Introduction
Necrotizing enterocolitis (NEC) is considered as one
of the most devastating diseases in neonates, especially those with
markedly low birth weight (<1,500 g) (1). NEC is characterized by necrosis of
intestinal mucosa and submucosal tissue, and is associated with
various pathogenic mechanisms, such as premature birth, hypoxia,
hyperosmotic artificial (namely formula) feeding, imbalance of the
gut microbial ecosystem, infection and inflammatory response
(1,2). NEC can be classified into three
stages: i) Stage 1 (suspect) is defined by temperature instability,
lethargy, apnea, bradycardia and mild abdominal distension; ii)
Stage 2 (definite) may be recognized as having the aforementioned
signs and symptoms plus persistent occult or gross gastrointestinal
bleeding and/or marked abdominal distension; iii) Stage 3
(advanced) is diagnosed as having the aforementioned signs and
symptoms plus deterioration of vital signs, evidence of septic
shock and/or marked gastrointestinal hemorrhage (3). The incidence of NEC in the USA
neonatal intensive care units is 3–11% (4–6).
From 1999 to 2009, the incidence of NEC in >600 hospitals in the
USA was significantly increased, while all the other neonatal
morbidities exhibited a stable or declining trend (4,6,7).
There is no effective prevention or treatment of NEC in the clinic
at present. Oral probiotic preparations, such as
Bifidobacterium and Lactobacillus species, have been
used to improve the gut microbial composition in neonates with NEC
(8).
The non-pathogenic yeast Saccharomyces
boulardii (SB) has been reported to be effective in the
prophylaxis and treatment of a wide range of enteropathies
(9). It was reported that this
probiotic yeast has beneficial effects on enteropathies, such as
improving the gut immune response and the intestinal barrier
(10,11). In addition, previous studies have
suggested that SB improves intestinal necrosis in neonatal mice
with NEC; however, the majority of the mechanisms require further
elucidation (12–14).
Sirtuin 1 (SIRT1) is a member of the sirtuin family,
and it is widely expressed in a variety of organisms, from yeast to
mammals (15). Previous results
indicated that SIRT1 is closely associated with inflammation
(16). Several inflammatory
transcription factors such as nuclear factor-κB (NF-κB) are
modulated by SIRT1. Overexpression of SIRT1 has been reported to
enhance NF-κB p65 subunit deacetylation, and suppress NF-κB
transcription and activity (17).
Once NF-κB is activated, it upregulates the expression of
downstream cytokines, such as tumor necrosis factor-α (TNF-α) and
interleukin-6 (IL-6), which will eventually lead to an inflammatory
response (18). However, the role
of the SIRT1/NF-κB pathway in neonatal mice with NEC, and whether
it can be regulated by SB treatment, is still not fully
understood.
Furthermore, the main microbial composition in the
intestine of mice is Eubacterium rectale/Clostridium
coccoides, Clostridium leptum, Lactobacillus species (sp.),
Bacteroides sp., mouse intestinal Bacteroides and
segmented filamentous bacteria, which are important for the
intestinal microenvironment (19).
Therefore, whether modulation of the gut microbiota is responsible
for the role of SB on neonatal mice with NEC should be
investigated.
The present study revealed that neonatal mouse
intestinal epithelial SIRT1 and its downstream NF-κB were
critically involved in the protective role of SB in mice with NEC.
Furthermore, the modulation of the gut microbiota was also
associated with the SB effect on NEC.
Materials and methods
Animals
A total of 76 newborn male and female C57BL/6 mice
(8 weeks old, weight 2–4 g) were obtained from the Experimental
Animal Center of Shanghai Jiao Tong University Affiliated Sixth
People's Hospital. Intestinal epithelial specific SIRT1 knockout
(KO) newborn mice (SIRT1KO, villin-cre+,
SIRT1flox/flox) and their littermate Flox
controls (villin-cre−, SIRT1flox/flox)
on a C57BL/6 background were generated as previously described
(20). The mice were socially
housed in accordance with the Guide for the Care and Use of
Laboratory Animals (NIH Publication, 8th Edition, 2011). All animal
experiments were approved by the Ethics Committee on Animal Care of
Shanghai Jiao Tong University Affiliated Sixth People's
Hospital.
Artificial feeding
The neonatal mice were placed into an incubator with
a constant temperature of 36°C and a humidity of 55% on the day of
birth, and were artificially fed using a formula previously
described (Table I) (21). The neonatal mice were intubated
with a sterile 1.9F silicone tube (BD Biosciences) every 4 h. The
skin around the mouth of the neonatal mice was cleaned with 0.9%
sodium chloride and 75% ethanol solution before feeding. The first
feeding volume was 0.1 ml, and 0.1–0.3 ml was added every 24 h.
 | Table I.Comparison of AF formula and main
components of mouse milk. |
Table I.
Comparison of AF formula and main
components of mouse milk.
|
| Substitute | Mouse |
|---|
| Ingredients | milk (1 l) | milk (1 l) |
| Sugar (g) | 28.04 | 26 |
| Protein (g) | 100 | 69-118 |
| Fat (g) | 105.7 | 93-175 |
| Calories (kJ) | 5,817.23 | 5,200-9,300 |
Induction of NEC model and
Saccharomyces boulardii (SB) treatment
Mice with NEC were established as previously
described (22). Briefly, neonatal
mice were fed with lipopolysaccharide (LPS; Sigma-Aldrich; Merck
KGaA) at 2 mg/kg in addition to artificial feeding for 3 days.
Furthermore, neonatal mice were exposed to hypoxia (99%
N2) for 1 min in a hypoxic chamber, followed by
hypothermia (4°C) for 10 min twice daily for an additional 3 days
(23). The NEC model was
successfully established when the animals appeared to be severely
ill with signs of cyanosis, lethargy and/or abdominal distension,
which was similar to the human NEC in stage II and III. The mice
continued to be artificially fed for 3 days and then sacrificed by
cervical dislocation. During this period, if the mouse exhibited
more serious symptoms, such as severe abdominal distention,
disappearance of bowel sounds, serious decrease of heart rate,
hypothermia, and/or shock, it was sacrificed by cervical
dislocation. After establishing the NEC model, the newborn mice
were orally administered with SB sachets (>1.3×109
CFU/g; Laboratoires Biocodex) once a day at 800 mg/kg/day for 3
days. After the gavage, the neonatal mice were returned to the
incubator, and the mice with NEC were sacrificed after 3 day SB
treatment. Notably, our previous study indicated that 800 mg/kg SB
significantly increased the body weight and survival cycle, and
decreased the pathological score of neonatal mice with NEC compared
with other SB doses (unpublished data). Therefore, 800 mg/kg SB was
used in the present study.
Material acquisition
Mice from the normal and control groups were
sacrificed 6 days after breastfeeding or artificial feeding. Mice
with NEC were sacrificed 6 days after the induction of NEC, while
NEC + SB mice were sacrificed 3 days after SB treatment. These
samples were all acquired at the same time among the experimental
groups. After euthanasia, mice were incised from the median region,
and the tissues and blood vessels were separated. Next, the
ileocecum (~5 cm) was obtained, fixed in 4% paraformaldehyde for 24
h at room temperature, and embedded in paraffin. In addition, a
section of ileocecal intestinal tissue was stored for subsequent
use in immunofluorescence staining, western blotting and reverse
transcription-quantitative PCR (RT-qPCR).
Hematoxylin and eosin (H&E)
staining and pathological score assessment
The embedded ileocecal intestinal tissue was sliced
at a thickness of 4 µm, and stained with an H&E Staining kit
(Beyotime Institute of Biotechnology) following the manufacturer's
instructions. The diagnosis of NEC was performed based on the
assessment of two independent pathologists, who were blinded in
this study. NEC was scored based on the severity of the damage from
the mucosa to the lamina propria as follows (24): i) 0, intact villi and epithelium
with normal tissue structure; ii) 1, mild separation or edema in
the submucosa and/or lamina propria; iii) 2, moderate separation or
edema in the submucosa and/or lamina propria; iv) 3, severe
separation or edema in the submucosa and/or lamina propria, and
villi shedding; and v) 4, complete absence of epithelial structures
and transmural necrosis. A pathological score ≥2 was classified as
NEC.
Immunofluorescence staining
The ileocecal tissue sections were fixed for 15 min
with 4% paraformaldehyde at room temperature, blocked with 5% BSA
(Beyotime Institute of Biotechnology) at room temperature for 30
min, and incubated with primary antibodies (SIRT1; 1:1,000; cat.
no. ab220807; Abcam) at 4°C overnight. Following which, tissues
were incubated at room temperature for 1 h with donkey anti-mouse
lgG Alexa Fluor 350-conjugated secondary antibody (cat. no. A10035)
and donkey anti-mouse lgG Alexa Fluor 594-conjugated secondary
antibody (cat. no. A32744; 1:1,000; both purchased from Thermo
Fisher Scientific, Inc.). Then, the sections were stained with DAPI
(Sigma-Aldrich; Merck KGaA) at room temperature for 10 min. After
final washing, the coverslips were mounted on slides using 50%
glycerin. The sections were then observed using a fluorescence
microscope (Olympus Corporation).
Western blotting
Cytoplasmic and nuclear proteins were extracted from
ileocecal tissues using a Nuclear and Cytoplasmic Protein
Extraction kit (Beyotime Institute of Biotechnology) according to
the manufacturer's instructions. Protein concentrations were
measured via bicinchoninic acid assay (Beyotime Institute of
Biotechnology). Proteins (40 µg) were loaded onto each lane and
separated by 8–12% SDS-PAGE and electro-transferred to
nitrocellulose membranes (EMD Millipore). Then, the membranes were
probed with antibodies against SIRT1 (cat. no. ab220807), NF-κB
(cat. no. ab16502), β-actin (cat. no. ab8226) or lamin B1 (cat. no.
ab16048; 1:1,000; all purchased from Abcam) at 4°C overnight,
followed by incubation with a LICOR IRDye® 800CW goat
anti-mouse lgG secondary antibody (cat. no. 926-32210) and IRDye
680RD goat anti-rabbit IgG (cat. no. 926-32221; 1:10,000; both
purchased from LI-COR Biosciences) at room temperature for 2 h.
Finally, the signals were detected by Odyssey Infrared Imaging
System (LI-COR Biosciences). Digitized images were determined by
ImageJ software (v1.47; National Institutes of Health). Protein
expression levels were calculated from the ratio of corresponding
protein to β-actin (cytoplasmic loading control) or lamin B1
(nuclear loading control) multiplied by 100%.
RT-qPCR and qPCR
Total RNA from ileocecal tissues was extracted with
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
following the manufacturer's protocol. The RNA was then subjected
to RT to synthesize cDNA using the PrimeScript RT Reagent kit with
gDNA Eraser (Takara Biotechnology Co., Ltd.) at 42°C for 15 min and
85°C for 5 min. Subsequently, 20-µl reactions with SIRT1, NF-κB or
β-actin primers (Genewiz, Inc.) were detected using a PikoReal 96
Real-Time PCR system (Thermo Fisher Scientific, Inc.) with SYBR
Green PCR Master mix (Applied Biosystems; Thermo Fisher Scientific,
Inc.). Relative quantitative analysis in the mRNA expression was
obtained using the 2−ΔΔCq method (25) and normalized to β-actin.
Total DNA from the feces and intestinal contents was
extracted using the QIAamp DNA Stool Mini kit (Qiagen GmbH). The
abundance of gut microbial composition was assessed by qPCR using
specific 16S ribosomal DNA (rDNA) primers (Genewiz, Inc.). The data
were quantified by calculating the abundance of bacterial
group-specific 16S rRNA genes and normalized to total bacterial
rRNA genes.
The sequences of the primers were as follows: SIRT1
forward, 5′-CAGCTCTGCTACAATTCATCGCGTC-3′ and reverse,
5′-AATCTCTGTAGAGTCCAGCGCGTGTG-3′; NF-κB forward,
5′-ACGAGCAGATGGTCAAGGAG-3′ and reverse, 5′-CTTCCATGGTCAGTGCCTTT-3′;
TNF-α forward, 5′-TACACCTCACCCACACAGTC-3′ and reverse,
5′-CGCACCTCACAGACTGTTTT-3′; IL-6 forward, 5′-GCCAGTTGCCTTCTTGGG-3′
and reverse, 5′-CGACTTGTGAAGTGGTATA-3′; β-actin forward,
5′-TAAAGACCTCTATGCCAACACAGT-3′ and reverse,
5′-CACGATGGAGGGCCGGACTCATC-3′; Eubacterium rectale/Clostridium
coccoides (Erec) forward, 5′-ACTCCTACGGGAGGCAGC-3′ and reverse,
5′-GCTTCTTAGTCAGGTACCGTCAT-3′; Clostridium leptum (Clept)
forward, 5′-GTTGACAAAACGGAGGAAGG-3′ and reverse,
5′-GACGGGCGGTGTGTACAA-3′; Lactobacillus sp. (Lact) forward,
5′-GACGGGCGGTGTGTACAA-3′ and reverse, 5′-CACCGCTACACATGGAG-3′;
Bacteroides sp. (Bact) forward, 5′-CACCGCTACACATGGAG-3′ and
reverse, 5′-GCTGCCTCCCGTAGGAGT-3′; mouse intestinal
Bacteroides (MIB) forward, 5′-CCAGCAGCCGCGGTAATA-3′ and
reverse, 5′-CCAGCAGCCGCGGTAATA-3′; segmented filamentous bacteria
(SFB) forward, 5′-GACGCTGAGGCATGAGAGCAT-3′ and reverse,
5′-GACGCTGAGGCATGAGAGCAT-3′; and total bacteria forward,
5′-ACTCCTACGGGAGGCAGCAGT-3′ and reverse,
5′-ATTACCGCGGCTGCTGGC-3′.
Statistical analysis
Data were expressed as the means ± standard error of
mean. Statistical analysis was performed by SPSS 23.0 (IBM Corp.).
Significant differences between two groups were analyzed via
Student's t-test, multiple comparisons of >two groups were
performed using Tukey's honest significant difference test, and
P<0.05 was considered to indicate a statistically significant
difference.
Results
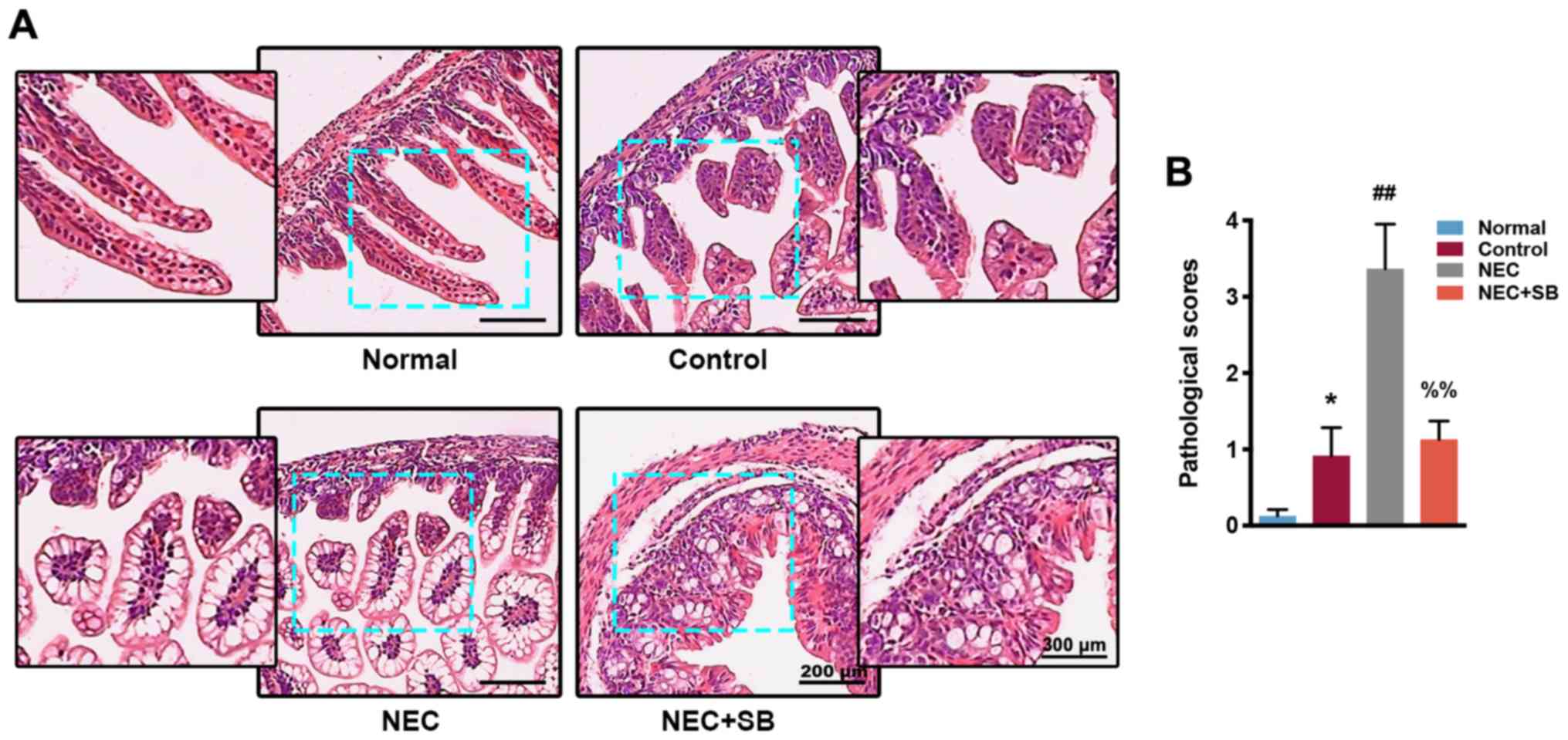
SB treatment improves NEC-induced
intestinal damage
Based on macroscopic observation of the intestinal
specimens, the intestine of normal neonatal mice was bright in
color, straight and smooth, without gas accumulation. The color of
the intestine in the control (artificial feeding) group was
slightly darkened, and the ileum was partially dilated. In the NEC
group, the intestine was severely dilated, blackened and congested.
In addition, the NEC mice had severe intestinal gas accumulation.
The intestine of the NEC + SB neonatal mice was visually relieved,
only showing mild edema and only slightly darkened in color, which
was significantly improved compared with the NEC group. H&E
staining revealed that the ileocecal intestinal tissue of the
normal group was intact with continuous epithelium, regular glands
and neat villi, and the mucosa, submucosa and lamina propria were
free of congestion and edema (Fig.
1A). The pathological score was low in the normal group
(Fig. 1B). The ileocecal mucosa,
submucosa and lamina propria of the control group exhibited mild
congestion and edema, and the glands were disorderly arranged,
resulting in an increase in pathological score compared with the
normal group (P<0.05). Furthermore, the ileocecum in the NEC
group had obvious villi degeneration and edema, disordered gland
arrangement, severe congestion, and edema of mucosa, submucosa and
lamina propria. The pathological score was significantly higher
than that of the control group (P<0.01). The NEC + SB group had
a mild edema in the mucosa, submucosa and lamina propria, and the
villi were uneven, although there were some arrangements, and the
glands were arranged regularly. The pathological score in the NEC +
SB group was significantly decreased compared with the NEC group
(P<0.01).
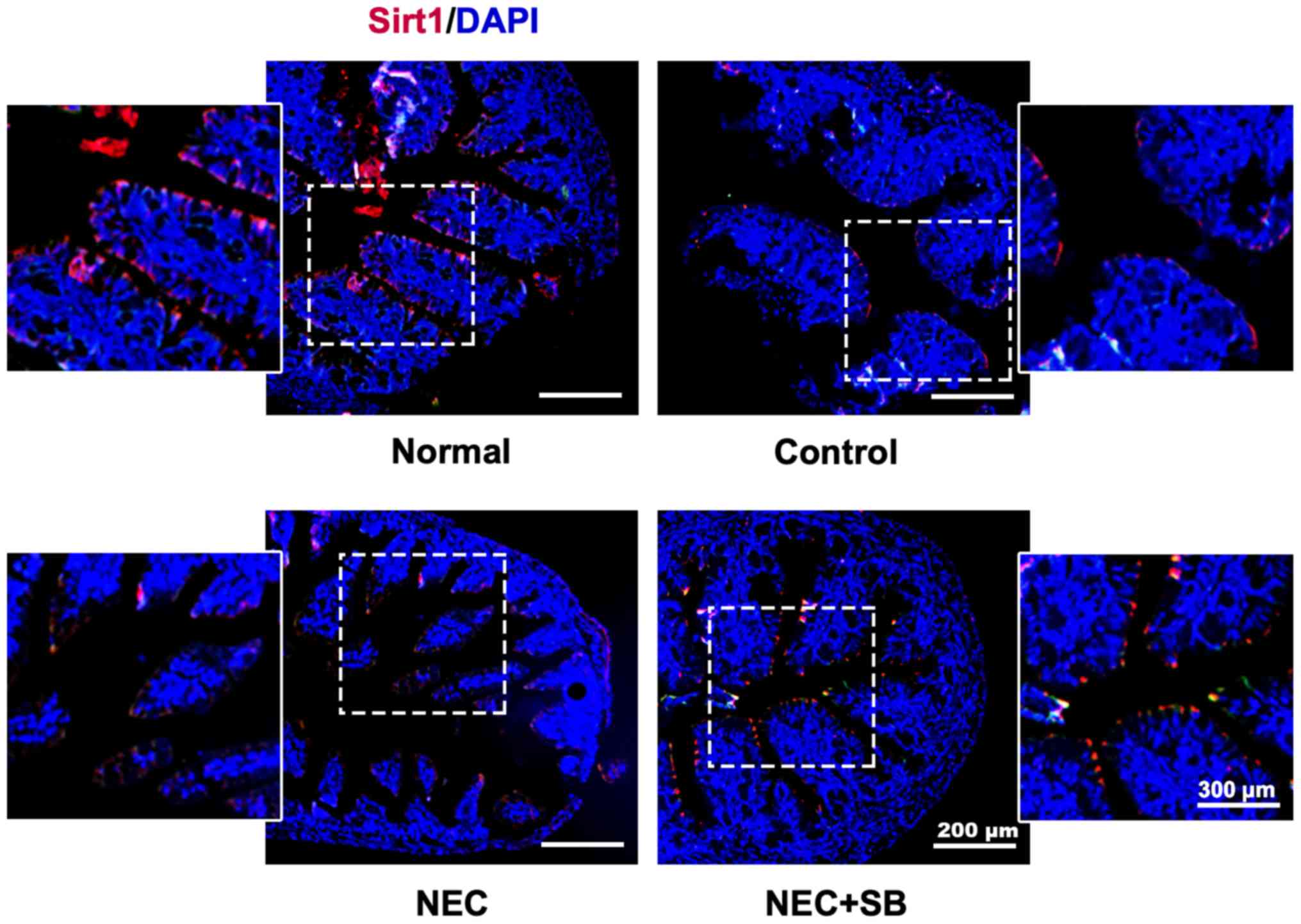
SB induces upregulation of SIRT1
expression in NEC neonatal mice
The results from immunofluorescence staining
indicated that a relatively high level of SIRT1 protein expression
was primarily located in the intestinal epithelial cells of the
normal group (Fig. 2). Artificial
feeding (control) could lead to a decrease in the expression of
SIRT1 in the ileocecum of neonatal mice, and there was a further
decrease in the level of SIRT1 in the NEC group compared with the
control group. Notably, NEC mice fed with SB could reverse the
decrease in SIRT1 and markedly enhance its expression level. These
results revealed that SB treatment enhanced SIRT1 protein
expression in NEC neonatal mice.
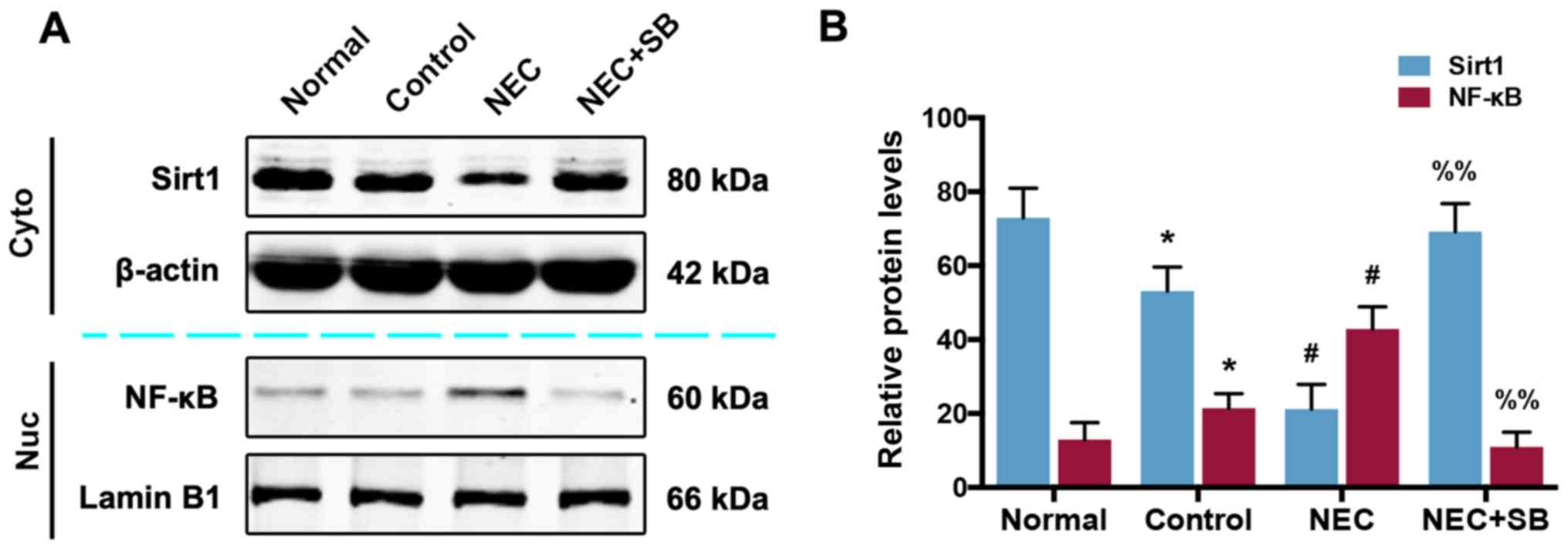
SB treatment in NEC mice results in an
activation of the SIRT1/NF-κB pathway
To determine whether the SIRT1/NF-κB pathway is
involved in the role of SB in NEC mice, western blotting was used
to analyze the protein expression of SIRT1 and NF-κB. As revealed
in Fig. 3A and B, the expression
of cytoplasmic SIRT1 and nuclear NF-κB proteins in the control
group was slightly decreased and increased, respectively, compared
with the normal group (P<0.05). NEC insult led to a further
decrease and increase of SIRT1 and NF-κB (P<0.01). SB treatment
in NEC mice reversed the decrease in SIRT1 and the increase in
NF-κB, and significantly promoted SIRT1 expression and reduced
NF-κB expression (P<0.01).
SB induces upregulation of SIRT1 and
downregulation of NF-κB mRNA levels in neonatal mice with NEC
Next, the effect of SB on SIRT1, NF-κB, TNF-α and
IL-6 mRNA expression in NEC mice was investigated. Similar to the
changes in SIRT1 and NF-κB at the protein level, a decrease in
SIRT1 and an increase in NF-κB mRNA was observed in the control
group compared with the normal group (P<0.05; Fig. 4A and B). NEC insult induced a
significant decrease in SIRT1 mRNA and an increase in NF-κB mRNA
levels compared with the control group (P<0.01). In addition,
the mRNA levels of TNF-α and IL-6 in the NEC group were
significantly increased compared with the control group (P<0.01;
Fig. 4C and D). SIRT1 mRNA
expression was significantly upregulated, and the mRNA levels of
the members of the NF-κB pathway were all downregulated in the NEC
+ SB group compared with the NEC group (P<0.01).
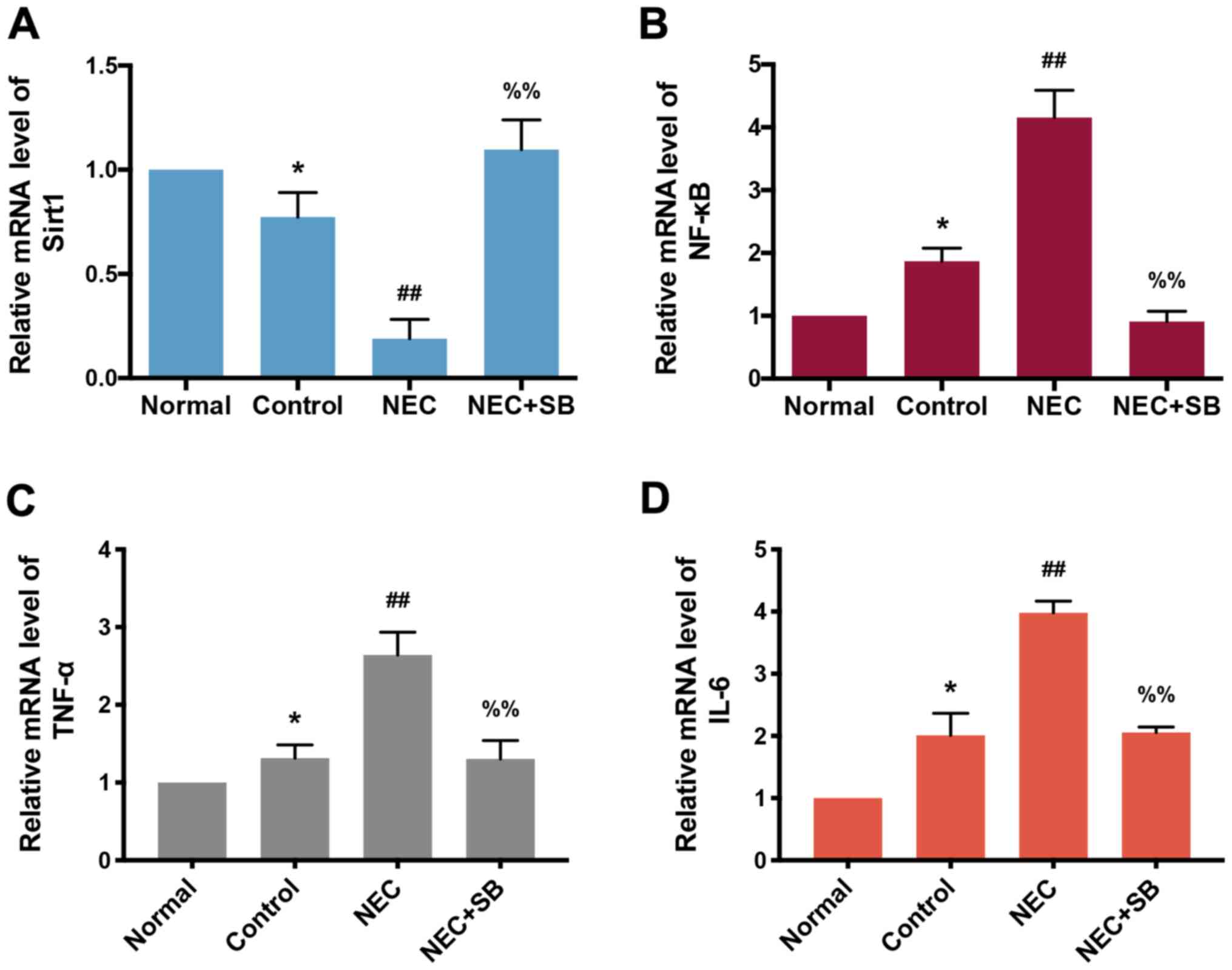 | Figure 4.SIRT1 and NF-κB mRNA levels in
ileocecal intestinal tissue of neonatal mice. (A) SIRT1 mRNA
expression in the normal, control, NEC and NEC + SB groups was
determined by RT-qPCR (n=7). (B) mRNA levels of NF-κB in the
normal, control, NEC and NEC + SB groups were analyzed by RT-qPCR
(n=7). (C and D) The mRNA levels of (C) TNF-α and (D) IL-6 in the
normal, control, NEC and NEC + SB groups were analyzed by RT-qPCR
(n=7). Data were compared using Tukey's honest significant
difference test. Error bars indicate the standard error of mean.
*P<0.05 vs. normal; ##P<0.01 vs. control;
%%P<0.01 vs. NEC. SIRT1, sirtuin 1; NF-κB, nuclear
factor-κB; NEC, necrotizing enterocolitis; SB, Saccharomyces
boulardii; RT-qPCR, reverse transcription-quantitative PCR;
TNF-α, tumor necrosis factor-α; IL-6, interleukin-6. |
Intestinal-specific SIRT1 KO
(SIRT1KO) inhibits the protective effect of SB on NEC
neonatal mice
To further testify the possible function of the
SIRT1 pathway in NEC neonatal mice, an intestinal
epithelium-specific SIRT1KO mouse model was generated,
in which the exon 4 of the mouse SIRT1 gene was deleted throughout
the length of the intestinal epithelium (20). There was no obvious intestinal
damage in SIRT1KO mice with breastfeeding or artificial
feeding (Fig. 5A). The ileocecal
lesions caused by NEC in SIRT1KO neonatal mice were
significantly increased compared with the control (co-housed paired
Flox) group, and SB treatment did not reverse this pathological
damage. Furthermore, the pathological score of SIRT1KO
with artificial feeding was increased compared with
SIRT1KO with breastfeeding (P<0.05). The score of
SIRT1KO in the NEC + SB group was also significantly
increased compared with the control group (P<0.01). In addition,
both the SIRT1 protein (Fig. 5B and
C) and mRNA (Fig. 5D)
expression in the SIRT1KO NEC + SB group were
significantly decreased compared with the control group
(P<0.01). Unexpectedly, the NF-κB protein (Fig. 5B and C) and mRNA (Fig. 5E) levels, and the mRNA levels of
its downstream partners TNF-α and IL-6 (Fig. 5F and G) were all significantly
increased in the SIRT1KO group compared with the control
group (P<0.01). All these results indicated that knockdown of
SIRT1 suppressed the effect of SB in NEC mice.
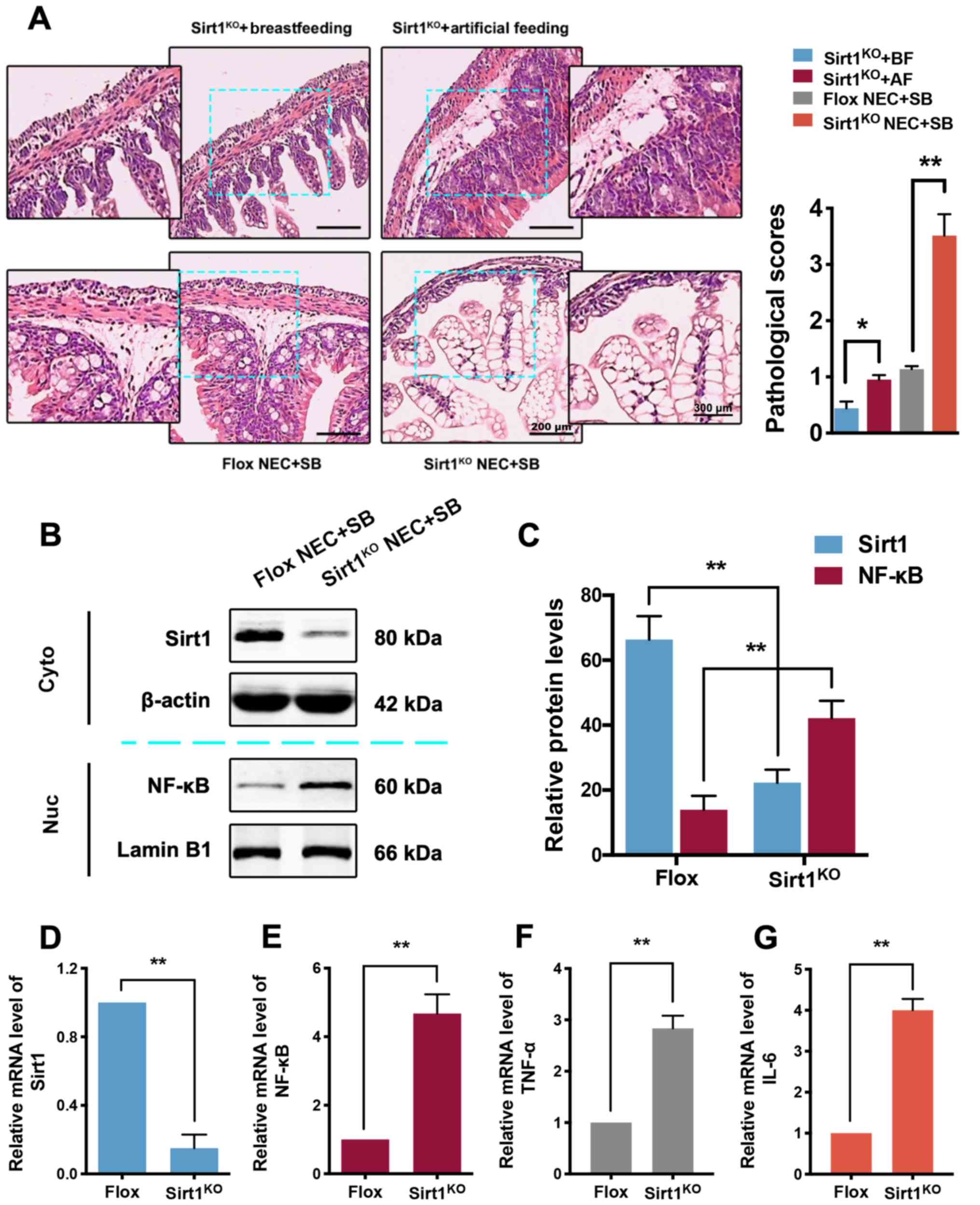 | Figure 5.Pathological changes, SIRT1 and NF-κB
expression in the ileocecum of the intestinal epithelium of
SIRT1KO mice. (A) Pathological changes (bar, 200 µm) and
scores (n=3) in the ileocecum of SIRT1KO mice with
breastfeeding, SIRT1KO mice with artificial feeding,
control + NEC + SB mice and SIRT1KO + NEC + SB mice
(n=7). *P<0.05, **P<0.01 vs. indicated group. (B) The protein
expression of SIRT1 and NF-κB in SIRT1KO and control
mice was determined by western blotting. (C) Western blot data are
presented by bar plots (n=3). The mRNA expression of (D) SIRT1, (E)
NF-κB, (F) TNF-α and (G) IL-6 in SIRT1KO and control
mice was analyzed by RT-qPCR (n=3). **P<0.01 vs. indicated
group. Data were compared using the Tukey's honest significant
difference test. Error bars indicate standard error of mean. SIRT1,
sirtuin 1; NF-κB, nuclear factor-κB; KO, knockout; NEC, necrotizing
enterocolitis; SB, Saccharomyces boulardii; BF,
breastfeeding; AF, artificial feeding; TNF-α, tumor necrosis
factor-α; IL-6, interleukin-6. |
Protective role of SB in NEC neonatal
mice is also associated with gut microbiota regulation
To evaluate whether SB-induced alterations in NEC
mice is associated with the regulation of the gut microbial
ecosystem, total fecal microbiota profiles of neonatal mice were
analyzed via 16S rRNA amplicon sequencing. Feces were collected,
and the proportions of microbiota were evaluated by qPCR. As
revealed in Fig. 6A, the abundance
of all gut microbial composition except Lact, including Erec,
Clept, Bact, MIB and SFB were decreased in the control group
compared with the normal group (P<0.05). Furthermore, NEC insult
induced a significant decrease in gut microbial abundance compared
with the control group (P<0.01 in Erec, Clept and Lact;
P<0.05 in Bact, MIB and SFB). SB treatment significantly
upregulated the abundance of gut microbiota in NEC mice compared
with the NEC group (P<0.01 in Erec, Clept, Lact and Bact;
P<0.05 in MIB and SFB). Notably, the gut microbial abundance in
the SIRT1KO NEC + SB group was significantly decreased
compared with the Flox NEC + SB group (P<0.01; P<0.05 in
Lactl; Fig. 6B). These results
revealed that the modulation of gut microbiota was also involved in
the role of SB in NEC neonatal mice.
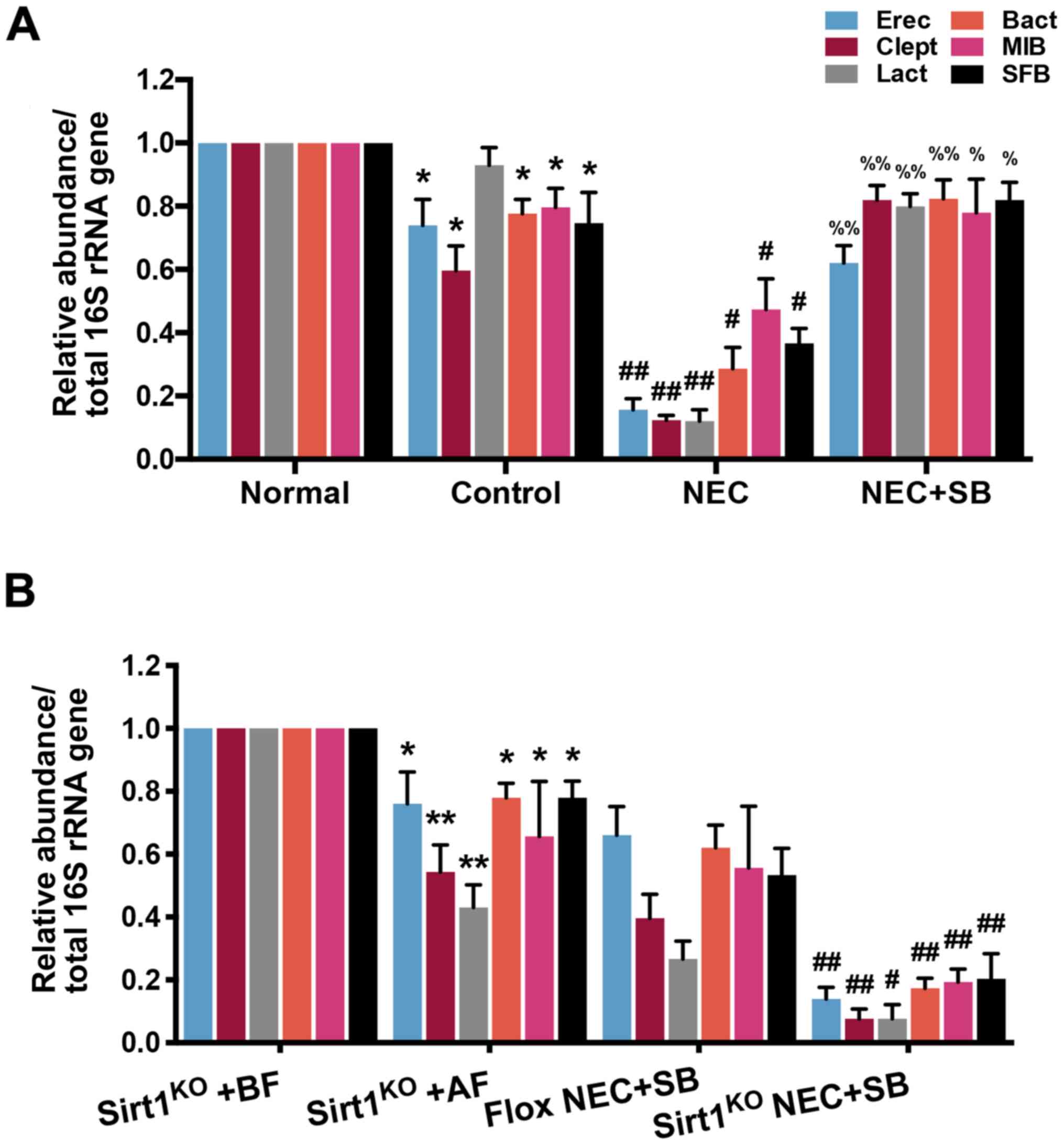 | Figure 6.Abundance of gut microbial
composition in neonatal mice. (A) The relative abundance of gut
microbiota in normal, control, NEC and NEC + SB mice was detected
by qPCR (n=7). (B) The relative abundance of gut microbiota in
SIRT1KO mice with BF, SIRT1KO mice with AF,
SIRT1KO NEC + SB mice and Flox NEC + SB mice was
determined by qPCR (n=3). *P<0.05, **P<0.01 vs. normal or
SIRT1KO + BF; #P<0.05,
##P<0.01 vs. control or Flox NEC + SB;
%P<0.05, %%P<0.01 vs. NEC. Data were
compared using Tukey's honest significant difference test. Error
bars indicate standard error of mean. NEC, necrotizing
enterocolitis; SB, Saccharomyces boulardii; qPCR,
quantitative PCR; SIRT1, sirtuin 1; KO, knockout; BF,
breastfeeding; Erec, Eubacterium rectale/Clostridium
coccoides; Clept, Clostridium leptum; Lact,
Lactobacillus sp.; Bact, bacteroides sp.; MIB, mouse
intestinal Bacteroides; SFB, segmented filamentous
bacteria. |
Discussion
NEC is one of the most common and devastating
gastrointestinal emergencies in markedly low birth weight
(<1,500 g) infants in neonatal intensive care units, with
reported mortality rates between 15 and 30% (1,26).
NEC is a multifactorial disease, and is a largely unpredictable
disease in newborns, the etiology of which remains unclear despite
the advances in research (1).
Multiple factors are reported to be responsible for the
pathogenesis of NEC, including immaturity of the gut,
hypoxia-ischemia, artificial (namely formula) feeding and gut
microbial dysbiosis, which play a role in inducing an inflammatory
response in the gut (27).
Inflammatory mediators such as NF-κB are activated to produce an
inflammatory cascade response, contributing to mucosal damage
(28). This inflammatory cascade
induces the activation of neutrophils, increases the permeability
of the vasculature and releases reactive oxygen species, leading to
vasoconstriction with ischemic-reperfusion injury (28). The mucosal barriers in the gut
continue to break down, resulting in a severe NEC, which could lead
to sepsis or even death (29).
The yeast SB is a known ‘generally regarded as safe’
microorganism with probiotic activity against various types of
microbial pathogens in the intestine (30). According to previous studies, SB
treatment in mice induces an immunomodulatory effect, and leads to
an increased level of secretory IgA and serum IgG (31,32),
as well as serum IgM (33). These
immunological roles, along with other probiotic features of SB,
such as bile and acid resistance, and an optimum growth at 37°C,
make SB a potential therapy with oral delivery in intestinal
diseases (30,34). However, whether SB has an effect on
the inflammatory response and microbial dysbiosis in the
pathogenesis of NEC mice remains to be explored.
SIRT1 is essential for cell survival,
differentiation, senescence and metabolism due to its role on
anti-inflammation and antioxidation (35). SIRT1 deacetylates NF-κB p65 subunit
at Lys310, which results in inhibition of NF-κB activity and
transcriptional silence of pro-inflammatory genes, such as IL-1β,
IL-6 and TNF-α (36,37). The function of SIRT1 and NF-κB in
the role of SB in NEC mice warrants further study.
The present study demonstrated that SB relieved the
NEC insult in neonatal mice through the SIRT1/NF-κB pathway and gut
microbial regulation. SIRT1 deficiency in the intestinal epithelium
results in increased pathological damage and NF-κB expression,
which in turn leads to a reduction in gut microbial composition.
The pathological injury and score in the control group were higher
than those of the normal group, and a further increase was observed
in the NEC group. Treatment with SB in the NEC group significantly
reduced both the pathological injury and score. Furthermore, the
expression and location of SIRT1 protein was verified by
immunofluorescence staining, and a relatively high expression of
SIRT1 protein expression, which was primarily located in the
intestinal epithelial cells of the normal group, was determined.
NEC insult induced a marked decrease in SIRT1 expression, which was
inhibited by SB treatment. Furthermore, the results from western
blotting and RT-qPCR revealed that the protein and mRNA expression
levels of SIRT1 were decreased, and the expression levels of NF-κB
and its downstream cytokines were all increased in the normal group
compared with the control group. A further decrease in SIRT1 and an
increase in the expression of molecules of the NF-κB pathway was
observed in NEC neonatal mice. SB treatment significantly relieved
the NEC role on the SIRT1/NF-κB pathway activation. In addition,
intestinal epithelium-specific SIRT1KO abolished the
effects of SB in NEC neonatal mice, including a decrease in
pathological damage and score, inhibition of SIRT1/NF-κB pathway
activation, and changes in the mRNA expression of SIRT1 and NF-κB.
Finally, the abundance of gut microbial composition, including
Erec, Clept, Bact, MIB and SFB (but not Lact) was decreased in the
control group compared with the normal group. NEC insult further
decreased the abundance of microbiota (including Lact) compared
with the control group. SB treatment reversed the decrease in all
gut microbiota in NEC neonatal mice. As anticipated, the effect of
SB on gut microbiota modulation was abolished in SIRT1KO
neonatal mice. These results indicated that the abundance of gut
microbial composition was modulated by SIRT1. Notably, there was a
mild gut dysbiosis in SIRT1KO mice with artificial
feeding (without NEC) compared with SIRT1KO mice with
breastfeeding. The present study hypothesized that the dysbiosis of
intestinal microbiota is more likely to be the cause rather than
the consequence of NEC, as there is still gut dysbiosis in the
absence of NEC. The possible mechanisms of the SB effect on SB
expression may be associated with a direct enhancement in SIRT1
gene transcription or with indirect regulation of specific
microRNAs and/or long noncoding RNAs (lncRNAs). The main
limitations of this study are the lack of exploration of the
specific mechanisms of the roles of SB on gut microbiota regulation
and the absence of relative human tissue samples. In addition,
since numerous other signaling pathways (including TLRs, JNK/STAT
and cAMP) also affect gut inflammation, further studies to verify
the association between SB and these pathways are required.
Furthermore, as NEC can be divided into three stages (namely I, II
and III), stages II and III were included in the present study;
however, the present study did not differentiate between the
therapeutic effect of SB in stages II and III.
Collectively, the present study provides a new
insight and further understanding of the mechanisms of the
protective role of SB on NEC. Furthermore, the present results
provide a theoretical basis for SB as a preventive and therapeutic
agent for NEC in the future.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Development Fund of Pudong New District (grant no.
PKJ2017-Y09).
Availability of data and materials
The datasets generated and/or analyzed during the
present study are available from the corresponding author on
reasonable request.
Authors' contributions
KZ designed the research, performed the experiments,
analyzed the data and wrote the paper. XZ designed and performed
experiments and analyzed the data. AL performed and analyzed
experiments. SF provided technical support and substantial
contributions to conception and design. JZ directed the study,
managed the project, interpreted the data and co-wrote the paper.
All authors read and approved the final manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All animal experiments were approved by the Ethics
Committee on Animal Care of Shanghai Jiao Tong University
Affiliated Sixth People's Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Bact
|
Bacteroides
|
|
Clept
|
Clostridium leptum
|
|
DAPI
|
4′,6-diamidino-2-phenylindole
|
|
Erec
|
Eubacterium rectale/Clostridium
coccoides
|
|
H&E
|
hematoxylin & eosin
|
|
IL-6
|
interleukin-6
|
|
Lact
|
Lactobacillus
|
|
LPS
|
lipopolysaccharide
|
|
MIB
|
mouse intestinal
bacteroides
|
|
NEC
|
necrotizing enterocolitis
|
|
NF-κB
|
nuclear factor-κB
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
SB
|
Saccharomyces boulardii
|
|
SDS-PAGE
|
sodium dodecyl sulfate-polyacrylamide
gel
|
|
SFB
|
segmented filamentous bacteria
|
|
SIRT1
|
sirtuin 1
|
|
sp.
|
species
|
|
TNF-α
|
tumor necrosis factor-α
|
References
|
1
|
Neu J and Walker WA: Necrotizing
enterocolitis. N Engl J Med. 364:255–264. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kafetzis DA, Skevaki C and Costalos C:
Neonatal necrotizing enterocolitis: An overview. Curr Opin Infect
Dis. 16:349–355. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bell MJ, Ternberg JL, Feigin RD, Keating
JP, Marshall R, Barton L and Brotherton T: Neonatal necrotizing
enterocolitis. Therapeutic decisions based upon clinical staging.
Ann Surg. 187:1–7. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guthrie SO, Gordon PV, Thomas V, Thorp JA,
Peabody J and Clark RH: Necrotizing enterocolitis among neonates in
the United States. J Perinatol. 23:278–285. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Holman RC, Stoll BJ, Clarke MJ and Glass
RI: The epidemiology of necrotizing enterocolitis infant mortality
in the United States. Am J Public Health. 87:2026–2031. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Holman RC, Stoll BJ, Curns AT, Yorita KL,
Steiner CA and Schonberger LB: Necrotising enterocolitis
hospitalisations among neonates in the United States. Paediatr
Perinat Epidemiol. 20:498–506. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Horbar JD, Carpenter JH, Badger GJ, Kenny
MJ, Soll RF, Morrow KA and Buzas JS: Mortality and neonatal
morbidity among infants 501 to 1500 grams from 2000 to 2009.
Pediatrics. 129:1019–1026. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aceti A, Gori D, Barone G, Callegari ML,
Di Mauro A, Fantini MP, Indrio F, Maggio L, Meneghin F, Morelli L,
et al: Probiotics for prevention of necrotizing enterocolitis in
preterm infants: Systematic review and meta-analysis. Ital J
Pediatr. 41:892015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hatoum R, Labrie S and Fliss I:
Antimicrobial and probiotic properties of yeasts: From fundamental
to novel applications. Front Microbiol. 3:4212012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Generoso SV, Viana M, Santos R, Martins
FS, Machado JA, Arantes RM, Nicoli JR, Correia MI and Cardoso VN:
Saccharomyces cerevisiae strain UFMG 905 protects against bacterial
translocation, preserves gut barrier integrity and stimulates the
immune system in a murine intestinal obstruction model. Arch
Microbiol. 192:477–484. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Buts JP and De Keyser N: Effects of
saccharomyces boulardii on intestinal mucosa. Dig Dis Sci.
51:1485–1492. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Serce O, Benzer D, Gursoy T, Karatekin G
and Ovali F: Efficacy of saccharomyces boulardii on necrotizing
enterocolitis or sepsis in very low birth weight infants: A
randomised controlled trial. Early Hum Dev. 89:1033–1036. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Demirel G, Erdeve O, Celik IH and Dilmen
U: Saccharomyces boulardii for prevention of necrotizing
enterocolitis in preterm infants: A randomized, controlled study.
Acta Paediatr. 102:e560–e565. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Akisu M, Baka M, Yalaz M, Huseyinov A and
Kultursay N: Supplementation with saccharomyces boulardii
ameliorates hypoxia/reoxygenation-induced necrotizing enterocolitis
in young mice. Eur J Pediatr Surg. 13:319–323. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Haigis MC and Sinclair DA: Mammalian
sirtuins: Biological insights and disease relevance. Annu Rev
Pathol. 5:253–295. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rajendrasozhan S, Yang SR, Kinnula VL and
Rahman I: SIRT1, an antiinflammatory and antiaging protein, is
decreased in lungs of patients with chronic obstructive pulmonary
disease. Am J Respir Crit Care Med. 177:861–870. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Deng Z, Jin J, Wang Z, Wang Y, Gao Q and
Zhao J: The metal nanoparticle-induced inflammatory response is
regulated by SIRT1 through NF-kappaB deacetylation in aseptic
loosening. Int J Nanomedicine. 12:3617–3636. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wen X, Chen X, Liang X, Zhao H, Li Y, Sun
X and Lu J: The small molecule NSM00191 specifically represses the
TNF-α/NF-кB axis in foot and ankle rheumatoid arthritis. Int J Biol
Sci. 14:1732–1744. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wellman AS, Metukuri MR, Kazgan N, Xu X,
Xu Q, Ren NSX, Czopik A, Shanahan MT, Kang A, Chen W, et al:
Intestinal epithelial sirtuin 1 regulates intestinal inflammation
during aging in mice by altering the intestinal microbiota.
Gastroenterology. 153:772–786. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kazgan N, Metukuri MR, Purushotham A, Lu
J, Rao A, Lee S, Pratt-Hyatt M, Lickteig A, Csanaky IL, Zhao Y, et
al: Intestine-Specific deletion of SIRT1 in mice impairs
DCoH2-HNF-1α-FXR signaling and alters systemic bile acid
homeostasis. Gastroenterology. 146:1006–1016. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Barna J, Renner E, Arszovszki A, Cservenák
M, Kovács Z, Palkovits M and Dobolyi A: Suckling induced activation
pattern in the brain of rat pups. Nutr Neurosci. 21:317–327. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Premkumar MH, Sule G, Nagamani SC,
Chakkalakal S, Nordin A, Jain M, Ruan MZ, Bertin T, Dawson B, Zhang
J, et al: Argininosuccinate lyase in enterocytes protects from
development of necrotizing enterocolitis. Am J Physiol Gastrointest
Liver Physiol. 307:G347–G354. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Radulescu A, Zhang HY, Yu X, Olson JK,
Darbyshire AK, Chen Y and Besner GE: Heparin-Binding epidermal
growth factor-like growth factor overexpression in transgenic mice
increases resistance to necrotizing enterocolitis. J Pediatr Surg.
45:1933–1939. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nadler EP, Dickinson E, Knisely A, Zhang
XR, Boyle P, Beer-Stolz D, Watkins SC and Ford HR: Expression of
inducible nitric oxide synthase and interleukin-12 in experimental
necrotizing enterocolitis. J Surg Res. 92:71–77. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yee WH, Soraisham AS, Shah VS, Aziz K,
Yoon W and Lee SK; Canadian Neonatal Network, : Incidence and
timing of presentation of necrotizing enterocolitis in preterm
infants. Pediatrics. 129:e298–e304. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Niemarkt HJ, De Meij TG, van Ganzewinkel
CJ, de Boer NKH, Andriessen P, Hütten MC and Kramer BW: Necrotizing
enterocolitis, gut microbiota, and brain development: Role of the
brain-gut axis. Neonatology. 115:423–431. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hagen PC and Skelley JW: Efficacy of
bifidobacterium species in prevention of necrotizing enterocolitis
in very-low birth weight infants. A systematic review. J Pediatr
Pharmacol Ther. 24:10–15. 2019.PubMed/NCBI
|
|
29
|
Gephart SM, McGrath JM, Effken JA and
Halpern MD: Necrotizing enterocolitis risk: State of the science.
Adv Neonatal Care. 12:77–87. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Czerucka D, Piche T and Rampal P: Review
article: Yeast as probiotics-saccharomyces boulardii. Aliment
Pharmacol Ther. 26:767–778. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rodrigues AC, Cara DC, Fretez SH, Cunha
FQ, Vieira EC, Nicoli JR and Vieira LQ: Saccharomyces boulardii
stimulates sIgA production and the phagocytic system of gnotobiotic
mice. J Appl Microbiol. 89:404–414. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Qamar A, Aboudola S, Warny M, Michetti P,
Pothoulakis C, LaMont JT and Kelly CP: Saccharomyces boulardii
stimulates intestinal immunoglobulin A immune response to
clostridium difficile toxin A in mice. Infect Immun. 69:2762–2765.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Stier H and Bischoff SC: Influence of
saccharomyces boulardii CNCM I-745on the gut-associated immune
system. Clin Exp Gastroenterol. 9:269–279. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kelesidis T and Pothoulakis C: Efficacy
and safety of the probiotic saccharomyces boulardii for the
prevention and therapy of gastrointestinal disorders. Therap Adv
Gastroenterol. 5:111–125. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Haigis MC and Guarente LP: Mammalian
sirtuins-emerging roles in physiology, aging, and calorie
restriction. Genes Dev. 20:2913–2921. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu TF, Vachharajani VT, Yoza BK and
McCall CE: NAD+-dependent sirtuin 1 and 6 proteins coordinate a
switch from glucose to fatty acid oxidation during the acute
inflammatory response. J Biol Chem. 287:25758–25769. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yoshizaki T, Milne JC, Imamura T, Schenk
S, Sonoda N, Babendure JL, Lu JC, Smith JJ, Jirousek MR and Olefsky
JM: SIRT1 exerts anti-inflammatory effects and improves insulin
sensitivity in adipocytes. Mol Cell Biol. 29:1363–1374. 2009.
View Article : Google Scholar : PubMed/NCBI
|