Introduction
Inflammatory bowel diseases (IBDs) such as
ulcerative colitis and Crohn's disease are refractory disorders
characterized by chronic relapsing inflammation of the
gastrointestinal tract. The mucosal lesions in IBD are generated by
an excessive or dysregulated immune response against commensal
microbes in the gut.
Genome-wide association studies have reported that
IBD-associated single nucleotide polymorphisms (SNPs) in genes such
as NOD2, CARD9, and ATG16L1 interact with the
microbiota and are involved in the pathogenesis of IBD (1). Several studies have revealed that
microbial imbalance causes pro-inflammatory immunological responses
and induces gut inflammation (2,3).
Further, it has been reported that specific commensal bacteria
affect the differentiation of mucosal immunocompetent cells. It is
possible that some commensal bacterial strains can also induce
anti-inflammatory immunological responses. To maintain intestinal
homeostasis it is important that the commensal bacterial flora
remains consistent (4–7) Microbial dysbiosis is essential for
driving inflammation in IBD (8),
and can be induced by several factors that affect the complexity
and stability of the microbiome, including genetics, birth route,
stress, nutrition, and drugs (9).
The gut commensal bacteria regulate mucosal inflammation by several
mechanisms, including inducing regulatory T cells, down-regulating
inflammatory cytokines, and regulating nuclear factor (NF)-κB
activation (4,10,11).
Lactobacillus bacteria, which constitute a significant
component of the gut microbiota, downregulate inflammatory
signaling in the mucosa of IBD patients and also ameliorate dextran
sulfate sodium (DSS)-induced colitis in mice by reducing the
production of pro-inflammatory cytokines (12).
The Lactobacillus plantarum (LP) 06CC2
strain, isolated from traditional Mongolian dairy products, has
shown probiotic potential in mice and has immune modulating
effects. Oral administration of the 06CC2 strain exerted a
significant immunomodulatory effect in influenza virus-infected
mice (13,14). However, the involvement of the
06CC2 strain in intestinal inflammation remains unknown. Therefore,
we assessed the impact of the 06CC2 strain in experimental colitis
and its impact on intestine-specific, antigen-presenting cell
phenotypes such as macrophages.
Materials and methods
Preparation of the LP 06CC2
strain
The 06CC2 strain, a lactic acid bacterium with
probiotic potential, was isolated from Mongolian dairy cheese
(Aaruul) as described previously (13). The 06CC2 cells were cultured at
37°C for 24 h in Man, Rogosa, and Sharpe (MRS) broth (Merck KGaA),
cells were harvested by centrifugation at 10,000 × g for 5 min,
washed twice with PBS, and then boiled for 1 h (14). The boiled 06CC2 cells were washed
again with PBS and lyophilized. The lyophilized 06CC2 cells were
suspended in Roswell Park Memorial Institute (RPMI)-1640 medium or
Dulbecco's modified Eagle's medium (DMEM) with 10% heat-inactivated
fetal bovine serum (FBS) for co-culture. For oral administration to
mice, heat-killed 06CC2 cells were lyophilized and powdered, and
the 06CC2 powder was suspended in distilled water before use.
Animals
Specific-pathogen-free C57BL/6 mice, 8 to 10 weeks
of age and weighing between 22 to 26 g, were sourced from Kyudo
(Saga). Mice were maintained under standard conditions (22°C,
50–60% humidity, and 12-h light/dark cycle) with free access to
standard mouse/rat chow diet (LabDiet Autoclavable Rodent Diet
5010; PMI Nutrition International,). We monitored the health of
mice daily, and all efforts were made to minimize animal suffering.
We monitored body and stool conditions and measured the body weight
of the mice each day. Mice were euthanized by cervical dislocation
following anesthesia intraperitoneally with pentobarbital (30–50
mg/kg), and the colonic tissue was collected following euthanasia.
Animal experimental procedures were approved by the institutional
animal care and use committees of Kagoshima University (permit no.
MD14106), and were performed in accordance with these committees'
guidelines for animal experiments.
Induction and assessment of
colitis
DSS-induced colitis is an established experimental
model that allows the investigation of the signs and symptoms of
UC, including diarrhea, weight loss, bloody stools, mucosal
ulceration, and shortening of the large intestine (15). The Acute experimental colitis was
induced using 1.0% DSS. DSS powder was manufactured by Wako Pure
Chemical Industries, Ltd., and the average molecular weight was
5,000 Da. Control mice were given distilled water. A 06CC2
suspension (20 mg per mouse) or PBS was inoculated twice a day into
experimental mice 3 days before oral DSS administration.
06CC2-treated mice were administered 1.0% DSS for 15 days and were
euthanized on day 20. Whole colon tissue was collected and lamina
propria mononuclear cells (LPMCs) were extracted.
In another experiment, a 06CC2 suspension or PBS was
inoculated into mice for 30 days. Beginning 3 days after
inoculation, 1.0% DSS was administered for 27 days, and
neutralizing anti-IL-10 monoclonal antibodies (clone JES5-16E3;
BioLegend) or control rat IgG2a (BioLegend) (0.4 mg/200 ml) was
injected intraperitoneally for the last 8 days. The DAI was
assessed blindly in both mouse groups. The DAI is based on clinical
scores for weight loss, stool consistency, and bleeding, as
previously described (16). Each
clinical parameter was scored on a scale from 0 to 4, and the
parameter values were summed.
Isolation of murine colonic LPMCs
Colonic LPMCs were isolated according to a previous
report (17). Briefly, colon
samples were cut into small pieces in
Ca2+/Mg2+-free 1.5% Hank's balanced salt
solution (1.5% HBSS). The colon samples were then placed in 1.5%
HBSS and washed three times at 37°C, then incubated with HBSS
containing 1 mM dithiothreitol (#15-508-031; Invitrogen; Thermo
Fisher Scientific, Inc.) and 5 mM EDTA (#15-575-020; Invitrogen;
Thermo Fisher Scientific, Inc.) for 30 min at 37°C to remove the
epithelial layer. The samples were then placed in digestion
solution containing 1.5% HBSS, 1.0–3.0 mg/ml collagenase A
(#11-088-793-001; Roche Diagnostics GmbH) and 0.1 mg/ml DNase I
(#11-284-932-001; Roche Diagnostics GmbH) for 1–2 h at 37°C. The
samples were passed through a 100-µm strainer and transferred to a
50-ml Falcon tube and centrifuged for 5 min at 4°C. The supernatant
was washed, and the pellets were resuspended in 40% Percoll and
overlaid on a 75% Percoll fraction. Mononuclear cells were
collected at the interphase, washed, and resuspended in RPMI-1640
medium (#11875-093; Life Science Products, Inc.) containing 10% FBS
and 1% penicillin/streptomycin.
Histological assessment and
evaluation
Colon tissue was fixed in 10% phosphate-buffered
formalin, and was embedded in paraffin, sectioned, and stained with
hematoxylin and eosin. Colitis stages were scored via blinded
histopathological analysis according to previously described
morphological criteria: Epithelial properties, depth of
inflammation, and degree of ulceration (18). Epithelial properties were scored as
follows: 0, intact crypt; 1, loss of the basal one-third of the
crypt; 2, loss of the basal two-thirds of the crypt; 3, loss of the
entire crypt but with intact surface epithelium; and 4, loss of the
entire crypt and surface epithelium (erosion). Depth of
inflammation was scored as follows: 0, no infiltration; 1, crypt
base; 2, mucosa; 3, submucosa; and 4, submucosa (extensive). The
degree of ulceration was scored as follows: 0, none; 2, positive;
and 4, positive (extensive). Each specimen was assigned a
histological score ranging from 0 to 12.
Cell separation of LPMCs by
magnetic-activated cell sorting (MACS)
Colonic LPMCs were separated by MACS. LPMCs with
specific CD antibodies were magnetically labeled with their
respective magnetic beads (CD11c, #130-108-338; CD11b,
#130-049-601; CD4, #130-049-201; Miltenyi Biotec). The cell
suspension was loaded onto a MACS® LS Column
(#130-042-401; Miltenyi Biotec), which was placed in the magnetic
field of a MACS Separator (#130-042-301; Miltenyi Biotec). The MACS
Separation Buffer contained a final BSA (Miltenyi Biotec)
concentration of 0.5%. After cells were separated, they were
analyzed for gene expression.
RNA isolation and gene expression
assay
Total RNA was extracted from colon tissues or LPMCs
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
Total RNA (0.5 µg) was reverse transcribed using a PrimeScript RT
Reagent kit (Takara Bio Inc.) at 37°C for 15 min and 85°C for 5
sec. Each reaction mixture for RT (total volume 20 µl) consisted of
total RNA (adjusted to 500 ng, 2.5 µl), oligo dT PCR primer (50 µM,
1 µl), 5X PrimeScript buffer (4 µl), PrimeScript RT enzyme mix (1
µl), random 6 mers (100 µM, 1 µl), and RNase free distilled
H2O (10.5 µl). The synthesized cDNA was amplified by
RT-qPCR using SYBR (Applied Biosystems; Thermo Fisher Scientific,
Inc.). Each reaction mixture for PCR (total volume 20 µl) consisted
of cDNA (adjusted to 500 ng, 2 µl), forward and reverse PCR primers
(10 µM, 0.8 µl each), SYBR (10X, 10 µl), ROX reference dye (50X,
0.4 µl), and distilled H2O (6 µl). The cycling conditions were as
follows: One cycle at 95°C for 30 sec followed by 35 cycles each at
95°C for 5 sec and 60°C for 34 sec. The relative expression levels
of target genes were normalized to rRNA (Applied Biosystems; Thermo
Fisher Scientific, Inc.) and were calculated using the
2−∆∆Cq method (19).
Primers for the following genes were used: GAPDH, IL-6, IL-10,
IL-12α, IL-23α, TGF-β, TNF-α, and Toll-like receptor (TLR)2, TLR3,
and TLR4 (Table I).
 | Table I.List of primers. |
Table I.
List of primers.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| IL-6 |
CCACTTCACAAGTCGGAGGCTTA |
CCAGTTTGGTAGCATCCATCATTTC |
| IL-10 |
GACCAGCTGGACAACATACTGCTAA |
GATAAGGCTTGGCAACCCAAGTAA |
| IL-12α | AATGTTCCA
GTGCCTCAACC | CTAGAGTTTGT
CTGGCCTTCTG |
| TNF-α |
TATGGCCCAGACCCTCACA |
GGAGTAGACAAGGTACAACCCATC |
| TLR2 |
CTCCCAGCAGGAACATCTGCTA |
CCAGGAATGAAGTCCCGCTTA |
| TLR3 |
TGTCTGGAAGAAAGGGACTTTGA |
GTTGAACTGCATGATGTACCTTGA |
| TLR4 |
CTGGGTGTGTTTCCATGTCTCA |
TGCGGACACACACACTTTCAAATA |
| Foxp3 |
AGTGCCTGTGTCCTCAATGGTC |
AGGGCCAGCATAGGTGCAAG |
| TGF-β |
GCTGAGCCAGCCAGATATAACAAGA |
ACAGGCAAGTAGCTGATCCCAAAC |
| GAPDH |
TGTGTCCGTCGTGGATCTGA |
TTGCTGTTGAAGTCGCAGGAG |
ELISA for IL-10
LPMCs were co-cultured with 06CC2 at concentrations
of 0, 1, 10 and 100 µg/ml, and the supernatant was collected. IL-10
concentrations were measured with an ELISA kit (#M1000B; R&D
Systems) according to the manufacturer's instructions, and analyzed
in duplicate using a microplate reader (Bio-Rad Laboratories, Inc.)
at 450 nm. The concentration of IL-10 in the supernatant was
calculated according to a standard curve.
DNA extraction from fecal samples and
intestinal bacteria assay
For quantification by quantitative PCR, DNA was
extracted from fecal samples obtained from 06CC2-treated and
control mice on day 20 using an ISOFECAL for Beads Beating kit
(Nippon Gene) according to the manufacturer's instructions as
described in a previous report (20). DNA was also extracted from standard
strains for each group, including Bifidobacterium longum JCM
1217 for the Bifidobacterium subspecies, the 06TCa19 strain
for the Lactobacillus group, Clostridium coccoides
JCM 1395 for the Clostridium group, and Bacteroides
fragilis JCM 11019 for the Bacteroides-Prevotella group.
Except for strain 06TCa19, these strains were cultured at 37°C for
24–48 h in GAM broth and on GAM agar (Nissui) containing 0.5%
glucose in jars with anaerobic packs for enumeration of
colony-forming units (CFUs). Strain 06TCa19 was cultured at 37°C
for 24 h in MRS broth and on MRS agar. Bifidobacterium
subspecies, C. coccoides, Bacteroides-Prevotella, and
Lactobacillus groups were detected by SYBR-Green
quantitative PCR. Specific primers are shown in Table II (20). Bacterial CFUs were calculated using
a standard regression curve of the threshold cycle values generated
from 10-fold serial dilutions of DNA samples from the standard
strains with known numbers of CFUs. Each fecal sample was diluted
103-fold and stained with 4′,6-diamidino-2-phenylindole
(DAPI) solution (Dojindo Laboratories), and the bacteria were
counted with a hemocytometer for bacteria (ASONE) using a
fluorescence microscope (Olympus).
| Table II.Primer list for target bacteria. |
Table II.
Primer list for target bacteria.
| Target
bacteria | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
|
Bifidobacterium subspecies |
TCGCGTCYGGTGTGAAAG |
CCACATCCAGCRTCCAC |
|
Lactobacillus group |
AGCAGTAGGGAATCTTCCA |
CACCGCTACACATGGAG |
|
Bcteroides-Prevotella group |
GAAGGTCCCCCACATTG |
CAATCGGAGTTCTTCGTG |
| Clostridium
coccoides group |
AAATGACGGTACCTGACTAA |
TTTGAGTTTCATTCTTGCGAA |
Statistical analysis
All of the data are expressed as the mean and
standard error of the mean. Differences between two or three groups
were appropriately analyzed using one-way ANOVA followed by the
Tukey HSD, Dunnet T3, Mann-Whitney U test, and Steel-Dwass tests
(IBM SPSS version 23.0). P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of genes for
inflammation-related cytokines and concentrations of LPMCs
co-cultured with LP 06CC2
The effect of 06CC2 on LPMCs isolated from whole
colon samples of mice without DSS-induced colitis was investigated.
The expression of genes for the following pro- and
anti-inflammatory cytokines was assessed: IL-12α, TNF-α, IL-4,
IL-23α, TGF-β, and IL-10. Gene expressions of IL-12α, TNF-α, TGF-β,
and IL-10 were significantly and dose-dependently higher in LPMCs
exposed to 06CC2. Since the gene expression of IL-10 was induced at
particularly high levels, we focused on results related to IL-10
(Fig. 1). LPMCs were separated by
MACS as follows: CD4-positive cells, CD11b-positive cells, and
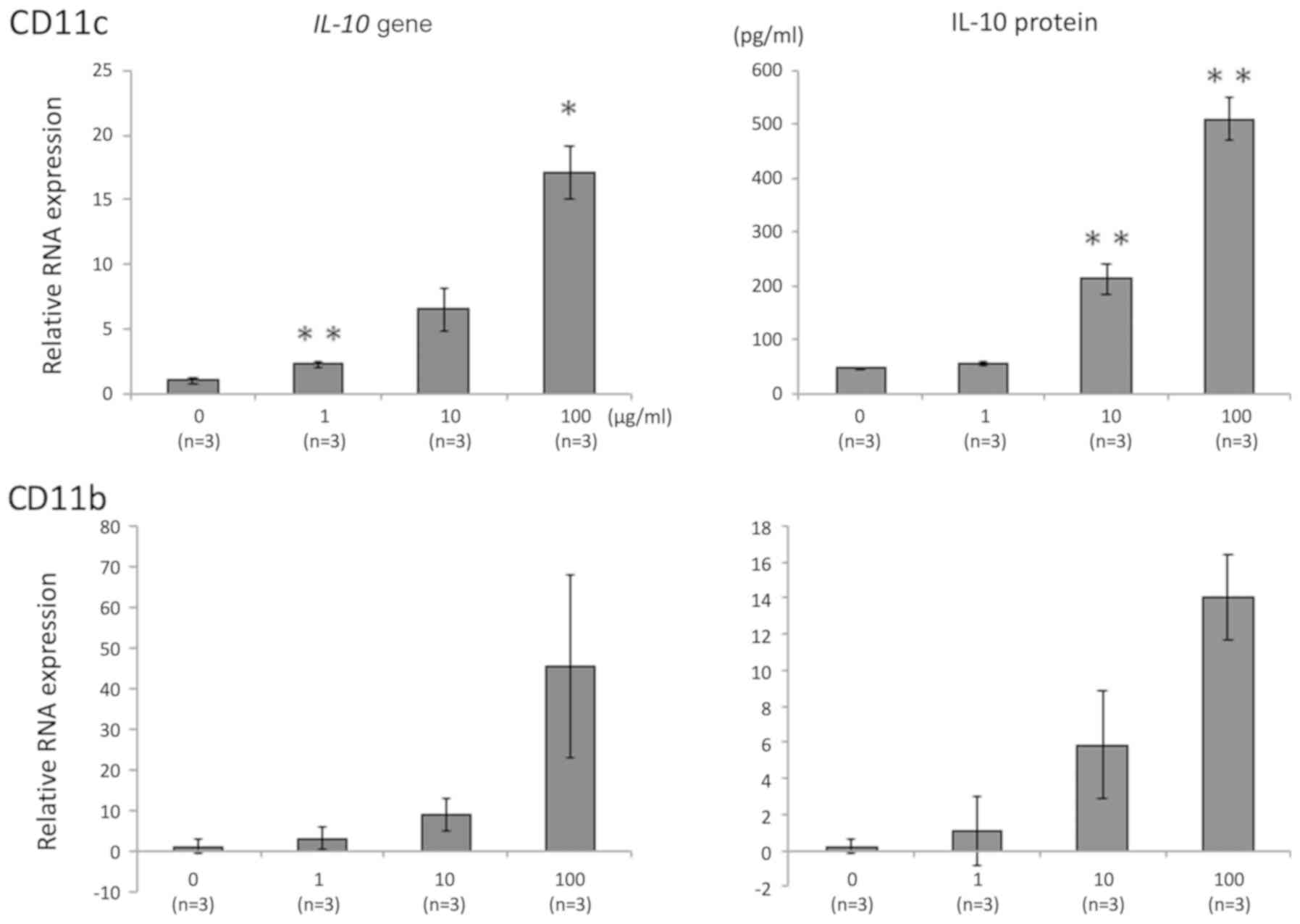
CD11c-positive cells. The gene expression and protein concentration
of IL-10 were increased by 06CC2 stimulation in a dose-dependent
manner in CD11b-positive and CD11c-positive cells (Fig. 2), but not in CD4-positive cells
(data not shown). The production of IL-10 was especially high in
CD11c-positive cells. These results show that 06CC2 induced
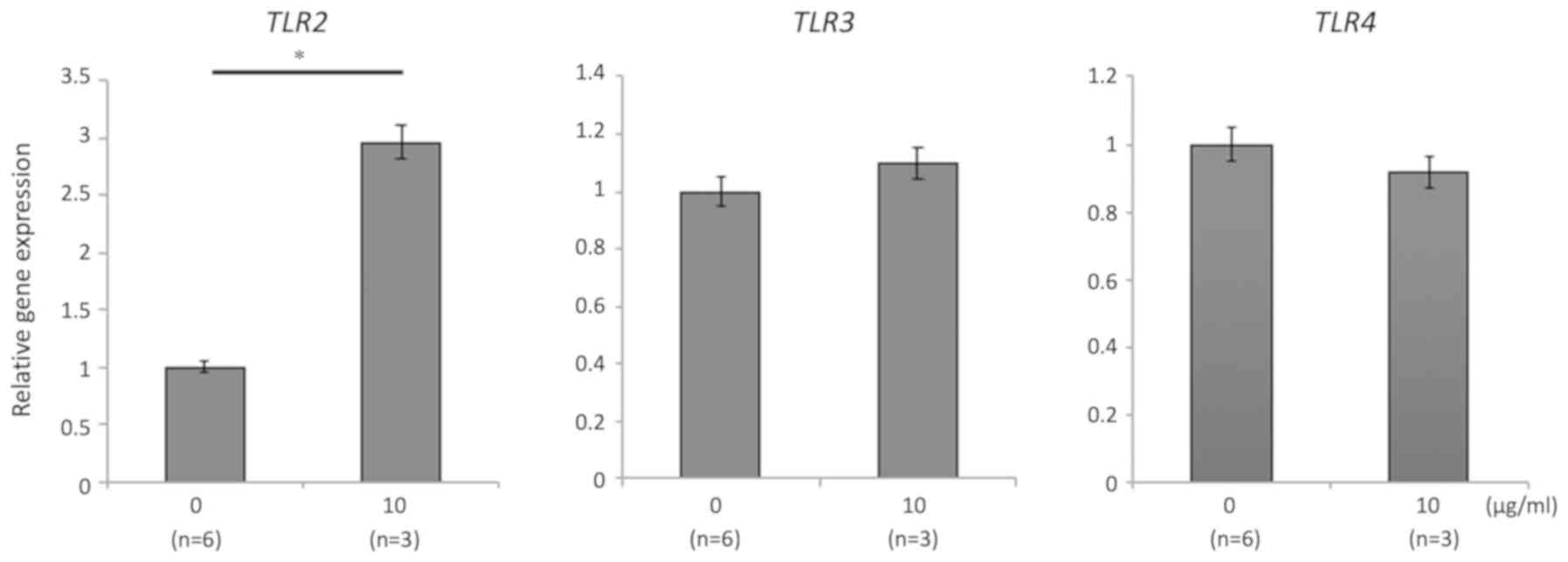
expression of a regulatory cytokine in LPMCs. TLR gene expression
was significantly increased by 06CC2 administration only in the
case of TLR2 in CD11c-positive cells; gene expressions of TLR3 and
TLR4 were unchanged (Fig. 3).
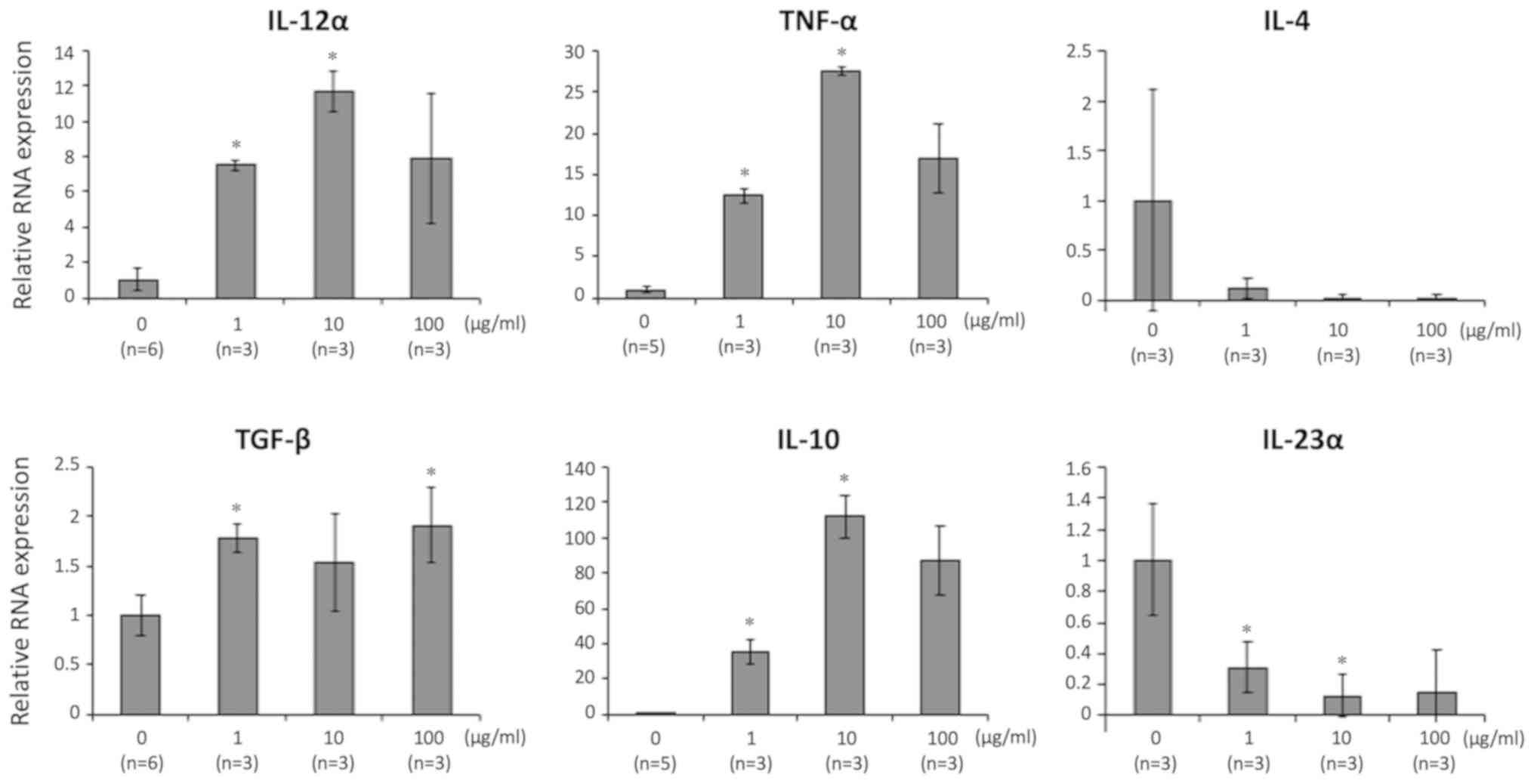 | Figure 1.LPMCs isolated from entire mouse
colons were co-cultured with 06CC2 (0, 1, 10 and 100 µg/ml). The
gene expression levels of IL-12α, TNF-α, TGF-β and IL-10 were
significantly and dose-dependently higher in LPMCs exposed to
06CC2; IL-10 gene expression was particularly high compared with
the other genes. IL-12α, IL-4 and IL-23α were analyzed using
Tukey's test. TNF-α, TGF-β and IL-10 were analyzed using Dunnet T3
test. *P<0.05 vs. 0 µg/ml. IL, interleukin; LPMCs, lamina
propria mononuclear cells; TGF-β, transforming growth factor β;
TNF-α, tumor necrosis factor α. |
Effects of oral administration of LP
06CC2 on the immune system of the murine large intestine
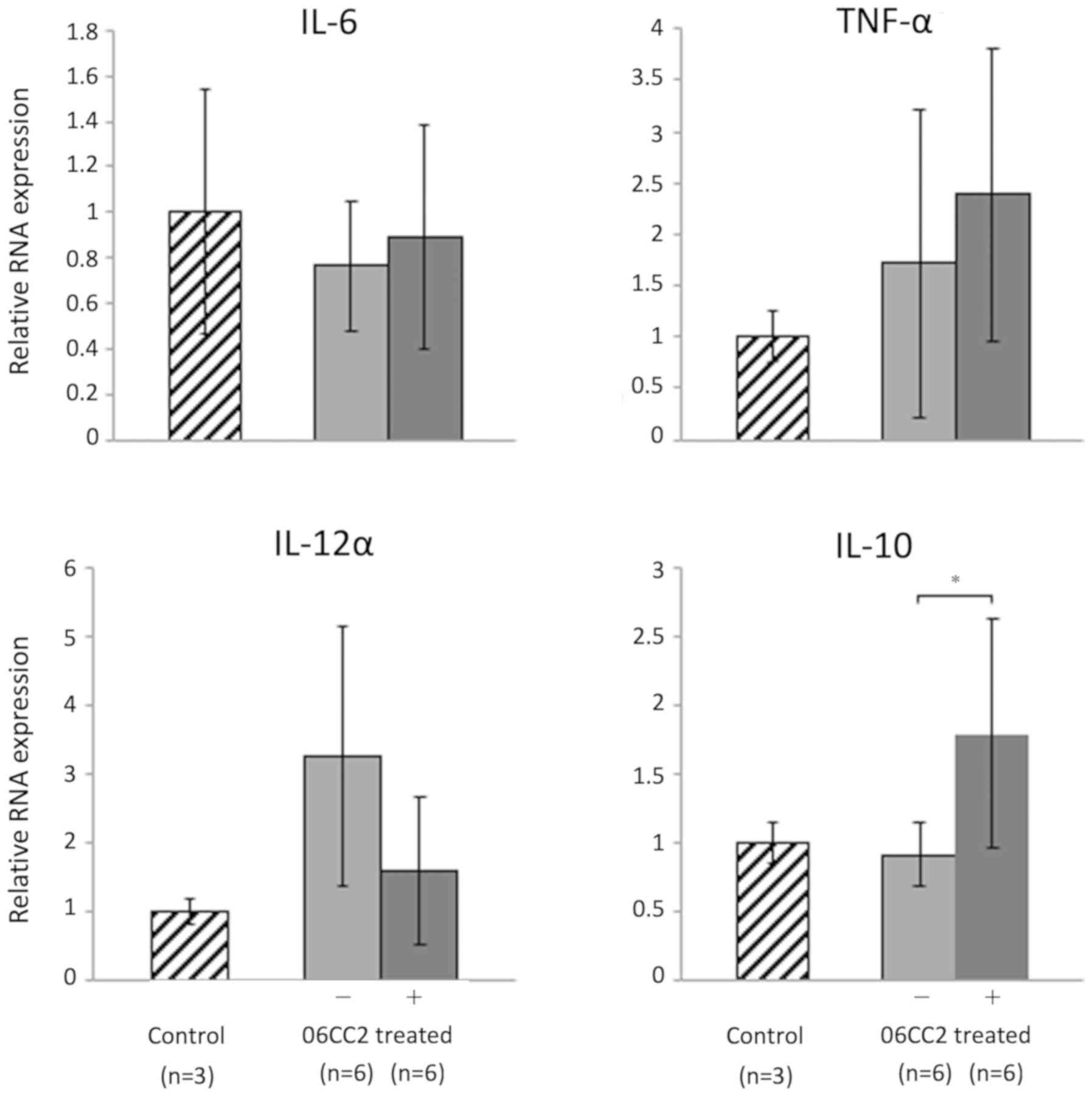
06CC2 was orally administered to mice twice a day
for 4 days; the mice were subsequently euthanized, and colon tissue
was collected for analysis. Pro-inflammatory and regulatory
cytokines in the large intestine were investigated by quantitative
PCR. The expression of IL-10 was significantly increased in mice
treated with 06CC2 compared to control mice (P=0.048). However,
administration of 06CC2 did not affect the expression of three
pro-inflammatory genes, namely IL-6, TNF-α, and IL-12α (Fig. 4).
Effects of oral administration of LP
06CC2 in mice with DSS-induced colitis
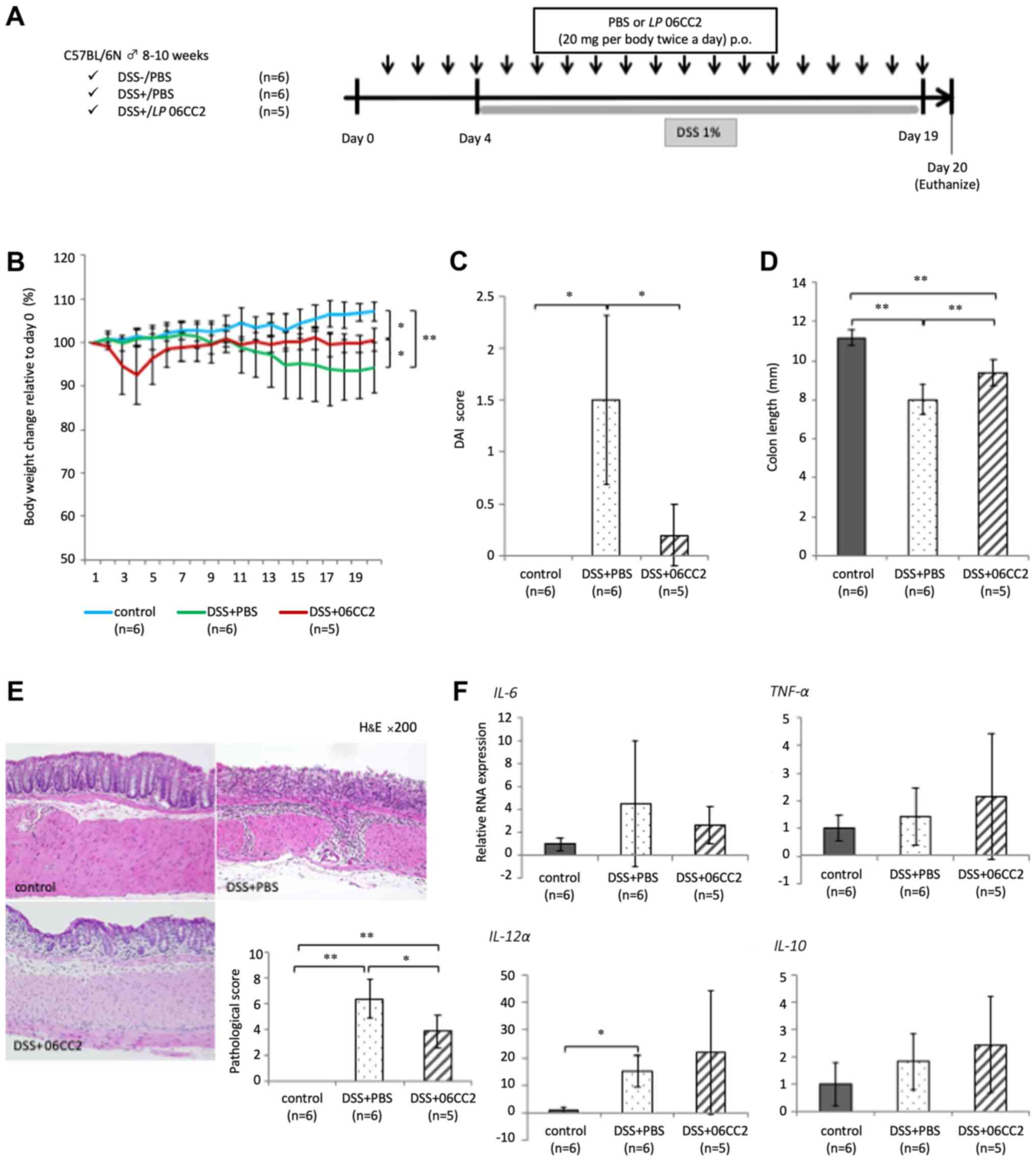
The ability of 06CC2 to prevent colitis was
investigated. After the administration of 06CC2 or
phosphate-buffered saline (PBS) as vehicle by oral gavage for 20
days. Approximately 20 mm of the distal colon was collected and
fixed by 10% phosphate-buffered formalin for histological analysis.
LPMCs were extracted from the remaining colon. Mice with
DSS-induced colitis that received vehicle demonstrated
significantly decreased body weight, while those administered 06CC2
showed marked attenuation of average body weight loss (a 6.5%
decrease in body weight relative to day 0 body weight) and reduced
colon shortening. Thus, we confirmed a transient decrease in the
body weights of DSS+06CC2 treated mice during our preliminary
tests. We performed these experiments several times, and this
phenomenon was easily reproducible, although we note that this
weight loss could also be induced by the physical irritation
associated with oral 06CC2 administration. The disease activity
index (DAI) and pathological score in the 06CC2 group were
significantly lower than in the vehicle group. The expression of
inflammatory cytokine genes such as IL-6 and TNF-α in colonic LPMCs
was elevated in colitis mouse models. The expression of IL-10 was
also slightly higher in the 06CC2 group, but these differences were
not significant (Fig. 5). The
symptoms of clinical and pathological colitis were lessened in the
06CC2 group, despite the fact that the expression of
proinflammatory cytokines such as IL-6 and TNF-α were increased.
Therefore, IL-10 expression may act to mitigate the symptoms of
colitis.
LP 06CC2-treated colitis model mice:
Effect of neutralizing anti-IL-10 antibody
To confirm whether IL-10 was required for the
protective effect of 06CC2 against DSS-induced colitis, mice were
orally administered PBS or 06CC2 twice a day before drinking DSS,
and neutralizing anti-IL-10 monoclonal antibody (100 mg/mouse, once
a day) or control antibody were intraperitoneally administered
after the second administration of PBS or 06CCs, as previously
described (21).
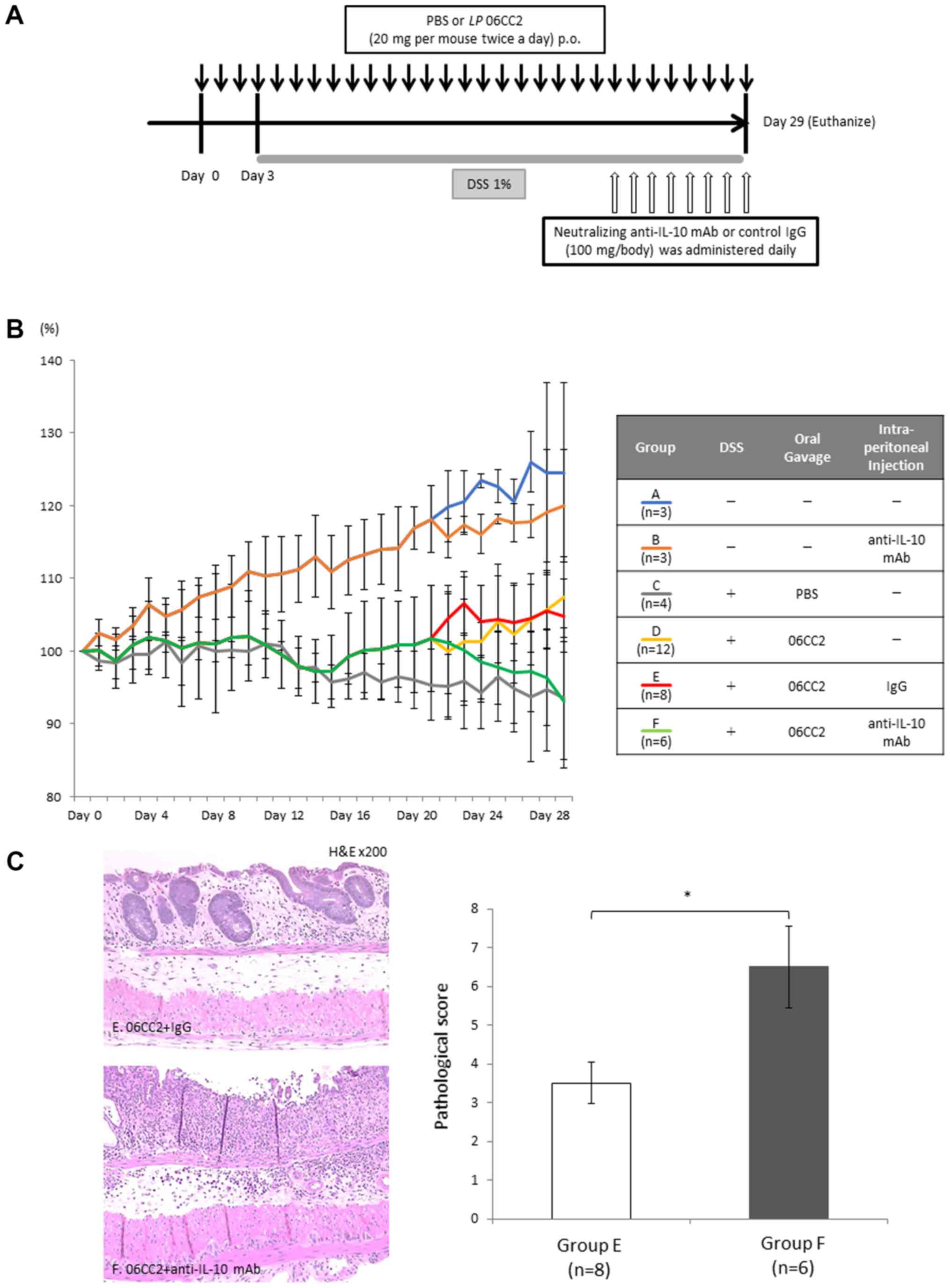
Group A constituted a control group that did not
receive DSS, 06CC2, or intraperitoneal antibody administration;
Group B received only anti-IL-10 neutralizing antibody without DSS
or 06CC2; Group C received DSS and PBS gavage; Group D received DSS
and 06CC2 gavage but no intraperitoneal injection; Group E received
DSS, 06CC2 gavage, and control IgG intraperitoneal injection; and
Group F received DSS, 06CC2 gavage, and intraperitoneal injection
of anti-IL-10 neutralizing antibody. Group F showed attenuated
average body weight loss (93.1% of day 0 body weight), while Group
E mice exhibited no significant changes in body weight.
Pathological examination showed that mucosal layer crypts still
existed in Group E, whereas in Group F these crypts had disappeared
and inflammatory cells had invaded the submucosal layer. The
pathology scores of Group F were worse than those of Group E. These
findings may indicate that the protective probiotic effect of 06CC2
against colitis depends on IL-10 (Fig.
6).
LP 06CC2-treated colitis model mice:
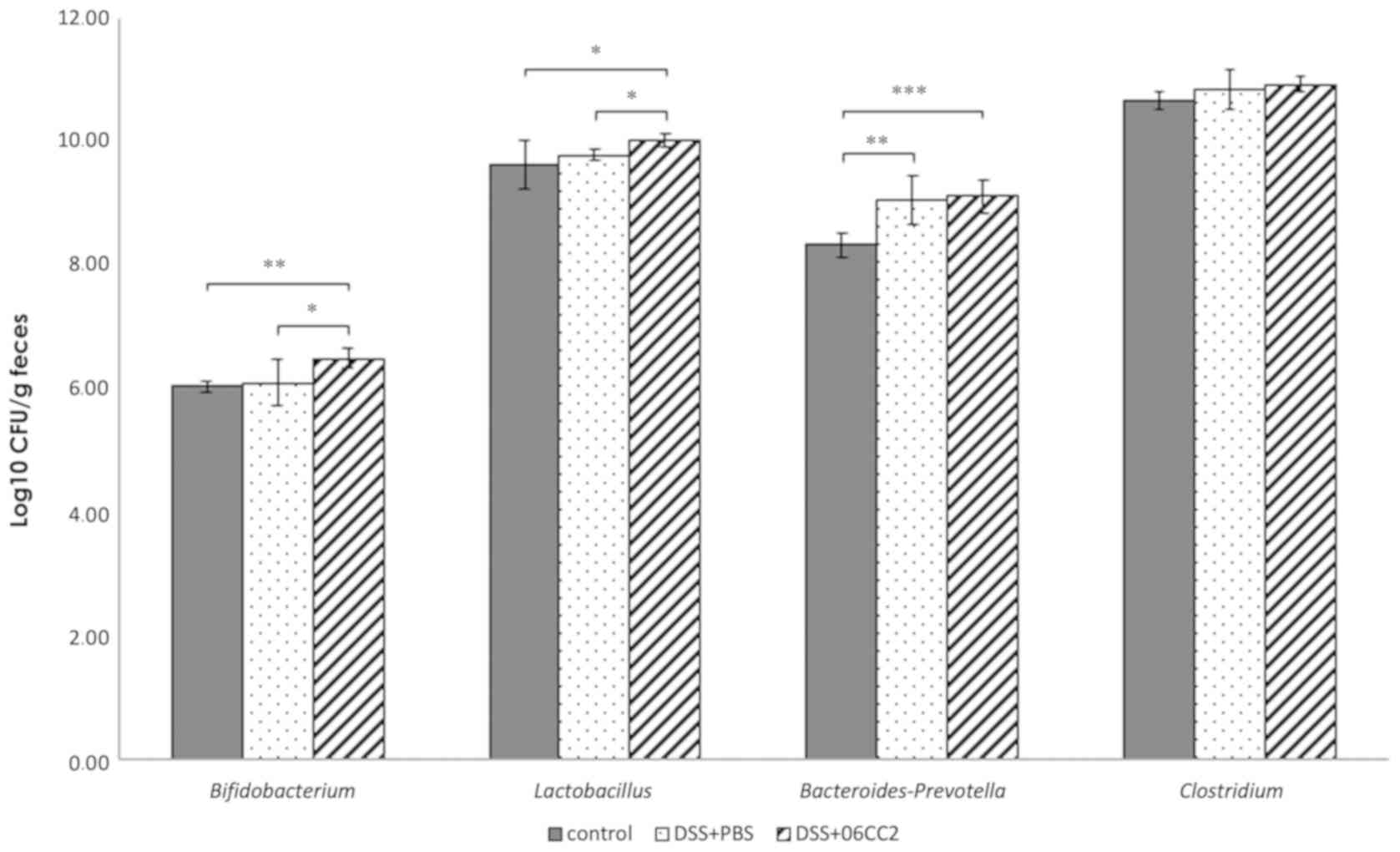
Alteration of the microbiome
The total bacterial counts measured by DAPI staining
did not differ between the control group, the colitis and PBS
group, and the colitis and 06CC2 group. The Lactobacillus
and the Bifidobacterium counts in feces from 06CC2-treated
colitis model mice were significantly higher than those in control
and PBS-treated colitis model mice. Bacteroides-Prevotella and
Clostridium counts were similar regardless of the
administration of 06CC2. Lactobacillus and
Bifidobacterium counts increased significantly in the 06CC2
group during the intake period (Fig.
7).
Discussion
We showed that the 06CC2 strain attenuated
DSS-induced colitis and stimulated the production of IL-10 from
CD11b-positive and CD11c-positive cells in the colon. The 06CC2
strain, which was isolated and identified from traditional
Mongolian dairy products, has beneficial probiotic properties
(13,14). 06CC2 is a useful probiotic lactic
acid bacteria strain for activating immunocompetent cells in the
large intestine. Our data regarding the beneficial effects of 06CC2
are the first to suggest that this strain may prevent colitis.
Several reports demonstrated that intestinal
bacteria in the Firmicutes phylum, which is divided
primarily into the Clostridia and Bacilli classes, also attenuated
DSS-induced colitis (22,23). In particular, Faecalibacterium
prausnitzii, a species in the Firmicutes phylum, was
shown to ameliorate DSS-induced colitis through the TLR2-dependent
modulation of IL-12 and IL-10 production (22). Clostridium cluster IV and XIVa
strains promoted the accumulation of IL-10-producing Treg cells in
the colon and induced resistance to acute DSS-induced colitis
(23). Clostridium
butyricum exerted a protective effect on the epithelial barrier
by maintaining the expression of tight junction proteins; it also
increased the expression of the anti-inflammatory cytokine IL-10,
and decreased levels of pro-inflammatory cytokines such as IL-1β,
TNF, and IL-13 (17,24). In terms of Lactobacillus
members of the Bacilli class, orally administered LP-K68
ameliorated DSS-induced colitis in BALB/c mice by reducing the
production of pro-inflammatory cytokines such as TNF-α, IL-1β, and
IL-6 (12). Lactobacillus
brevis K65 reduced the levels of the pro-inflammatory cytokines
TNF-α, IL-6, and IL-1β (25).
Lactobacillus reuteri F-9-35 prevented DSS-induced colitis
by inhibiting the expression of several pro-inflammatory genes,
namely those for TNF-α, cyclooxygenase-2, and IL-6 (26). Therefore, we focused on 06CC2 under
the hypothesis that its probiotic effect would augment intestinal
immunity in DSS-induced colitis model mice.
We analyzed the levels of anti-inflammatory and
pro-inflammatory cytokines, namely IL-6, TNF-α, IL-12α, and IL-10,
in mice without DSS-induced colitis that were orally administered
06CC2. Regulation of the production of pro-inflammatory mediators
is critical for normal physiological immune function and for
controlling inflammation. The expression levels of IL-6, TNF-α,
IL-12α, and IL-10 showed no significant difference between
DSS-induced colitis mice treated with and without 06CC2 (Fig. 5). Although the expressions of IL-6,
TNF-α, and IL-12 genes were not changed by 06CC2 stimulation, the
expression of IL-10 was significantly increased in 06CC2-treated
mice compared to controls (Fig.
4). Furthermore, the expression of the inflammatory cytokine
genes IL-12α and TNF-α were significantly associated with 06CC2
concentration (expression was 10 to 30 times higher relative to the
0 ng/ml). However, the expression of IL-10 following 06CC2 culture
was much higher relative to the control (80 to 120 times higher),
as shown in Fig. 1. Therefore, we
considered that IL-10 played a key role in these experiments, and
focused on how its expression was altered by 06CC2 stimulation.
It has generally been reported that IL-10 is
produced by CD11b-, CD11c-, and CD4-positive cells, including Treg
cells. We investigated CD11b-, CD11c-, and CD4-positive colonic
LPMCs co-cultured with 06CC2. CD11b- and CD11c-positive cells
produced IL-10 following stimulation by 06CC2. IL-10 production was
much higher in CD11c-positive cells relative to CD11b-positive
cells (Fig. 2), whereas IL-10
production was unaffected in CD4-positive cells (data not shown).
In addition, the transcription factor Foxp3 is specifically
expressed by naturally occurring Treg cells and plays a key role in
their development and function (27–29).
As we observed no changes in the expression of Foxp3 following the
co-culturing of colonic LPMCs with 06CC2 in this study (data not
shown), we suggest that the number of Treg cells in LPMCs was
unaffected by co-culturing, and that other cells were responsible
for IL-10 production. We believe that the main locus of IL-10
production may be CD11c-positive cells.
The function of IL-10 is important for regulating
gut homeostasis during host defense, and IL-10 suppresses the
function of T-lymphocytes and mononuclear cells (30,31).
A genome-wide association study in humans showed that a single
nucleotide polymorphism in the IL-10 gene was closely
associated with IBD (32,33). Thus, the association between IL-10
and IBD has been demonstrated by several findings in both humans
and animal models (34). In fact,
IL-10 knockout mice spontaneously developed colitis in the presence
of commensal bacteria (35). There
are reports that various immune cell types, including regulatory T
cells (Treg cells), macrophages, and dendritic cells, are a major
source of IL-10 in the gut (36–38).
This study used a colitis mouse model to investigate
the effects of prolonged DSS treatment. We used prolonged DSS
treatment because, in terms of the duration of DSS treatment, the
short-term administration of DSS has been used as an acute colitis
model in previous studies, while low-dose and long-term
administration for periods of around 10 days have often been used
as model for chronic colitis (39,40).
We found no differences between the PBS group and the 06CC2 group
after DSS administration for 10 days in our preliminary
experiments. We decided that a 10-day DSS treatment period would be
sufficient to identify any changes in mouse weight.
In this study we co-cultured LPMCs with 06CC2 and
investigated which cells produced IL-10. LPMCs were separated by
MACS as follows: CD11b-positive cells, CD11c-positive cells, and
CD4-positive cells. It has been reported that IL-10 is produced by
CD11b-positive cells, CD11c-positive cells, and Treg cells. We
confirmed that 06CC2 exposure dose-dependently increased the gene
expression of IL-10 in CD11c-positive cells, and to a lesser
extent, CD11b-positive cells (Fig.
2), but not CD4-positive cells (data not shown). Since CD11b
and CD11c are mainly expressed by macrophages and dendritic cells,
respectively, we believe that 06CC2 mainly affected dendritic
cells.
The 06CC2 administered to mice with DSS-induced
colitis resulted in increased numbers of Lactobacillus and
Bifidobacterium bacteria compared with mice not treated with
06CC2. This result occurred despite the fact that heat-killed 06CC2
was administered. Lactobacillus and Bifidobacterium, both of which
have probiotic characteristics, play a pivotal role in the
intestinal environment by preventing pathogen colonization and
maintaining normal mucosal immunity. Increased numbers of
Lactobacillus and Bifidobacterium lead to
downregulation of the gene expression of pro-inflammatory cytokines
such as TNF-α, IL-6, and IL-1β (25–41).
Thus, as a result of changing the composition of the intestinal
microflora, heat-killed 06CC2 may indirectly prevent colitis.
There were some limitations to this study. First, we
demonstrated that 06CC2 suppressed colon inflammation by altering
IL-10 production, but this should be confirmed using genetically
engineered animals such as IL-10 knockout mice. However,
administration of a neutralizing IL-10 antibody to DSS-induced
colitis model mice demonstrated that the protective effects of
06CC2 were mediated by IL-10. Further study should aim to determine
whether the number of IL-10-producing LPMCs is increased in mice
following 06CC2 treatment. Unfortunately, we did not count the
total number of LPMCs because we focused primarily on LPMC gene
expression. However, we believe that our results are valid because
of the number of experimental replicates we performed. MACS could
not be used to strictly classify LPMCs, though CD11b-positive cells
and CD11c-positive cells were considered to be macrophages and
dendritic cells, respectively. It is thus not exactly clear how
06CC2 directly or indirectly affects immune cells, although
TLR2 gene expression in CD11c-positive cells was increased
by 06CC2 stimulation. We showed that 06CC2 may act on macrophages
or dendritic cells to induce IL-10 via TLR2 in vitro, but we
have not demonstrated how the mechanism of 06CC2 differs between
in vivo and in vitro experiments. Data concerning
06CC2 monotherapy in mice is almost non-existent. However, we have
performed a similar study in humans. Participants ingested strain
06CC2 daily for 28 days, resulting in the elevation of fecal
Bifidobacterium abundance (data not shown). Although our
study was limited to only humans, 06CC2 may facilitate
Bifidobacterium abundance in other organisms.
In conclusion, oral administration of the 06CC2
strain suppressed colon inflammation by altering IL-10 production
in colonic dendritic cells and macrophages. We expect that 06CC2
will be available clinically as a therapeutic agent in the
future.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
AT and SK were responsible for the experimental
design. AT, SK, YM and KK performed the animal experiments. AT and
YM performed the colonic histochemical examinations, reverse
transcription-quantitative PCR assays, and colonic ELISAs and were
involved in the analysis and interpretation of data. AT, YM, MT and
ST extracted DNA from fecal samples, performed the intestinal
bacteria assays, and were involved in the analysis and
interpretation of data. SA, FS, YN, ST and SH were responsible for
statistical analyses. AI made substantial contributions to the
conception and design of the study and the drafting of the
manuscript. AT and SK wrote the manuscript. All authors were
involved in manuscript revisions.
Ethics approval and consent to
participate
All experiments were approved by the Animal Care and
Use Committee of Kagoshima University (approval no. MD14106).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Khor B, Gardet A and Xavier RJ: Genetics
and pathogenesis of inflammatory bowel disease. Nature.
474:307–317. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Garrett WS, Lord GM, Punit S,
Lugo-Villarino G, Mazmanian SK, Ito S, Glickman JN and Glimcher LH:
Communicable ulcerative colitis induced by T-bet deficiency in the
innate immune system. Cell. 131:33–45. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Elinav E, Strowig T, Kau AL, Henao-Mejia
J, Thaiss CA, Booth CJ, Peaper DR, Bertin J, Eisenbarth SC, Gordon
JI and Flavell RA: NLRP6 inflammasome regulates colonic microbial
ecology and risk for colitis. Cell. 145:745–757. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Atarashi K, Tanoue T, Oshima K, Suda W,
Nagano Y, Nishikawa H, Fukuda S, Saito T, Narushima S, Hase K, et
al: Treg induction by a rationally selected mixture of clostridia
strains from the human microbiota. Nature. 500:232–236. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Atarashi K, Tanoue T, Shima T, Imaoka A,
Kuwahara T, Momose Y, Cheng G, Yamasaki S, Saito T, Ohba Y, et al:
Induction of colonic regulatory T cells by indigenous Clostridium
species. Science. 331:337–341. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Round JL and Mazmanian SK: Inducible
Foxp3+ regulatory T-cell development by a commensal bacterium of
the intestinal microbiota. Proc Natl Acad Sci USA. 107:12204–12209.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Round JL, Lee SM, Li J, Tran G, Jabri B,
Chatila TA and Mazmanian SK: The Toll-like receptor 2 pathway
establishes colonization by a commensal of the human microbiota.
Science. 332:974–977. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sartor RB: Microbial influences in
inflammatory bowel diseases. Gastroenterology. 134:577–594. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kostic AD, Xavier RJ and Gevers D: The
microbiome in inflammatory bowel disease: Current status and the
future ahead. Gastroenterology. 146:1489–1499. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Llopis M, Antolin M, Carol M, Borruel N,
Casellas F, Martinez C, Espín-Basany E, Guarner F and Malagelada
JR: Lactobacillus casei downregulates commensals'
inflammatory signals in Crohn's disease mucosa. Inflammatory Bowel
Dis. 15:275–283. 2009. View Article : Google Scholar
|
|
11
|
Kelly D, Campbell JI, King TP, Grant G,
Jansson EA, Coutts AG, Pettersson S and Conway S: Commensal
anaerobic gut bacteria attenuate inflammation by regulating
nuclear-cytoplasmic shuttling of PPAR-gamma and RelA. Nat Immunol.
5:104–112. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu YW, Su YW, Ong WK, Cheng TH and Tsai
YC: Oral administration of Lactobacillus plantarum K68
ameliorates DSS-induced ulcerative colitis in BALB/c mice via the
anti-inflammatory and immunomodulatory activities. Int
Immunopharmacol. 11:2159–2166. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Takeda S, Yamasaki K, Takeshita M, Kikuchi
Y, Tsend-Ayush C, Dashnyam B, Ahhmed AM, Kawahara S and Muguruma M:
The investigation of probiotic potential of lactic acid bacteria
isolated from traditional Mongolian dairy products. Anim Sci J.
82:571–579. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takeda S, Takeshita M, Kikuchi Y, Dashnyam
B, Kawahara S, Yoshida H, Watanabe W, Muguruma M and Kurokawa M:
Efficacy of oral administration of heat-killed probiotics from
Mongolian dairy products against influenza infection in mice:
Alleviation of influenza infection by its immunomodulatory activity
through intestinal immunity. Int Immunopharmacol. 11:1976–1983.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Egger B, Bajaj-Elliott M, MacDonald TT,
Inglin R, Eysselein VE and Büchler MW: Characterisation of acute
murine dextran sodium sulphate colitis: Cytokine profile and dose
dependency. Digestion. 62:240–248. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Murthy SN, Cooper HS, Shim H, Shah RS,
Ibrahim SA and Sedergran DJ: Treatment of dextran sulfate
sodium-induced murine colitis by intracolonic cyclosporin. Dig Dis
Sci. 38:1722–1734. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hayashi A, Sato T, Kamada N, Mikami Y,
Matsuoka K, Hisamatsu T, Hibi T, Roers A, Yagita H, Ohteki T, et
al: A single strain of Clostridium butyricum induces
intestinal IL-10-producing macrophages to suppress acute
experimental colitis in mice. Cell Host Microbe. 13:711–722. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cooper HS, Murthy SN, Shah RS and
Sedergran DJ: Clinicopathologic study of dextran sulfate sodium
experimental murine colitis. Lab Invest. 69:238–249.
1993.PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Takeda S, Fujimoto R, Takenoyama SI,
Takedhita M, Kikuchi Y, Tsend-Ayush C, Dashnyam B, Muguruma M and
Kawahara S: Application of probiotics from Mongolian dairy products
to fermented dairy products and its effects on human defecation.
Food Sci Technol Res. 19:245–253. 2013. View Article : Google Scholar
|
|
21
|
Mabalirajan U, Dinda AK, Kumar S, Roshan
R, Gupta P, Sharma SK and Ghosh B: Mitochondrial structural changes
and dysfunction are associated with experimental allergicasthma. J
Immunol. 181:3540–3548. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rossi O, Khan MT, Schwarzer M, Hudcovic T,
Srutkova D, Duncan SH, Stolte EH, Kozakova H, Flint HJ, Samsom JN,
et al: Faecalibacterium prausnitzii strain HTF-F and its
extracellular polymeric matrix attenuate clinical parameters in
DSS-induced colitis. PLoS One. 10:e01230132015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Atarashi K and Honda K: Microbiota in
autoimmunity and tolerance. Curr Opin Immunol. 23:761–768. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li H, Gong Y, Xie Y, Sun Q and Li Y:
Clostridium butyricum protects the epithelial barrier by
maintaining tight junction protein expression and regulating
microflora in a murine model of dextran sodium sulfate-induced
colitis. Scand J Gastroenterol. 53:1031–1042. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu YW, Ong WK, Su YW, Hsu CC, Cheng TH
and Tsai YC: Anti-inflammatory effects of Lactobacillus
brevis K65 on RAW 264.7 cells and in mice with dextran sulphate
sodium-induced ulcerative colitis. Benef Microbes. 7:387–396. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun MC, Zhang FC, Yin X, Cheng BJ, Zhao
CH, Wang YL, Zhang ZZ, Hao HW, Zhang TH and Ye HQ: Lactobacillus
reuteri F-9-35 prevents DSS-induced colitis by inhibiting
proinflammatory gene expression and restoring the gut Microbiota in
mice. J Food Sci. 83:2645–2652. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hori S, Nomura T and Sakaguchi S: Control
of regulatory T cell development by the transcription factor Foxp3.
Science. 299:1057–1061. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fontenot JD, Gavin MA and Rudensky AY:
Foxp3 programs the development and function of CD4+CD25+ regulatory
T cells. Nat Immunol. 4:330–336. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khattri R, Cox T, Yasayko SA and Ramsdell
F: An essential role for Scurfin in CD4+CD25+ T regulatory cells.
Nat. Immunol. 4:337–342. 2003.
|
|
30
|
Maloy KJ and Powrie F: Intestinal
homeostasis and its breakdown in inflammatory bowel disease.
Nature. 474:298–306. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sydora BC, MacFarlane SM, Lupicki M,
Dmytrash AL, Dieleman LA and Fedorak RN: An imbalance in mucosal
cytokine profile causes transient intestinal inflammation following
an animal's first exposure to faecal bacteria and antigens. Clin
Exp Immunol. 161:187–196. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Franke A, Balschun T, Karlsen TH,
Sventoraityte J, Nikolaus S, Mayr G, Domingues FS, Albrecht M,
Nothnagel M, Ellinghaus D, et al: Sequence variants in IL10, ARPC2
and multiple other loci contribute to ulcerative colitis
susceptibility. Nat Genet. 40:1319–1323. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Coelho T, Andreoletti G, Ashton JJ,
Pengelly RJ, Gao Y, RamaKrishnan A, Batra A, Beattie RM, Williams
AP and Ennis S: Immuno-genomic profiling of patients with
inflammatory bowel disease: A systematic review of genetic and
functional in vivo studies of implicated genes. Inflamm Bowel Dis.
20:1813–1819. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Santana G, Bendicho MT, Santana TC, Reis
LB, Lemaire D and Lyra AC: The TNF-α-308 polymorphism may affect
the severity of Crohn's disease. Clinics (Sao Paulo). 66:1373–1378.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hedges SR, Sibley DA, Mayo MS, Hook EW III
and Russell MW: Cytokine and antibody responses in women infected
with Neisseria gonorrhoeae: Effects of concomitant infections. J
Infect Dis. 178:742–751. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Honda K and Littman DR: The microbiome in
infectious disease and inflammation. Annu Rev Immunol. 30:759–795.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Maynard CL, Harrington LE, Janowski KM,
Oliver JR, Zindl CL, Rudensky AY and Weaver CT: Regulatory T cells
expressing interleukin 10 develop from Foxp3+ and Foxp3- precursor
cells in the absence of interleukin 10. Nat Immunol. 8:931–941.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Rubtsov YP, Rasmussen JP, Chi EY, Fontenot
J, Castelli L, Ye X, Treuting P, Siewe L, Roers A, Henderson WR Jr,
et al: Regulatory T cell-derived interleukin-10 limits inflammation
at environmental interfaces. Immunity. 28:546–558. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Okayasu I, Hatakeyama S, Yamada M, Ohkusa
T, Inagaki Y and Nakaya R: A novel method in the induction of
reliable experimental acute and chronic ulcerative colitis in mice.
Gastroenterology. 98:694–702. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Alex P, Zachos NC, Nguyen T, Gonzales L,
Chen TE, Conklin LS, Centola M and Li X: Distinct cytokine patterns
identified from multiplex profiles of murine DSS and TNBS-induced
colitis. Inflammatory Bowel Dis. 15:341–352. 2009. View Article : Google Scholar
|
|
41
|
Hsieh MC, Tsai WH, Jheng YP, Su SL, Wang
SY, Lin CC, Chen YH and Chang WW: The beneficial effects of
Lactobacillus reuteri ADR-1 or ADR-3 consumption on type 2
diabetes mellitus: A randomized, double-blinded, placebo-controlled
trial. Sci Rep. 8:167912018. View Article : Google Scholar : PubMed/NCBI
|