Introduction
Chlamydia is a genus of gram-negative
obligate intracellular bacteria consisting of nine recognized
species; each species exhibits specific tissue tropism and disease
pathology (1). The global impact
of Chlamydia trachomatis (C. trachomatis) is
considerable, and of significant medical concern in humans
(2). C. trachomatis exists
as 19 serovars; serovars A-C are responsible for trachoma, the
leading infectious cause of blindness worldwide (3). Serovars D-K primarily infect the
genital mucosae, causing numerous commonly-diagnosed sexually
transmitted diseases, including hydrosalpinx, a
laparoscope-detectable marker of tubal factor infertility (4). Moreover, C. trachomatis is
also a major risk factor in the transmission of human
immunodeficiency virus (5). To the
best of our knowledge, no study to date has determined why C.
trachomatis can cause infectious blindness, or how infection of
the lower genital tract can result in tubal fibrosis and
hydrosalpinx. Therefore, the aim of the present study was to
determine the molecular mechanisms of C. trachomatis
pathogenicity, and to guide the design of live-attenuated vaccine
strains for the prevention of chlamydial diseases.
As with all other chlamydia, C. trachomatis
possesses a unique intracellular growth cycle with a distinct
biphasic developmental cycle, alternating between an infectious
elementary body (EB) and a replicating, metabolically-active
reticulate body (RB) (6). EBs
differentiate into RBs within a non-acidified vacuole, the
chlamydial inclusion (7). At ~18 h
post-infection, the generated progeny differentiate back into EBs.
Later in the developmental cycle, EBs are released from the host
cell to initiate a new cycle of infection (2).
Host inflammatory responses triggered by chlamydial
intracellular survival and replication contribute to
chlamydia-induced pathologies; secretory proteins, including
chlamydia protease-like activity factor (CPAF), have been
hypothesized to play important roles in this process (8). CPAF, secreted into the cytosol of
C. trachomatis-infected cells, is involved in the immune
evasion mechanism that may block NF-κB p65 nuclear translocation,
resulting in decreased interferon-β and pro-inflammatory cytokine
synthesis (9). Despite extensive
research into CPAF, the mechanism of C.
trachomatis-associated trachoma and infertility following
infection remains unknown. Asmost chlamydia-infected host cells
secrete chlamydial proteins, other secreted proteins are also
likely to notably contribute tothe pathogenicity of chlamydial
infections in humans.
In a previous study, in fixed C.
trachomatis-infected HeLa cells analyzed with antibodies
against chlamydia glycogen metabolism-related enzymes, only C.
trachomatis glycogen synthase (GlgA) was found to be secreted
into the host cell cytosol (10).
It was first revealed to be associated with chlamydial inclusion
bodies at 12 h post-infection, and secretion into the cytosol was
detectable at ~24 h post-infection. However, since glycogen was
only monitored in the inclusion bodies, and not the cytosol, it is
unclear whether GlgA secretion into the host cell cytosol is
necessary for the induction of chlamydial diseases.
GlgA expression is dependent on a cryptic plasmid;
removal of this plasmid results in the loss of GlgA expression and
attenuated pathogenicity in both C. trachomatis serovar A
and C. muridarum (11).
These findings indicate that GlgA may play an essential role in
chlamydial pathogenesis.
The yeast two-hybrid system enables the in
vivo detection of interacting proteins in order to reveal the
biological roles of a known protein (12). Following a series of optimizations
and development by Fields and Song (13), the yeast two-hybrid system was
considered to bea classical method ofidentifying and studying
protein-protein interactions. In a recent studyusing the yeast
two-hybrid system, the C. trachomatis inclusion membrane
protein MrcAwas found tointeract with inositol 1,4,5-trisphosphate
receptor type 3 to regulate extrusion formation (14). Thus, due to its inexpensive and
time-saving nature, the yeast two-hybrid system is a powerful
method for the analysis of protein-protein interactions.
In the present study, the yeast two-hybrid system
was used to identify proteins that interact with GlgA. This
strategy involved screening 13 potential clones, which following
cDNA identification, were confirmed via rotary validation and
co-immunoprecipitation. The results indicated that prohibitin (PHB)
interacts with GlgA, which may provide novel insight into the
understanding of GlgA in chlamydial biology and pathogenesis.
Materials and methods
Bait plasmid construction
The Matchmaker two-hybrid system (Clontech
Laboratories, Inc.) was used to confirm the potential interaction
partners of GlgA (CT798). The gene sequence of CT798 (WP-100139618)
was obtain from the National Centre for Biotechnology Information
database (https://www.ncbi.nlm.nih.gov/) and amplified by PCR
using the following primers, which contained SfiI sites at
both ends: CT798 forward,
5′-AAGGCCATTACGGCCATGAAAATTATTCACACAGCTATCG-3′ and reverse,
5′-CCGGCCGAGGCGGCCTTGTTTATAAATTTCTAAATATTTATTGGC-3′.
PCR was performed under the following conditions:
Initial denaturation at 98°C for 5 min, amplification (30 cycles)
at 98°C for 30 sec, 55°C for 30 sec and 72°C for 42 sec, and afinal
extension at 72°C for 5 min (Qiagen). The FastPfu DNA Polymerase
was used (Beijing Transfen Biotech Co., Ltd.). The orientation and
authenticity of the plasmid sequence was confirmed by sequencing
(Sangon Biotech Co., Ltd.) and restriction endonuclease digestion
by 8% SfiI digestion (Fermentas) at 50°C for 24 h.
Recombinant plasmid confirmation
The CT798 plasmid was cloned into the GAL4 binding
domain of the pGBKT7 vector (Hitech Bio-Technology Co., Ltd). CT798
plasmids were treated with SfiI in a 25-µl reaction system
including 20 µl CT798, 2 µl SfiI restriction enzyme, 2.5 µl
10X buffer and 0.5 µl sterile water. pGBKT7 was treated with
SfiI in a 20-µl reaction system including 10 µl CT798, 1 µl
SfiI restriction enzyme, 2 µl 10X buffer and 7 µl sterile
water. The digested reaction mixtures were then combined and
incubated at 16°C for 1 h, consisting of 5 µl Ligation High Buffer
(Toyobo Life Sciences), 5 µl CT798/SfiI and 0.5 µl
pGBKT7/SfiI. The recombinant plasmid wasthenused to
transform TOP10 Escherichia coli. Escherichia coli was
supplied by American Type Culture Collection and cultured in
Luria-Bertani medium (1% NaCl; 1% Polypeptone; 0.5% Yeast extract)
in a humidified incubator at 37°C with 5% CO2. Then, it
was inoculated onto Luria-Bertani medium plates (1% NaCl; 1%
Polypeptone; 0.5% Yeast extract; 2% agar) containing 50 µg/ml
kanamycin (LB-Kanr+) overnight at 37°C. A total of six
bacterial colonies were selected and further cultured at 16°C
overnight with agitation (250 × g). The plasmids of the cultured
bacteria were extracted using a Plasmid Miniprep kit (Qiagen, Inc.)
and verified by sequencing.
Agarose gel electrophoresis
Agarose gel electrophoresis is a standard method for
identification DNA fragments (15). In the present study, 0.7% agarose
gel was used. Then, 0.14 g agarose and 20 ml 1X Tris-acetate-EDTA
buffer in a flask were heated in a microwave for 5 min at 100°C.
Then, 2 µl ethidium bromide was added (Thermo Fisher Scientific,
Inc.), and poured onto a taped plate with casting combs. Then, 2 µl
DNA sample from Escherichia coli was added to the 5X agarose
gel, and underwent electrophoresis at 120 mA for 40 min at 25°C
until separation had been achieved. The DNA fragments were
visualized using a long wave UV light box.
Self-activation test
pGADT7, pGBKT7-p53, pGBKT7-laminC and the
pGADT7-170297-LUcDNA plasmid genome were provided from Hitech
Bio-Technology Co., Ltd. The pGADT7-170297-LUcDNA plasmid genome
was referred to as ‘pGADT7-largeT’. As a positive control,
pGBKT7-p53 and pGADT7-largeT were used to transform AH109 yeast
cells. As a negative control, AH109 cells were also transformed
using pGBKT7-lamin C with pGADT7-large T. AH109 were purchased from
the American Type Culture Collection and cultured in yeast peptone
dextrose adenine (YPDA; 1% Yeast extract; 2% Tryptone; 2% Glucose;
0.02% Adenine) with 5% CO2. Synthetic defined
(SD)/-tryptophan (Trp), SD/-leucine (Leu), SD/-Trp/-Leu,
SD/-Trp/-Leu/-histidine (His) and SD/-Trp/-Leu/-His/-adenine (Ade)
media were purchased from Clontech Laboratories, Inc.
To test whether the prey plasmid was able to
auto-activate the reporter genes, pGADT7 and pGBKT7-CT798 were used
to co-transform AH109 cells according to the lithium acetate
transformation method (16). The
cells were then cultured in SD medium lacking various amino acids
(SD/-Trp/-Leu) at 30°C for 4 days. The isolated yeast clones were
tested for the activation of the third reporter gene, including
imidazoleglycerol-phosphate dehydratase (HIS3),
phosphoribosylaminoimidazole carboxylase (ADE2) and LacZ.
Yeast two-hybrid screening
The pGADT7-170297-LUcDNA plasmid genome from HeLa
cells was used for the yeast two-hybrid screen (17). AH109 yeast cells containing the
pGBKT7-CT798 plasmid were plated and selected using SD agar plates
without tryptophan. The positive clones were inoculated in 50 ml
SD/-Trp medium and cultured at 30°C (225 × g) for 18 h. The cells
were then transferred to 500 ml YPDA medium and cultured at 30°C
(agitation at 225 × g) for 4 h until OD600=0.6. Next,
the cultured cells were centrifuged at 4,000 × g for 5 min at room
temperature; the supernatant was discarded and the pellet was
resuspended in 0.1 MLiAc of 20 ml. The cells were then centrifuged
once more under the same conditions and suspended in 10 ml LiAc
(0.1 M). Then, 9.6 ml 50% PEG3350, 1.44 ml LiAc, 300 µl
single-stranded DNA and 25 µg cDNA library plasmid were added, and
the cells were cultured at 30°C for 30 min. The mixture was heated
in a water bath at 42°C for 25 min, followed by resuscitation at
30°C for 1 h. The cell pellet was resuspended in 8 ml sterile water
following centrifugation at 4,000 × g for 5 min at room
temperature. The cells were cultured on SD/-Leu/-Trp/-His and
3-amino-1,2,4-triazole (3AT) plates with 200 µl solution/plate, for
a total of 40 plates. In order to determine library transformation
efficiency, 20 µl cell cultures from 8 ml solution were diluted
10-fold by gradient and cultured on SD/-Leu/-Trp plates. The cells
were cultured for 3–4 days at 30°C, with observation ofthe
conversion results and recording of conversion efficiency. The
following calculations were used: Total number of transformants =
(n1/20+n2/2+n3/0.2)×1/3×8,000).
Where n1, the number of colonies growing on the plate
after 10-fold dilution; n2, the number of colonies
growing on the plate after 100-fold dilution; n3, the
number of colonies growing on the plate after 1,000-fold dilution.
Transformation efficiency = Total number of transformants/25 µg
cDNA plasmid.
Positive clone verification
After culturing on SD/-Leu/-Trp/-His and 3AT plates,
the positive clones were plated onto SD/-Leu/-Trp plates. To select
for clones which potentially interacted with CT798, the clones were
inoculated onto SD/-Leu/-Trp/-His+3AT and SD/-Leu/-Trp/-Ade plates
to assess the transcriptional activation of the HIS3 and ADE2
reporter genes, and resuspended in sterile water to detect LacZ
expression. Concurrently, positive and negative control
transformation experiments were conducted, for the purpose of
selecting true positive colonies. The positive clone plasmids were
subsequently extracted using the Plasmid Miniprep kit. Following
sequencing, a BLAST search was conducted using GenBank (National
Center for Biotechnology Information) to determine the associated
genes.
Rotary validation
The selected clones were used to transform AH109
yeast cells containing pGBKT7-CT798 as aforementioned, and these
cells were cultured on SD/-Leu/-Trp plates. Positive clones were
transferred onto SD/-Leu/-Trp/-His/-Ade+3AT and SD/-Leu/-Trp/-Ade
plates to determine the transcriptional activation of the HIS3 and
ADE2 reporter genes, and resuspended in sterile water to detect
LacZ expression.
Co-immunoprecipitation
HeLa cells were provided from the University of
South of China and cultured in complete DMEM (Gibico; Thermo Fisher
Scientific, Inc), containing 10% FBS (Gibico; Thermo Fisher
Scientific, Inc.) in a humidified incubator at 37°C with 5%
CO2. HeLa cells were infected with C.
trachomatis. C. trachomatis was added into HeLa cell. At
48 h post-infection, the cells were washed twice with cold PBS, and
lysed with RIPA buffer (Epizyme, Inc.) for 15 min at 4°C. The
lysate was pre-clarified by rotation for 10 min, centrifuged at
14,000 × g for 15 min (both 4°C), and incubated with protein-A
agarose beads. Cell-free lysates were incubated with a rabbit
anti-PHB antibody (1:1,000; Cell Signaling Technology, Inc.; cat.
no. ab75766.) or IgM antibody (1:1,000; Cell Signaling Technology,
Inc.) at 4°C overnight. Protein-A beads were added for 2 h at room
temperature, and the lysates were subsequently washed three
timeswith PBS. The proteins were eluted into 5X SDS-PAGE sample
buffer, separated by 12% SDS-PAGE gel for immunoblotting.
Western blotting
HeLa cells were infected with C. trachomatis
and harvested at 48 h post-infection. Cells were washed with cold
PBS and lysed with RIPA buffer (Epizyme, Inc.) with protease
inhibitor and phosphatase inhibitor. Then, 10 µl protein extracts
and cell lysates (Input control) were separated by 12% SDS-PAGE gel
and electrophoretically transferred to nitrocellulose membranes.
After blocking in 5% non-fat dry milk with TBST (50 mM Tris-HCl; pH
7.4; 5.36 mM KCl; 274 mM NaCl; 0.1% Tween-20) for 2 h at room
temperature, the membranes were probed with a mouse anti-CT798
antibody (1:1,000; The University of South China) overnight at 4°C.
Then, the membrane waswashed four times for 5 min in TBST and
incubated with anti-mouse horseradish peroxidase-conjugated
secondary antibody (1:3,000; Servicebio, Inc.; cat. no. GB23303)
for 1 h at 37°C Blots were detected by Western Blot system G: BOX
Chemi XXX9 (Syngene International Ltd.).
Results
Construction of the pGBKT7-CT798 bait
plasmid
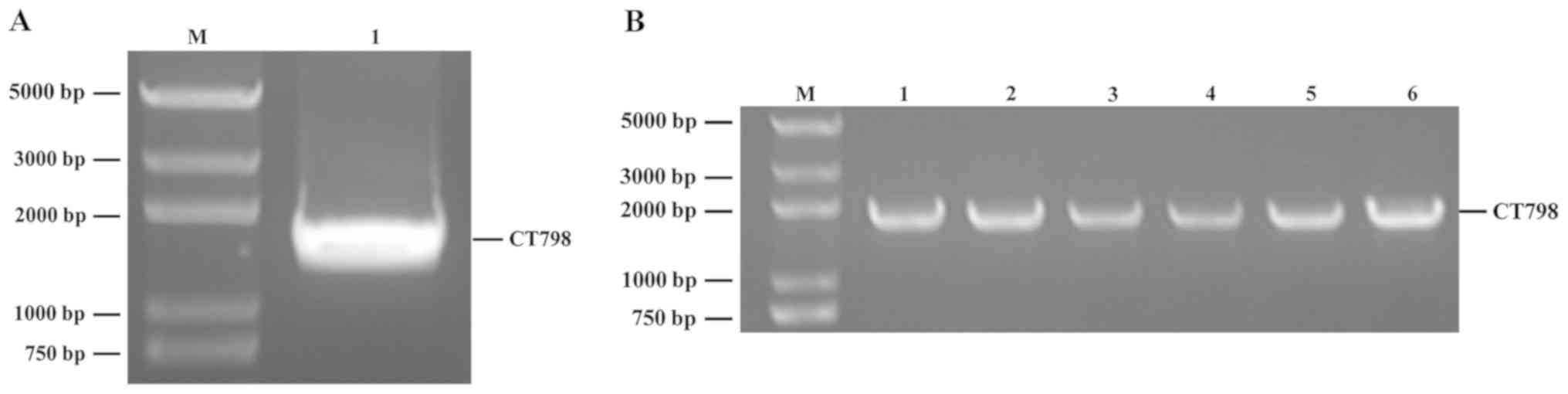
CT798 is 53 kDa; the full length of CT798 (1,440 bp)
was successfully amplified from C. trachomatis genomic DNA
viaPCR, and analyzed using restriction enzyme digestion (Fig. 1A). The PCR products were
subsequently cloned into the bait vector pGBKT7. The construct was
transformed into E. coli and inoculated onto
LB-kanr+ plates. Following incubation, six positive
transformants were randomly picked and identified by PCR (Fig. 1B), and the results of recombinant
plasmid sequencing were consistent with that of pGBKT7-CT798.
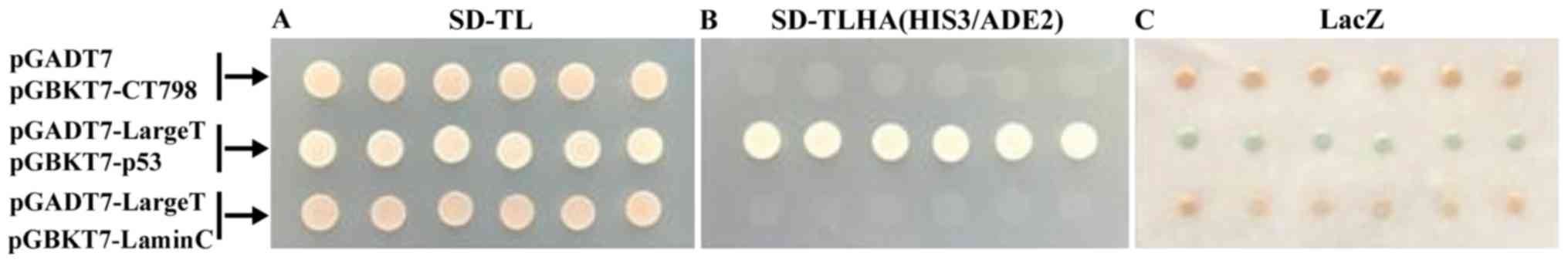
Validity of the bait plasmid
pGBKT7-CT798
The AH109 yeast strain contains various nutritional
reporter genes. These include: i) ADE2, coding for
phosphoribosylaminoimidazole carboxylase, an important enzyme for
adenine synthesis; ii) HIS3, encoding imidazoleglycerol-phosphate
dehydratase, another key enzyme for adenine synthesis; and iii)
LacZ, which encodes β-galactosidase enzymes. Therefore, AHI09
allows for the control of strong nutritional selection, whilst
reducing the incidence of false positives. To determine the
efficiency of the system, pGBKT7-CT798 and pGADT7 were used to
transform AH109 cells, and pGBKT7-p53 and pGBKT7-laminC were used
as the positive and negative controls, respectively. AH109 cells
were then cultured on SD/-Leu/-Trp plates, and six transformants
were used to test for self-activation, including pointing-plate
culturing on SD/-Leu/-Trp/-His/-Ade plates to test for the
expression of HIS3 and ADE2, and a colorimetric assay to determine
the expression of LacZ. Control strains both grow on the
SD/-Leu/-Trp plate, but only the positive control cells are able to
grow on the SD/-Leu/-Trp/-His/-Ade plate. In the present study,
AH109 cells containing the pGBKT7/CT798 plasmid grew on the
SD/-Leu/-Trp plate, but not on the SD/-Leu/-Trp/-His/-Ade plate.
Furthermore, the results of the colorimetric assay were consistent
with those of the negative control (Fig. 2). These results suggest that CT798
does not possess self-activation function in AH109 cells.
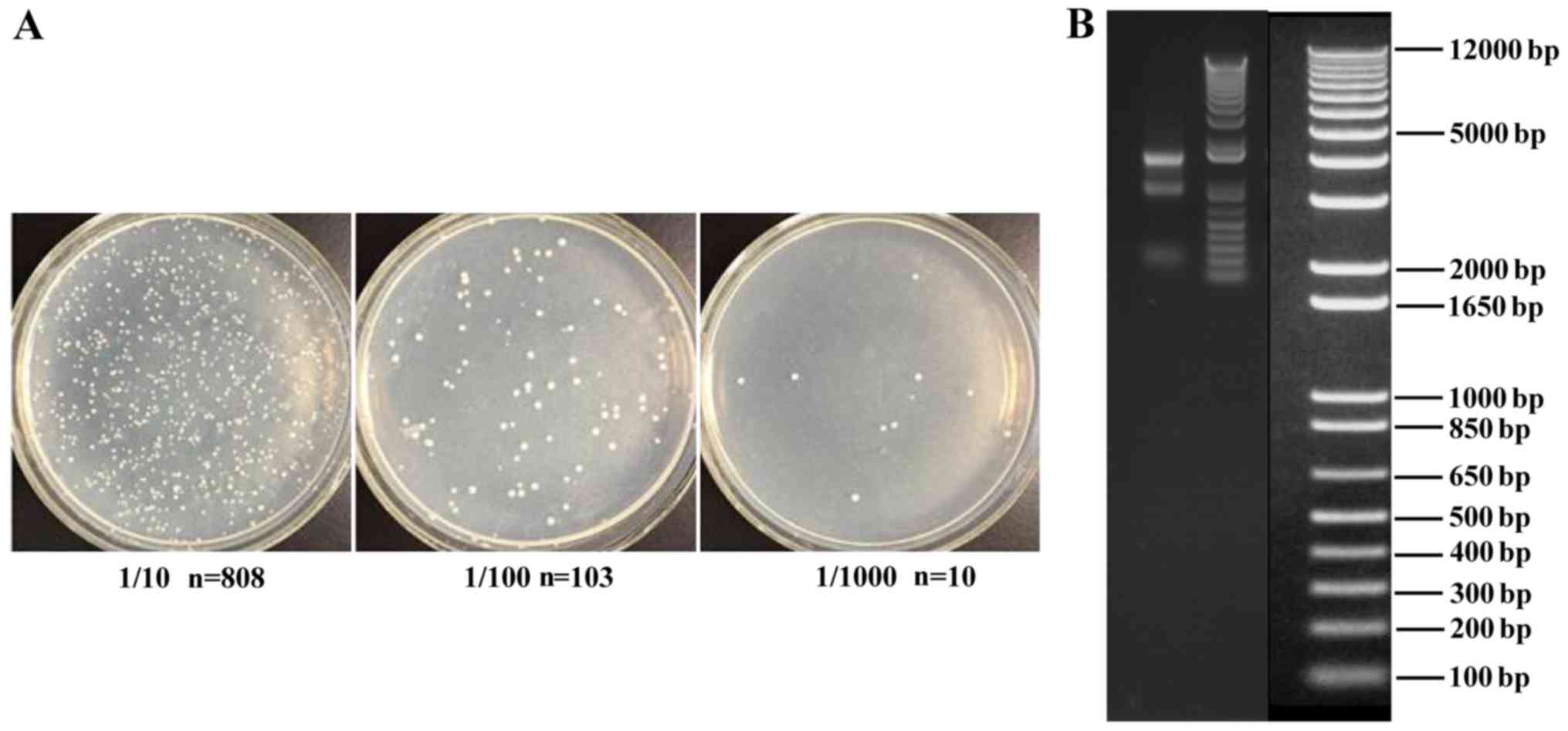
cDNA library screening
pGADT7-170297-LUcDNA library used for screening was
supplied by Hitech Bio-Technology Co., Ltd. Plasmids contained only
pGBKT7-CT798 as bait for screening the pGADT7-170297-LUcDNA
library. Library plasmids were transformed into AH109 cells
containing bait plasmid and cultured on SD/-Leu/-Trp/-His and 3AT
plates. AH109 cells were also cultured on SD/-Leu/-Trp plates
(Fig. 3), and transformation
efficiency (which is important for the success of the yeast
two-hybrid system) was determined. The results revealed a total of
3.78×105 transformants, indicating a transformation
efficiency of 1.51×104/µg. At the same time, the quality
of the yeast two-hybrid library was assessed. Agarose gel
electrophoresis of the PCR products indicated the average length of
the insertion product to be ~1.5 kb, and the positive success rate
of the library was 100%.
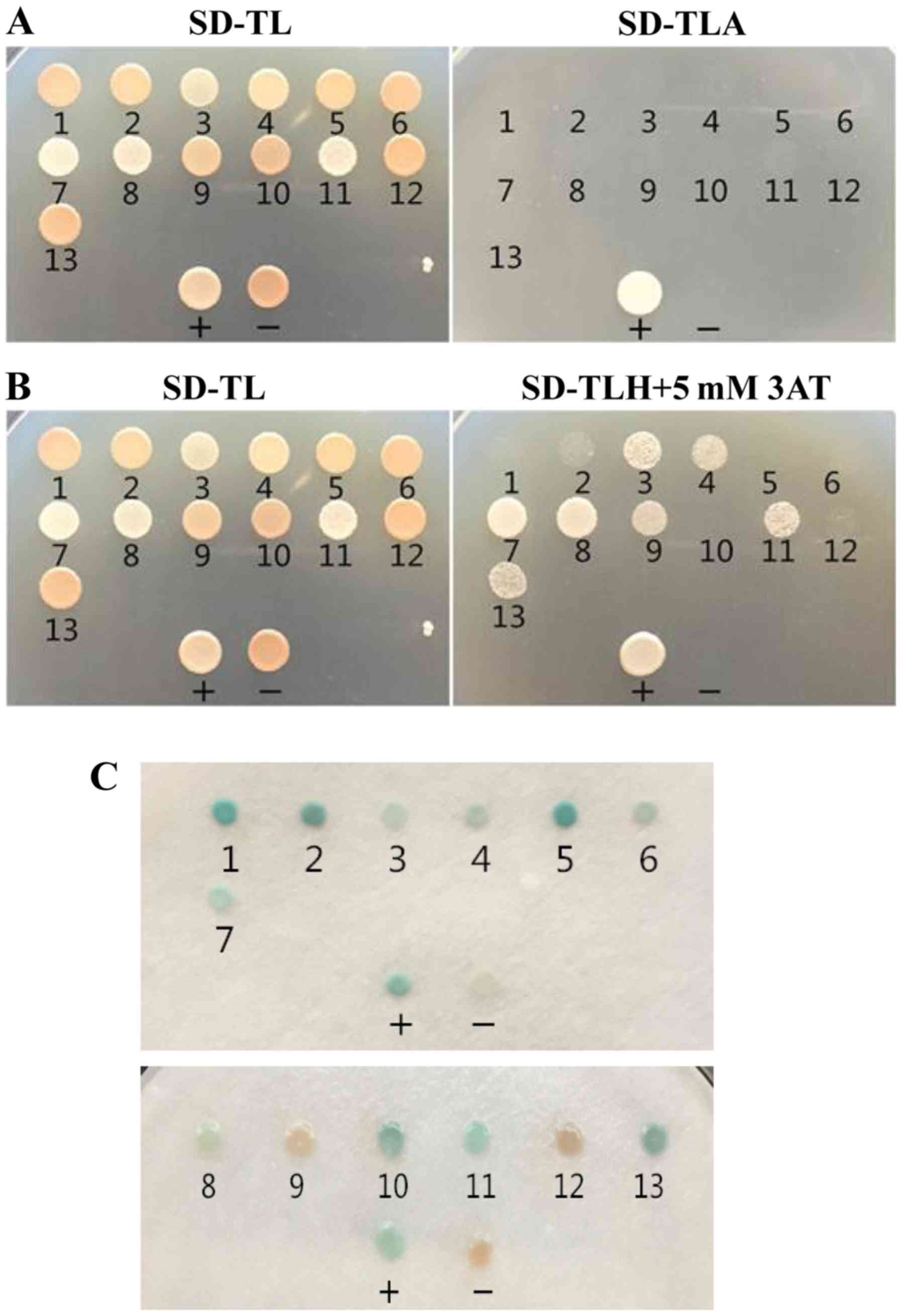
Identification of proteins interacting
with the bait protein
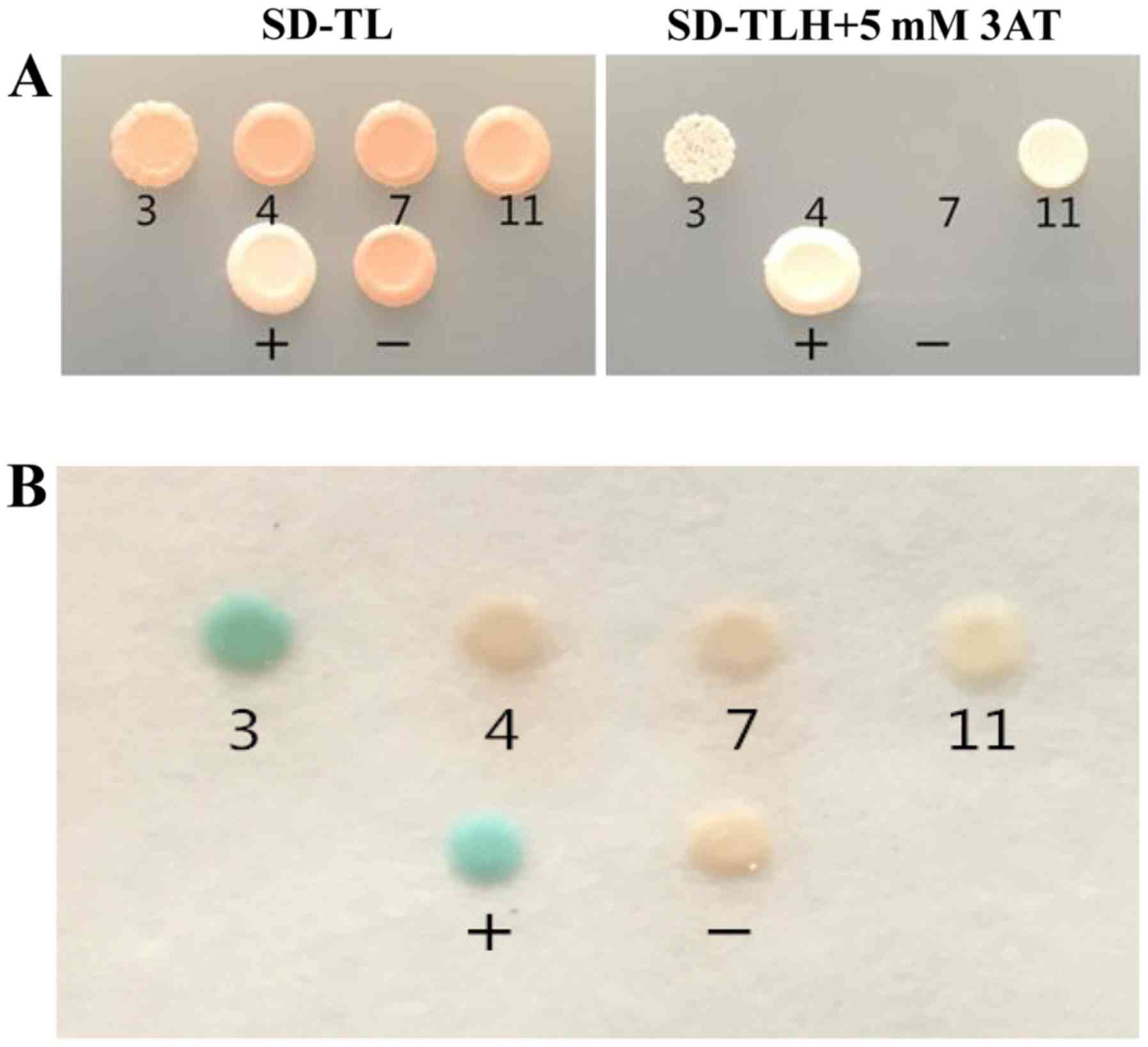
In the present study, 13 positive clones were
obtained from two rounds of selection, which were designated A1-13.
These clones were not capable of growing on SD/-Leu/-Trp/-Ade
plates, and seven grew on SD/-Leu/-Trp/-His and 3AT plates
(Fig. 4). In addition, 11 of the
13 clones expressed LacZ. These results indicated that the HIS3
gene was activated by the seven positive clones, and further
suggested that A3, A4, A7, A8, A9, A11 and A13 interacted with
CT798 to a certain degree. The positive clones were subsequently
sequenced and analyzed using BLAST software. The results indicated
that the seven positive clones possessed a high degree of
similarity to known genes encoding four different proteins
(Table I). Finally, for rotary
validation, the resulting four plasmids were used to transform
AH109 cells containing the pGBKT7-CT798 plasmid (Fig. 5). The results revealed that clones
A3 and A11 were able to active the HIS3 gene, and that only A3
activated the LacZ gene.
 | Table I.Biological information of the seven
positive clones. |
Table I.
Biological information of the seven
positive clones.
| Clone(s) | NCBI accession | NCBI
description |
|---|
| A3 | XM_017025718.1 | PREDICTED: Homo
sapiens CXXC finger protein 1, transcript variant X2, mRNA |
| A4, A9 | XM_017024763.1 | PREDICTED: Homo
sapiens prohibitin, transcript variant X2, mRNA |
| A7, A8 | NM_001256139.1 | Homo sapiens
capping actin protein, gelsolin-like, transcript variant 2,
mRNA |
| A11, A13 | AK298222.1 | Homo sapiens
cDNA FLJ56357 complete cds, highly similar to Homo sapiens
apolipoprotein A-I binding protein, mRNA |
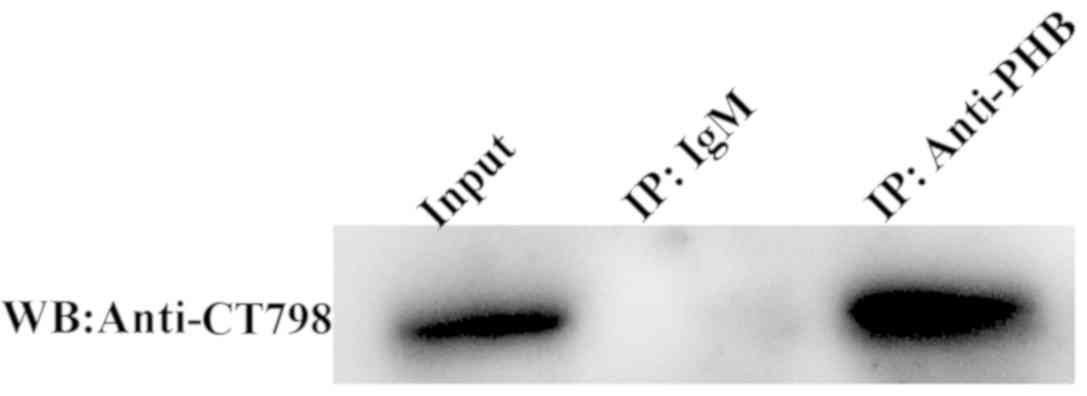
PHB interacts with CT798
Co-immunoprecipitation was performed to validate the
interaction between CT798 and PHB (Fig. 6). CT798 was co-precipitated by
anti-PHB from the lysates of cells infected with C.
trachomatis, but not by precipitation with IgM. This suggested
that PHB interacts with CT798.
Discussion
GlgA is a key enzyme involved in glycogen
metabolism, which catalyzes the glucose moiety of UDP-glucose
transferred to the non-reducing end of a linear α-1, 4-glucan
primer. Previous studies have reported that the inactivation of
GlgA not only inhibits glycogen synthesis, but also reduces
bacterial stress resistance (18,19).
It is possible that GlgA is associated with adaptive environmental
survival, colonization and virulence. In addition, the average
chlamydial genome size is ~1.0 Mb, only 1/4 of that of E.
coli (20). Notably, the
expression and secretion of GlgA is highly conserved among
different serovars of C. trachomatis and C.
muridarum, a separate, but related species (21). The importance of GlgA in chlamydial
growth and development is well established (10). However, the effects of GlgA
secretion into the host cell and its involvement in chlamydial
metabolism (which may serve a role in the association of GlgA
secretion with the inflammatory response) are yet to be
investigated. Thus, screening for binding partners of GlgA using
the yeast two-hybrid system will enhance current understanding of
the role of GlgA in chlamydial biology and pathogenesis.
Yeast two-hybrid technology, an effective means of
detecting unidentified protein-protein interactions, has allowed
for further exploration of biological pathways and their regulation
(22). In the present study, not
only were positive and negative controls used, but promoters
modulating HIS3, ADE2 and LacZ reporter gene expression in the
AH109 yeast strain were also included to exclude false-positive
results. Although the yeast two-hybrid assay is associated with
high sensitivity and easy operation, it exhibits obvious drawbacks,
and false positives are still easily obtained. Hence, other in
vitro assays (such as immunoprecipitation) should be conducted
to verify these results and improve data credibility.
To enhance the probability of identifying
unambiguous interactions, stringent media were used, which lacked
the amino acid histidine or the nucleic acid adenine (23). Growth-dependent selection enables
the most effective identification of interaction partners.
Therefore, only bait-prey interactions result in HIS3 or ADE2
reporter gene expression; LacZ expression provides a reference to
reflect the strength of the interactions. The identified
interaction proteins were Homo sapiens CXXC finger protein 1
(CFP1), PHB, gelsolin-like actin-capping protein (CAPG) and
apolipoprotein A-I binding protein (ApoA-I).
CFP1, an unmethylated CpG-binding protein, is a
component of the mammalian set1 histone methyltransferase complex,
which is involved in histone methylation, regulating T-cell
development and promoting cellular differentiation (24). Anti-chlamydial T cells may
represent a double-edged sword, and may be responsible for
initiating the pathological changes associated with chlamydial
infection (25). Thus, T-cell
secretion of GlgA and CFP1 may promote chlamydial persistence and
immunopathology.
PHB is characterized as an anti-proliferative gene,
and is widely distributed in different cellular compartments, such
as the plasma membrane, nucleus and mitochondria (26). PHB family members possess two
subunits, PHB1 and PHB2; PHB1 binds to the transcription factor
p53, inducing p53-inducible gene 3 promoter binding and promoting
p53-dependent cell apoptosis (27). PHB2 can translocate from the
mitochondria to the nucleus during capsaicin-induced apoptosis
(28). PHB1 and 2 are conserved
mitochondrial proteins with diverse functions, which include roles
in cell proliferation and mitochondrial integrity (29). As obligate intracellular pathogens,
chlamydia have been proposed to be anti-apoptotic to complete their
development cycle (30).
Accordingly, PHB is hypothesized to shuttle between the
mitochondria and the nucleus during the cell cycle, and GlgA is
secreted into the cytosol at ~24 h post-infection (31). These findings indicate that at the
beginning of the developmental cycle, the manipulation of GlgA by
C. trachomatis may prevent premature release of inclusion
bodies from the cell via its interaction with PHB.
The actin-binding protein CAPG was originally
identified in the cytoplasm of alveolar macrophages (32). CAPG interacts with cytoplasmic and
nuclear proteins that are closely associated with cell
proliferation and motility (33).
It promotes restructuring of the actin cytoskeleton by capping the
fast-growing end of actin filaments in a Ca2+ and
phosphatidylinositol 4,5-bisphosphate-dependent manner, but does
not initiate actin filament severing (34). Furthermore, CAPG may be involved in
mediating cross-talk between the actin cytoskeleton and the
microtubule-based organelles that regulate cell division (35). C. trachomatis has evolved
exclusion mechanisms of host-cell escape, through which
membrane-bound inclusions are released from the host cell viaa
process resembling exocytosis (36). This process is hypothesized to be
dependent on cytoskeletal activities, including actin
polymerization, as well as neural Wiskott-Aldrich syndrome protein,
Rho GTPase and the myosin motor complex (37). The results of the present study
demonstrated that CAPG may interact with GlgA to regulate extrusion
formation, which facilitates evasion of the localized inflammatory
response at the site of infection.
ApoA-I is the primary constituent of plasma
high-density lipoprotein (HDL) and plays a central role in reverse
cholesterol transport (RCT) (38).
The process of RCT, which transports cellular cholesterol back to
the liver, is considered to be one of the most important
anti-atherogenic properties of HDL (39). Several experimental strategies
aimed at overexpressing ApoA-I are associated with the
anti-atherogenic effects of HDL (40). Besides upper respiratory infection,
chlamydial challenge has also been associated with chronic
inflammatory conditions, including atherosclerosis (41). The interaction between GlgA and
ApoA-I has been speculated as a novel target for the treatment and
prevention of severe long-term sequelae of chlamydial infections.
As these interacting proteins are known to be associated with
immunoregulation, apoptosis and tumor progression, it is possible
that GlgA operates in a similar manner.
In the present study, four potential
GlgA-interaction proteins were identified. However, three plasmids
did not pass rotary validation, which may be due to unsuccessful
transformation of prey or bait plasmids into AH109 or weak
protein-protein interactions. Here, full-length of PHB was found to
interact with GlgA via co-immunoprecipitation. But unfortunately,
the part of PHB that interacts with GlgA was not determined in this
study, even though PHB1 and PHB2 belong to the PHB family. Thus,
subsequent studies will investigate the interacting part of GlgA to
further clarify the relation between C. trachomatis
infections and GlgA secretion into the host cell cytosol. In
summary, the yeast two-hybrid system was successfully exploited to
screen for GlgA interaction partners, indicating further regulatory
functions of GlgA.
Acknowledgements
Not applicable.
Funding
This project was supported by the Natural Science
Foundation of China (grant nos. 81471969, 81202374 and 31600150),
the construct program of Hunan Provincial Key Laboratory for
Special Pathogens Prevention and Control Foundation (grant no.
2014-5), and Hunan Province Cooperative Innovation Center for
Molecular Target New Drug Study (grant no. 2015-351).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZS, YS, YL, XL, HC, HW and BP performed the
experiments. ZS wrote the paper. CL designed the study, coordinated
the data analysis, reviewed and edited the manuscript. All authors
read and approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wang Y, Cutcliffe LT, Skilton RJ, Ramsey
KH, Thomson NR and Clarke IN: The genetic basis of plasmid tropism
between Chlamydia trachomatis and Chlamydia
muridarum. Pathog Dis. 72:19–23. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weber MM, Lam JL, Dooley CA, Noriea NF,
Hansen BT, Hoyt FH, Carmody AB, Sturdevant GL and Hackstadt T:
Absence of specific Chlamydia trachomatis inclusion membrane
proteins triggers premature inclusion membrane lysis and host cell
death. Cell Rep. 19:1406–1417. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Whitcher JP, Srinivasan M and Upadhyay MP:
Corneal blindness: A global perspective. Bull World Health Organ.
79:214–221. 2001.PubMed/NCBI
|
|
4
|
Brunham RC and Rey-Ladino J: Immunology of
Chlamydia infection: Implications for a Chlamydia
trachomatis vaccine. Nat Rev Immunol. 5:149–161. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yang C, Starr T, Song L, Carlson JH,
Sturdevant GL, Beare PA, Whitmire WM and Caldwell HD: Chlamydial
lytic exit from host cells is plasmid regulated. mBio. 6:e01648–15.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moulder JW: Interaction of chlamydiae and
host cells in vitro. Microbiol Rev. 55:143–190. 1991.PubMed/NCBI
|
|
7
|
Stephens RS, Myers G, Eppinger M and
Bavoil PM: Divergence without difference: Phylogenetics and
taxonomy of Chlamydia resolved. FEMS Immunol Med Microbiol.
55:115–119. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhong G: Chlamydial plasmid-dependent
pathogenicity. Trends Microbiol. 25:141–152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Patton MJ, McCorrister S, Grant C,
Westmacott G, Fariss R, Hu P, Zhao K, Blake M, Whitmire B, Yang C,
et al: Chlamydial protease-like activity factor and type III
secreted effectors cooperate in inhibition of p65 nuclear
translocation. mBio. 7(pii): e01427–16. 2016.PubMed/NCBI
|
|
10
|
Lu C, Lei L, Peng B, Tang L, Ding H, Gong
S, Li Z, Wu Y and Zhong G: Chlamydia trachomatis GlgA is
secreted into host cell cytoplasm. PLoS One. 8:e687642013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
O'Connell CM, Ingalls RR, Andrews CW Jr,
Scurlock AM and Darville T: Plasmid-deficient Chlamydia
muridarum fail to induce immune pathology and protect against
oviduct disease. J Immunol. 179:4027–4034. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ma RR, Sun J, Fang WH, Dong YP, Ruan JM,
Yang XL and Hu K: Identification of Carassius auratus
gibelio liver cell proteins interacting with the
GABAA receptor γ2 subunit using a yeast two-hybrid
system. Fish Physiol Biochem. 45:199–208. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fields S and Song O: A novel genetic
system to detect protein-protein interactions. Nature. 340:245–246.
1989. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nguyen PH, Lutter EI and Hackstadt T:
Chlamydia trachomatis inclusion membrane protein MrcA
interacts with the inositol 1,4,5-trisphosphate receptor type 3
(ITPR3) to regulate extrusion formation. PLoS Pathog.
14:e10069112018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee PY, Costumbrado J, Hsu CY and Kim YH:
Agarose gel electrophoresis for the separation of DNA fragments. J
Vis Exp:. (pii): 39232012.PubMed/NCBI
|
|
16
|
Gietz RD and Schiestl RH: High-efficiency
yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat
Protoc. 2:31–34. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhao X, Li P, An K, Jia X, Cheng Y and Jia
T: Chlamydia pneumoniae inclusion membrane protein Cpn0147
interacts with host protein CREB3. PLoS One. 12:e01855932017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Velmurugan R and Incharoensakdi A:
Disruption of polyhydroxybutyrate synthesis redirects carbon flow
towards glycogen synthesis in synechocystis sp. PCC 6803
overexpressing glgC/glgA. Plant Cell Physiol. 59:2020–2029. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Preiss J: Bacterial glycogen synthesis and
its regulation. Annu Rev Microbiol. 38:419–458. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Stephens RS, Kalman S, Lammel C, Fan J,
Marathe R, Aravind L, Mitchell W, Olinger L, Tatusov RL, Zhao Q, et
al: Genome sequence of an obligate intracellular pathogen of
humans: Chlamydia trachomatis. Science. 282:754–759. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Everett KD, Bush RM and Andersen AA:
Emended description of the order Chlamydiales, proposal of
Parachlamydiaceae fam. nov. and Simkaniaceae fam.
nov., each containing one monotypic genus, revised taxonomy of the
family Chlamydiaceae, including a new genus and five new species,
and standards for the identification of organisms. Int J Syst
Bacteriol. 49:415–440. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stynen B, Tournu H, Tavernier J and Van
Dijck P: Diversity in genetic in vivo methods for protein-protein
interaction studies: From the yeast two-hybrid system to the
mammalian split-luciferase system. Microbiol Mol Biol Rev.
76:331–382. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peterson TA, Stamnes MA and Piper RC: A
yeast 2-hybrid screen in batch to compare protein interactions. J
Vis Exp. 2018. View
Article : Google Scholar
|
|
24
|
Cao W, Guo J, Wen X, Miao L, Lin F, Xu G,
Ma R, Yin S, Hui Z, Chen T, et al: CXXC finger protein 1 is
critical for T-cell intrathymic development through regulating H3K4
trimethylation. Nat Commun. 7:116872016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wizel B, Nystrom-Asklin J, Cortes C and
Tvinnereim A: Role of CD8(+)T cells in the host response to
Chlamydia. Microbes Infect. 10:1420–1430. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang J, Li B and He QY: Significance of
prohibitin domain family in tumorigenesis and its implication in
cancer diagnosis and treatment. Cell Death Dis. 9:5802018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Guan X, Liu Z, Wang L, Johnson DG and Wei
Q: Identification of prohibitin and prohibiton as novel factors
binding to the p53 induced gene 3 (PIG3) promoter (TGYCC)(15)
motif. Biochem Biophys Res Commun. 443:1239–1244. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kuramori C, Azuma M, Kume K, Kaneko Y,
Inoue A, Yamaguchi Y, Kabe Y, Hosoya T, Kizaki M, Suematsu M and
Handa H: Capsaicin binds to prohibitin 2 and displaces it from the
mitochondria to the nucleus. Biochem Biophys Res Commun.
379:519–525. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Signorile A, Sgaramella G, Bellomo F and
De Rasmo D: Prohibitins: A critical role in mitochondrial functions
and implication in diseases. Cells. 8(pii): E712019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fischer SF, Vier J, Kirschnek S, Klos A,
Hess S, Ying S and Häcker G: Chlamydia inhibit host cell apoptosis
by degradation of proapoptotic BH3-only proteins. J Exp Med.
200:905–916. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yu Y: Prohibitin shuttles between
mitochondria and the nucleus to control genome stability during the
cell cycle. Plant Physiol. 179:1435–1436. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu FX, Johnston PA, Südhof TC and Yin HL:
gCap39, a calcium ion- and polyphosphoinositide-regulated actin
capping protein. Science. 250:1413–1415. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li T, Hong X, Zhao J, Teng Y, Zheng J,
Chen H, Chen H and Li H: Gelsolin-like actin-capping protein is
associated with patient prognosis, cellular apoptosis and
proliferation in prostate cancer. Biomark Med. 10:1251–1260. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Papala A, Sylvester M, Dyballa-Rukes N,
Metzger S and D'Haese J: Isolation and characterization of human
CapG expressed and post-translationally modified in Pichia
pastoris. Protein Expr Purif. 134:25–37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hubert T, Van Impe K, Vandekerckhove J and
Gettemans J: The actin-capping protein CapG localizes to
microtubule-dependent organelles during the cell cycle. Biochem
Biophys Res Commun. 380:166–170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hybiske K and Stephens RS: Mechanisms of
host cell exit by the intracellular bacterium Chlamydia.
Proc Natl Acad Sci USA. 104:11430–11435. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lutter EI, Barger AC, Nair V and Hackstadt
T: Chlamydia trachomatis inclusion membrane protein CT228
recruits elements of the myosin phosphatase pathway to regulate
release mechanisms. Cell Rep. 3:1921–1931. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
White CR, Datta G, Wilson L, Palgunachari
MN and Anantharamaiah GM: The apoA-I mimetic peptide 4F protects
apolipoprotein A-I from oxidative damage. Chem Phys Lipids.
219:28–35. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kingwell BA, Chapman MJ, Kontush A and
Miller NE: HDL-targeted therapies: Progress, failures and future.
Nat Rev Drug Discov. 13:445–464. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Rye KA and Barter PJ: Cardioprotective
functions of HDLs. J Lipid Res. 55:168–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Campbell LA and Kuo CC: Chlamydia
pneumoniae-an infectious risk factor for atherosclerosis? Nat
Rev Microbiol. 2:23–32. 2004. View Article : Google Scholar : PubMed/NCBI
|