Introduction
According to a recent report from the World Health
Organization, ischemic stroke is the second leading cause of
mortality worldwide (1). In 2016
alone, ~5.5 million people succumbed to an ischemic stroke and an
estimated 116.4 million people suffered from permanent disabilities
following a stroke event worldwide (2). Undoubtedly, these mortalities and
disabilities have a huge effect, both emotionally and financially,
on families and society.
Ischemic stroke is primarily caused by the blockage
of cerebrovascular blood flow. Previous studies have reported that
the incidence of early mortality following ischemic stroke (between
21 days to 1 month) in both high-income and low-middle-income
countries increased gradually from 2000 to 2008, from 13 to 23% and
13 to 19%, respectively (3,4).
Currently, intravenous thrombolysis and mechanical thrombectomy are
the only effective means of treating ischemic stroke (5,6);
however, these treatment options are limited by the
short-recommended treatment window (6). In addition, a series of reperfusion
injuries may occur following ischemic reperfusion (6,7); in
particular, inflammation and oxidative stress are the main causes
of reperfusion injuries, and the latter can induce the former
(6–9). An increasing amount of evidence has
demonstrated that continuous neuroinflammation can damage neurons
and the blood-brain barrier (BBB) during the process of cerebral
infarction, and this in turn results in tissue destruction and
worsens the functional outcome (10–12).
In addition, it has been demonstrated that the inhibition of
neuroinflammation decreased the neurological deficit in
experimental stroke models (12,13).
Therefore, it is evident that neuroinflammation serves a crucial
role in ischemic stroke-induced brain injury.
Microglia are important cells that initiate the
inflammatory response in the brain following various brain injuries
(14), including ischemic stroke
(15). Accumulating evidence has
indicated that activated microglia are the primary source of
immunomodulatory molecules, such as cytokines, chemokines and free
radicals, in the brain (16–18);
these molecules are closely associated with secondary brain injury
and brain tissue repair in ischemic stroke (16–18).
In addition, some molecules, including soluble Fas ligand (19) and endothelial nitric oxide synthase
(20,21), which are generated by other cells,
such as neurons and endothelial cells, have also been identified to
affect the stroke outcome through interacting with microglia in the
brain following ischemic stroke (19). Therefore, investigating microglial
changes and their function is crucial to understanding ischemic
stroke pathophysiology.
To develop new and effective therapies for ischemic
stroke, it is also necessary to investigate the cellular and
molecular mechanisms underlying brain damage and tissue repair in
ischemic stroke. Despite the fact that microglia-induced
inflammation can aggravate brain damage upon ischemic stroke, the
modulation of the microglial phenotype has also been observed to
promote brain repair; for example, through decreasing brain edema,
improving white matter integrity and recovering motor function
(13). Therefore, investigating
the mechanisms underlying microglial phenotypic transformation is
crucial for the development of novel treatments for patients with
ischemic stroke.
The present review aimed to describe the important
advances in the current understanding of the changes, functions and
modulators of microglia during ischemic stroke. The roles of
autophagy in the modulation of microglia polarization are
discussed, in addition to the future research directions of
ischemic stroke, which may help identify novel treatment strategies
for ischemic stroke.
Microglia
Microglia are small, macrophage-like glial cells
that account for 10–15% of cells in the central nervous system
(CNS). In response to various stresses, microglia are rapidly
activated to differentiate into either the M1 or the M2 phenotype,
which are involved in tissue damage and repair, respectively
(22–24).
M1 microglia and changes
M1 microglia exist in a proinflammatory state and
secrete proinflammatory cytokines, which have been identified to
promote brain damage (13,19). Following ischemic stroke, M1
microglia are activated and subsequently serve a detrimental role.
Briefly, upon experiencing ischemia/hypoxia, NF-κB is activated
within microglia and translocates from the cytoplasm to the
nucleus; this activates the release of proinflammatory cytokines,
which causes secondary brain damage (16–18),
such as interleukin (IL)-1β (24),
IL-6 (25) and tumor necrosis
factor-α (TNF-α) (26), in
addition to the production of inducible nitric oxide synthase
(iNOS) (27,28). For example, in middle cerebral
artery occlusion (MCAO) model mice, TNF-α secreted by M1 microglia
was identified to increase endothelial necroptosis and BBB leakage
following ischemic stroke (21,29).
This further promotes neuroinflammation and brain edema, resulting
in a poor outcome (29). Moraes
et al (30) demonstrated
that IL-1β, which is generated by M1 microglia, decreased the
number of synapses in the hippocampus and cortex, which aggravated
cognitive impairment. In oxygen-glucose-deprived (OGD) microglia,
the increase in IL-17A levels aggravated neuronal death (31). In addition, C-C motif chemokine
ligand 2 (CCL2) generated by microglia was observed to recruit
CD8+ γδ T cells to the ischemic brain, which was
identified to be the primary factor aggravating brain infarction
(32,33). In fact, besides ischemic stroke, M1
microglia activation occurs in multiple other neurological
diseases; for example, in an RD1 mouse model of retinal
degeneration, flow cytometry analysis revealed that the percentage
of T-lymphocyte activation antigen CD86 (CD86)+ M1
microglia was markedly increased in the rapid degeneration phase
(34); paraquat stimulation was
revealed to activate M1 microglia, which subsequently activated the
toll-like receptor-4 (TLR4)-mediated NF-κB signaling pathway and
resulted in the release of proinflammatory cytokines (35); and the number of low affinity
immunoglobulin gamma Fc region receptor III-A/b
(CD16/32)+ M1 microglia was significantly increased in
the brains of subarachnoid hemorrhage model mice through the
activation of the myeloid differentiation primary response protein
MyD88 (MyD88)/mitogen-activated protein kinase pathway, whereas
decreasing the percentage of M1 microglia significantly improved
the neurological deficits (36).
Other results from the literature have indicated that M1 microglia
activation can also aggravate brain damage in other types of
neurological disease (34–36).
The activation of M1 microglia can be determined by
detecting the expression of their surface markers (Table I); these classical markers include
integrin alpha-M (CD11b) (37),
CD16 (38), CD32 (39) and CD86 (40). However, these markers cannot
distinguish between microglia and macrophages in the brain, as they
can be expressed by both M1 microglia and macrophages. For example,
Liu et al (41) reported
that both CD16+/ionized calcium-binding adapter molecule
1 (Iba1)+ M1 microglia/macrophages were detected using
double immunofluorescence staining. To distinguish resident
microglia from blood-derived macrophages in the brain, Satoh et
al (42) suggested that
TMEM119 may be used as a marker of resting microglia in the human
brain; however, the challenge remains to discriminate activated
microglia from infiltrated macrophages in the brain.
 | Table I.Markers of M1 and M2 microglia. |
Table I.
Markers of M1 and M2 microglia.
| Phenotype | Marker | Type | Effect | (Refs.) | Author, year |
|---|
| M1 | IL-1β | Cytokine |
Proinflammatory | (25) | Zhu et al,
2019 |
|
| IL-6 | Cytokine |
Proinflammatory | (59) | Shu et al,
2016 |
|
| TNF-α | Cytokine |
Proinflammatory | (25) | Zhu et al,
2019 |
|
| iNOS | Metabolic
enzyme | Oxidative
damage | (59) | Shu et al,
2016 |
|
| ROS | Metabolite | Oxidative
damage | (7) | Xu et al,
2018 |
|
| CD11b | receptor | Induce
proinflammatory signaling | (37,81) | Liu et al,
2018; Hyakkoku et al, 2010 |
|
| CD16 | Receptor | Induce
proinflammatory signaling | (38) | Jiang et al,
2018 |
|
| CD32 | Receptor | Induce
proinflammatory signaling | (37) | Liu et al,
2018 |
|
| CD68 | Receptor | Induce
proinflammatory signaling | (27) | Boddaert et
al, 2018 |
|
| CD86 | Receptor | Induce
proinflammatory signaling | (44) | Hu et al,
2012 |
|
| NF-кB | Transcription | Induce M1
phenotype, elevate the expression of proinflammatory cytokines | (16) | Yang et al,
2019 |
|
| STAT1 | Transcription | Elevate the
expression of NF-κB | (70) | Butturini et
al, 2019 |
|
| STAT3 | Transcription | Elevate the
expression of NF-κB | (72) | Ding et al,
2019 |
|
| Hv1 | Ion channel
protein | Elevate the
expression of ROS and NF-κB, induce M1 phenotype | (94,96) | Tian et al,
2016; Yu et al, 2018 |
|
| Kv1.3 | Ion channel
protein | Proinflammatory,
induce M1 phenotype | (83,97) | Di Lucente et
al, 2018; Nguyen et al, 2017 |
|
| H19 | Gene | Proinflammatory,
induce M1 phenotype | (98) | Wang et al,
2017 |
|
| miRNA-155 | Gene | Proinflammatory,
induce M1 phenotype | (18) | Zheng et al,
2018 |
| M2 | IL-4 | Cytokine | Anti-inflammatory,
induce M2 phenotype | (23) | Liu et al,
2016 |
|
| IL-10 | Cytokine |
Anti-inflammatory | (25) | Zhu et al,
2019 |
|
| TGF-β | Cytokine | Anti-inflammatory,
regeneration | (44) | Hu et al,
2012 |
|
| Ym1/2 | Protein | Anti-inflammatory,
regeneration | (25) | Zhu et al,
2019 |
|
| Arg-1 | Protein | Anti-inflammatory,
regeneration | (44) | Hu et al,
2012 |
|
| CCL22 | Chemokine | Recruits regulatory
T cells | (44) | Hu et al,
2012 |
|
| CD206 | Receptor | Induce
anti-inflammatory signaling | (39) | Jin et al,
2014 |
|
| STAT6 | Transcription | Inhibit the
expression of NF-κB | (74) | Yang et al,
2017 |
|
| Nrf2 | Transcription | Anti-inflammatory,
antioxidant stress | (7) | Xu et al,
2018 |
|
| PPARγ | Transcription | Inhibit the
expression of NF-κB, promote the expression of Nrf2 | (26,78) | Liu et al,
2018; Cai et al, 2017 |
|
| miRNA-124 | Gene | Anti-inflammatory,
induce M2 phenotype | (99) | Hamzei et
al, 2016 |
|
| FAM19A3 | Gene | Anti-inflammatory,
induce M2 phenotype | (100) | Shao et al,
2015 |
Changes in the activation of M1 microglia can affect
stroke prognosis and changes in the expression levels of M1
microglia biomarkers, including surface markers and proinflammatory
cytokines, are also common in ischemic stroke (13,25,43).
Based on these changes, it is possible to draw some useful
conclusions regarding ischemic stroke. For example, in MCAO mice,
the expression levels of certain proinflammatory cytokines, such as
IL-1β, TNF-α and IL-6, in addition to iNOS, were observed to
increase from 12 h, peak at day 14, then subsequently decrease and
remain constant in the ischemic boundary zone until day 28
following MCAO (25,43). In addition, the study (25) also observed that CD32 expression
levels increased in a similar manner. Another previous study drew
similar conclusions, witnessing a significant increase in CD16
expression levels from day 3, which remained elevated until day 14
in the cortex and striatum at the inner boundary of the infarct
(44). These conclusions were
further confirmed in both a bilateral common carotid artery
ligation (BCCA) model and a photothrombotic (PT) stroke model. In
the hippocampal CA1 region of the BCCA ligation mice, the M1
microglia marker CD74 was observed to be expressed from day 3
post-ischemic insult and elevated from day 5 post-ischemia
(45). Additionally, CD16 and iNOS
expression levels were identified to be increased in infarct
regions at day 42 following BCCA (46). In PT stroke model mice, CD16 and
CD32 expression levels were detected at day 1 and remained elevated
until day 14 following PT cortical ischemia (47). In addition, numerous inflammatory
cytokines, such as TNF-α, IL-1β and IL-6, remained highly expressed
at days 14 and 21 in the ischemic region following a PT stroke
(48,49). Other associated studies reported
that age was also associated with microglia polarization, and the
trend of the expression levels of M1 microglial markers in ischemic
stroke was consistent with the former studies (25,44)
in both young adult and older mice (50,51).
Based on these changes, it is suggested that the
proinflammatory state may be dominant in the acute stage of
ischemic stroke; however, the changes in the expression levels of
proinflammatory cytokines during the chronic phase of ischemic
stroke, and the interactions between them and brain tissue repair,
remain unclear.
M2 microglia and changes
M2 microglia exist in an anti-inflammatory state and
secrete anti-inflammatory cytokines and neurotrophic factors to
promote brain repair (23,25,52).
M2 microglia are activated and reported to serve a beneficial role
following ischemic stroke; for example, in conditions of
ischemia/hypoxia, peroxisome proliferator-activated receptor γ
(PPARγ), a transcription factor with anti-inflammatory properties,
is reportedly activated and mobilized from the nucleus to the
cytoplasm in microglia (26). This
leads to the activation of M2 microglia, which release
anti-inflammatory cytokines that improve stroke outcome. For
example, Liu et al (23)
demonstrated that IL-4 secreted by M2 microglia decreased the
infarct size following ischemic stroke and improved the long-term
functional recovery. Zhu et al (25) observed that M2 microglia-induced
chitinase-3-like protein 3 (Ym1/2), IL-10 and transforming growth
factor-β (TGF-β) secretion promoted angiogenesis, and thereby
decreased the BBB leakage and improved stroke outcome. In addition,
Choi et al (53) confirmed
that M2 microglia promoted the proliferation and neuronal
differentiation of neural stem/progenitor cells in the ipsilateral
subventricular zone following ischemic stroke through upregulating
the expression levels of TGF-α; this may provide an effective
therapy for neurogenesis. Additionally, macrophages recruited by
microglia were identified to enhance M2 microglia polarization and
improve stroke outcome (54,55).
Similar to M1 microglia, besides from ischemic stroke, M2 microglia
activation occurs in numerous other types of neurological disease;
for example, in both spinal cord injury and intracerebral
hemorrhage mouse models, M2 microglia were identified to be
activated, which released anti-inflammatory cytokines downstream of
cAMP response element-binding protein (CREB)-associated signaling
pathways (56,57). In neonatal germinal matrix
hemorrhage rats, Rh-Chemerin promoted the accumulation and
proliferation of M2 microglia in periventricular regions and
suppressed the inflammatory response through nuclear factor
erythroid 2-related factor 2 (Nrf2)-associated signaling pathways
(58). Previous studies
demonstrated that M2 microglia activation can also promote brain
tissue repair in other neurological diseases (56–58).
The activation of M2 microglia can be assessed by
determining the expression levels of their surface markers
(Table I), for example, macrophage
mannose receptor 1 (CD206) (59),
and the secretion of anti-inflammatory cytokines, including IL-4
(23), IL-10 (59), arginase-1 (Arg-1) (60), Ym1 (28) and TGF-β (59). Changes in the percentage of
activated M2 microglia can affect stroke prognosis and based on
these changes, it is possible to draw some useful conclusions
regarding ischemic stroke; for example, in MCAO model mice
(44), the mRNA expression levels
of cytokines generated by M2 microglia, including Arg-1,
Ym1/2, CCL22, IL-10 and TGF-β, were detected from
days 1 to 3 following MCAO and peaked at days 3 to 5 post-injury in
ischemic regions (44). The
expression levels of the majority of M2 microglia genes was
observed to begin to decrease at days 5–7 after MCAO and by day 14,
were restored to pre-injury levels in ischemic regions (44). In addition, the number of
CD206+ cells followed a similar trend (44); CD206 remained highly expressed
until day 21 in the cortex and striatum following ischemia
(61). Previous studies have
further validated these results in a PT stroke model (47–49);
in the ischemic cortex, CD206 expression levels rapidly increased
at day 1, peaked at day 7 (47)
and remained highly expressed until day 21 following a PT stroke
(49). In addition, certain
anti-inflammatory cytokines were also observed to be highly
expressed until days 14 and 21 following a PT stroke; for example,
IL-4 and IL-10 remained highly expressed until day 14 (47) and TGF-β remained highly expressed
until day 21 in ischemic regions (49). Furthermore, another previous study
(44) revealed that the M2
phenotype was the dominant phenotype for the first 7 days following
ischemic stroke and could promote neuronal survival following
OGD.
Based on these data regarding the changes in the
expression levels of M1 and M2 markers, several conclusions were
drawn (Fig. 1); it was
hypothesized that microglia are activated following ischemic stroke
and the M1/M2-type pattern of expression changes dynamically
post-injury, exhibiting an early beneficial M2 phenotype, followed
by the transition to a detrimental M1 phenotype at the later
stages. These results suggested that regulating microglia
transformation from the M1 to M2 phenotype may be a crucial method
of treating ischemic stroke.
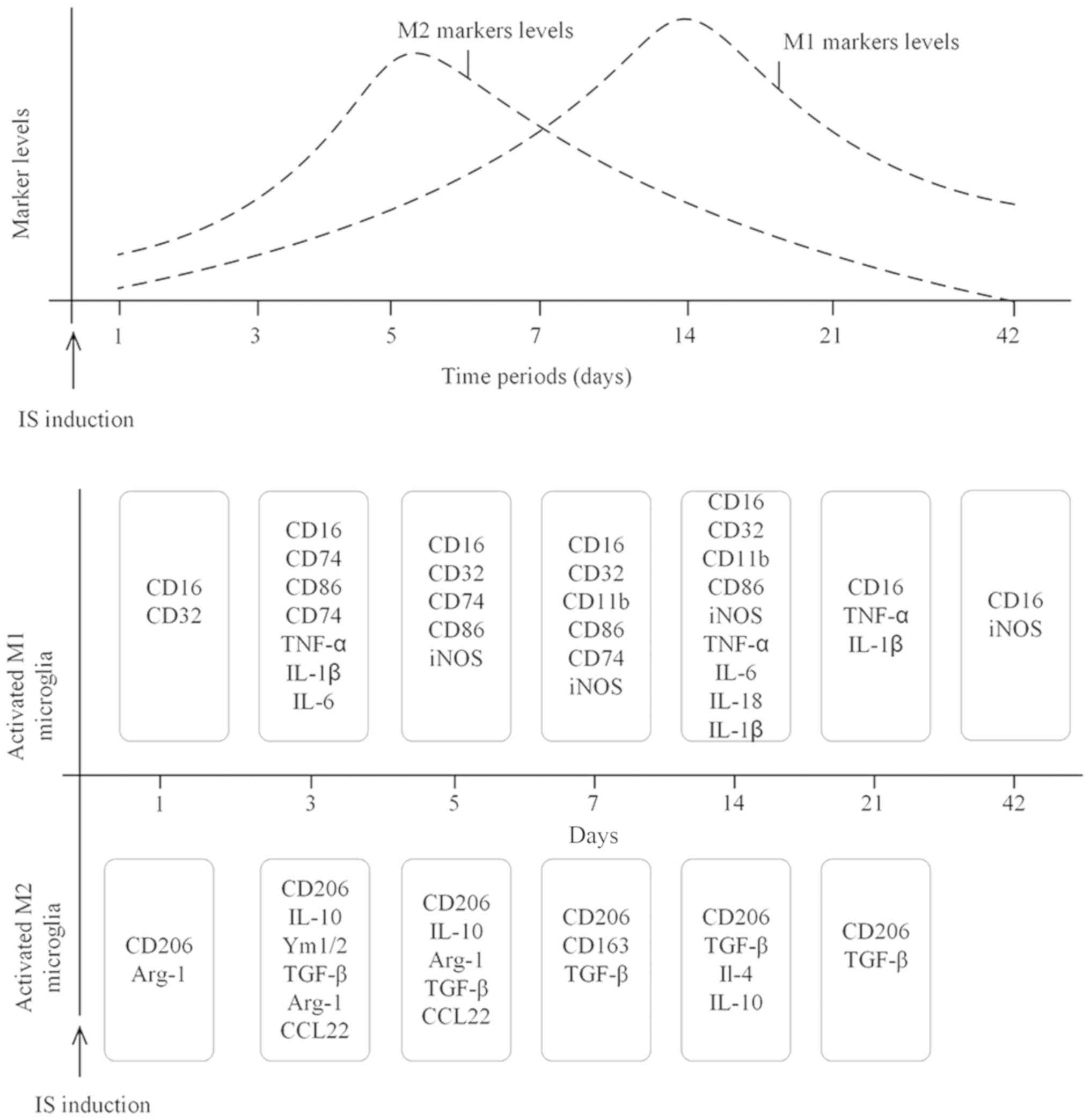 | Figure 1.Dynamic changes in microglia marker
levels over time following IS. Top panel: M1 microglia markers
demonstrated an increasing trend during the first 14 days following
IS, after which they decreased. Expression levels of M2 microglia
markers increased from day 1, peaked at days 5–7 and decreased
until day 42. Bottom panel: Microglia exhibited an M2-like response
as early as 1 day following IS, which manifested as increased
expression levels of CD206 Arg-1. The expression levels of TGF-β
increased from day 3 until day 21. With regards to the M1 markers,
the expression levels of the proinflammatory cytokines, TNF-α, IL-6
and IL-1β increased from day 3. The levels of M1 surface markers
CD16 and CD32 increased from day 1 and CD16 and iNOS expression
levels remained increased until day 42 after IS. IS, ischemic
stroke; Arg-1, arginase 1; TGF-β, transforming growth factor-β;
TNF-α, tumor necrosis factor α; IL, interleukin; iNOS, inducible
nitric oxide synthase; CCL, C-C motif chemokine ligand; Ym1/2,
chitinase-3-like protein 3. |
Modulatory mechanisms of microglia
polarization
It has been suggested that ischemic stroke may be
effectively treated through modulating microglia polarization; in
general, these modulators include four categories: Transcription
factors; receptors; ion channels; and gene modulators.
Transcription factors
Transcription factors are proteins that bind to DNA
and regulate the transcriptional activity of genes. NF-κB, members
of the STAT family, nuclear factor erythroid 2-related factor 2
(Nrf2) and PPARγ are 4 important transcription factors that have
been associated with the mechanisms underlying microglia
polarization (16,19,26,62,63)
(Fig. 2). The first 2
transcription factors are closely associated with M1 polarization
(16,19,62),
whereas the other 2 are associated with M2 polarization (26,63).
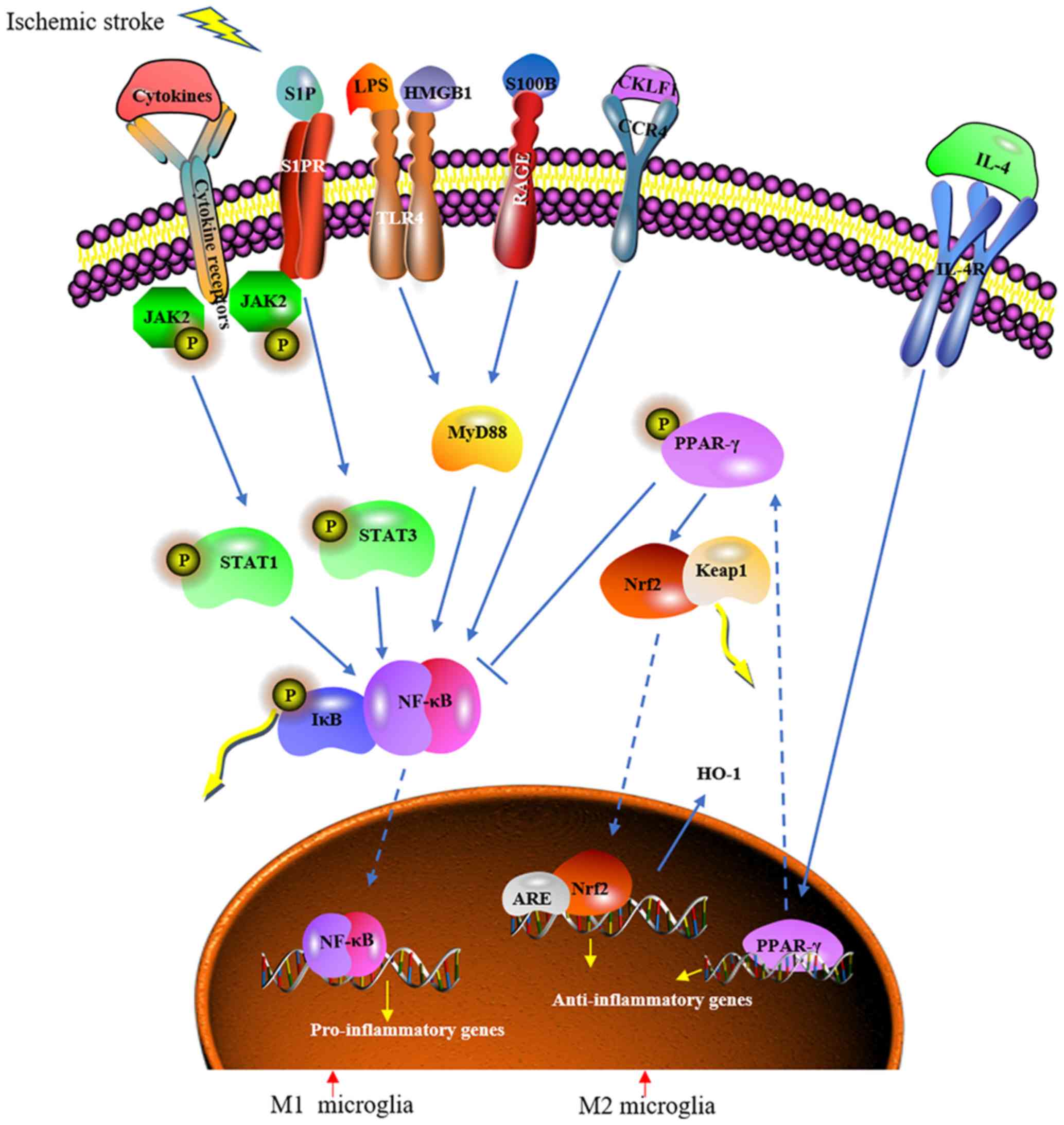 | Figure 2.Modulatory mechanisms of microglia
polarization following ischemic stroke. Microglia are activated
following ischemic stroke. M1 microglia activation involves
different factors, including the cytokines S1P, LPS, HMGB1, S100B
and CKLF1. These factors bind to membrane receptors, such as the
cytokine receptors S1PR, RAGE, TLR4, IL-4R and CCR4, to trigger
proinflammatory cellular signaling pathways. These pathways include
the JAK2/STAT1/NF-κB, JAK2/STAT3/NF-κB and MyD88/NF-κB pathways.
The downstream target NF-κB is released and translocates from the
cytoplasm to the nucleus following the phosphorylation of IκB,
where it initiates the transcription of proinflammatory genes. M2
microglia can be activated by IL-4, which promotes PPARγ
mobilization from the nucleus to the cytoplasm. Subsequently, PPARγ
inhibits the activation of NF-κB and promotes the activation of
Nrf2 to induce the transcription of anti-inflammatory genes.
Additionally, the activated Nrf2 promotes the expression of HO-1,
which scavenges reactive oxygen species and nitric oxide. S1P,
sphingosine 1 phosphate; LPS, lipopolysaccharide; IL, interleukin;
HMGB1, high mobility group protein B1; S100B, protein S100-B;
CKLF1, chemokine-like factor; SIPR, sphingosine 1 phosphate
receptor; TRL4, toll-like receptor 4; CCR4, C-C chemokine receptor
type 4; RAGE; JAK2, Janus activated kinase 2; MyD88, myeloid
differentiation primary response protein MyD88; IκB, NF-κB
inhibitor; PPARγ, peroxisome proliferator-activated receptor γ;
Nrf2, nuclear factor erythroid 2-related factor 2; HO-1, heme
oxygenase 1; Keap 1, Kelch-like ECH-associated protein 1; STAT,
signal transducer and activator of transcription. |
NF-κB
The transcription factor NF-κB has been identified
to activate microglia, and to transform these activated cells into
the M1 phenotype (16,62). In fact, it has been confirmed that
NF-κB can be activated in response to detrimental stresses and
induce inflammatory responses in the brain (16,62).
Upon ischemia or hypoxia, the expression levels of some
damage-associated molecules, including protein S100-B (11,64),
chemokine-like factor 1 (65–67)
and CXC motif ligand 12 (43) are
reportedly increased; these molecules bind to membrane receptors,
such as receptor for advanced glycation end products, C-C chemokine
receptor 4 and CXC-chemokine receptor 4 to activate NF-κB (43,64–66).
The activated NF-κB is subsequently translocated to the nucleus,
where it triggers inflammatory responses (16). NF-κB inhibitor α (IκBα), a protein
bound to NF-κB, can inhibit the activation of NF-κB; however, upon
phosphorylation, it can release NF-κB, which translocates to the
nucleus from the cytoplasm (16,68).
The activated NF-κB can promote the secretion of proinflammatory
cytokines from M1 microglia, such as IL-18, IL-6 and TNF-α
(16). For example, in MCAO model
mice, a significant increase in NF-κB (p65) expression levels were
detected in the ischemic brain, which was accompanied by an
increase in IL-18, IL-6 and TNF-α expression levels, whereas an
inhibitor of NF-κB significantly inhibited the release of these
cytokines (16). In addition,
inhibiting the increased upregulation of activated NF-κB (p65)
decreased the production of hypoxia-induced factor-1α, which then
attenuated the neuroinflammatory responses in ischemic stroke
(69).
STAT family members
STAT1-STAT6 exist in the cytoplasm and serve an
important role in the regulation of inflammatory responses. In
hypoxia-activated BV2 microglia, the phosphorylation levels of
STAT1 were significantly increased, which was accompanied by
increased expression levels of M1 microglia markers, such as
cluster of differentiation 68 and iNOS (70); however, the expression levels of
these markers were significantly decreased in STAT1−/−
BV2 cells following hypoxia. Several previous studies have also
identified that STAT3 is also associated with the regulation of M1
microglia polarization in both an MCAO-induced and a bilateral
common carotid arteries stenosis (BCAS)-induced model of ischemic
stroke (19,71–73).
Ding et al (72) identified
that the inhibition of the Janus kinase 2 (JAK2)/STAT3 signaling
pathway promoted the transition from resting microglia to the M2
phenotype to exert anti-inflammatory effects; briefly, the
phosphorylation levels of JAK2 and STAT3 were increased in the
acute phase of cerebral ischemia in the mice, which was accompanied
by a significant increase in M1 microglia markers, including CD16,
IL-1β and TNF-α, and a mild increase in M2 microglia markers, such
as CD206 and IL-10. However, SC99, an inhibitor targeting the
JAK2/STAT3 signaling pathway, inhibited the production of M1
markers and increased the expression of M2 markers, exerting
significant anti-inflammatory effects both in vivo and in
vitro. Similarly, another study demonstrated that AG490 (a
tyrosine kinase inhibitor of JAK2) decreased the expression levels
of M1 markers, iNOS, IL-1β, TNF-α, CD16 and CD32, the
phosphorylation of IκBα and translocation of p50/p65 in serum
soluble tumor necrosis factor ligand superfamily member 6 (Fas
ligand)-induced M1 microglia (19). Conversely, another previous study
demonstrated that the activated STAT3 and STAT6 pathways could
promote the polarization of nonactivated microglia to the M2
phenotype (74). Therefore, based
on these contradictory observations, further studies are required
to elucidate whether STATs and their associated signaling pathways
can regulate microglia polarization.
Nrf2
The redox-sensitive transcription factor Nrf2
demonstrates both antioxidant and anti-inflammatory properties
(75,76), and Nrf2 and its associated
signaling pathways have been identified to exert a protective
effect following ischemic stroke; for example, in
Nrf2−/− mice, the neurological deficit and
cerebral infarct volume were both aggravated following MCAO
compared with the wild-type mice (75). Meanwhile, the beneficial effects of
sirtuin 6, a member of the sirtuin family of
NAD+-dependent deacetylases, were abolished in
Nrf2−/− mice following ischemic stroke, in that
the reactive oxygen species (ROS) levels and the number of dead
neuro-2A cells in Nrf2−/− mice were increased
(75). In addition, another
previous study demonstrated that L-F001, a novel multifunctional
Rho-associated protein kinase inhibitor, increased the expression
levels of the M2 microglia marker CD206 through the activation of
Nrf2 signaling pathways in vitro (63). These beneficial results have also
been validated in vitro; in oxygen-glucose
deprivation/reperfusion (OGDR)-activated BV2 microglia, Nrf2 was
observed to be activated and dissociated from the repressor
protein, Kelch ECH-associating protein 1. This facilitated its
translocation from the cytoplasm to the nucleus, where it bound to
the antioxidant response element (ARE) and induced the expression
of downstream genes, such as NADPH quinone oxidoreductase and heme
oxygenase 1, which scavenge ROS and nitric oxide (7,68,77).
Additionally, the activation of Nrf2 was identified to decrease
ROS, NOD like receptor family pyrin domain containing 3 (NLRP3) and
IL-1β expression levels in BV2 microglia (7). However, despite the evident
protective role of Nrf2, further studies are required to elucidate
whether this transcription factor and its signaling pathways may
regulate M2 microglia polarization and function following ischemic
stroke.
PPARγ
PPARγ is a ligand-activated transcription factor
that is considered to be the primary mediator of inflammatory
responses. In acute ischemic stroke, PPARγ is reported to be
activated, which has been demonstrated to directly decrease tissue
damage through inhibiting the NF-κB pathway, decreasing
inflammation and stimulating the Nrf2/ARE axis to decrease
oxidative stress (78). A previous
study observed that 10-O-(N,N-dimethylaminoethyl)-ginkgolide B
methanesulfonate, a novel derivative of Ginkgolide B, could
transform polarized BV2 microglia from the M1 to the M2 phenotype
through promoting PPARγ translocation from the nucleus to the
cytoplasm following OGD; this subsequently promoted the decreased
expression levels of TNF-α and the increased expression levels of
TGF-β1 (26). Additionally, IL-4
generated by neurons was identified to bind to the IL-4 receptor
expressed on the surface of microglia and activate M2 microglia
through regulating the PPARγ signaling pathway (52); the activated M2 microglia were
observed to phagocytose damaged cells and secrete brain-derived
neurotrophic factor (BDNF) to decrease ischemic brain damage
(52). Based on these studies, it
was suggested that the modulation of PPARγ and its associated
signaling pathways may represent a potential target for the
treatment of ischemic stroke.
Receptors
Receptors are proteins, usually cell surface
receptors, that recognize and bind to ligands and serve an
important role in the signaling pathways involved in inducing local
inflammation, the recruitment of new effector cells, the
containment of local infection and the initiation of an adaptive
immune response. Previous studies have identified that 3 important
membrane receptors, TLR4, sphingosine 1 phosphate receptors (S1PRs)
and thromboxane A2 receptor (TXA2R), are closely associated with
microglia polarization (31,70,79,80).
TLR4
TLR4, which is predominantly expressed by microglia,
is an important regulator of inflammatory responses (79). Increased protein expression levels
of TLR4 are reportedly associated with poor outcomes in patients
with ischemic stroke (81).
Furthermore, in ischemic stroke, TLR4 has been observed to
recognize damage-associated molecules, such as lipopolysaccharide
(LPS) and high mobility group box-1, and trigger inflammatory
responses (Fig. 2) (82,83).
In mice, the number of TLR4+ cells, as well as
NF-κB+ and IL-1β+ cells, was significantly
increased at 72 h following MCAO; the number of TLR4+
cells was identified to be closely positively correlated with the
number of IL-1β+ cells and the infarct volume (84). In addition, in
TLR4−/− mice, the infarct volume was decreased
compared with TLR4+/+ mice following MCAO (85). These results have also been
validated in vitro; in BV2 microglia, a significant increase
in TLR4 expression levels were detected following OGD, which was
accompanied by an increase in proinflammatory cytokines, including
TNF-α, IL-1β and IL-6 (86);
however, in TLR4−/− mice, expression levels of
these cytokines generated by M1 microglia were observed to be
significantly decreased through the inhibition of the MyD88/NF-κB
signaling pathway (79,87). These results suggested that the
overexpression of TLR4 may activate and promote M1 microglia to
trigger neuroinflammatory responses in ischemic stroke, which
subsequently aggravates the brain damage. However, in another
study, TLR4 was demonstrated to be positively associated with brain
tissue repair (85): TLR4
activation regulated cell proliferation and neuroblast migration in
the subventricular zone on days 1, 7 and 14 following MCAO, which
suggested an effective means to modulate neurogenesis following
ischemic stroke. Based on this conflicting evidence, further
studies on the effects of TLR4 in different cells and at different
time points in ischemic stroke are required.
S1PRs
S1PR1-S1PR5 are G-protein-coupled receptors that are
expressed in abundance in the microglia and are demonstrated to
regulate inflammatory responses following ischemic stroke (88). S1P is the ligand for S1PR and has
been identified to bind to S1PR1, S1PR2 and S1PR3 to trigger
neuroinflammatory responses in ischemic stroke (Fig. 2) (89–91).
An in vitro study reported that upon the addition of S1P to
the culture medium of primary microglia, the expression levels of
IL-17 were increased (31).
Additionally, the exogenous administration of S1P to OGDR microglia
aggravated OGDR-induced neuronal apoptosis (31). In MCAO model mice, sphingosine
kinase 1 (Sphk1) was demonstrated to phosphorylate sphingosine to
S1P, and then S1P bound to S1PR3 to activate M1 microglia, which
released proinflammatory cytokines and aggravated brain damage.
This culminated in an increase in the expression levels of S1P and
M1 microglial markers, CD11b, CD16, CD32, CD86, TNF-α and IL-1β, as
well as an increased infarct volume and neurological deficit
(81). CAY10444, an inhibitor of
S1PR3, and PF-543, an inhibitor of Sphk1, were identified to
partially reverse the result to increase the expression levels
(31,91); however, Qin et al (71) described opposing results, following
the observation that Fingolimod, an S1PR agonist, inhibited M1
microglia-induced neuroinflammation and shift microglia toward M2
polarization through regulating STAT3 signaling in a BCAS-induced
model of ischemic stroke. Therefore, additional studies are
required to determine the role of S1P and S1PRs in the regulation
of microglia polarization following ischemic stroke.
TXA2R
TXA2R, another G-protein-coupled receptor, can
promote platelet activation and aggregation, which regulates
thrombosis/hemostasis and inflammatory responses (92,93).
In ischemia/reperfusion mice, significant TXA2R expression levels
were detected in microglia/macrophages by immunofluorescence double
staining (80). In addition, the
expression levels of M1 microglia markers, such as CD16, CD86,
IL-1β, IL-6 and TNF-α, were significantly increased in ischemic
brains (80). However, SQ29548, a
TXA2R antagonist, inhibited the activation of M1 microglia and
subsequent inflammatory responses, which culminated in a decrease
in CD16, CD86, IL-1β, IL-6 and TNF-α expression levels.
Ion channels
The expression of ion channels changes in response
to voltage and pH gradients in the microenvironment, which induces
intracellular signal transduction. Currently, 2 important ion
channels, Hv1 and Kv1.3, have been reported to be closely
associated with microglia polarization (83,94).
Hv1
Hv1 is a voltage-gated proton channel that enables
NADPH oxidase (NOX) to produce ROS by sensing both voltage and pH
gradients (95). ROS have been
identified to be one of the main factors contributing to the
induction of M1 microglia to release proinflammatory cytokines
(96). For example, a previous
study demonstrated that an increase in Hv1 expression levels
induced neuroinflammatory responses through the Hv1/NOX/ROS
pathway, thereby exacerbating brain damage in ischemic stroke
(95). Furthermore, in both an
MCAO-induced and a PT-induced model of ischemic stroke, large
Hv1-mediated currents were detected in microglia, but not in
neurons or astrocytes (94,95);
this was accompanied by increased expression levels of ROS in the
brain, which activated its downstream target NF-κB to trigger the
secretion of proinflammatory cytokines generated by M1 microglia,
further aggravating brain damage following ischemic stroke. In
Hv1−/− mice, the brain infarct volume, neuronal
damage and motor deficits were attenuated compared with wild-type
mice following PT induction, which was accompanied by decreased
expression levels of the M1 microglia markers, iNOS and CD16
(94). Additionally, the study
also confirmed that more ROS could be released by M1 microglia
following ischemic stroke, and in turn, these increased levels of
ROS promoted M1 microglia polarization, resulting in a vicious
circle of neuroinflammation, which exacerbated neuronal death.
Another study demonstrated that the regenerative rate of
oligodendrocyte precursor cells was increased, and subsequently the
periventricular leukomalacia was reduced in
Hv1−/− mice, confirming that Hv1 may aggravate
brain damage (96). These results
indicated that Hv1 may aggravate brain damage through increasing
the expression levels of ROS and proinflammatory cytokines
generated by M1 microglia; however, whether Hv1 affects M2
microglia polarization remains unclear.
Kv1.3
Previous studies have identified that changes in the
expression levels of K+ channels also occur in ischemic
stroke and this can regulate microglia polarization (83,97).
K+ channels are membrane proteins that permit the rapid
and selective flow of K+ ions across the cell membrane,
thus generating electrical signals in cells. In response to brain
ischemia, the expression levels of Kv1.3 were observed to be
increased in microglia of both rodents and humans, which modulated
Ca2+ signaling and induced neuroinflammation (97). In addition, in an LPS-induced
microglia activated mouse model, M1 microglia were activated,
accompanied by an increase in the current density and mRNA
expression levels of Kv1.3 and inflammatory cytokines, such
as TNF-α, IL-1β and IL-6, in addition to iNOS (83). Notably, Kv1.3 knockout or
treatment with the Kv1.3 inhibitor 5-(4-phenoxybutoxy)psoralen
(PAP-1) could partially reverse these results; in LPS-activated
microglia the expression levels of Kv1.3 and M1 microglia markers,
such as TNF-α, IL-1β and nitric oxide were significantly increased;
however, PAP-1 was observed to decrease the polarization of M1
microglia, as well as the expression levels of proinflammatory
cytokines (83). These data
suggested that Kv1.3 may be one of the main mediators of M1
microglia polarization.
Modulators of gene expression
H19 imprinted maternally expressed transcript
(H19), microRNA (miRNA)−155, miRNA-124 and
chemokine-like protein TAFA-3 (FAM19A3) have been reported
to be closely associated with inflammatory responses; the first 2
factors are closely associated with M1 polarization, whereas the
latter 2 factors are associated with M2 polarization (18,98–100).
H19
H19 is a long non-coding RNA (lncRNA) that
has been demonstrated to induce the onset of ischemic stroke and
participate in the chronic regeneration process following ischemic
stroke (98,101–103). In fact, H19 has been
demonstrated to promote M1 microglia polarization and induce
neuroinflammation in ischemic stroke through regulating histone
deacetylases (HDACs) (98,104,105). In clinical experiments (98,101), H19 levels were identified
to be significantly increased in the plasma at 3 h, 7, 30 and 90
days post-ischemic stroke, in addition to being positively
correlated with the National Institute of Health Stroke Scale score
(106) and TNF-α expression
levels. Furthermore, in MCAO model mice, H19 expression
levels were increased in both the plasma and the brain (98). Notably, H19 knockdown
decreased the infarct volume and brain edema and decreased TNF-α
and IL-1β expression levels in the brain following ischemic stroke
(98). Furthermore, the study also
demonstrated that H19 knockdown could inhibit OGD-induced M1
microglia polarization in BV2 cells, which manifested as a decrease
in the expression levels of TNF-α and CD11b (98). In addition, H19 knockdown
reversed the OGD-induced upregulation of HDAC1, whereas HDAC1
overexpression reversed the effects of H19 knockdown and
promoted microglia polarization toward the M1 phenotype (98). These data could be used to regulate
M1 microglia polarization through the H19/HDAC1 signaling
pathway.
miRNA-155
Increased expression levels of miRNA-155
have been demonstrated to be associated with the neuroinflammatory
responses (20,107). In LPS-activated microglia,
miRNA-155 expression levels were significantly increased,
which could target suppressor of cytokine signaling 1 to trigger M1
microglia-mediated inflammatory responses and aggravate brain
damage (18). In addition,
miRNA-155−/− mice were reported to partially
reverse LPS-induced neurological deficits and microglia activation
(107); however, the roles of
miRNA-155 in microglia polarization in ischemic stroke
require additional clarification.
miRNA-124
miRNA-124 is the most abundant miRNA present
in the CNS and is involved extensively in neuroprotective
mechanisms (108,109). In fact, it has been identified to
regulate microglia activation and polarization in ischemic stroke
(99,110). A previous study demonstrated that
miRNA-124 induced neuroprotection and functional improvement
through regulating M2 microglia polarization in ischemic stroke
(99); upon injection of
miRNA-124 into the mouse ipsilateral hemisphere, the number
of M2 microglia/macrophages was markedly increased, contributing to
neuronal survival by releasing the trophic factor Arg-1 following
MCAO. miRNA-124 treatment also resulted in the accelerated
infiltration of reactive astrocytes into the lesion area, which
subsequently resulted in a tighter glial scar border; however,
whether the activation and polarization of microglia affects
astrocyte function through modulating miRNA-124 remains
unclear.
FAM19A3
FAM19A3, a member of the TAFA family of
genes (TAFA1-5), is predominantly expressed in the CNS
(111). Shao et al
(100) suggested that
FAM19A3 expression levels were significantly increased in
the microglia of MCAO model mice, which promoted M2 microglia
polarization and ameliorated cerebral ischemia. Notably, within
these mice, FAM19A3 expression levels in the brain were
elevated 1–3 days following MCAO, peaked at day 5, began to
decrease at day 7 and returned to pre-injury levels by day 14,
which is consistent with the pattern of M2 microglia markers,
CD206, IL-10, Ym1/2 and TGF-β discussed previously. Additionally,
FAM19A3 was observed to be secreted by M2 microglia and in turn,
FAM19A3 increased the expression levels of M2 microglia markers in
a dose-dependent manner (100).
These results suggested that FAM19A3 may be a genetic marker
for M2 microglia; however, the underlying molecular mechanisms
remains to be investigated.
Drug modulators
In clinical and experimental studies, a large
number of drugs have been demonstrated to serve important roles as
modulators of neuroinflammation and microglia polarization; for
example, in both a PT-induced stroke model and BCAS-induced model,
Fingolimod, a United States of America Food and Drug
Administration-approved drug for the treatment of
relapsing-remitting multiple sclerosis, modulated microglia toward
M2 polarization, protected against ischemic damage and promoted
angiogenesis through suppressing the STAT3 signaling pathway
(47,71). In MCAO model mice, rosiglitazone
decreased the numbers of Iba1+/CD16+ M1
microglia and increased the numbers of
Iba1+/CD206+ M2 microglia following stroke,
which improved the overall long-term white matter integrity
(61,112). In addition to western medicines,
some traditional Chinese medicines can also be used to regulate
microglia polarization (16,25,113); a study by Yang et al
(16) demonstrated that baicalein
significantly decreased the expression levels of the M1 markers,
CD16 and CD86, and increased the expression levels of the M2
markers, CD163 and CD206. Additionally, expression levels of the
proinflammatory factors, IL-6, IL-18 and TNF-α were decreased upon
inhibition of NF-κB signaling following baicalein treatment in MCAO
model mice (16). Zhu et al
(25) demonstrated that berberine
could inhibit M1 polarization and promote M2 polarization to
protect against ischemic stroke through the AMPK signaling pathway.
Wang et al (46) reported
that xinnao shutong capsules, a Chinese medicine produced in China,
decreased the number of M1 microglia and increased the number of M2
microglia, which improved memory impairments in a BCCA-induced
model of ischemic stroke (46).
Nonetheless, despite the effects of these drugs on microglia
polarization being confirmed in experimental studies, clinical
studies are still lacking.
Autophagy
Autophagy is a cellular metabolic pathway through
which damaged organelles and misfolded proteins are degraded and
recycled to maintain cellular homeostasis. Accumulating evidence
has reported that autophagy is activated in brain cells such as
neurons, endothelial cells and microglia in ischemic stroke, and
interfering with autophagy can aggravate brain damage (114–116). For example, in an in vitro
study, the inhibition of autophagic flux with NH4Cl was
demonstrated to aggravate the damage to brain cells, as it
stimulated M1 microglia polarization and the release of TNF-α, iNOS
and cyclooxygenase-2 (117). In
addition, the expression levels of M2 markers, including IL-10,
Arg-1 and BDNF were decreased following the treatment of
NH4Cl in OGD microglia. mTOR, a 289-kDa serine/threonine
protein kinase, was demonstrated to inhibit autophagy through
regulating its downstream signaling pathways (118), whereas Li et al (119) observed that mTOR complex 1
(mTORC1), a form of mTOR, prompted microglia to shift towards the
M1 phenotype. Moreover, the inhibition of mTORC1 decreased the
infarct volume and improved motor function, which lead to a
decrease in the production of proinflammatory cytokines, including
iNOS, TNF-α and IL-1β, and chemokines, C-C motif chemokine 2 and
C-C motif chemokine ligand 22, by M1 microglia. However, another
previous study proposed the opposite view and suggested that
autophagy may stimulate a microglial shift toward the M1 phenotype
to exacerbate cerebral ischemia (120). In an in vitro study, the
expression levels of the autophagy markers microtubule-associated
proteins 1A/1B light chain 3B lipid modified form, beclin-1 and
autophagy protein 5 were increased, which was accompanied by
increased expression levels of the proinflammatory factors IL-1β,
TNF-α and iNOS in primary microglia under OGD (120); however, these proinflammatory
factors and the number of M1 microglia were downregulated by the
autophagy inhibitor 3-methyladenine (3-MA). Moreover, 3-MA also
prevented M2 microglia polarization through inhibiting the
activation of CREB. Thus, based on these contradictory results, the
roles of autophagy in microglia polarization in ischemic stroke
require further investigation.
Conclusions
Microglia activation and polarization is considered
to be important in brain damage and repair following ischemic
stroke. The present review provided an overview of the activation
and changes of M1 and M2 microglia following ischemic stroke. This
is useful for determining the optimal time point at which to target
microglia to treat ischemic stroke. Additionally, studies
investigating the modulators of microglia activation and
polarization were analyzed, as well as the role of autophagy in
microglia polarization in ischemic stroke. This is of importance as
it provides therapeutic targets to target microglia in ischemic
stroke and it is also critical for our understanding of the
pathophysiology and future investigations of ischemic stroke. It is
worth noting that according to the American Heart
Association/American Stroke Association guidelines intracerebral
hemorrhage and ischemic stroke are 2 different types of stroke with
different pathogenesis and treatments (106,121). Lan et al (122) have already reviewed the
modulation of microglial activation and polarization in
intracerebral hemorrhage from 3 aspects: Microglial activation and
function in intracerebral hemorrhage; modulators of microglial
polarization; and microglial interactions with other cells.
However, the present review differs from the review by Lan et
al (122), as the present
study investigated the changes and modulation of microglia in
ischemic stroke. Additionally, Lan et al (122) primarily focused on the modulation
of microglial polarization by the transcription factors and
receptors. In addition to these typical modulation factors, which
also serve a similar role in the modulation of microglia in
ischemic stroke, the present review also discussed other factors
associated with the modulation of microglial polarization, such as
ion channels, genes and drugs. Notably, the time and spatial
changes of microglia in ischemic stroke were further examined in
this review. In practice, this review suggested future research
directions to provide novel strategies for the treatment of
ischemic stroke.
Based on the data presented in the present review,
several issues can be selected for further investigation. Firstly,
the experimental models of ischemic stroke need to be improved and
more data on the experimental models needs to be collected and
collated, to decrease the discrepancies reported between studies.
Secondly, beneficial factors identified that regulated the
polarization of microglia towards the M2 phenotype in animals with
ischemic stroke require further investigations. Thirdly, further
investigations are required into the role of autophagy in microglia
polarization to improve our understanding of the pathology of
ischemic stroke. In addition, regarding the extensive roles of
microglia in ischemic stroke, investigations into the interaction
between microglia and other cells will be important to identify the
effective pathophysiological pathways that prevent brain damage.
Finally, the focus of this review was on the pathophysiological
regulatory mechanisms of microglia polarization. Therefore, the
range of potential drug targets has not been fully explored. Our
future studies will focus on other aspects of the roles of
microglia in ischemia.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant nos. 81874416 and 81774174), the
Hunan Education Department's Science & Research Project (grant
no. 16K063) and the Department of Integrated Traditional Chinese
and Western Medicine, Hunan University of Traditional Chinese
Medicine (grant no. 2018ZXYJH01).
Availability of data and materials
Not applicable.
Authors' contributions
CTJ collected the literature, performed the
analysis and interpretation and wrote the manuscript. WFW, YHD and
JWG contributed to literature collection, and analysis and
interpretation of the results. CTJ, YHD and JWG revised the
manuscript. All authors reviewed the final draft of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
World Health Organization (WHO), . Top 10
global causes of deaths, 2016. https://www.who.int/en/news-room/fact-sheets/detail/the-top-10-causes-of-deathWHO;
Geneva: 2016 May 24–2018
|
|
2
|
GBD 2016 Stroke Collaborators, . Global,
regional, and national burden of stroke, 1990–2016: A systematic
analysis for the Global Burden of Disease Study 2016. Lancet
Neurol. 18:439–458. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Feigin VL, Lawes CMM, Bennett DA,
Barker-Collo SL and Parag V: Worldwide stroke incidence and early
case fatality reported in 56 population-based studies: A systematic
review. Lancet Neurol. 8:355–369. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Navis A, Garcia-Santibanez R and Skliut M:
Epidemiology and outcomes of ischemic stroke and transient ischemic
attack in the adult and geriatric population. J Stroke Cerebrovasc
Dis. 28:84–89. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ozaki T, Nakamura H and Kishima H:
Therapeutic strategy against ischemic stroke with the concept of
neurovascular unit. Neurochem Int. 126:246–251. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thiebaut AM, Gauberti M, Ali C, Martinez
De Lizarrondo S, Vivien D, Yepes M and Roussel BD: The role of
plasminogen activators in stroke treatment: Fibrinolysis and
beyond. Lancet Neurol. 17:1121–1132. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu XJ, Zhang L, Ye XC, Hao Q, Zhang T, Cui
GY and Yu M: Nrf2/ARE pathway inhibits ROS-induced NLRP3
inflammasome activation in BV2 cells after cerebral ischemia
reperfusion. Inflamm Res. 67:57–65. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Azedi F, Mehrpour M, Talebi S, Zendedel A,
Kazemnejad S, Mousavizadeh K, Beyer C, Zarnani A and Joghataei MT:
Melatonin regulates neuroinflammation ischemic stroke damage
through interactions with microglia in reperfusion phase. Brain
Res. 1723:1464012019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhuang P, Wan Y, Geng S, He Y, Feng B, Ye
Z, Zhou D, Li D, Wei H, Li H, et al: Salvianolic Acids for
Injection (SAFI) suppresses inflammatory responses in activated
microglia to attenuate brain damage in focal cerebral ischemia. J
Ethnopharmacol. 198:194–204. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chamorro Á, Dirnagl U, Urra X and Planas
AM: Neuroprotection in acute stroke: Targeting excitotoxicity,
oxidative and nitrosative stress, and inflammation. Lancet Neurol.
15:869–881. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou SJ, Zhu WQ, Zhang Y, Pan SP and Bao
JH: S100B promotes microglia M1 polarization and migration to
aggravate cerebral ischemia. Inflamm Res. 67:937–949. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao HR, Wan LH, Chen Y, Zhang H, Xu Y and
Qiu SW: FasL incapacitation alleviates CD4(+) T cells-induced brain
injury through remodeling of microglia polarization in mouse
ischemic stroke. J Neuroimmunol. 318:36–44. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xiong XY, Liu L and Yang QW: Functions and
mechanisms of microglia/macrophages in neuroinflammation and
neurogenesis after stroke. Prog Neurobiol. 142:23–44. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Neher JJ and Cunningham C: Priming
microglia for innate immune memory in the brain. Trends Immunol.
40:358–374. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li B, Concepcion K, Meng XM and Zhang LB:
Brain-immune interactions in perinatal hypoxic-ischemic brain
injury. Prog Neurobiol. 159:50–68. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang S, Wang H, Yang Y, Wang R, Wang Y, Wu
C and Du G: Baicalein administered in the subacute phase
ameliorates ischemia-reperfusion-induced brain injury by reducing
neuroinflammation and neuronal damage. Biomed Pharmacother.
117:1091022019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Weng L, Wu Z, Zheng W, Meng H, Han L, Wang
S, Yuan Z and Xu Y: Malibatol A enhances alternative activation of
microglia by inhibiting phosphorylation of Mammalian Ste20-like
kinase1 in OGD-BV-2 cells. Neurol Res. 38:342–348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zheng X, Huang H, Liu J, Li M, Liu M and
Luo T: Propofol attenuates inflammatory response in LPS-activated
microglia by regulating the miR-155/SOCS1 pathway. Inflammation.
41:11–19. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Meng HL, Li XX, Chen YT, Yu LJ, Zhang H,
Lao JM, Zhang X and Xu Y: Neuronal soluble Fas ligand drives
M1-microglia polarization after cerebral ischemia. CNS Neurosci
Ther. 22:771–781. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Greco R, Demartini C, Zanaboni AM,
Blandini F, Amantea D and Tassorelli C: Endothelial nitric oxide
synthase inhibition triggers inflammatory responses in the brain of
male rats exposed to ischemia-reperfusion injury. J Neurosci Res.
96:151–159. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ji J, Xiang P, Li T, Lan L, Xu X, Lu G, Ji
H, Zhang Y and Li Y: NOSH-NBP, a novel nitric oxide and hydrogen
sulfide- releasing hybrid, attenuates ischemic stroke-induced
neuroinflammatory injury by modulating microglia polarization.
Front Cell Neurosci. 11:1542017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang J, Xing H, Wan L, Jiang X, Wang C and
Wu Y: Treatment targets for M2 microglia polarization in ischemic
stroke. Biomed Pharmacother. 105:518–525. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu X, Liu J, Zhao S, Zhang H, Cai W, Cai
M, Ji X, Leak R, Gao Y, Chen J and Hu X: Interleukin-4 is essential
for microglia/macrophage M2 polarization and long-term recovery
after cerebral ischemia. Stroke. 47:498–504. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Feng YK, He XF, Luo SJ, Chen XF, Long SM,
Liang FY, Shi TT, Pei Z and Li ZD: Chronic colitis induces meninges
traffic of gut-derived T cells, unbalances M1 and M2
microglia/macrophage and increases ischemic brain injury in mice.
Brain Res. 1707:8–17. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu J, Cao D, Guo C, Liu M, Tao Y, Zhou J,
Wang F, Zhao Y, Wei J, Zhang Y, et al: Berberine facilitates
angiogenesis against ischemic stroke through modulating microglial
polarization via AMPK signaling. Cell Mol Neurobiol. 39:751–768.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu R, Diao J, He S, Li B, Fei Y, Li Y and
Fang W: XQ-1H protects against ischemic stroke by regulating
microglia polarization through PPARgamma pathway in mice. Int
Immunopharmacol. 57:72–81. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Boddaert J, Bielen K, 's Jongers B,
Manocha E, Yperzeele L, Cras P, Pirici D and Kumar-Singh S: CD8
signaling in microglia/macrophage M1 polarization in a rat model of
cerebral ischemia. PLoS One. 13:e01869372018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Collmann FM, Pijnenburg R, Hamzei-Taj S,
Minassian A, Folz-Donahue K, Kukat C, Aswendt M and Hoehn M:
Individual in vivo profiles of microglia polarization after stroke,
represented by the genes iNOS and Ym1. Front Immunol. 10:12362019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen AQ, Fang Z, Chen XL, Yang S, Zhou YF,
Mao L, Xia YP, Jin HJ, Li YN, You MF, et al: Microglia-derived
TNF-alpha mediates endothelial necroptosis aggravating blood
brain-barrier disruption after ischemic stroke. Cell Death Dis.
10:4872019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moraes CA, Santos G, de Sampaio e Spohr
TC, D'Avila JC, Lima FR, Benjamim CF, Bozza FA and Gomes FC:
Activated microglia-induced deficits in excitatory synapses through
IL-1β: Implications for cognitive impairment in sepsis. Mol
Neurobiol. 52:653–663. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lv M, Zhang D, Dai D, Zhang W and Zhang L:
Sphingosine kinase 1/sphingosine-1-phosphate regulates the
expression of interleukin-17A in activated microglia in cerebral
ischemia/reperfusion. Inflamm Res. 65:551–562. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xie L, Sun F, Wang J, Mao X, Xie L, Yang
S, Su D, Simpkins J, Greenberg D and Jin K: mTOR signaling
inhibition modulates macrophage/microglia-mediated
neuroinflammation and secondary injury via regulatory T cells after
focal ischemia. J Immunol. 192:6009–6019. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang SW, Zhang H and Xu Y: Crosstalk
between microglia and T cells contributes to brain damage and
recovery after ischemic stroke. Neurol Res. 38:495–503. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhou T, Huang Z, Sun X, Zhu X, Zhou L, Li
M, Cheng B, Liu X and He C: Microglia polarization with M1/M2
phenotype changes in rd1 mouse model of retinal degeneration. Front
Neuroanat. 11:772017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang M, Li Y, Wu K, Yan W, Tian T, Wang Y
and Yang H: Paraquat modulates microglia M1/M2 polarization via
activation of TLR4-mediated NF-κB signaling pathway. Chem Biol
Interact. 310:1087432019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Huang XP, Peng JH, Pang JW, Tian XC, Li
XS, Wu Y, Li Y, Jiang Y and Sun XC: Peli1 contributions in
microglial activation, neuroinflammatory responses and neurological
deficits following experimental subarachnoid hemorrhage. Front Mol
Neurosci. 10:3982017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu LQ, Liu XR, Zhao JY, Yan F, Wang RL,
Wen SH, Wang L, Luo YM and Ji XM: Brain-selective mild hypothermia
promotes long-term white matter integrity after ischemic stroke in
mice. CNS Neurosci Ther. 24:1275–1285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jiang M, Liu X, Zhang D, Wang Y, Hu X, Xu
F, Jin M, Cao F and Xu L: Celastrol treatment protects against
acute ischemic stroke-induced brain injury by promoting an
IL-33/ST2 axis-mediated microglia/macrophage M2 polarization. J
Neuroinflammation. 15:782018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jin Q, Cheng J, Liu Y, Wu J, Wang X, Wei
S, Zhou X, Qin Z, Jia J and Zhen X: Improvement of functional
recovery by chronic metformin treatment is associated with enhanced
alternative activation of microglia/macrophages and increased
angiogenesis and neurogenesis following experimental stroke. Brain
Behav Immun. 40:131–142. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li R, Liu WC, Yin J, Chen YC, Guo SQ, Fan
HY, Li XF, Zhang X, He XY and Duan CZ: TSG-6 attenuates
inflammation-induced brain injury via modulation of microglial
polarization in SAH rats through the SOCS3/STAT3 pathway. J
Neuroinflammation. 15:2312018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu Z, Ran Y, Huang S, Wen S, Zhang W, Liu
X, Ji Z, Geng X, Ji X, Du H, et al: Curcumin protects against
ischemic stroke by titrating microglia/macrophage polarization.
Front Aging Neurosci. 9:2332017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Satoh J, Kino Y, Asahina N, Takitani M,
Miyoshi J, Ishida T and Saito Y: TMEM119 marks a subset of
microglia in the human brain. Neuropathology. 36:39–49. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huang M, Wan Y, Mao L, He QW, Xia YP, Li
M, Li YN, Jin HJ and Hu B: Inhibiting the migration of M1 microglia
at hyperacute period could improve outcome of tMCAO rats. CNS
Neurosci Ther. 23:222–232. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hu X, Li P, Guo Y, Wang H, Leak R, Chen S,
Gao Y and Chen J: Microglia/macrophage polarization dynamics reveal
novel mechanism of injury expansion after focal cerebral ischemia.
Stroke. 43:3063–3070. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hwang IK, Park JH, Lee TK, Kim DW, Yoo KY,
Ahn JH, Kim YH, Cho JH, Kim YM, Won MH and Moon SM:
CD74-immunoreactive activated M1 microglia are shown late in the
gerbil hippocampal CA1 region following transient cerebral
ischemia. Mol Med Rep. 15:4148–4154. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang L, Wang R, Chen Z, Zhao H and Luo Y:
Xinnao Shutong modulates the neuronal plasticity through regulation
of microglia/macrophage polarization following chronic cerebral
hypoperfusion in rats. Front Physiol. 9:5292018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shang K, He J, Zou J, Qin C, Lin L, Zhou
LQ, Yang LL, Wu LJ, Wang W, Zhan KB and Tian DS: Fingolimod
promotes angiogenesis and attenuates ischemic brain damage via
modulating microglial polarization. Brain Res. 1726:1465092020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Yang L, Tucker D, Dong Y, Wu C, Lu Y, Li
Y, Zhang J, Liu TC and Zhang Q: Photobiomodulation therapy promotes
neurogenesis by improving post-stroke local microenvironment and
stimulating neuroprogenitor cells. Exp Neurol. 299:86–96. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zong X, Dong Y, Li Y, Yang L, Li Y, Yang
B, Tucker L, Zhao N, Brann DW, Yan X, et al: Beneficial effects of
theta-burst transcranial magnetic stimulation on stroke injury via
improving neuronal microenvironment and mitochondrial integrity.
Transl Stroke Res. Sep 12–2019.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhao SC, Wang C, Xu H, Wu WQ, Chu ZH, Ma
LS, Zhang YD and Liu F: Age-related differences in interferon
regulatory factor-4 and −5 signaling in ischemic brains of mice.
Acta Pharmacol Sin. 38:1425–1434. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Suenaga J, Hu X, Pu H, Shi Y, Hassan SH,
Xu M, Leak RK, Stetler RA, Gao Y and Chen J: White matter injury
and microglia/macrophage polarization are strongly linked with
age-related long-term deficits in neurological function after
stroke. Exp Neurol. 272:109–119. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zhao X, Wang H, Sun G, Zhang J, Edwards NJ
and Aronowski J: Neuronal interleukin-4 as a modulator of
microglial pathways and ischemic brain damage. J Neurosci.
35:11281–11291. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Choi JY, Kim JY, Kim JY, Park J, Lee WT
and Lee JE: M2 phenotype microglia-derived cytokine stimulates
proliferation and neuronal differentiation of endogenous stem cells
in ischemic brain. Exp Neurobiol. 26:33–41. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Perego C, Fumagalli S, Zanier ER, Carlino
E, Panini N, Erba E and De Simoni MG: Macrophages are essential for
maintaining a M2 protective response early after ischemic brain
injury. Neurobiol Dis. 96:284–293. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zheng Y, He RY, Wang P, Shi YJ, Zhao L and
Liang J: Exosomes from LPS-stimulated macrophages induce
neuroprotection and functional improvement after ischemic stroke by
modulating microglial polarization. Biomater Sci. 7:2037–2049.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang Q, Bian G, Chen P, Liu L, Yu C, Liu
F, Xue Q, Chung SK, Song B, Ju G and Wang J: Aldose reductase
regulates microglia/macrophages polarization through the cAMP
response element-binding protein after spinal cord injury in mice.
Mol Neurobiol. 53:662–676. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li L, Yihao T, Zhou F, Yin N, Qiang T,
Haowen Z, Qianwei C, Jun T, Yuan Z, Gang Z, et al: Inflammatory
regulation by driving microglial M2 polarization: Neuroprotective
effects of cannabinoid receptor-2 activation in intracerebral
hemorrhage. Front Immunol. 8:1122017.PubMed/NCBI
|
|
58
|
Zhang Y, Xu N, Ding Y, Zhang Y, Li Q,
Flores J, Haghighiabyaneh M, Doycheva D, Tang J and Zhang JH:
Chemerin suppresses neuroinflammation and improves neurological
recovery via CaMKK2/AMPK/Nrf2 pathway after germinal matrix
hemorrhage in neonatal rats. Brain Behav Immun. 70:179–193. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shu ZM, Shu XD, Li HQ, Sun Y, Shan H, Sun
XY, Du RH, Lu M, Xiao M, Ding JH and Hu G: Ginkgolide B protects
against ischemic stroke via modulating microglia polarization in
mice. CNS Neurosci Ther. 22:729–739. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Moretti R, Leger PL, Besson VC, Csaba Z,
Pansiot J, Di Criscio L, Gentili A, Titomanlio L, Bonnin P, Baud O
and Charriaut-Marlangue C: Sildenafil, a cyclic GMP
phosphodiesterase inhibitor, induces microglial modulation after
focal ischemia in the neonatal mouse brain. J Neuroinflammation.
13:952016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Han L, Cai W, Mao L, Liu J, Li P, Leak RK,
Xu Y, Hu X and Chen J: Rosiglitazone promotes white matter
integrity and long-term functional recovery after focal cerebral
ischemia. Stroke. 46:2628–2636. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xiang B, Xiao C, Shen T and Li XF:
Anti-inflammatory effects of anisalcohol on
lipopolysaccharide-stimulated BV2 microglia via selective
modulation of microglia polarization and down-regulation of
NF-kappaB p65 and JNK activation. Mol Immunol. 95:39–46. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chen J, Yin W, Tu Y, Wang S, Yang X, Chen
Q, Zhang X, Han Y and Pi R: L-F001, a novel multifunctional ROCK
inhibitor, suppresses neuroinflammation in vitro and in vivo:
Involvement of NF-κB inhibition and Nrf2 pathway activation. Eur J
Pharmacol. 806:1–9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Xiao L, Sun W, Lan W, Xiong Y, Duan Z,
Zhang Z, Fan W, Xu L, Xie X, Ma N, et al: Correlation between
cerebral microbleeds and S100B/RAGE in acute lacunar stroke
patients. J Neurol Sci. 340:208–212. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Chen C, Chu SF, Ai QD, Zhang Z, Guan FF,
Wang SS, Dong YX, Zhu J, Jian WX and Chen NH: CKLF1 Aggravates
focal cerebral ischemia injury at early stage partly by modulating
microglia/macrophage toward M1 polarization through CCR4. Cell Mol
Neurobiol. 39:651–669. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Chen C, Ai Q, Chu S, Zhang Z, Zhou X, Luo
P, Liu Y and Chen N: IMM-H004 protects against oxygen-glucose
deprivation/reperfusion injury to BV2 microglia partly by
modulating CKLF1 involved in microglia polarization. Int
Immunopharmacol. 70:69–79. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kong LL, Wang ZY, Hu JF, Yuan YH and Chen
NH: Inhibition of chemokine-like factor 1 improves blood-brain
barrier dysfunction in rats following focal cerebral ischemia.
Neurosci Lett. 627:192–198. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Kwon YW, Cheon SY, Park SY, Song J and Lee
JH: Tryptanthrin suppresses the activation of the LPS-treated BV2
microglial cell line via Nrf2/HO-1 antioxidant signaling. Front
Cell Neurosci. 11:182017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu R, Liao XY, Pan MX, Tang JC, Chen SF,
Zhang Y, Lu PX, Lu LJ, Zou YY, Qin XP, et al: Glycine exhibits
neuroprotective effects in ischemic stroke in rats through the
inhibition of M1 microglial polarization via the NF-κB p65/Hif-1α
signaling pathway. J Immunol. 202:1704–1714. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Butturini E, Boriero D, Carcereri de Prati
A and Mariotto S: STAT1 drives M1 microglia activation and
neuroinflammation under hypoxia. Arch Biochem Biophys. 669:22–30.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Qin C, Fan WH, Liu Q, Shang K, Murugan M,
Wu LJ, Wang W and Tian DS: Fingolimod protects against ischemic
white matter damage by modulating microglia toward M2 polarization
via STAT3 pathway. Stroke. 48:3336–3346. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Ding Y, Qian J, Li H, Shen H, Li X, Kong
Y, Xu Z and Chen G: Effects of SC99 on cerebral ischemia-perfusion
injury in rats: Selective modulation of microglia polarization to
M2 phenotype via inhibiting JAK2-STAT3 pathway. Neurosci Res.
142:58–68. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Zhou K, Chen J, Wu J, Wu Q, Jia C, Xu YXZ,
Chen L, Tu W, Yang G, Kong J, et al: Atractylenolide III
ameliorates cerebral ischemic injury and neuroinflammation
associated with inhibiting JAK2/STAT3/Drp1-dependent mitochondrial
fission in microglia. Phytomedicine. 59:1529222019. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Yang X, Xu S, Qian Y and Xiao Q:
Resveratrol regulates microglia M1/M2 polarization via PGC-1alpha
in conditions of neuroinflammatory injury. Brain Behav Immun.
64:162–172. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zhang W, Wei R, Zhang L, Tan Y and Qian C:
Sirtuin 6 protects the brain from cerebral ischemia/reperfusion
injury through NRF2 activation. Neuroscience. 366:95–104. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Xue F, Huang JW, Ding PY, Zang HG, Kou ZJ,
Li T, Fan J, Peng ZW and Yan WJ: Nrf2/antioxidant defense pathway
is involved in the neuroprotective effects of Sirt1 against focal
cerebral ischemia in rats after hyperbaric oxygen preconditioning.
Behav Brain Res. 309:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Alfieri A, Srivastava S, Siow RC, Modo M,
Fraser PA and Mann GE: Targeting the Nrf2-Keap1 antioxidant defence
pathway for neurovascular protection in stroke. J Physiol.
589:4125–4136. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Cai W, Yang T, Liu H, Han LJ, Zhang K, Hu
XM, Zhang XJ, Yin KJ, Cao YQ, Bennett MVL, et al: Peroxisome
proliferator-activated receptor γ (pparγ): A master gatekeeper in
CNS injury and repair. Prog Neurobiol. 163-164:27–28. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Yao X, Liu S, Ding W, Yue P, Jiang Q, Zhao
M, Hu F and Zhang H: TLR4 signal ablation attenuated neurological
deficits by regulating microglial M1/M2 phenotype after traumatic
brain injury in mice. J Neuroimmunol. 310:38–45. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Yan A, Zhang T, Yang X, Shao J, Fu N, Shen
FX, Fu Y and Xia W: Thromboxane A2 receptor antagonist SQ29548
reduces ischemic stroke-induced microglia/macrophages activation
and enrichment, and ameliorates brain injury. Sci Rep. 6:358852016.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Hyakkoku K, Hamanaka J, Tsuruma K,
himazawa M, Tanaka H, Uematsu S, Akira S, Inagaki N, Nagai H and
Hara H: Toll-like receptor 4 (TLR4), but not TLR3 or TLR9,
knock-out mice have neuroprotective effects against focal cerebral
ischemia. Neuroscience. 171:258–267. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Le K, Chibaatar Daliv E, Wu SS, Qian FY,
Ali AI, Yu DF and Guo Y: SIRT1-regulated HMGB1 release is partially
involved in TLR4 signal transduction: A possible
anti-neuroinflammatory mechanism of resveratrol in neonatal
hypoxic-ischemic brain injury. Int Immunopharmacol. 75:1057792019.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Di Lucente J, Nguyen HM, Wulff H, Jin LW
and Maezawa I: The voltage-gated potassium channel Kv1.3 is
required for microglial pro-inflammatory activation in vivo. Glia.
66:1881–1895. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhao Y, Wang H, Chen W, Chen L, Liu D and
Wang X and Wang X: Melatonin attenuates white matter damage after
focal brain ischemia in rats by regulating the TLR4/NF-kappaB
pathway. Brain Res Bull. 150:168–178. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Palma-Tortosa S, Hurtado O, Pradillo JM,
Ferreras-Martin R, García-Yébenes I, García-Culebras A, Moraga A,
Moro MA and Lizasoain I: Toll-like receptor 4 regulates
subventricular zone proliferation and neuroblast migration after
experimental stroke. Brain Behav Immun. 80:573–582. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Xiang HF, Cao DH, Yang YQ, Wang HQ, Zhu
LJ, Ruan BH, Du J and Wang MC: Isoflurane protects against injury
caused by deprivation of oxygen and glucose in microglia through
regulation of the Toll-like receptor 4 pathway. J Mol Neurosci.
54:664–670. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Luo Y, Wang C, Li WH, Liu J, He HH, Long
JH, Yang J, Sui X, Wang S, You Z and Wang YA: Madecassoside
protects BV2 microglial cells from oxygen-glucose
deprivation/reperfusion-induced injury via inhibition of the
toll-like receptor 4 signaling pathway. Brain Res. 1679:144–154.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Spiegel S and Milstien S:
Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat Rev Mol
Cell Biol. 4:397–407. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Gaire BP, Lee CH, Sapkota A, Lee SY, Chun
J, Cho HJ, Nam TG and Choi JW: Identification of Sphingosine
1-Phosphate Receptor Subtype 1 (S1P1) as a pathogenic
factor in transient focal cerebral ischemia. Mol Neurobiol.
55:2320–2332. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kim GS, Yang L, Zhang G, Zhao H, Selim M,
McCullough LD, Kluk MJ and Sanchez T: Critical role of
sphingosine-1-phosphate receptor-2 in the disruption of
cerebrovascular integrity in experimental stroke. Nat Commun.
6:78932015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Gaire BP, Song MR and Choi JW: Sphingosine
1-phosphate receptor subtype 3 (S1P3) contributes to
brain injury after transient focal cerebral ischemia via modulating
microglial activation and their M1 polarization. J
Neuroinflammation. 15:2842018. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Chen H: Role of thromboxane A2 signaling
in endothelium-dependent contractions of arteries. Prostaglandins
Other Lipid Mediat. 134:32–37. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Huang JS, Ramamurthy SK, Lin X and Breton
GCL: Cell signalling through thromboxane A2 receptors. Cell Signal.
16:521–533. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Tian DS, Li CY, Qin C, Murugan M, Wu LJ
and Liu JL: Deficiency in the voltage-gated proton channel Hv1
increases M2 polarization of microglia and attenuates brain damage
from photothrombotic ischemic stroke. J Neurochem. 139:96–105.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Wu LJ, Wu G, Akhavan Sharif MR, Baker A,
Jia Y, Fahey FH, Luo HR, Feener EP and Clapham DE: The
voltage-gated proton channel Hv1 enhances brain damage from
ischemic stroke. Nat Neurosci. 15:565–573. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yu Y, Yu ZY, Xie MJ, Wang W and Luo X: Hv1
proton channel facilitates production of ROS and pro-inflammatory
cytokines in microglia and enhances oligodendrocyte progenitor
cells damage from oxygen-glucose deprivation in vitro. Biochem
Biophys Res Commun. 498:1–8. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Nguyen HM, Grössinger EM, Horiuchi M,
Davis KW, Jin LW, Maezawa I and Wulff H: Differential Kv1.3,
KCa3.1, and Kir2.1 expression in ‘classically’ and ‘alternatively’
activated microglia. Glia. 65:106–121. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Wang J, Zhao H, Fan Z, Li G, Ma Q, Tao Z,
Wang R, Feng J and Luo Y: Long noncoding RNA H19 promotes
neuroinflammation in ischemic stroke by driving histone deacetylase
1-dependent M1 microglial polarization. Stroke. 48:2211–2221. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Hamzei Taj S, Kho W, Riou A, Wiedermann D
and Hoehn M: MiRNA-124 induces neuroprotection and functional
improvement after focal cerebral ischemia. Biomaterials.
91:151–165. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Shao Y, Deng T, Zhang T, Li P and Wang Y:
FAM19A3, a novel secreted protein, modulates the
microglia/macrophage polarization dynamics and ameliorates cerebral
ischemia. FEBS Lett. 589:467–475. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Wang J, Cao B, Zhao H, Gao Y, Luo Y, Chen
Y and Feng J: Long noncoding RNA H19 prevents neurogenesis in
ischemic stroke through p53/Notch1 pathway. Brain Res Bull.
150:111–117. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Huang Y, Wang L, Mao Y and Nan G: Long
noncoding RNA-H19 contributes to atherosclerosis and induces
ischemic stroke via the upregulation of acid phosphatase 5. Front
Neurol. 10:322019. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Wang J, Cao B, Han D, Sun M and Feng J:
Long non-coding RNA H19 induces cerebral ischemia reperfusion
injury via activation of autophagy. Aging Dis. 8:71–84. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Huang Y, Zheng Y, Jin C, Li X, Jia L and
Li W: Long non-coding RNA H19 inhibits adipocyte differentiation of
bone marrow mesenchymal stem cells through epigenetic modulation of
histone deacetylases. Sci Rep. 6:288972016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Han CL, Ge M, Liu YP, Zhao XM, Wang KL,
Chen N, Meng WJ, Hu W, Zhang JG, Li L and Meng FG: LncRNA H19
contributes to hippocampal glial cell activation via JAK/STAT
signaling in a rat model of temporal lobe epilepsy. J
Neuroinflammation. 15:1032018. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
William JP, Alejandro AR, Teri A, Opeolu
MA, Nicholas CB, Kyra B, José B, Michael B, Bart MD, Brian H, et
al: 2018 Guidelines for the early management of patients with acute
ischemic stroke. Stroke. 49:e46–e99. 2018.PubMed/NCBI
|
|
107
|
Woodbury ME, Freilich RW, Cheng CJ, Asai
H, Ikezu S, Boucher JD, Slack F and Ikezu T: miR-155 is essential
for inflammation-induced hippocampal neurogenic dysfunction. J
Neurosci. 35:9764–9781. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Sun Y, Luo ZM, Guo XM, Su DF and Liu X: An
updated role of microRNA-124 in central nervous system disorders: A
review. Front Cell Neurosci. 9:1932015. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zhang W and Meng A: MicroRNA-124
expression in the brains of rats during early cerebral ischemia and
reperfusion injury is associated with cell apoptosis involving
STAT3. Exp Ther Med. 17:2870–2876. 2019.PubMed/NCBI
|
|
110
|
Hamzei Taj S, Kho W, Aswendt M, Collmann
FM, Green C, Adamczak J, Tennstaedt A and Hoehn M: Dynamic
modulation of microglia/macrophage polarization by miR-124 after
focal cerebral ischemia. J Neuroimmune Pharmacol. 11:733–748. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Tom Tang Y, Emtage P, Funk WD, Hu T,
Arterburn M, Park EE and Rupp F: TAFA: A novel secreted family with
conserved cysteine residues and restricted expression in the brain.
Genomics. 83:727–734. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Wen L, You W, Wang H, Meng Y, Feng J and
Yang X: Polarization of microglia to the M2 phenotype in a
peroxisome proliferator-activated receptor gamma-dependent manner
attenuates axonal injury induced by traumatic brain injury in mice.
J Neurotrauma. 35:2330–2340. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Liu X, Wen S, Yan F, Liu K, Liu L, Wang L,
Zhao S and Ji X: Salidroside provides neuroprotection by modulating
microglial polarization after cerebral ischemia. J
Neuroinflammation. 15:392018. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Wang P, Shao BZ, Deng ZQ, Chen S, Yue ZY
and Miao CY: Autophagy in ischemic stroke. Prog Neurobiol.
163-164:98–117. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Yang Z, Zhong L, Zhong S, Xian R and Yuan
B: Hypoxia induces microglia autophagy and neural inflammation
injury in focal cerebral ischemia model. Exp Mol Pathol.
98:219–224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
He HY, Ren L, Guo T and Deng YH: Neuronal
autophagy aggravates microglial inflammatory injury by
downregulating CX3CL1/fractalkine after ischemic stroke. Neural
Regen Res. 14:280–288. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Xia CY, Zhang S, Chu SF, Wang ZZ, Song XY,
Zuo W, Gao Y, Yang PF and Chen NH: Autophagic flux regulates
microglial phenotype according to the time of oxygen-glucose
deprivation/reperfusion. Int Immunopharmacol. 39:140–148. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Jiang J, Dai JC and Cui H: Vitexin
reverses the autophagy dysfunction to attenuate MCAO-induced
cerebral ischemic stroke via mTOR/Ulk1 pathway. Biomed
Pharmacother. 99:583–590. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Li D, Wang C, Yao Y, Chen L, Liu G, Zhang
R, Liu Q, Shi F and Hao J: mTORC1 pathway disruption ameliorates
brain inflammation following stroke via a shift in microglia
phenotype from M1 type to M2 type. FASEB J. 30:3388–3399. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Jiang M, Wang H, Jin M, Yang X, Ji H,
Jiang Y, Zhang H, Wu F, Wu G, Lai X, et al: Exosomes from
mir-30d-5p-adscs reverse acute ischemic stroke-induced,
autophagy-mediated brain injury by promoting m2
microglial/macrophage polarization. Cell Physiol Biochem.
47:864–878. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Hemphill JC III, Greenberg SM, Anderson
CS, Becker K, Bendok BR, Cushman M, Fung GL, Goldstein JN,
Macdonald RL, Mitchell PH, et al: Guidelines for the management of
spontaneous intracerebral hemorrhage. A Guideline for Healthcare
Professionals From the American Heart Association/American Stroke
Association. Stroke. 46:2032–2060. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Lan X, Han X, Li Q, Yang QW and Wang J:
Modulators of microglial activation and polarization after
intracerebral haemorrhage. Nat Rev Neurol. 13:420–433. 2017.
View Article : Google Scholar : PubMed/NCBI
|














