Introduction
Inflammatory bowel disease (IBD) is a chronic,
relapsing, non-specific inflammatory intestinal condition, which is
characterized by acute exacerbation followed by remission. IBD
includes ulcerative colitis (UC) and Crohn's disease (CD). UC is a
chronic inflammatory disorder of the colon, which causes bloody
diarrhea, abdominal pain and weight loss (1). The underlying pathogenic mechanisms
of IBD remain unclear, although they have been attributed partially
to dysregulation of immune reactions that respond to intestinal
flora, and to genetic and environmental factors (1).
Dextran sulfate sodium (DSS)-induced colitis is a
well-established experimental model that presents with various
signs and symptoms of human UC, including diarrhea, weight loss,
bloody stools, mucosal ulceration, and shortening of the large
intestine (2). Previous research
has shown that mice with acute DSS-induced colitis display similar
expression profiles of cytokines and histological changes to those
observed in human IBD, particularly UC (2).
The incidence of IBD in China is increasing annually
(3). The role of intestinal
microbiota in the pathophysiology of IBD has recently garnered
attention. Previous studies have demonstrated the intestinal
microbiota serves an important role in gut inflammation, as
follows: i) In the cluster of differentiation 45Rbhigh
transfer IBD model, the transfer of naive helper T cells to
Rag2−/− mice leads to microbiota-dependent intestinal
inflammation, whereas in germ-free mice, transfer of naive helper T
cells do not develop colitis (4,5). ii)
It has been reported that fecal stream diversion can improve
intestinal inflammation in CD (6).
iii) Antibiotics are effective in the treatment of IBD to some
extent (7). iv) Antibiotics, such
as ciprofloxacin and metronidazole, are also available for anal
lesions and the prevention of recurrence in CD (8). v) Many IBD susceptibility genes are
involved in the identification and processing of microbiota
(9). However, the role of normal
intestinal flora in the recovery of colitis has received little
attention, and has not been reported in the literature. The present
study investigated this topic and aimed to clarify the role of
intestinal E. coli in the recovery process of colitis. The
results indicated that the presence of normal intestinal flora may
be a necessary condition for the recovery of colitis, potentially
through activation of the Toll-like receptor 4 (TLR4)/nuclear
factor-κB (NF-κB) signaling pathway, which provides evidence for
the use of intestinal flora to treat IBD.
Materials and methods
Reagents and animals
DSS solution (3.5%; Sigma Healthcare, Melbourne,
Australia) was dissolved in sterile, distilled water and freshly
prepared every other day (10).
Antibiotic solutions were composed of kanamycin (8 mg/ml),
gentamicin (0.7 mg/ml), polymyxin 34,000 U/ml, metronidazole (4.3
mg/ml) and vancomycin (0.9 mg/ml).
A total of 40 adult BALB/c (8-week-old females;
19.20±2 g), 10 C57BL/6 (8-week-old females; 18.8±2 g) and 10
TLR4−/− mice (8-week-old females; 19±2 g) were purchased
from the Animal Experimental Center of Guangdong Academy of Medical
Sciences (Guangzhou, China). Animals were maintained under standard
conditions, and fed rodent food and water, according to the Guide
for the Care and Use of Laboratory Animals (11). The mice were maintained as follows:
Temperature, 20–22°C; relative humidity, 55±5%, 12-h light/dark
cycle, and ad libitum access to food and water, Mice were
fed in cages containing an average of six mice/cage. The protocol
was approved by the Review Board of the Institute of Medical Animal
Laboratory at the Guangzhou Medical University (Guangzhou,
China).
DSS-induced colitis model and
bacteria-depleted (BD) mice
Mice were administered 3.5% DSS in drinking water
for 5 days, followed by normal drinking water for 14 days,
according to a method described previously by Chen et al
(10).
Prior to the experiment, 6-week-old adult mice were
depleted from intestinal bacteria by administration of antibiotic
solution (100 µl/day/mouse) for 2 weeks. Fresh mice fecal samples
were harvested daily and cultured in aerobic and anaerobic
conditions. When bacterial growth could not be detected in the
culture media, the mice were classified as BD mice.
Group classification
The mice were randomly divided into 4 groups as
follows: i) Group A, normal drinking water; ii) group B,
DSS-induced colitis only; iii) group C, DSS-induced colitis in BD
mice; and iv) group D, DSS-induced colitis in BD mice treated with
E. coli (fed E. coli 1×109 CFU once every
other day feeding). The animals received treatments for 14 days.
The experiments were carried out using BALB/c mice, and
subsequently repeated with C57BL/6 and TLR4−/− mice.
In the TLR4−/− experiment, the mice were
randomly divided into three groups as follows: i) Control group
(Group A1); ii) Group B1, DSS-induced colitis mice in BD C57BL/6
mice treated with E. coli as described previously in Group
D; Group C1, DSS-induced colitis mice in BD TLR4−/− mice
treated with E. coli as described previously in Group D.
Outcome measures
Clinical indicators
Animal body weight, stool characteristics and occult
blood were recorded and evaluated daily. These parameters were
scored by a trained observer blinded to the protocol, in order to
calculate disease activity index (DAI), as described in Table I. The mice were sacrificed on day
14 using CO2, and total colon specimens were collected.
The samples were stained with hematoxylin and eosin (H&E),
which was used for the pathological score evaluation of the colon
tissues. The histological severity of colitis was classified into
mucosal damage (D) and extent of disease (E), according to the
scoring criteria proposed by Cooper et al (12). The criteria were as follows: D: 0
points, none; 1 point: 1/3 of the crypt near the basement membrane
was lost; 2 points, 2/3 of the crypt near the basement membrane was
lost; 3 points, all crypts were destroyed, leaving only the surface
mucosal epithelium; 4 points: Crypt epithelium was lost. E: 0
points, none; 1 point, focal lesions; 2 points, lesions were
present in ~1/3 mucosa; 3 points, lesions were present in 2/3
mucosa; 4 points, lesions were present in all mucosal tissues. The
histological score (HS) is equal to the product of D and E, i.e.,
HS=D × E.
 | Table I.DAI scoring system. |
Table I.
DAI scoring system.
| Score | Stool | Hematochezia | Weight loss
(%) |
|---|
| 0 | Normal form | Occult blood
negative | 0 |
| 1 |
|
| 1-5 |
| 2 | Loose and not
forming | Occult blood
positive | 5-10 |
| 3 |
|
| 10-15 |
| 4 | Watery stool | Eye blood | >15 |
Histopathology
The distal ileum, colon and rectum were removed and
cleaned with cold sterile saline. The end of the colon (1 cm from
anus) was divided into 0.5 cm sections. The histological specimens
were fixed in 10% neutral formalin at room temperature for 12 h,
paraffin embedded, sliced and stained with hematoxylin (Mayer) for
5 min and 0.5% eosin for 1–3 min at room temperature (H&E
staining). A blinded method was used in order to score the
pathological sections according to the severity of colitis, as
previously described (13).
Colonic myeloperoxidase (MPO)
detection
Tissue MPO activity was determined using the MPO
Peroxidation Assay kit (cat. no. KA1338; Abnova, Taipei, Taiwan),
as described previously (14). MPO
is an index used to evaluate neutrophil infiltration (15). Colon tissue weights were measured
prior to cooling in liquid nitrogen and stored at −80°C. The
tissues were homogenized four times (5 sec each) with 10 sec
intervals by ultrasonic pulverization (14 kHz) at 4°C, and the
supernatant was removed by centrifugation at 14,000 × g for 20 min
at 4°C. After 5 min, the absorbance value was measured at 460 nm to
determine the MPO activity of each sample.
Immunohistochemical method for the
detection of NF-κB
The application of the standard three-step antibody
method was used to detect NF-κB expression levels (16,17).
Antibody I [NF-κB p65 (F-6), cat. no. sc-8008; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA] was a monoclonal antibody
specific for the NF-κΒ p65 B subunit. Antibody II [goat anti-mouse
immunoglobulin G (IgG)-B: sc-2039, Santa Cruz Biotechnology, Inc.]
was a biotin-conjugated mouse IgG antibody specific for the goat
anti-mouse epitope. Antibody I (10 mg/ml) was incubated with the
sections for 1 h in a humid chamber at room temperature.
Subsequently, Antibody II (2 mg/ml) was incubated with the sections
for 1 h in a humid chamber at room temperature. Finally, the
sections were incubated with avidin-biotin complex reagent
containing horseradish peroxidase (cat. no. 554058; BD Biosciences,
Franklin Lakes, NJ, USA) for 30 min at room temperature. NF-κB
activation was scored at ×400 magnification using a fluorescence
microscope (IX71; Olympus Corporation, Tokyo, Japan) with
visualization of red fluorescence in five fields of view and a mean
value was calculated. NF-κB was scored according to the percentage
of positive cells, as described previously (10): 0 points, 0–1%; 1 point, 2–5%; 2
points, 6–10%; 3 points, 11–25%.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) for the detection of TLR4
expression
RNA was extracted using MiniBEST Universal RNA
Extraction kit (cat. no. 9767; Takara Biotechnology Co., Ltd.,
Dalian, China), SuperScript™ IV First Chain Synthesis system (cat.
no. 18091050; Thermo Fisher Scientific, Inc., Waltham, MA, USA) was
used for RT, and TaqMan™ Fast Advanced Master Mix (cat. no.
4444557; Thermo Fisher Scientific, Inc.) was used for PCR. Total
RNA was isolated from colon tissues and qPCR was performed as
previously described (18,19). Gene expression was normalized to
GAPDH using the 2−ΔΔCq method (20). The primers used were as follows:
GAPDH (253 bp), 5′-ACAGCAACAGGGTGGTGGAC-3′ (forward) and
5′-TTTGAGGGTGCAGCGAACTT-3′ (reverse); TLR4 (239 bp),
5′-CCAGAGCCGTTGGTGTATCT-3′ (forward) and 5′-TCAAGGCTTTTCCATCCAAC-3′
(reverse); and NF-κB p65 (251 bp), 5′-GGCAGCACTCCTTATCAACC-3′
(forward) and 5′-GAGGTGTCGTCCCATCGTAG-3′ (reverse). The data were
representative of at least three independent experiments.
Statistical analysis
The results were analyzed using GraphPad statistical
software (v.5.0; GraphPad Software, Inc., La Jolla, CA, USA).
Experiments were repeated three times. The results were presented
as the mean ± standard deviation. One-way analysis of variance
followed by Bonferroni's correction was used for comparisons
between groups. Kruskal-Wallis test and Steel-Dwass test were
performed to compare histological scores among the test groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Clinical findings
The body weight of mice decreased following
ingestion of DSS. When DSS was ceased after 5 days, the weight of
the mice slightly decreased until day 6. In the BALB/c experiment,
the non-E. coli BD treatment group mice continued to
decrease in weight after DSS treatment was ceased. The body weight
of the mice in the E. coli treatment group declined slightly
(Fig. 1A). In the experiment using
TLR4−/− mice, the E. coli treatment BD
TLR4−/− group retained the weight decrease. The body
weight of the E. coli-treated DSS-induced BD C57BL/6 mice
declined slightly, but increased following the cessation of DSS
treatment (Fig. 1B).
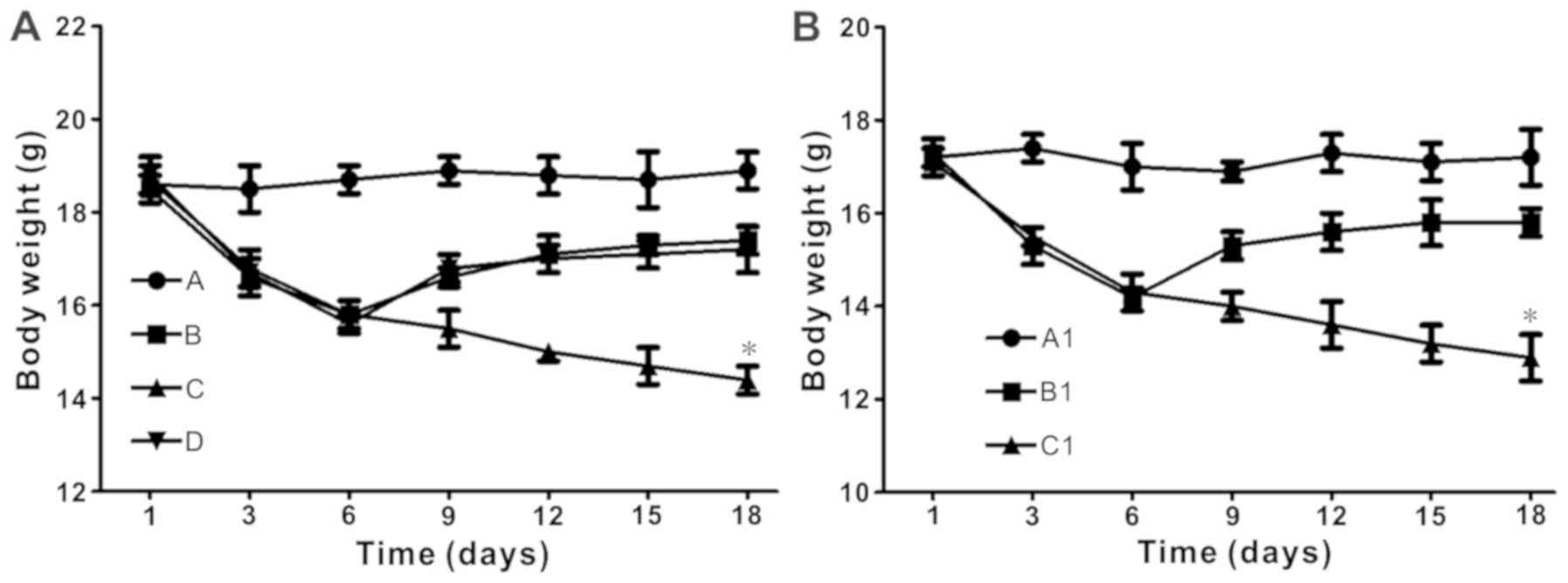 | Figure 1.Escherichia coli treatment
significantly increases body weight in DSS-induced BD mice, but not
in TLR4−/− mice. (A) Body weight over time in BALB/c
mice. *P<0.05 vs. group D at day 18. Group A, control; Group B,
DSS; Group C, DSS + BD; Group D, DSS + BD + E. coli. (B)
Body weight over time in the TLR4−/− mice experiment.
*P<0.05 vs. group B1 at day 18. Group A1, control (C57); Group
B1. DSS + BD + E. coli (C57); Group C1, DSS + BD + E.
coli (TLR4−/−). DSS, dextran sulfate sodium; BD,
bacteria-depleted; TLR4, Toll-like receptor 4. |
In BALB/c mice, the survival rate at the end of the
experiment was 100% for groups A, B and D. Group C exhibited a
survival rate of 60% (Fig. 2A),
suggesting that bacterial depletion followed by DSS-induced colitis
resulted in increased mortality. In the experiment using C57BL/6
and TLR4−/− mice, Groups A1 and B1 had a survival rate
of 100%, whereas C1 had a survival rate of 60%, suggesting that
treatment with E. coli could promote recovery in BD
wild-type mice, but not in TLR4−/− mice. This finding
demonstrated that E. coli may have promoted recovery through
the TLR4 pathway (Fig. 2B).
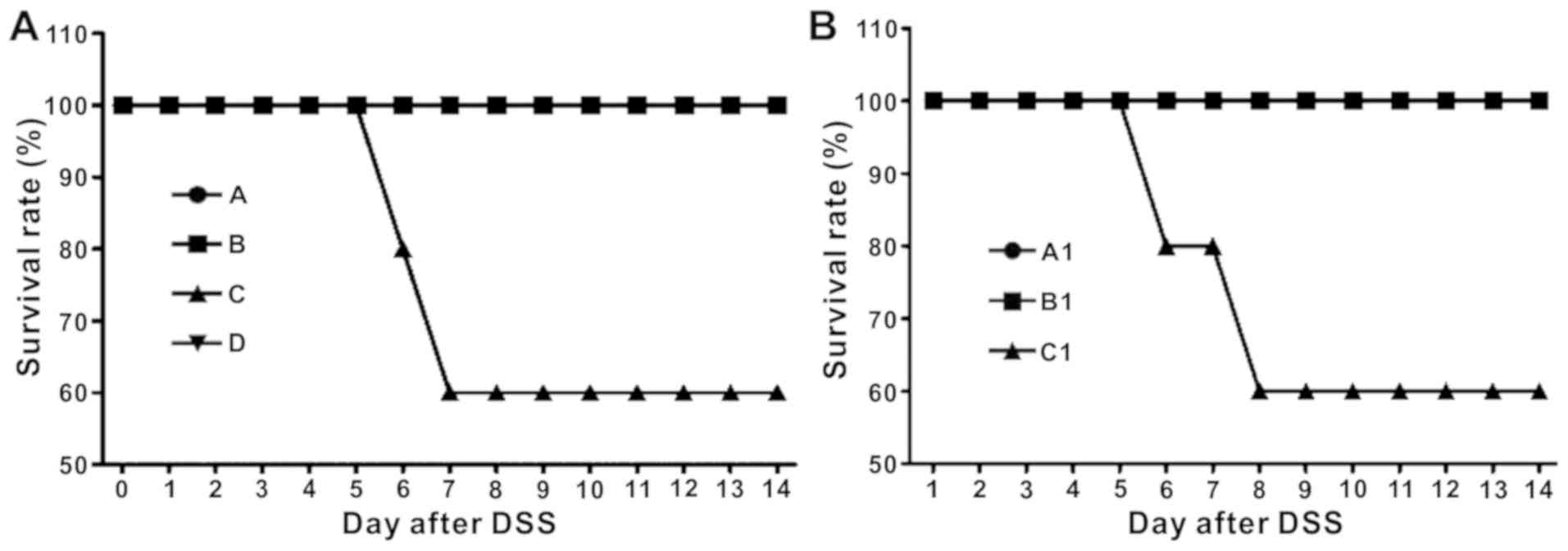 | Figure 2.Survival rate is decreased in
DSS-induced BD BALB/c mice and DSS-induced BD TLR4−/−
mice treated with Escherichia coli. (A) Survival rate over
time in BALB/c mice (n=10/group). At the end of the experiment, the
survival rate of groups A, B and D was 100%, whereas the survival
rate of group C was 60%. Group A, control; Group B, DSS; Group C,
DSS + BD; Group D, DSS + BD + E. coli. (B) Survival rate
over time in the TLR4−/− mice experiment (n=10/group).
The survival rate of groups A1 and B1 was 100%, whereas the
survival rate of group C1 was 60%. Group A1, control (C57); Group
B1, DSS + BD + E. coli (C57); Group C1, DSS + BD + E.
coli (TLR4−/−). DSS, dextran sulfate sodium; BD,
bacteria-depleted; TLR4, Toll-like receptor 4. |
DAI was used to assess the severity of colitis in
the DSS-induced colitis mice. All mice with DSS-induced colitis
underwent fecal occult blood tests from day 3 of DSS ingestion. In
BALB/c mice, Groups B and D had significantly lower DAI scores
compared with Group C (P<0.05; Fig.
3A). In the TLR4−/− experiment, the DAI score of
groups A1 and B1 declined slowly from day 10 of the experiment.
However, the clinical score of the TLR4−/− group
exhibited a continuous increase (Fig.
3B).
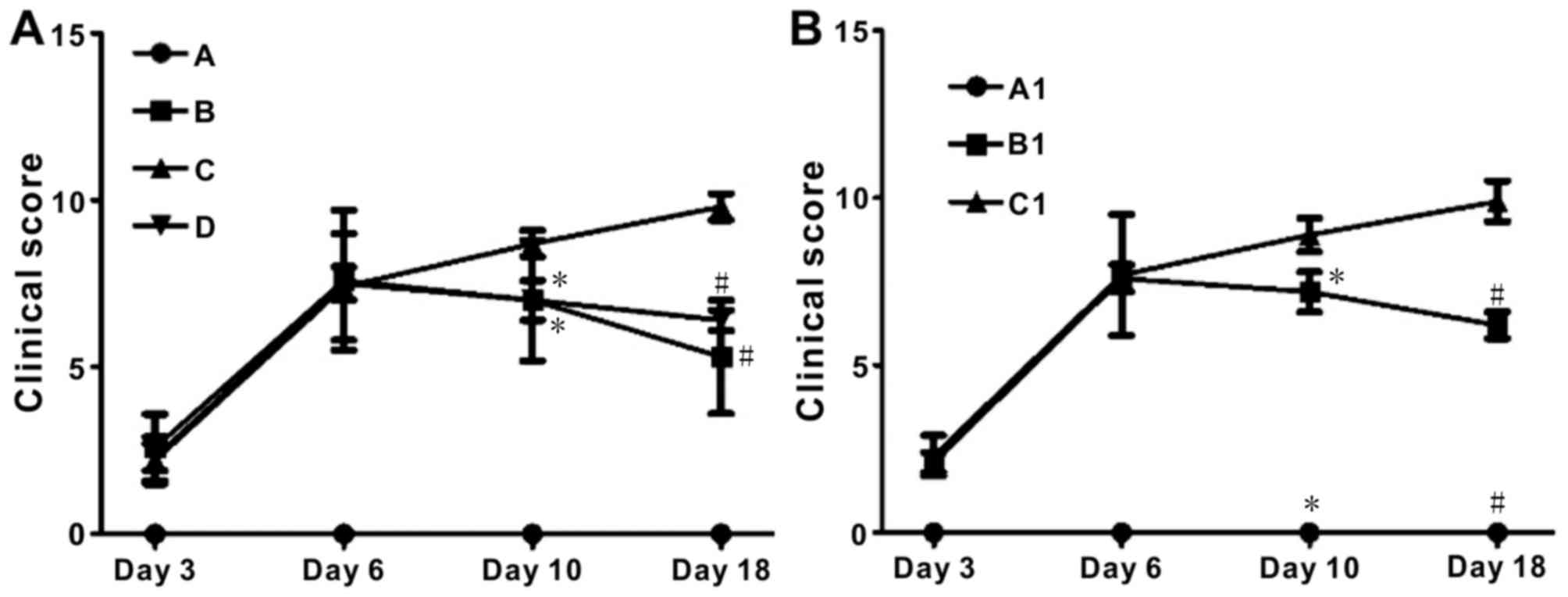 | Figure 3.Escherichia coli treatment
significantly decreases disease activity index in DSS-induced BD
mice, but not TLR4−/− mice. (A) Clinical scores in
BALB/c mice. Group D vs. Group C (day 10, *P<0.05; day 18,
#P<0.05); group B vs. group C (day 10, *P<0.05;
day 18, #P<0.05). Group A, control; Group B, DSS;
Group C, DSS + BD; Group D, DSS + BD + E. coli. (B) Clinical
scores in the TLR4−/− mice experiment. Group A1 vs.
Group C1 (day 10, *P<0.05; day 18, #P<0.05); group
B1 vs. Group C1 (day 10, *P<0.05; day 18,
#P<0.05). Group A1, control (C57); Group B1, DSS + BD
+ E. coli (C57); Group C1, DSS + BD + E. coli
(TLR4−/−). DSS, dextran sulfate sodium; BD,
bacteria-depleted; TLR4, Toll-like receptor 4. |
Histopathology
In BALB/c mice with DSS-induced colitis (Group B),
the colon was shorter than that noted in the mice of group A, and
showed extensive mucosal and glandular defects with crypt
destruction and a large amount of inflammatory cell infiltration
(Fig. 4A and B). In Group D, the
colonic mucosa defects were partially repaired, compared with group
C (Fig. 4C and D). Concomitantly,
the crypt damage decreased and inflammatory cell infiltration and
depth of inflammation were reduced. There was a significant
reduction in tissue damage in group D compared with group C
(P=0.03; Fig. 4E).
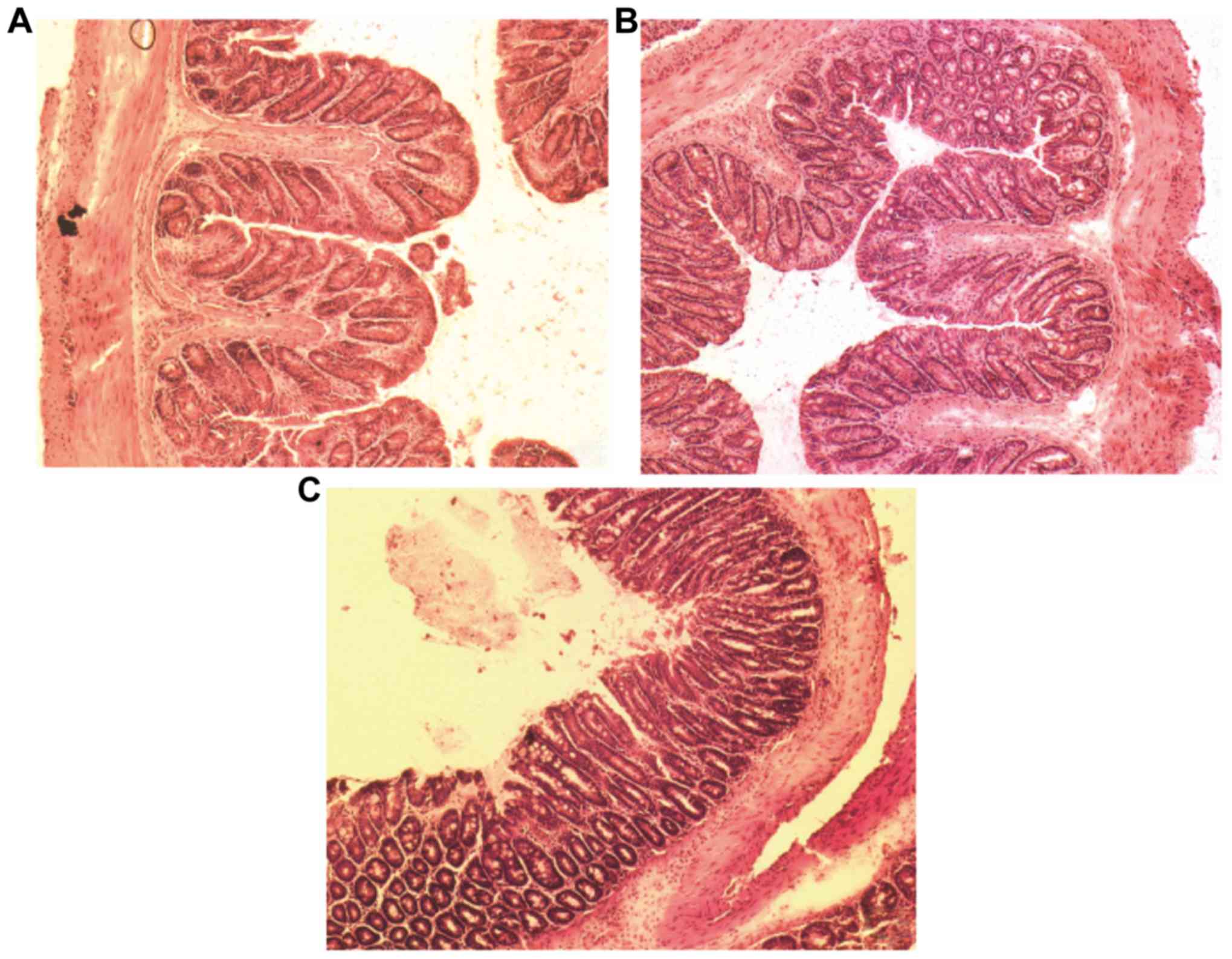
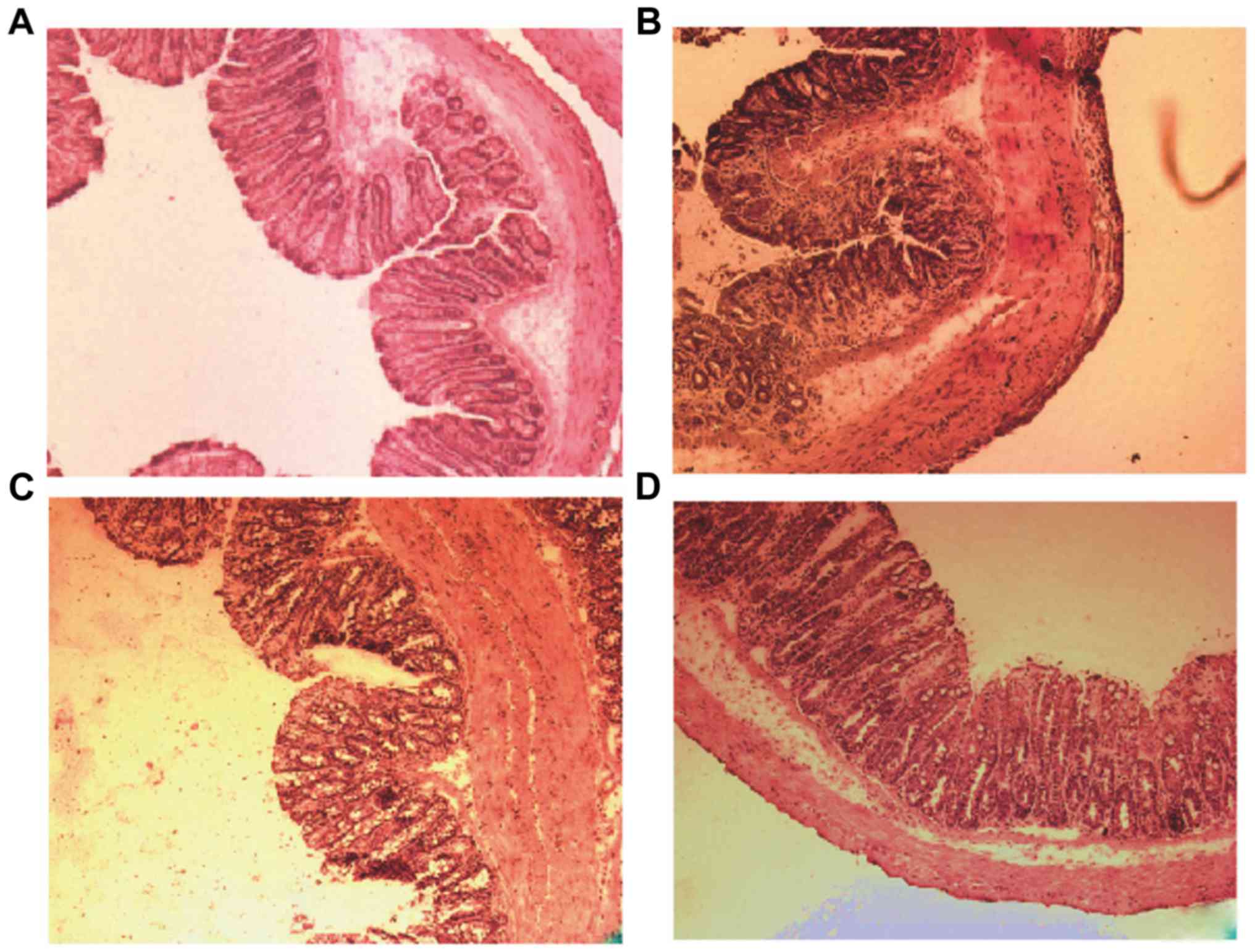 | Figure 4.Histological analysis of mice colon
sections in BALB/c mice. (A) Hematoxylin and eosin staining of the
mouse colon section in control, (B) DSS-induced, (C) DSS-induced BD
and (D) E. coli-treated DSS-induced BD mice. Magnification,
×100. DSS-induced BD mice without E. coli treatment showed
extensive mucosal and glandular defects, crypt destruction and a
large number of inflammatory cell infiltration in the recovery
phase of colonic tissue. Following E. coli treatment, the
colon mucosa defects were partially repaired, the crypt damage
decreased, inflammatory cell infiltration and depth of inflammation
was reduced. (E) Colon sections were histologically scored. *P=0.02
and #P=0.03. DSS, dextran sulfate sodium; BD,
bacteria-depleted; E. coli, Escherichia coli. |
In the TLR4−/− experiments (Fig. 5), no signs of inflammatory response
were noted in Group A1 (Fig. 5A).
Group C1 indicated extensive mucosal and glandular defects, crypt
destruction and inflammatory cell infiltration (Fig. 5C). In the E. coli-treated BD
C57BL/6 mice with DSS-induced colitis (Group B1), crypt damage
decreased, and inflammatory cell infiltration and depth of
inflammation were reduced, thus indicating that the colonic mucosal
defects were partially repaired compared with in group C1 (Fig. 5B). The differences noted between
TLR4−/− and wild type C57BL/6 group were significant
(P<0.05; Fig. 5D).
MPO activity
In wild-type BALB/c mice, the intestinal mucosal MPO
activity in Group C was significantly reduced compared with that in
Groups B and D (P<0.05). Notably, the intestinal mucosal MPO
activity in the E. coli-treated BD TLR4−/− group
was significantly reduced compared with that noted in the E.
coli-treated BD C57BL/6 group (P<0.05; Fig. 6).
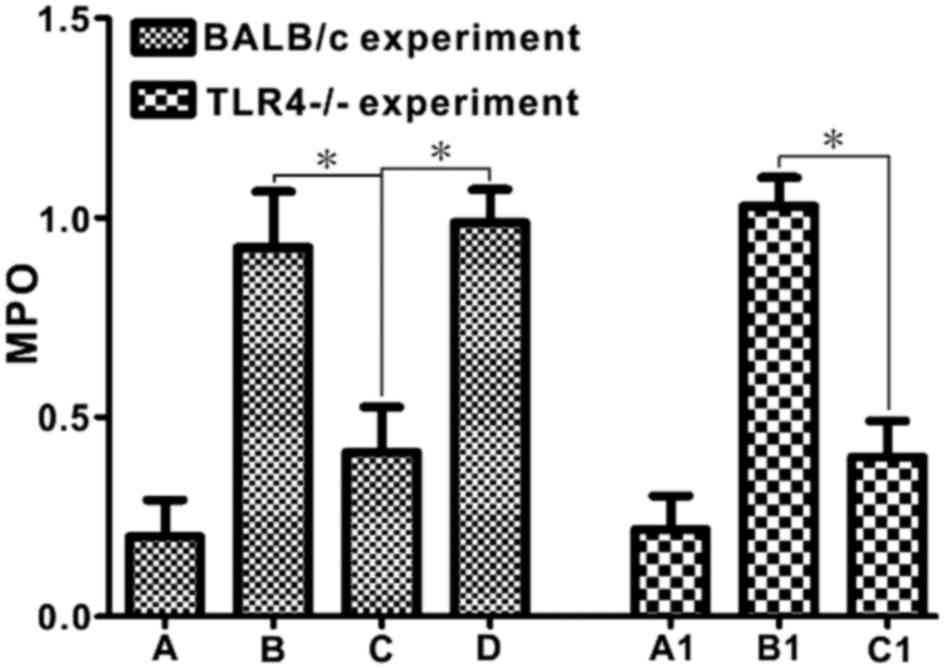 | Figure 6.Escherichia coli treatment
decreases MPO activity in DSS-induced BD mice, but not DSS-induced
BD TLR4−/− mice. MPO was measured in order to evaluate
neutrophil infiltration. *P<0.05. Group A, control; Group B,
DSS; Group C, DSS + BD; Group D, DSS + BD + E. coli; Group
A1, control (C57); Group B1, DSS + BD + E. coli (C57); Group
C1, DSS + BD + E. coli (TLR4−/−). MPO,
myeloperoxidase; DSS, dextran sulfate sodium; BD,
bacteria-depleted; TLR4, Toll-like receptor 4. |
NF-κB activation
In BALB/c mice, activated NF-κB was mainly localized
in the lamina propria, as demonstrated by brown nuclear staining
under high magnification (data not shown). Groups B and D had
higher NF-κB expression, whereas group C had a significantly lower
expression level of NF-κB, compared with these groups (P<0.05).
In addition, the results indicated that the TLR4−/−
group exhibited less NF-κB expression compared with the E.
coli-treated BD C57BL/6 group (P<0.05; Fig. 7), suggesting that TLR4 knockout
decreased NF-κB cell activation by E. coli.
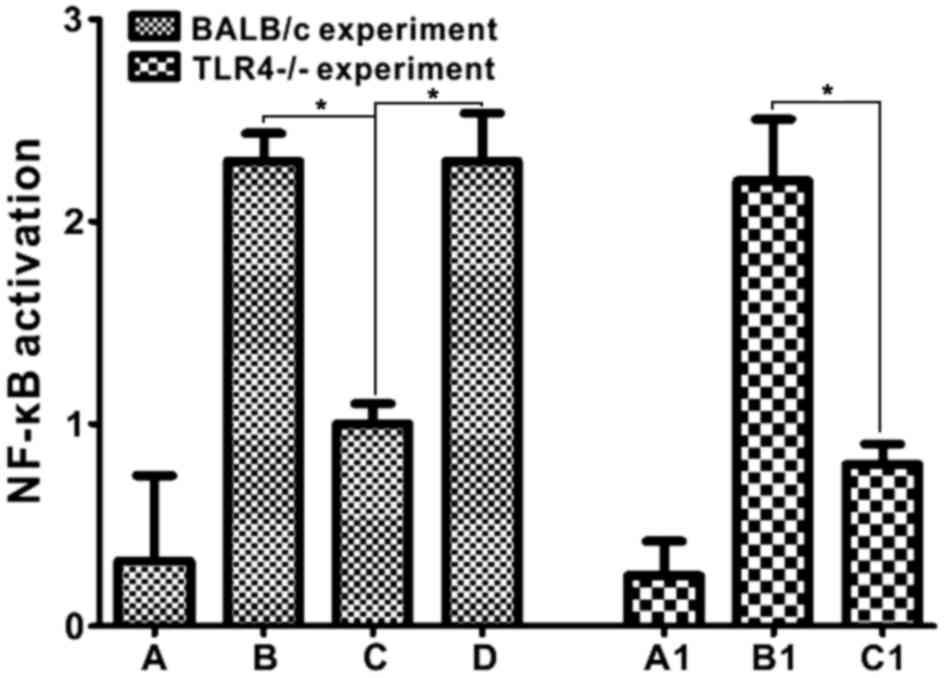 | Figure 7.Escherichia coli treatment
increases NF-κB activity in DSS-induced BD mice (group D), but not
DSS-induced BD TLR4−/− mice (group C1). Group D vs.
group C (*P<0.05); NF-κB activity in TLR4−/− mice
experiment: Group C1 vs. group B1 (*P<0.05). Group A, control;
Group B, DSS; Group C, DSS + BD; Group D, DSS + BD + E.
coli; Group A1, control (C57); Group B1, DSS + BD + E.
coli (C57); Group C1, DSS + BD + E. coli
(TLR4−/−). DSS, dextran sulfate sodium; BD,
bacteria-depleted; NF-κB, nuclear factor-κB; TLR4, Toll-like
receptor 4. |
TLR4 and NF-κB mRNA expression
In control and DSS-induced BD BALB/c mice, the mRNA
expression levels of TLR4 and NF-κB was relatively low. Following
administration of E. coli, TLR4 and NF-κB mRNA expression
levels increased significantly (P<0.05; Fig. 8A).
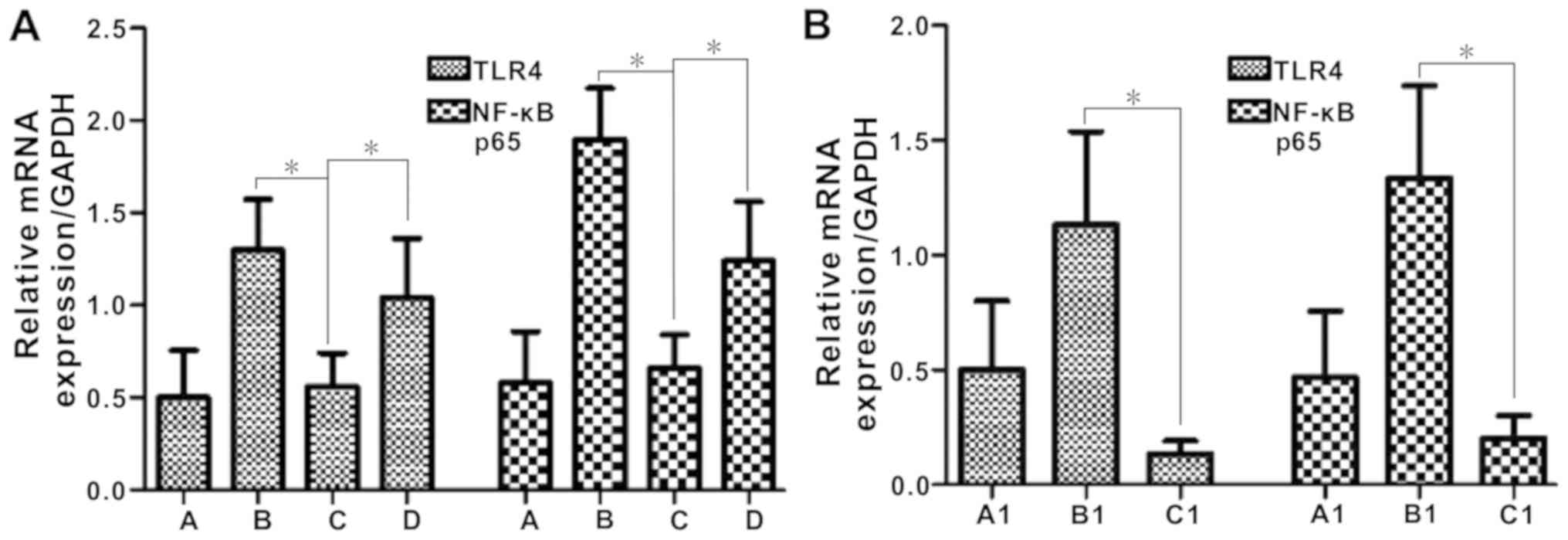 | Figure 8.TLR4 and NF-κB p65 mRNA expression is
increased by Escherichia coli in DSS-induced BD mice (group
D), but not DSS-induced BD TLR4−/− mice (group C1). (A)
TLR4 mRNA and NF-kB P65 mRNA expression in BALB/c mice and (B) the
TLR4−/− mice experiment. *P<0.05. Group A, control;
Group B, DSS; Group C, DSS + BD; Group D, DSS + BD + E.
coli; Group A1, control (C57); Group B1, DSS + BD + E.
coli (C57); Group C1, DSS + BD + E. coli
(TLR4−/−). DSS, dextran sulfate sodium; BD,
bacteria-depleted; TLR4, Toll-like receptor 4; NF-κB, nuclear
factor-κB. |
In the experiment using TLR4−/− mice,
Group C1 exhibited significantly decreased mRNA levels of TLR4 and
NF-κB compared with group B1 (P<0.05; Fig. 8B).
Discussion
IBD is thought to be caused by an imbalance of the
immune response and the intestinal flora, although the mechanism by
which the intestinal flora affects the development of this disease
remains unclear (21,22). The intestinal microbiome has been
shown to have an important role in the development of IBD (13,23).
The mutual communication between intestinal microbes and the
intestinal mucosa regulates the expression of intestinal genes
(23).
Several studies in various animal colitis models
have demonstrated the importance of the resident luminal flora in
the initiation and perpetuation of intestinal inflammation
(24). Non-pathogenic
microorganisms in the intestinal mucosa may alter the immune
response in the intestinal epithelial barrier function in
vitro (24). Feces within the
intestinal lumen can contain up to 1×1011 bacteria per
gram, with E. coli as the predominant species (25). E. coli is the most populous
bacteria in the gut of humans and various animals (25). To the best of our knowledge, no
relevant reports exist in regards to the involvement of the normal
intestinal E. coli in experimental colitis recovery.
In general, mouse colitis caused by DSS induction
heals itself (26). In contrast to
this outcome, BD mice that were orally administered antibiotics for
2 weeks in the present study were unable to recover from damage
caused by DSS induction, indicating that host intestinal microbial
interactions were necessary for recovery. The results demonstrated
that commensal E. coli reduced mortality in BD mice with
DSS-induced colitis, and promoted colitis recovery. This suggested
that the presence of normal intestinal E. coli is necessary
for recovery.
Inflammation and necrosis of intestinal mucosa leads
to colon shortening and these alterations reflect the degree of
damage to the intestinal tissue. In the current study, the colon
length of DSS-induced mice was shorter than that of normal mice.
E. coli treatment could prevent colon shortening, which may
be associated with the ability of E. coli to reduce
intestinal inflammation and scarring (27). With regard to the histopathological
findings, the E. coli treatment groups had restored
intestinal mucosal integrity, reduced intestinal hemorrhage and
inflammatory cell infiltration, and had a lower histological score
than DSS-induced BD mice without E. coli treatment. However,
in TLR4−/− mice, E. coli treatment could not
reduce the colon tissue damage. These results indicated that E.
coli promoted recovery of DSS-induced colonic inflammation and
that this effect was potentially mediated, at least partially,
through the TLR4 receptor.
NF-κB is a transcription factor that regulates the
expression of a series of inflammatory factors (28). In the present study, E.
coli-treated DSS-induced BD mice expressed more NF-κB compared
with untreated DSS-induced BD mice, which suggested that E.
coli promoted NF-κB activation and accelerated inflammation
recovery. By contrast, TLR4−/− mice showed no
significant recovery from colitis, suggesting that E. coli
promoted DSS-induced colitis recovery through activation of the
TLR4/NF-κB signaling pathway.
In conclusion, the presence of normal intestinal
flora may be a necessary condition for the recovery of colitis,
potentially through activation of the TLR4/NF-κB signaling pathway.
This indicated that host intestinal microbial interactions were
critical to colitis recovery. This study provided evidence for the
use of intestinal flora to treat IBD. In the future, more studies
are required to identify the mechanisms and confirm its
efficacy.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from Guangzhou
Science Technology and Innovation Commission (grant no.
2014Y2-00074), the Natural Science Foundation of Guangdong (grant
no. 2015A030313729) and the Guangzhou Science Technology and
Innovation Commission (grant no. 201804010073).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL and JC were responsible for experimental design.
JL and BD performed the experiments. AC, FH, BP and ZW were
responsible for statistical analysis. JL and BD wrote the
manuscript. All authors were involved in manuscript revision.
Ethics approval and consent to
participate
The protocol was approved by The Review Board of the
Institute of Medical Animal Laboratory at the Guangzhou Medical
University (Guangzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Baumgart DC and Sandborn WJ: Inflammatory
bowel disease: Clinical aspects and established and evolving
therapies. Lancet. 369:1641–1657. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Egger B, Bajaj-Elliott M, MacDonald TT,
Inglin R, Eysselein VE and Büchler MW: Characterisation of acute
murine dextran sodium sulphate colitis: Cytokine profile and dose
dependency. Digestion. 62:240–248. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Prideaux L, Kamm MA, De Cruz PP, Chan FK
and Ng SC: Inflammatory bowel disease in Asia: A systematic review.
J Gastroenterol Hepatol. 27:1266–1280. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sellon RK, Tonkonogy S, Schultz M,
Dieleman LA, Grenther W, Balish E, Rennick DM and Sartor RB:
Resident enteric bacteria are necessary for development of
spontaneous colitis and immune system activation in
interleukin-10-deficient mice. Infect Immun. 66:5224–5231.
1998.PubMed/NCBI
|
|
5
|
Blum AM, Metwali A, Elliott DE, Berg DJ
and Weinstock JV: CD4+ T cells from IL-10-deficient mice transfer
susceptibility to NSAID-induced Rag colitis. Am J Physiol
Gastrointest Liver Physiol. 287:G320–G325. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rutgeerts P, Goboes K, Peeters M, Hiele M,
Penninckx F, Aerts R, Kerremans R and Vantrappen G: Effect of
faecal stream diversion on recurrence of Crohn's disease in the
neoterminal ileum. Lancet. 338:771–774. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Khan KJ, Ullman TA, Ford AC, Abreu MT,
Abadir A, Marshall JK, Talley NJ and Moayyedi P: Antibiotic therapy
in inflammatory bowel disease: A systematic review and
meta-analysis. Am J Gastroenterol. 106:661–673. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ananthakrishnan AN, Hur C, Juillerat P and
Korzenik JR: Strategies for the prevention of postoperative
recurrence in Crohn's disease: Results of a decision analysis. Am J
Gastroenterol. 106:2009–2017. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jostins L, Ripke S, Weersma RK, Duerr RH,
McGovern DP, Hui KY, Lee JC, Schumm LP, Sharma Y, Anderson CA, et
al: Host-microbe interactions have shaped the genetic architecture
of inflammatory bowel disease. Nature. 491:119–124. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen J, Xie L, Toyama S, Hünig T, Takahara
S, Li XK and Zhong L: The effects of Foxp3-expressing regulatory T
cells expanded with CD28 superagonist antibody in DSS-induced mice
colitis. Int Immunopharmacol. 11:610–617. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals: Guide for the Care and Use of Laboratory Animals. 8th.
National Academies Press (US); Washington, DC: 2011
|
|
12
|
Cooper HS, Murthy SN, Shah RS and
Sedergran DJ: Clinicopathologic study of dextran sulfate sodium
experimental murine colitis. Lab Invest. 69:238–249.
1993.PubMed/NCBI
|
|
13
|
Loher F, Schmall K, Freytag P, Landauer N,
Hallwachs R, Bauer C, Siegmund B, Rieder F, Lehr HA, Dauer M, et
al: The specific type-4 phosphodiesterase inhibitor mesopram
alleviates experimental colitis in mice. J Pharmacol Exp Ther.
305:549–556. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moriyama I, Ishihara S, Rumi MA, Aziz MD,
Mishima Y, Oshima N, Kadota C, Kadowaki Y, Amano Y and Kinoshita Y:
Decoy oligodeoxynucleotide targeting activator protein-1 (AP-1)
attenuates intestinal inflammation in murine experimental colitis.
Lab Invest. 88:652–663. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Krawisz JE, Sharon P and Stenson WF:
Quantitative assay for acute intestinal inflammation based on
myeloperoxidase activity. Assessment of inflammation in rat and
hamster models. Gastroenterology. 87:1344–1350. 1984.PubMed/NCBI
|
|
16
|
Becker C, Neurath MF and Wirtz S: The
intestinal microbiota in inflammatory bowel disease. ILAR J.
56:192–204. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bantel H, Berg C, Vieth M, Stolte M, Kruis
W and Schulze-Osthoff K: Mesalazine inhibits activation of
transcription factor NF-kappaB in inflamed mucosa of patients with
ulcerative colitis. Am J Gastroenterol. 95:3452–3457. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang T, Su J, Guo B, Wang K, Li X and
Liang G: Apigenin protects blood-brain barrier and ameliorates
early brain injury by inhibiting TLR4-mediated inflammatory pathway
in subarachnoid hemorrhage rats. Int Immunopharmacol. 28:79–87.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang CX, Xie GB, Zhou CH, Zhang XS, Li T,
Xu JG, Li N, Ding K, Hang CH, Shi JX and Zhou ML: Baincalein
alleviates early brain injury after experimental subarachnoid
hemorrhage in rats: Possible involvement of TLR4/NF-κB-mediated
inflammatory pathway. Brain Res 1594. 245–255. 2015. View Article : Google Scholar
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ray A and Dittel BN: Interrelatedness
between dysbiosis in the gut microbiota due to immunodeficiency and
disease penetrance of colitis. Immunology. 146:359–368. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Loh G and Blaut M: Role of commensal gut
bacteria in inflammatory bowel diseases. Gut Microbes. 3:544–555.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vigsnaes LK, van den Abbeele P, Sulek K,
Frandsen HL, Steenholdt C, Brynskov J, Vermeiren J, van de Wiele T
and Licht TR: Microbiotas from UC patients display altered
metabolism and reduced ability of LAB to colonize mucus. Sci Rep.
3:11102013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Schultz M, Veltkamp C, Dieleman LA,
Grenther WB, Wyrick PB, Tonkonogy SL and Sartor RB: Lactobacillus
plantarum 299V in the treatment and prevention of spontaneous
colitis in interleukin-10-deficient mice. Inflamm Bowel Dis.
8:71–80. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Amar J, Chabo C, Waget A, Klopp P, Vachoux
C, Bermúdez-Humarán LG, Smirnova N, Bergé M, Sulpice T, Lahtinen S,
et al: Intestinal mucosal adherence and translocation of commensal
bacteria at the early onset of type 2 diabetes: Molecular
mechanisms and probiotic treatment. EMBO Mol Med. 3:559–572. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhang R, Ito S, Nishio N, Cheng Z, Suzuki
H and Isobe KI: Dextran sulphate sodium increases splenic
Gr1(+)CD11b(+) cells which accelerate recovery from colitis
following intravenous transplantation. Clin Exp Immunol.
164:417–427. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fabrega MJ, Rodriguez-Nogales A,
Garrido-Mesa J, Algieri F, Badía J, Giménez R, Gálvez J and Baldomà
L: Intestinal Anti-inflammatory effects of outer membrane vesicles
from Escherichia coli nissle 1917 in DSS-experimental colitis in
mice. Front Microbiol. 8:12742017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
French N and Pettersson S: Microbe-host
interactions in the alimentary tract: The gateway to understanding
inflammatory bowel disease. Gut. 47:162–163. 2000. View Article : Google Scholar : PubMed/NCBI
|