro P,
Zaccagnini G, Greco S, Sarra-Ferraris G, Antonini A, Martelli F and
Capogrossi MC: miR-200c is upregulated by oxidative stress and
induces endothelial cell apoptosis and senescence via ZEB1
inhibition. Cell Death Differ. 18:1628–1639. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Qiao J, Lee S, Paul P, Theiss L, Tiao J,
Qiao L, Kong A and Chung DH: miR-335 and miR-363 regulation of
neuroblastoma tumorigenesis and metastasis. Surgery. 154:226–233.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yao L, Li M, Hu J, Wang W and Gao M:
MiRNA-335-5p negatively regulates granulosa cell proliferation via
SGK3 in PCOS. Reproduction. 2018.(Epub ahead of print). View Article : Google Scholar :
|
|
27
|
Wang J, Wang X, Liu F and Fu Y:
microRNA-335 inhibits colorectal cancer HCT116 cells growth and
epithelial-mesenchymal transition (EMT) process by targeting
Twist1. Pharmazie. 72:475–481. 2017.PubMed/NCBI
|
|
28
|
Zhou XM, Sun R, Luo DH, Sun J, Zhang MY,
Wang MH, Yang Y, Wang HY and Mai SJ: Upregulated TRIM29 promotes
proliferation and metastasis of nasopharyngeal carcinoma via
PTEN/AKT/mTOR signal pathway. Oncotarget. 7:13634–13650.
2016.PubMed/NCBI
|
|
29
|
DaSilva-Arnold S, James JL, Al-Khan A,
Zamudio S and Illsley NP: Differentiation of first trimester
cytotrophoblast to extravillous trophoblast involves an
epithelial-mesenchymal transition. Placenta. 36:1412–1418. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li XL, Dong X, Xue Y, Li CF, Gou WL and
Chen Q: Increased expression levels of E-cadherin, cytokeratin 18
and 19 observed in preeclampsia were not correlated with disease
severity. Placenta. 35:625–631. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Du L, Kuang L, He F, Tang W, Sun W and
Chen D: Mesenchymal-to-epithelial transition in the placental
tissues of patients with preeclampsia. Hypertens Res. 40:67–72.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fedorova L, Gatto-Weis C, Smaili S,
Khurshid N, Shapiro JI, Malhotra D and Horrigan T: Down-regulation
of the transcription factor snail in the placentas of patients with
preeclampsia and in a rat model of preeclampsia. Reprod Biol
Endocrinol. 10:152012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee MS, Byun HJ, Lee J, Jeoung DI, Kim YM
and Lee H: Tetraspanin CD82 represses Sp1-mediated Snail expression
and the resultant E-cadherin expression interrupts nuclear
signaling of β-catenin by increasing its membrane localization.
Cell Signal. 52:83–94. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He P, Chen Z, Sun Q, Li Y, Gu H and Ni X:
Reduced expression of 11β-hydroxysteroid dehydrogenase type 2 in
preeclamptic placentas is associated with decreased PPARgamma but
increased PPARalpha expression. Endocrinology. 155:299–309. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu F, Tian FJ, Lin Y and Xu WM: Oxidative
Stress: Placenta function and dysfunction. Am J Reprod Immunol.
76:258–271. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Aouache R, Biquard L, Vaiman D and
Miralles F: Oxidative stress in preeclampsia and placental
diseases. Int J Mol Sci. 19:14962018. View Article : Google Scholar :
|
|
37
|
Rudov A, Balduini W, Carloni S, Perrone S,
Buonocore G and Albertini MC: Involvement of miRNAs in placental
alterations mediated by oxidative stress. Oxid Med Cell Longev.
2014:1030682014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cross CE, Tolba MF, Rondelli CM, Xu M and
Abdel-Rahman SZ: Oxidative stress alters miRNA and gene expression
profiles in villous first trimester trophoblasts. Biomed Res Int.
2015:2570902015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu B, Chen Y and St Clair DK: ROS and
p53: A versatile partnership. Free Radic Biol Med. 44:1529–1535.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Scarola M, Schoeftner S, Schneider C and
Benetti R: miR-335 directly targets Rb1 (pRb/p105) in a proximal
connection to p53-dependent stress response. Cancer Res.
70:6925–6933. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sharp AN, Heazell AE, Baczyk D, Dunk CE,
Lacey HA, Jones CJ, Perkins JE, Kingdom JC, Baker PN and Crocker
IP: Preeclampsia is associated with alterations in the p53-pathway
in villous trophoblast. PLoS One. 9:e876212014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Monteiro LJ, Cubillos S, Sanchez M,
Acuña-Gallardo S, Venegas P, Herrera V, Lam EW, Varas-Godoy M and
Illanes SE: Reduced FOXM1 expression limits trophoblast migration
and angiogenesis and is associated with preeclampsia. Reprod Sci.
26:580–590. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Niu ZR, Han T, Sun XL, Luan LX, Gou WL and
Zhu XM: MicroRNA-30a-3p is overexpressed in the placentas of
patients with preeclampsia and affects trophoblast invasion and
apoptosis by its effects on IGF-1. Am J Obstet Gynecol.
218:249.e1–249.e12. 2018. View Article : Google Scholar
|