Introduction
Ovarian cancer is the 8th leading cause of
gynecological cancer type mortalities and cancer-associated
incidence rates worldwide (1). The
long-term survival rates of patients with ovarian cancer vary
between 40–45% depending on diverse pathological types and clinical
stages (2). Difficulties in
effectively treating ovarian cancer with regards to its formation,
development and progression remain challenging due to the
multivariate risk factors of this disease, the diagnosis at a
terminal stage and poor prognosis (3). Moreover, advances in treatment
strategies, including hormonotherapy, immunotherapy, surgical
operation, chemotherapy and radiological intervention, have not
increased the long-term survival rate of patients with ovarian
cancer (4). Therefore, effective
anticancer therapies with few side effects are urgently
required.
Propofol, a short-acting intravenous sedative
hypnotic reagent used during and after operation, is one of the
most widely accepted and commonly used intravenous sedative
hypnotic agents and has multiple advantages over other types of
these reagents in protecting neurocytes from oxidative stress and
hypoxia injury (5), as well as
exerting several non-anesthetic effects (6). Previous studies have reported that
suppressive effects of propofol are observed in different cancer
types. For example, it has been revealed that propofol suppressed
the proliferative ability and epithelial-mesenchymal transition
cascades by enhancing the expression of microRNA (miRNA/miR)-1284
in A549 lung cancer cells (7). In
endometrial cancer, Du et al (8) reported that propofol decreased
cellular proliferation, migration and invasion, but induced
apoptosis of endometrial cancer cells by regulating Sox4.
Previous studies investigating the tumor-suppressing
effects of miR-125a-5p have shown that downregulated miR-125a-5p is
found in multiple types of cancer (9,10).
Zhang et al (11) revealed
that lin-28 homologue B (LIN28B) was a direct target of miR-125a-5p
in melanoma. Furthermore, Yong et al (12) demonstrated that LIN28B expression
was increased in ovarian cancer, which could promote the cellular
proliferation and migration. LIN28B has a close association with
proliferation, apoptosis, metastasis, growth and oncogenesis in
colon cancer, non-small cell lung cancer, esophageal cancer,
prostate cancer, peripheral T-cell lymphoma, breast cancer,
nasopharyngeal carcinoma, renal cell cancer, hepatocellular
carcinoma and ovarian cancer (13–15).
Therefore, it was hypothesized that propofol could inhibit the
expression of LIN28B to reduce proliferation and metastasis by
regulating miR-125a-5p in ovarian cancer. The present study may
offer a novel and important insight into the use of anesthetics in
the surgical operation of ovarian cancer treatment.
Materials and methods
Bioinformatic analysis
Bioinformatic analysis was performed on the
GSE119055 dataset (https://www.ncbi.nlm.nih.gov/gds/?term=) that was
submitted by Dong et al (16) which contained six ovarian cancer
tissues and three healthy ovarian tissues, and was based on the
GPL21572 (miRNA-4) Affymetrix Multispecies miRNA-4 Array (ProbeSet
ID version). Subsequently, the raw data were analyzed using several
packages (pheatmap and ggplot2) of R statistical software (version
3.3.2; http://www.r-project.org/) (17) to obtain a heat map, volcano plots.
Visualization of Gene Ontology (GO; http://geneontology.org/) Slim summary, Gene Set
Enrichment Analysis (GSEA), and Kyoto Encyclopedia of Genes and
Genomes analyses (KEGG) pathway enrichment (https://www.kegg.jp/) were performed (http://www.linkedomics.org/login.php),
which are efficient methods to visualize large amounts of genomic
information (18). TargetScan
(http://www.targetscan.org) (19), miRTarBase (http://mirtarbase.mbc.nctu.edu.
tw/php/index.php) (20), and
miRDB (http://www.mirdb.org/) (21) were used to predict potential
targets of miR-125a-5p.
Cell culture
Human ovarian cancer cell lines, A2780 and SKOV3
(American Type Culture Collection), were cultured in RPMI-1640
medium (Sigma-Aldrich; Thermo Fisher Scientific, Inc.) supplemented
with 1%(v/v) Penicillin-Streptomycin mixture (Thermo Fisher
Scientific, Inc.) and 10% (v/v) fetal bovine serum (FBS; Gibco;
Invitrogen; Thermo Fisher Scientific, Inc.). Cells were treated
with propofol (Sigma-Aldrich; Merck KGaA; 1–10 µg/ml) for 48 h at
37°C, or were cultured at different time points (0, 12, 24, 48 h)
at 10 µg/ml. All cells were cultured in a humidified atmosphere of
5% CO2 and 95% air at 37°C.
Cell viability assays
For MTT assay, cells were seeded in 96-well
(1×103 cells/well) and treated with 0, 1, 5, 10 µg/ml of
propofol for 48 h at 37°C. A total of 5×103 cells/well
were seeded into 96-well plates and cultured for 1, 2, 3, 4 and 5
days at 37°C. Twenty microliters MTT solution (5 mg/ml,
Sigma-Aldrich; Merck KGaA) was incubated for 4 h at 37°C. Then, 150
µl DMSO was added to dissolve the precipitates and the effect of
cell number on absorbance at 490 nm was measured using a microplate
reader (Molecular Devices, LLC). The experiments were performed in
triplicate.
Wound healing assay
For wound healing assay, A2780 and SKOV3 cells were
plated in a 6-well plate (5×105 cells/well) and were
cultured to confluence as a monolayer at 5% CO2 and 95%
air at 37°C. The monolayer was scratched with a 10 µl plastic
pipette tip and washed twice with PBS to remove injured cells,
which were cultured with DMEM without serum and treated with 0, 1,
5, 10 µg/ml of propofol for 48 h at 37°C. The images of the wounded
cell monolayer were captured and evaluated using a light microscope
(magnification, ×40; Nikon Corporation). Then, the data of wound
healing area were analyzed by image analysis software (ImageJ
v1.8.0, National Institutes of Health).
Transwell migration and invasion
assays
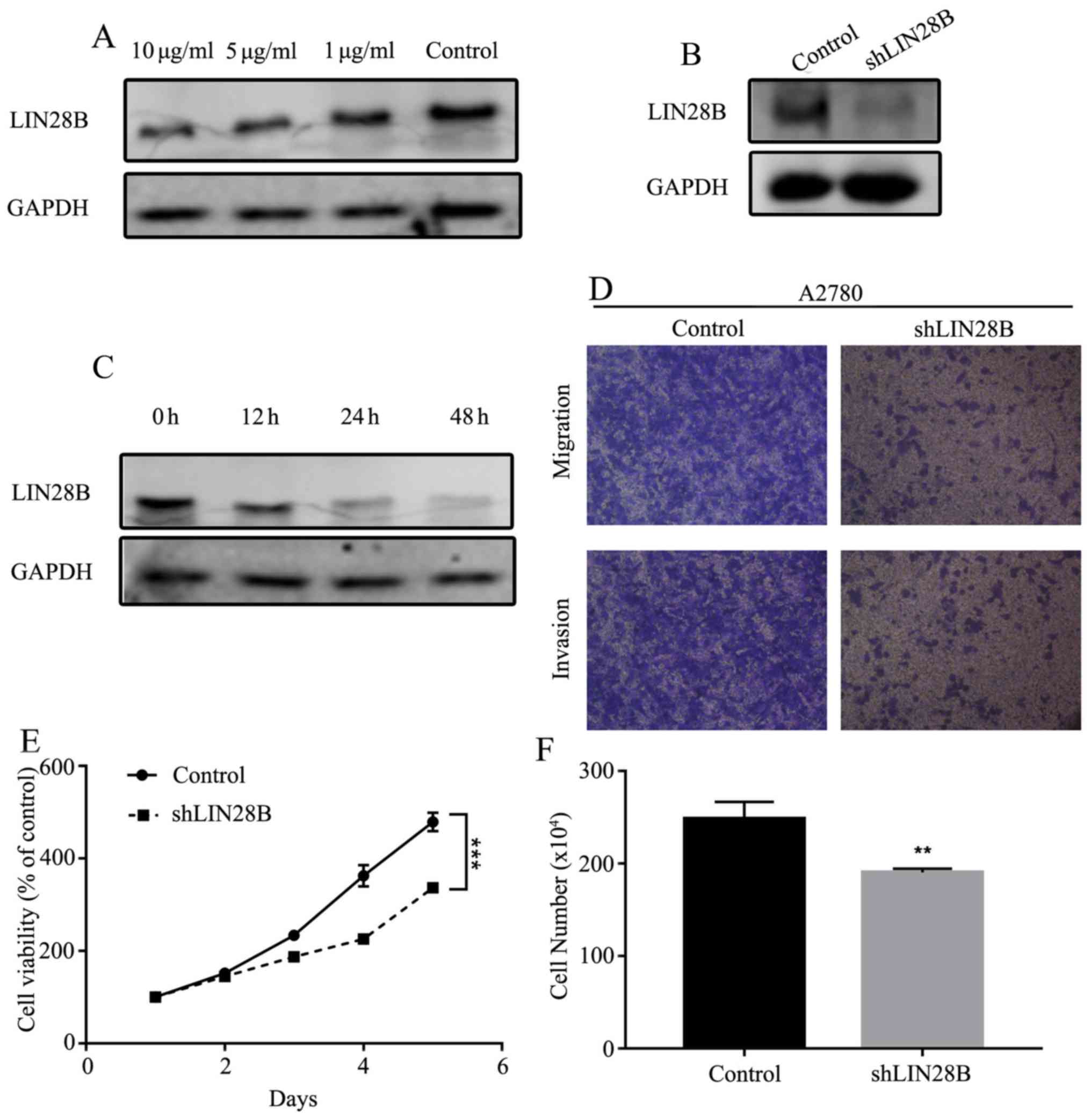
For cell transfection, LIN28B-shRNA (no. 1, no. 2;
Sigma-Aldrich; Merck KGaA) and scramble-shRNA plasmids (no 1;
Sigma-Aldrich; Merck KGaA) were transfected into A2780 cells. Cells
were transfected with 2 µg of each plasmid in six-well plates with
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.).
Following cell culture in the medium with 2 µg/ml puromycin for 72
h, cell monoclonal culture from single cell was performed. After 12
days of cell culture, the knockdown efficiency was examined by
Western blot analysis. Scramble-shRNA transfected cells were used
as the negative control for LIN28B-knockdown cells. A Transwell
assay was utilized to demonstrate the migratory and invasive
abilities of A2780 and SKOV3 cells after transfection with LIN28B
short hairpin (sh)RNA or co-incubation with propofol in 0, 1, 5 and
10 µg/ml at 37°C for 72 h. Cells were added to the Transwell
chamber (8 µm pore size; BD Biosciences). The number of cells that
migrated through the membrane was determined 8 h later. For
invasion assay, Matrigel (BD Biosciences) was mixed with 1640
medium at a ratio of 1:6 and was placed into upper chambers at 37°C
for 30 min in a 5% CO2 incubator overnight prior to
addition of cells. For migration assay, Matrigel was not used.
Then, 150 µl cell suspension (2.5×104 cells) was plated
to the upper chamber with serum-free medium and cell medium with
10% FBS was placed into lower chambers, which was incubated at 37°C
for 24 h. Migrated and invaded cells were determined by crystal
violet staining at 37°C for 15 min and were subsequently counted by
an inverted light microscope (magnification, ×40; Nikon
Corporation) using ImageJ v1.8.0 (National Institutes of
Health).
Reverse transcription-quantitative PCR
(RT-qPCR)
A2780 cells were treated with 0, 1, 5 and 10 µg/ml
of propofol for 48 h at 37°C and A2780 cells administered 10 µg/ml
propofol for 0, 12, 24 and 48 h were analyzed by RT-qPCR. Total
cellular RNA was extracted with TRIzol® (Invitrogen;
Thermo Fisher Scientific, Inc.) following the manufacturer's
instructions. Then, 1,000 ng total RNA was reverse transcribed
using TaqMan miRNA RT kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.) for 1 h at 37°C. qPCR (SYBR® Premix Ex
Taq II; Takara Biotechnology Co., Ltd.) was performed on ABI 7500
Sequence Detection system (Thermo Fisher Scientific, Inc.). The
thermocycling conditions were as follows: 2 min at 50°C and 10 min
at 95°C followed by 40 cycles of 95°C for 3 sec and 60°C for 30
sec. The expression level of miRNA was normalized by U6. The
expression level of RNA was normalized to β-actin. The following
primers were used: β-actin forward, 5′-ACCCTGAAGTACCCCATCGAG-3′ and
reverse, 5′-AGCACAGCCTGGATAGCAAC-3′; U6 forward,
5′-CTCGCTTCGGCAGCACATATACT-3′ and reverse,
5′-ACGCTTCACGAATTTGCGTGTC-3′; miR-125a-5p forward,
5′-CGATTCCCTGAGACCCTTTAA-3′ and reverse,
5′-TATGGTTTTGACGACTGTGTGAT-3′; and LIN28B forward,
5′-GTCAATACGGGTAACAGGAC-3′ and reverse, 5′-TTCTTTGGCTGAGGAGGTAG-3′
The expression level of miR-125a-5p was calculated using the
2−ΔΔCq method (22),
whereby ΔCq=Cq (miR-125a-5p)-Cq (U6). The expression level of
LIN28B was calculated using the 2−ΔΔCq method, whereby
ΔCq=Cq (LIN28B)-Cq (β-actin).
Luciferase reporter assays
Luciferase assays were used in A2780 cells.
Wild-type 3′-UTR of LIN28B and mutant controls were constructed and
inserted into the psiCheck2 Luciferase vector (Promega
Corporation). The psi-CHECK2 vector (cat. no. C8021) was purchased
from Promega Corporation. The miR-125a-5p mimics and negative
control (NC) mimics, and miR-125a-5p inhibitor and NC inhibitor
were synthesized by Guangzhou RiboBio Co., Ltd., and transfected
into 24-well plate at 50 nM using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions. Next, cells were harvested. Firefly
and Renilla luciferase activities were measured with the
dual-luciferase assay kit (Promega Corporation) at 48 h after
transfection. Then, cells were collected and lysed. The luciferase
activity was subsequently measured using a Dual Luciferase Reporter
Assay System Kit (Promega Corporation) on a Luminometer TD-20/20
detector (E5311; Promega Corporation), and the relative luciferase
activity was expressed by Firefly/Renilla luciferase
activity. The mimics sequence (5′-UCCCUGAGACCCUUUAACCUGUGA-3′),
inhibitors sequence (5′-TCACAGGUUAAAGGGTCTCAGGGA-3′), mimic NC
sequence (5′-GGACCAAATCTCGAGATTTGG-3′) and inhibitor NC sequence
(5′-UUCUCCGAACGUGUCACGUU-3′) were all designed as chemically
modified double strands.
Western blot analysis
Cells were homogenized and sonicated in RIPA buffer
(Sigma-Aldrich; Merck KGaA) on ice. Western blotting was conducted
according to our previous report (23). Determination of protein content: A
small amount of the supernatant was taken with a pipette, and the
absorbance was measured by a visible spectrophotometer at a
wavelength of 590 nm according to the Bradford method. Using
solvent as the blank control and Bovine Serum Albumin (BSA) as the
standard control a curve was drawn, and the content of the protein
in the extracted sample was estimated based on the standard curve.
The extracted protein was collected, denatured and electrophoresed
through a 10% SDS-polyacrylamide gel. Samples were loaded and
electrophoresis was performed for 60–90 min followed by transfer to
PVDF membranes and blocking in 5% skimmed milk at 37°C. After
shaking for 2 h, elution was performed. Following this, membranes
were incubated with the primary antibodies (LIN28B; Abcam, cat. no.
ab229628, 1:1,000 dilution) at 4°C overnight with shaking.
Membranes were then incubated in secondary antibodies (conjugated
goat anti rabbit IgG; CWBIO, cat. no. CW0103S, 1:2,000 dilution,)
at room temperature for 2 h and washed in TBST three times for 15
min. The Western blot analysis was detected by a chemiluminescent
method. The membranes were then incubated in Super Signal ECL-HRP
detection reagent (ComWin Biotech) for 1 min followed by exposure
to film in a visualizer.
Statistical analysis
Data are presented as the mean ± SD. All assays were
tested in three independent experiments. Experimental data were
analyzed using SPSS statistical software (version 12.0; SPSS Inc.).
The significance of the group difference was evaluated by one-way
analysis of variance followed by Tukey's post hoc test. Multiple
comparisons between the groups were performed using
Student-Newman-Keuls method. P<0.05 was considered to indicate a
statistically significant difference.
Results
Propofol suppresses the proliferation
of ovarian cancer cells
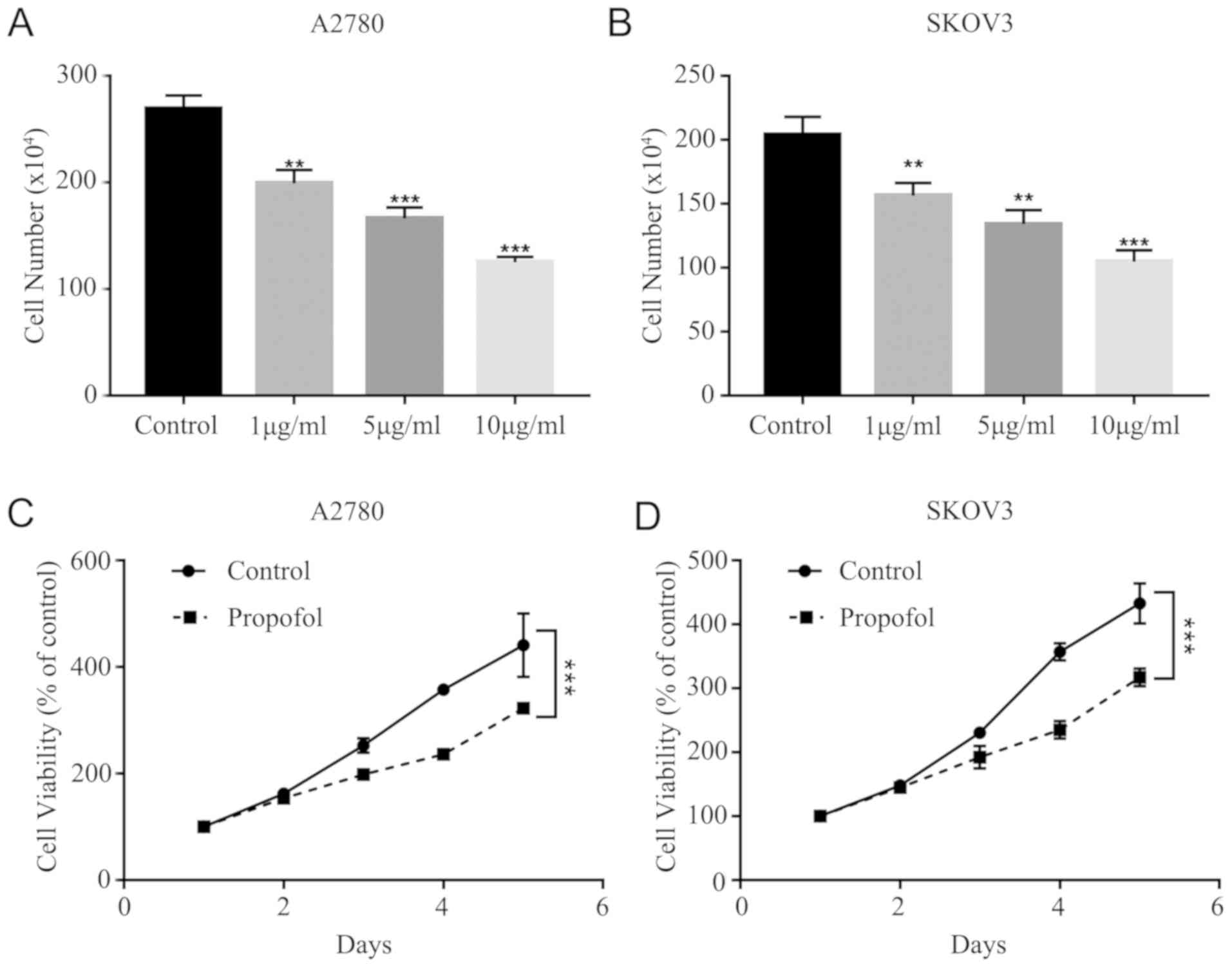
To assess whether the effects of 1, 5 and 10 µg/ml
propofol suppress cell viability, the cytotoxicity of propofol was
tested in A2780 and SKOV3 cells. It was demonstrated that propofol
decreased A2780 and SKOV3 cell viability in a dose-dependent manner
(Fig. 1A and B). It was found that
10 µg/ml propofol had the strongest inhibitory effect on A2780 and
SKOV3 cells. Furthermore, the MTT assay results indicated that
propofol treatment (10 µg/ml) significantly inhibited the
proliferation of A2780 and SKOV3 cells (Fig. 1C and D).
Migration and invasion of ovarian
cancer cells are attenuated by propofol
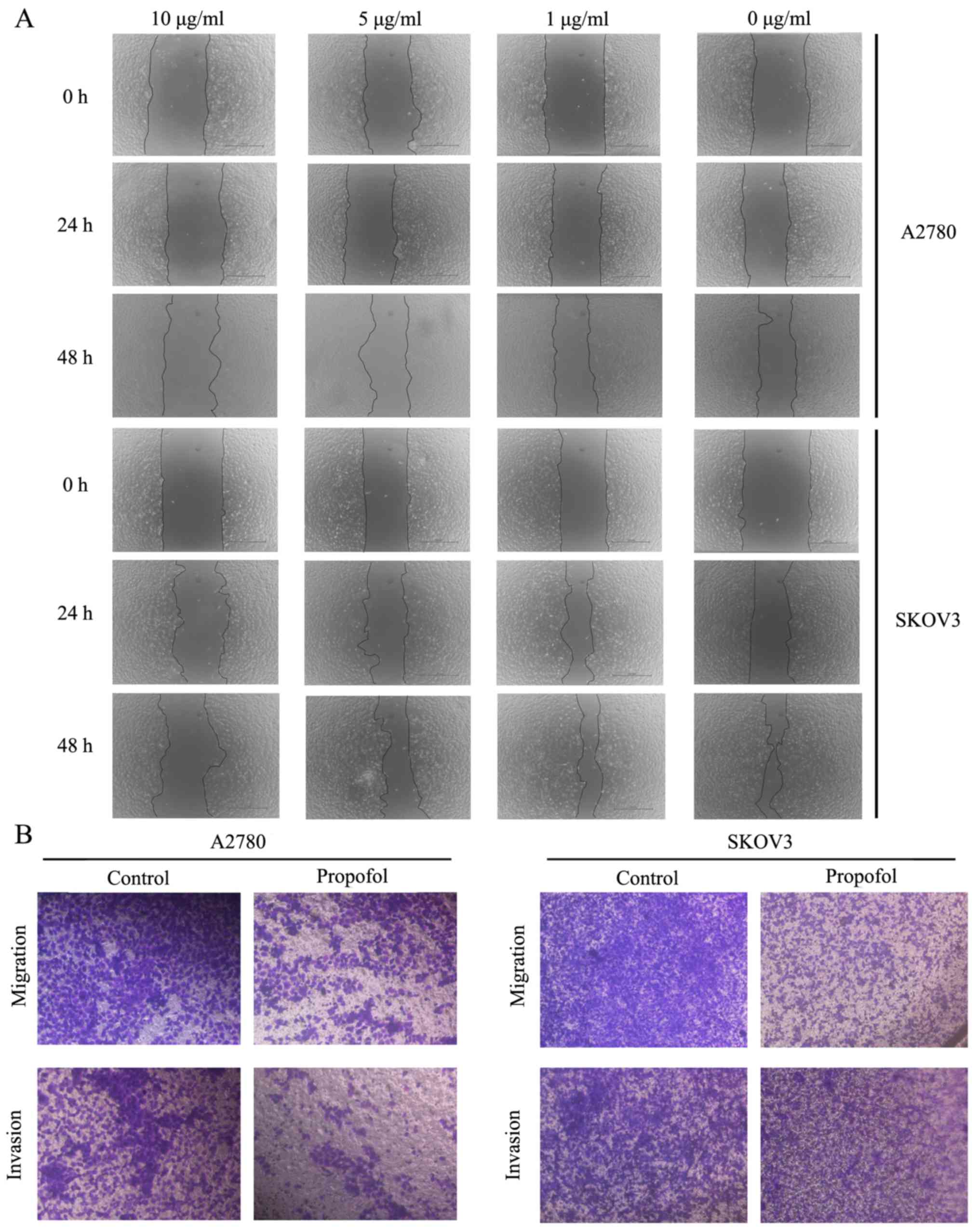
Subsequently, the effects of propofol on A2780 and
SKOV3 cell migratory and invasive abilities were examined. The
wound healing assay results suggested that treatment with 10 µg/ml
propofol markedly inhibited the migration of A2780 and SKOV3 cells
compared to NC group (Fig. 2A). To
further demonstrate the suppressive effects of propofol on cell
migration and invasion, Transwell assays were performed. These
findings also indicated that propofol notably suppressed the
migration and invasion of ovarian cancer cells (Fig. 2B).
Bioinformatics analysis of women with
ovarian cancer
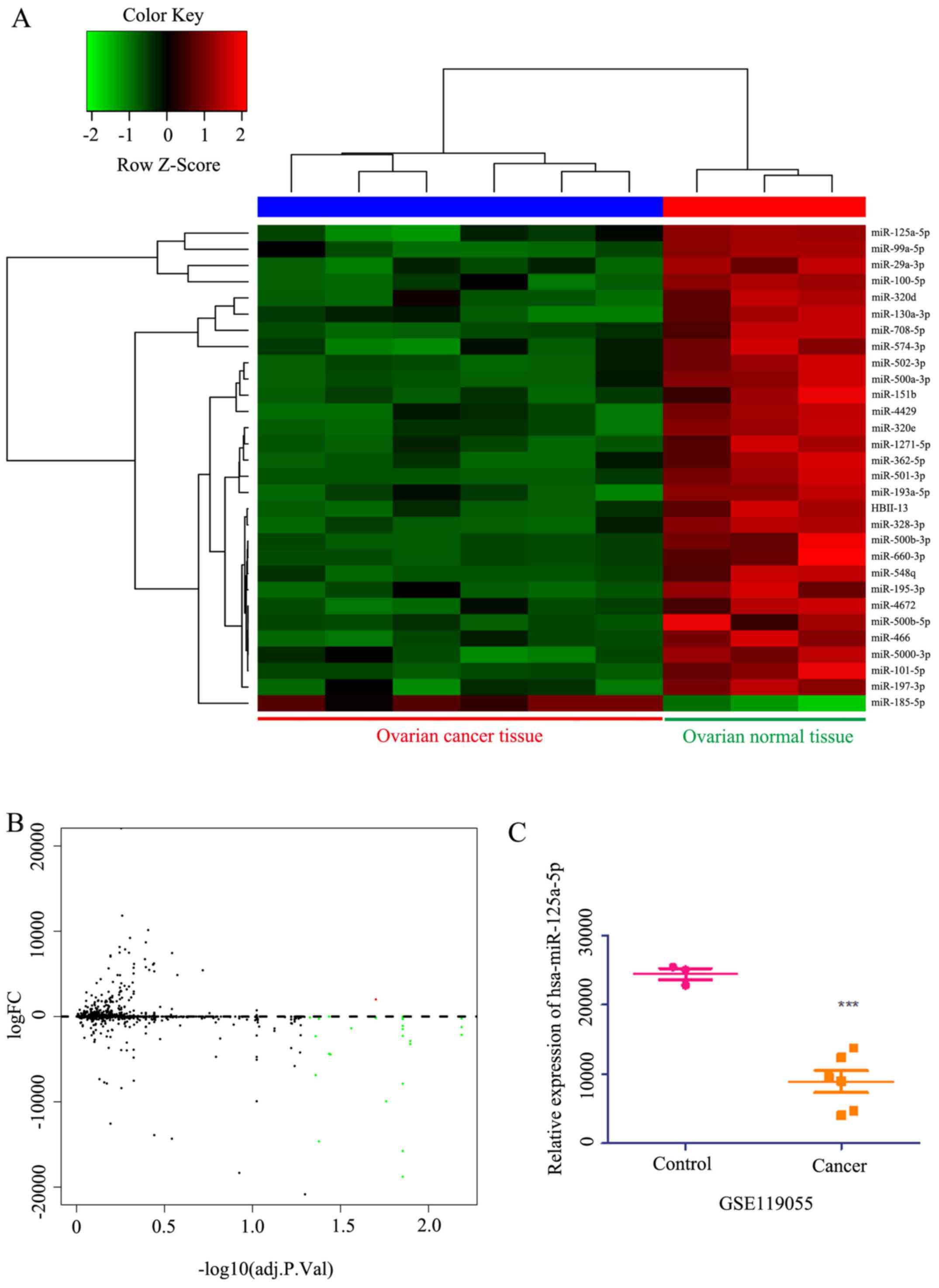
miRNA expression profiles were downloaded from the
Gene Expression Omnibus database (ID, GSE119055) to analyze
differentially expressed miRNAs. This analysis identified 45
microRNAs with differential expression in ovarian cancer compared
with healthy ovarian tissue (Fig. 3A
and B). Among these, the expression of miR-125a-5p was lower in
patients with ovarian cancer compared with healthy ovarian tissue
(Fig. 3C). Within the top 30
candidate miRNAs analyzed, it was found that the expression of
miR-125a-5p was lower in cancer tissues compared with healthy
tissues (Fig. 3A). Collectively,
these results indicated that miR-125a-5p expression was lower in
ovarian cancer, resulting in a poor prognosis.
GO analysis and GSEA of co-expressed
genes correlated with miR-125a-5p in ovarian cancer
The present study analyzed mRNA sequencing data from
453 patients with ovarian cancer in The Cancer Genome Atlas (TCGA).
As shown in the heat map and volcano plot (Fig. 4A and B), 3,395 genes (dark red
dots) were significantly upregulated along with miR-125a-5p, while
3,031 genes (dark green dots) were significantly downregulated
(false discovery rate<0.01). In total, 50 hub genes were
positively and negatively associated with miR-125a-5p, as presented
in the heat map (Fig. 4A). These
results indicated a significant and wide pathophysiological effect
of miR-125a-5p on gene transcription. The list of co-expressed
genes associated with miR-125a-5p contained 6,426 gene IDs of which
5,188 gene IDs were unambiguously mapped to 5,188 unique Entrez
gene IDs and 1,238 user IDs could not be mapped to any Entrez gene
ID.
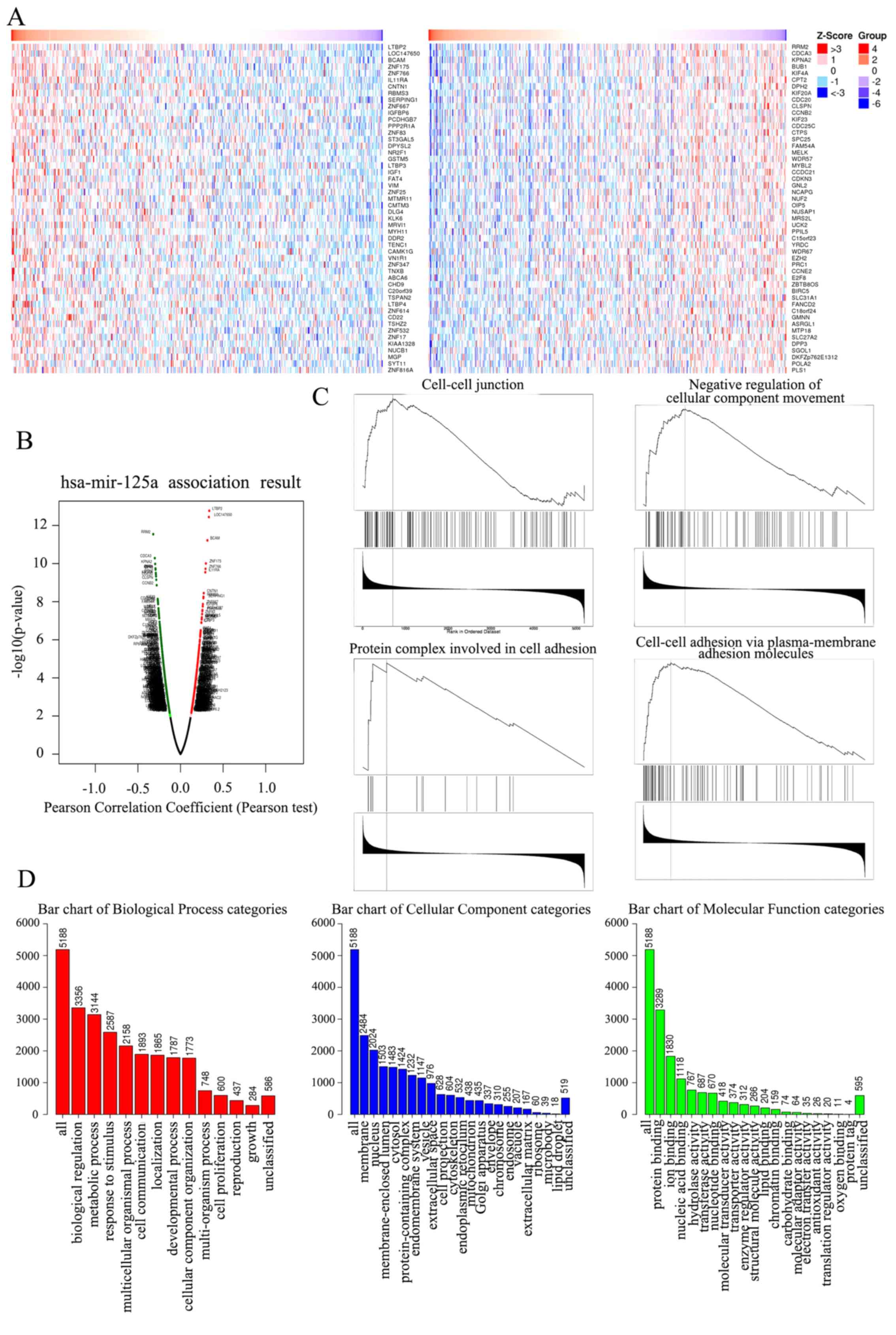 | Figure 4.Heat map, Volcano plot, GSEA and GO
enrichment assay in ovarian of key genes targeted by miR-125a-5p.
(A) Heat map of the positive DEGs (left) and negative DEGs (right)
targeted by miR-125a-5p in TCGA database. (B) Volcano plot of the
DEGs targeted by miR-125a-5p in TCGA database, the red dots and
green dots represent the upregulated and downregulated,
respectively. (C) The four enrichment plots from the GSEA results,
including ‘cell-cell junction’, ‘negative regulation of cellular
component movement’, ‘protein complex involved in cell adhesion’
and ‘cell-cell adhesion via plasma-membrane adhesion molecules’.
(D) GO analysis results of DEGs. GO, Gene Ontology; miR, microRNA;
DEGs, differentially expressed genes; GSEA, Gene Set Enrichment
Analysis; TCGA, The Cancer Genome Atlas. |
The GO Slim summary indicated a notable quantity
variance and significance level difference among the 5,188
differentially expressed genes (DEGs) that were enriched in
biological processes, molecular functions and cellular components.
For biological processes, these 5,188 genes regulated by
miR-125a-5p were mainly enriched in ‘biological regulation’,
including ‘cell proliferation’, ‘growth’, ‘metabolic process’ and
‘cell communication’, thus suggesting that miR-125a-5p could act as
an anticarcinogen to inhibit ovarian cancer by mediating biological
regulation. For cellular components, the genes were largely
enriched in the ‘membrane’, including ‘cell projection’,
‘cytoskeleton’ and ‘extracellular matrix’, indicating that
miR-125a-5p could regulate membrane components to control cell
movement and organization. For molecular functions, the genes were
mainly enriched in ‘protein binding’ and ‘enzyme activity’,
demonstrating that miR-125a-5p could act as a regulator to mediate
ovarian cancer progression (Fig.
4D).
GSEA of the 5,188 DEGs at the whole gene expression
level was performed, and was used to assess the cell migration and
invasion effects of DEGs that played roles in the whole gene
expression level. The results demonstrated that the 5,188 DEGs
regulated by miR-125a-5p were largely enriched in pathways
including ‘cell-cell junction’, ‘negative regulation of cellular
component movement'’, ‘protein complex involved in cell adhesion’
and ‘cell-cell adhesion via plasma-membrane adhesion molecules’,
which indicated that miR-125a-5p could mediate cell migration and
invasion in ovarian cancer (Fig.
4C).
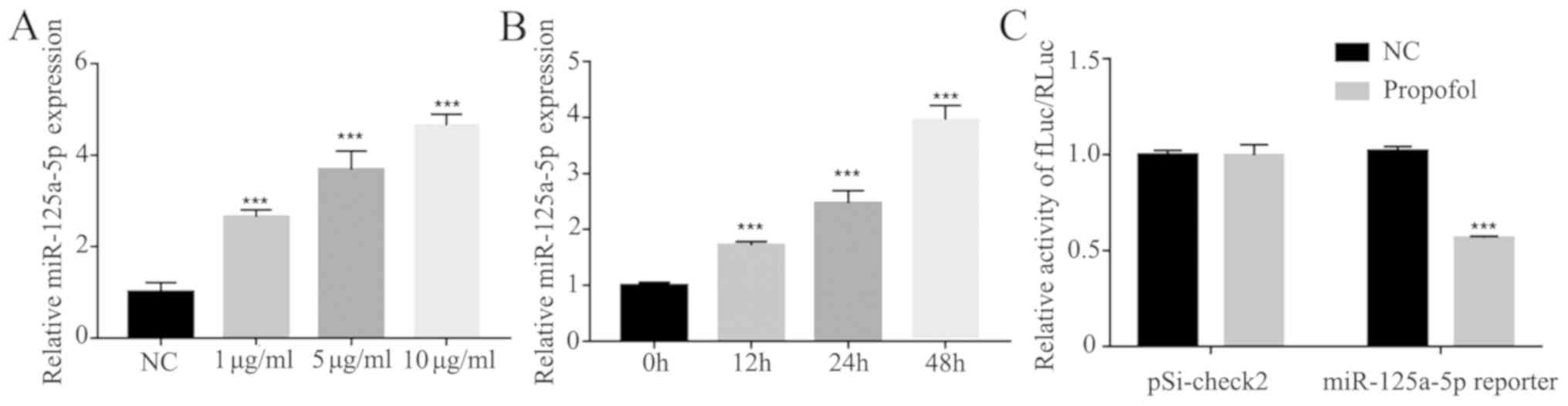
Expression of miR-125a-5p is enhanced
by propofol in ovarian cancer cells
It has been revealed that propofol can upregulate
the expression levels of miR-199a (24) and miR-143 (25), suggesting that propofol may
regulate a number of miRNAs. Therefore, the present study detected
the effects of propofol on miR-125a-5p involved in ovarian cancer
cells. The expression of miR-125a-5p was measured under different
concentrations of propofol (1, 5 and 10 µg/ml), and it was found
that the expression of miR-125a-5p was enhanced in a dose- and
time-dependent manner (Fig. 5A and
B). Furthermore, the positive effect of propofol on miR-125a-5p
transcriptional activity was further tested using a luciferase
assay and it was found that propofol could significantly increase
the transcriptional activity of miR-125a-5p compared to NC group
(Fig. 5C).
miR-125a-5p targets LIN28B to
downregulate ovarian cancer cell proliferation
TargetScan (http://www.targetscan.org), miRTarBase (http://mirtarbase.mbc.nctu.edu.
tw/php/index.php), miRDB (http://www.mirdb.org/) and TCGA (https://cancergenome.nih.gov/) were used to predict
potential targets of miR-125a-5p (Fig.
6A). A variety of cell proliferation-, migration- and
invasion-related genes were predicted, and the level of Lin28B in
these genes was subsequently assessed by RT-qPCR. It was
demonstrated that LIN28B mRNA expression was significantly
inhibited by miR-125a-5p mimics in A2780 and SKOV3 cells (Fig. 6B). In both ovarian cancer cell
lines, miR-125a-5p mimics also significantly decreased the protein
expression of LIN28B compared with the controls (Fig. 6C).
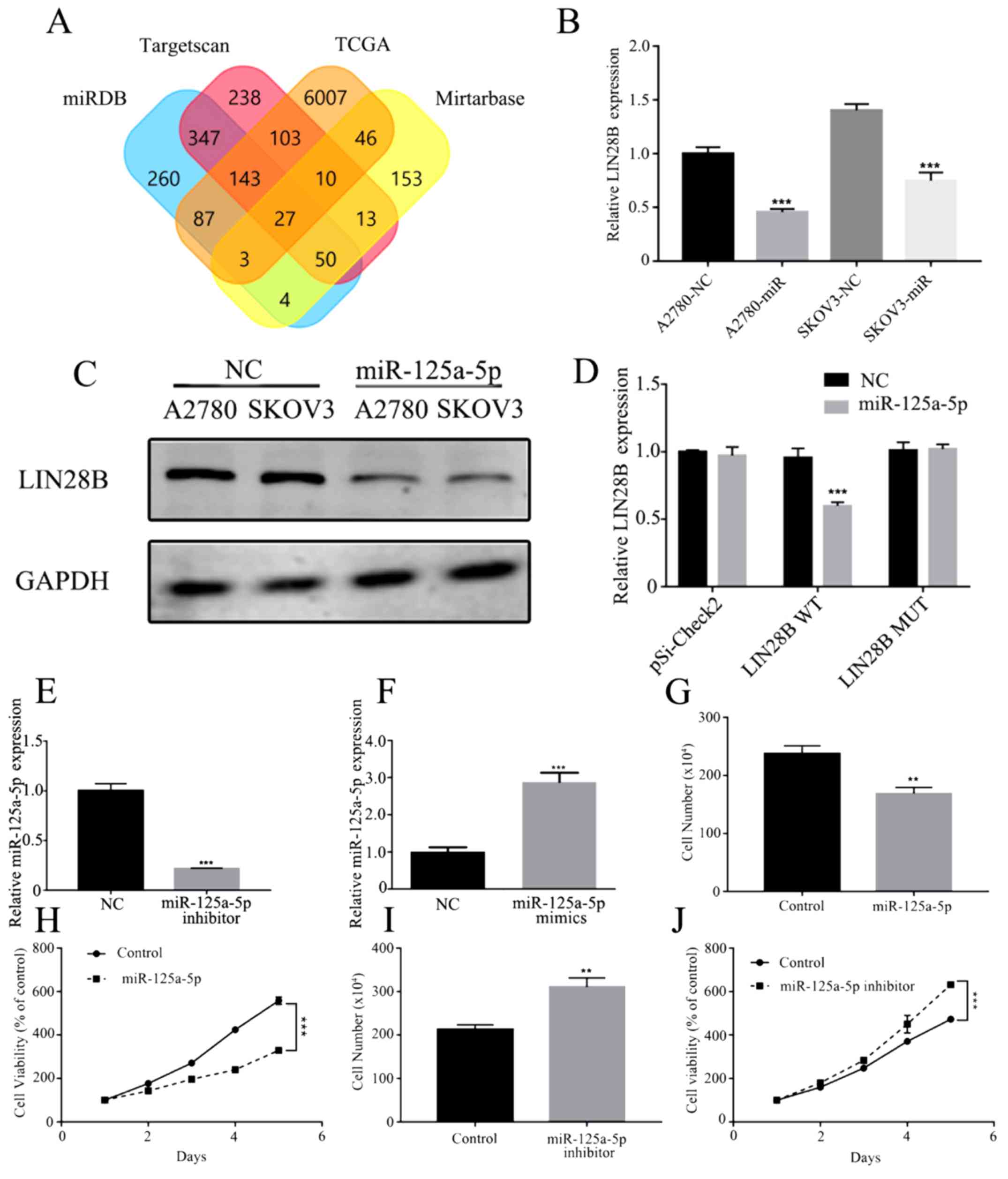 | Figure 6.miR-125a-5p directly targets LIN28B
and repress proliferation ability in ovarian cancer. (A) Prediction
potential targets of miR-125a-5p assessed by TargetScan,
miRTarBase, miRDB and TCGA. (B) mRNA expression of LIN28B regulated
by miR-125a-5p in A2780 and SKOV3 cells was measured using RT-qPCR.
(C) Expression of LIN28B in A2780 and SKOV3 cells transfected with
miR-125a-5p were detected by Western blotting. (D) Luciferase
reporter activities driven by wild-type or MUT LIN28B 3’
untranslated regions were examined in A2780 cells transfected with
miR-125a-5p mimics or NC. (E) Expression of miR-125a-5p in A2780
cells transfected with miR-125a-5p inhibitor was evaluated by
RT-qPCR. (F) miR-125a-5p expression in A2780 cells transfected with
miR-125a-5p mimics was measured by RT-qPCR. (G) Proliferation of
A2780 cells transfected with miR-125a-5p was assessed by cell
counts assay. (H) Viability of A2780 cells transfected with
miR-125a-5p was tested by MTT assay. (I) Proliferation ability of
A2780 cells transfected with miR-125a-5p inhibitor was determined
by cell counts assay. (J) Viability of A2780 cells transfected with
miR-125a-5p inhibitor was tested by MTT assay. Data are presented
as the mean ± SD. **P<0.01, ***P<0.001 vs. control. miR,
microRNA; MUT, mutant; RT-qPCR, reverse transcription-quantitative
PCR; NC, negative control; LIN28B, lin-28 homolog B; TCGA, TCGA,
The Cancer Genome Atlas. |
To identify the direct action of miR-125a-5p on
LIN28B a luciferase assay was performed. miR-125a-5p significantly
suppressed the expression of LIN28B, while LIN28B with mutant
binding sites was not regulated by miR-125a-5p mimics (Fig. 6D). Furthermore, A2780 cells were
transfected with miR-125a-5p inhibitor and mimics, which
significantly affected the expression of miR-125a-5p (Fig. 6E and F). Cellular proliferation and
viability were also inhibited by the miR-125a-5p mimic group
compared with the NC group (Fig. 6G
and H). It was found that inhibition of miR-125a-5p could
significantly enhance cell proliferation and viability compared
with the control group (Fig. 6I and
J).
Expression of LIN28B is decreased by
propofol in ovarian cancer cells
To determine whether the expression of LIN28B could
be affected by propofol, the expression of LIN28B was measured
under different concentrations of propofol (1, 5 and 10 µg/ml). The
results indicated that the expression of LIN28B was inhibited in a
dose- and time-dependent manner (Fig.
7A and C). Mechanistically, it was identified that LIN28B
knockdown (KD; Fig. 7B) inhibited
cellular migration and invasion (Fig.
7D). Furthermore, cell viability and numbers could be
suppressed by LIN28B KD compared with the control cell line
(Fig. 7E and F).
Discussion
The present study evaluated the effects of propofol
on ovarian cancer A2780 and SKOV3 cells. It was found that exposure
to propofol markedly inhibited proliferation, migration and
invasion in ovarian cancer cells. The underlying mechanisms were
investigated, and several miRNAs related to cell migration and
invasion were detected. After propofol treatment, the expression of
miR-125a-5p was significantly increased in a dose- and
time-dependent manner in A2780 and SKOV3 cells. Previous studies
have reported that propofol can upregulate the expression levels of
miR-199a (24) and miR-143
(25), indicating that propofol
can regulate miRNAs involved in cell migration and invasion. The
present results also identified that LIN28B was a target of
miR-125a-5p. Thus, it was speculated that propofol serves crucial
roles in the inhibition of ovarian cancer cell proliferation,
migration and invasion by enhancing miR-125a-5p, which targets
LIN28B. Moreover, KD of LIN28B markedly enhanced cellular migration
and invasion compared with the control cell line.
Ovarian cancer is one of the most fatal gynecologic
tumor types (26). Currently,
therapeutic methods involve multiple approaches that include
debulking surgery, chemotherapy and radiotherapy in ovarian cancer,
but the curative treatment is surgical resection (27). Anesthesia agents must be used
during the surgical processes. However, the role that anesthetics
serve in the development of cancer is not fully understood.
Propofol, a commonly used intravenous sedative-hypnotic agent
administered to induce and maintain anesthesia, has been used since
the late 1980s (5). Propofol not
only has an anesthetic effect but has also been reported to possess
antioxidant, neuroprotective, immunomodulatory, analgesic,
antiemetic and anticancer effects (6,28,29).
Previous studies have also reported that propofol can suppress
tumor growth (25,30,31).
Another study revealed that propofol increased the expression of
caspase-3 in A549 cells and LoVo cells to induce cell apoptosis
(32). Du et al (8) also showed that propofol inhibited
cell proliferation, migration and invasion, and induced apoptosis
by regulating Sox4 in endometrial cancer cells. Moreover, Peng and
Zhang (33) reported that propofol
possessed anti-proliferative and pro-apoptotic properties in
gastric cancer via the upregulation of miR-451. Propofol also has
an inhibitory role in A549 lung cancer cells by downregulating
miR-372 and inactivating the mTOR and Wnt/β-catenin pathways
(34). Propofol inhibits
proliferation, migration and invasion, and induces apoptosis in
gastric cancer cells by upregulating miR-195 and miR-451 (33,35).
Cellular proliferation can also be inhibited by propofol via its
regulation of several miRNAs, including miR-372, miR-1284 and
miR-486, in lung cancer cells (7,36).
Overall, propofol inhibits cell viability functions in numerous
cancer types. Huang et al (37) revealed that propofol upregulated
miR-9 expression and led to the inhibition of cell proliferation
and invasion in ovarian cancer ES-2 cells. Moreover, Sun et
al (31) found that propofol
could decrease miR-374a expression to increase the expression of
forkhead box O1, resulting in inhibition of cisplatin resistance
and proliferation. The present study evaluated the effects of
different concentrations of propofol on two ovarian cell lines. It
was identified that propofol had inhibitory effects on cell
viability, migration and invasion in a dose- and time-dependent
manner. Moreover, the present results are consistent with the
aforementioned previous studies, suggesting that propofol can
suppress tumors.
miRNAs, which are non-coding RNAs consisting of
18–25 nucleotides, act as oncogenes or tumor suppressors and can be
used as potential biological markers to diagnose, predict and treat
different cancer types (38).
Previous studies have reported that several miRNAs are regulated by
propofol (25,31). The present study analyzed mRNA
sequencing data from 453 patients with ovarian cancer in TCGA. For
biological processes, these 5,188 genes regulated by miR-125a-5p
were mainly enriched in ‘biological regulation’, including ‘cell
proliferation’, ‘growth’, ‘metabolic process’ and ‘cell
communication’, indicating that miR-125a-5p could act as an
anticarcinogen to inhibit ovarian cancer by mediating biological
regulation. The expression of miR-125a-5p was measured under
several concentrations of propofol, and the results indicated that
the expression of miR-125a-5p was enhanced in a dose- and
time-dependent manner. Furthermore, the positive effect of propofol
on miR-125a-5p transcriptional activity was further identified by a
luciferase assay. Wu and Belasco (39) revealed the human LIN28 mRNA as a
regulatory target of human miR-125b and its homologue miR-125a
during neuronal differentiation of embryonal carcinoma cells.
Moreover, Zhang et al (11)
showed that miR-125a-5p directly targeted LIN28B and suppressed
melanoma growth. In patients with colon cancer, it has been
revealed that propofol anesthesia is associated with improved
survival during colon cancer surgery (40). However, another study suggested
that the prognosis and survival of patients treated with propofol
and desflurane were not significantly different after breast cancer
surgery (30). However, these
conflicting results could be attributed to the different types of
cancer or to different effects of propofol in vitro and
in vivo. The present results demonstrated a significant
decrease in LIN28B expression with propofol in a dose- and
time-dependent manner. Further investigations revealed that LIN28B
KD significantly inhibited cell proliferation and viability. LIN28B
was also identified as a direct target of miR-125a-5p, and propofol
treatment reduced LIN28B expression. Thus, the results indicated
that LIN28B KD led to significantly decreased cell proliferation
and viability in ovarian cancer.
In conclusion, the results demonstrated that
propofol plays an important role in inhibiting the proliferation,
migration and invasion of ovarian cancer cells. Propofol also
upregulates the expression of miR-125a-5p to inhibit LIN28B.
Therefore, propofol in the surgical treatment of ovarian cancer may
be a potential target for improving cancer prognosis and survival
(24,31,41).
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
Conception and design: JZe, JZo and YKL. Acquisition
and assembly of data: WJT, XZ, FFQ, CYC and TZ. Data analysis and
interpretation: CYC, FFQ and XZ. Manuscript writing: JZe, WJT, YKL
and FFQ. Paper revision: JZo and WJT. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D, Jemal A and
Ahmedin J: Cancer statistics, 2013. CA Cancer J Clin. 63:11–30.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arnold M, Rutherford MJ, Bardot A, Ferlay
J, Andersson TM, Myklebust TÅ, Tervonen H, Thursfield V, Ransom D,
Shack L, et al: Progress in cancer survival, mortality, and
incidence in seven high-income countries 1995-2014 (ICBP
SURVMARK-2): A population-based study. Lancet Oncol. 20:1493–1505.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Coleman RL, Fleming GF, Brady MF, Swisher
EM, Steffensen KD, Friedlander M, Okamoto A, Moore KN, Efrat
Ben-Baruch N, Werner TL, et al: Veliparib with First-Line
Chemotherapy and as Maintenance Therapy in Ovarian Cancer. N Engl J
Med. 381:2403–2415. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hsing CH, Lin MC, Choi PC, Huang WC, Kai
JI, Tsai CC, Cheng YL, Hsieh CY, Wang CY, Chang YP, et al:
Anesthetic propofol reduces endotoxic inflammation by inhibiting
reactive oxygen species-regulated Akt/IKKβ/NF-κB signaling. PLoS
One. 6:e175982011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Vasileiou I, Xanthos T, Koudouna E, Perrea
D, Klonaris C, Katsargyris A and Papadimitriou L: Propofol: A
review of its non-anaesthetic effects. Eur J Pharmacol. 605:1–8.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu WZ and Liu N: Propofol inhibits lung
cancer A549 cells growth and epithelial-mesenchymal transition
process by up-regulation of microRNA-1284. Oncol Res. 27:1–8. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Du Q, Liu J, Zhang X, Zhang X, Zhu H, Wei
M and Wang S: Propofol inhibits proliferation, migration, and
invasion but promotes apoptosis by regulation of Sox4 in
endometrial cancer cells. Braz J Med Biol Res. 51:e68032018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang LB, Feng L, He J, Liu B and Sun JG:
MiR-125a-5p inhibits the proliferation and invasion of breast
cancer cells and induces apoptosis by targeting GAB2. Math Biosci
Eng. 16:6923–6933. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cao Y, Shen T, Zhang C, Zhang QH and Zhang
ZQ: MiR-125a-5p inhibits EMT of ovarian cancer cells by regulating
TAZ/EGFR signaling pathway. Eur Rev Med Pharmacol Sci.
23:8249–8256. 2019.PubMed/NCBI
|
|
11
|
Zhang Z, Zhang S, Ma P, Jing Y, Peng H,
Gao WQ and Zhuang G: Lin28B promotes melanoma growth by mediating a
microRNA regulatory circuit. Carcinogenesis. 36:937–945. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yong W, Yu D, Jun Z, Yachen D, Weiwei W,
Midie X, Xingzhu J and Xiaohua W: Long noncoding RNA NEAT1,
regulated by LIN28B, promotes cell proliferation and migration
through sponging miR-506 in high-grade serous ovarian cancer. Cell
Death Dis. 9:8612018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou J, Ng SB and Chng WJ: LIN28/LIN28B:
An emerging oncogenic driver in cancer stem cells. Int J Biochem
Cell Biol. 45:973–978. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Huang J, Cao D, Sha J, Zhu X and Han S:
DLL3 is regulated by LIN28B and miR-518d-5p and regulates cell
proliferation, migration and chemotherapy response in advanced
small cell lung cancer. Biochem Biophys Res Commun. 514:853–860.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang C, Gu Y, Zhang E, Zhang E, Zhang K,
Na Qin N, Dai J, Zhu M, Liu J, Xie K, et al: A cancer-testis
non-coding RNA LIN28B-AS1 activates driver gene LIN28B by
interacting with IGF2BP1 in lung adenocarcinoma. Oncogene.
38:1611–1624. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dong S, Wang R, Wang H, Ding Q, Zhou X,
Wang J, Zhang K, Long Y, Lu S, Hong T, et al: HOXD-AS1 promotes the
epithelial to mesenchymal transition of ovarian cancer cells by
regulating miR-186-5p and PIK3R3. J Exp Clin Cancer Res.
38:1102019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
R Core Team, . R: A language and
environment for statistical computing. R Foundation for Statistical
Computing, Vienna, 2014.
|
|
18
|
Vasaikar SV, Straub P, Wang J and Zhang B:
LinkedOmics: Analyzing multi-omics data within and across 32 cancer
types. Nucleic Acids Res. 46:D956–D963. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Agarwal V, Bell GW, Jin-Wu Nam JW and
Bartel DP: Predicting effective microRNA target sites in mammalian
mRNAs. Elife. 4:e050052015. View Article : Google Scholar
|
|
20
|
Chou CH, Shrestha S, Yang CD, Chang NW,
Lin YL, Liao KW, Huang WC, Sun TH, Tu SJ, Lee WH, et al: miRTarBase
update 2018: A resource for experimentally validated
microRNA-target interactions. Nucleic Acids Res. 46:D296–D302.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Y and Wang X: miRDB: An online
database for prediction of functional microRNA targets. Nucleic
Acids Res. 48:D127–D131. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zou J, Li H, Huang Q, Liu X, Qi X, Wang Y,
Lu L and Liu Z: Dopamine-induced SULT1A3/4 promotes EMT and cancer
stemness in hepatocellular carcinoma. Tumour Biol.
39:10104283177192722017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang J, Zhang D, Wu GQ, Feng ZY and Zhu
SM: Propofol inhibits the adhesion of hepatocellular carcinoma
cells by upregulating microRNA-199a and downregulating MMP-9
expression. Hepatobiliary Pancreat Dis Int. 12:305–309. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ye Z, Jingzhong L, Yangbo L, Lei C and
Jiandong Y: Propofol inhibits proliferation and invasion of
osteosarcoma cells by regulation of microRNA-143 expression. Oncol
Res. 21:201–207. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wijnen JA and Rosenshein NB: Surgery in
ovarian cancer. Arch Surg. 115:863–868. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park TW and Kuhn WC: Neoadjuvant
chemotherapy in ovarian cancer. Expert Rev Anticancer Ther.
4:639–647. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Z, Zang M, Wang S and Wang C:
Effects of propofol on human cholangiocarcinoma and the associated
mechanisms. Exp Ther Med. 17:472–478. 2019.PubMed/NCBI
|
|
29
|
Wang J, Cheng CS, Lu Y, Ding X, Zhu M,
Miao C and Chen J: Novel findings of anti-cancer property of
propofol. Anticancer Agents Med Chem. 18:156–165. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang YH, Lee MS, Lou YS, Lai HC, Yu JC,
Lu CH, Wong CS and Wu ZF: Propofol-based total intravenous
anesthesia did not improve survival compared to desflurane
anesthesia in breast cancer surgery. PLoS One. 14:e02247282019.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun Y, Peng YB, Ye LL, Ma LX, Zou MY and
Cheng ZG: Propofol inhibits proliferation and cisplatin resistance
in ovarian cancer cells through regulating the microRNA
374a/forkhead box O1 signaling axis. Mol Med Rep. 21:1471–1480.
2020.PubMed/NCBI
|
|
32
|
Song J, Shen Y, Zhang J and Lian Q: Mini
profile of potential anticancer properties of propofol. PLoS One.
9:e1144402014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Peng Z and Zhang Y: Propofol inhibits
proliferation and accelerates apoptosis of human gastric cancer
cells by regulation of microRNA-451 and MMP-2 expression. Genet Mol
Res. 15:152016. View Article : Google Scholar
|
|
34
|
Sun H and Gao D: Propofol suppresses
growth, migration and invasion of A549 cells by down-regulation of
miR-372. BMC Cancer. 18:12522018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang W, Wang Y, Zhu Z, Zheng Y and Song
B: Propofol inhibits proliferation, migration and invasion of
gastric cancer cells by up-regulating microRNA-195. Int J Biol
Macromol. 120:975–984. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang N, Liang Y, Yang P, Yang T and Jiang
L: Propofol inhibits lung cancer cell viability and induces cell
apoptosis by upregulating microRNA-486 expression. Braz J Med Biol
Res. 50:e57942017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huang X, Teng Y, Yang H and Ma J: Propofol
inhibits invasion and growth of ovarian cancer cells via regulating
miR-9/NF-κB signal. Braz J Med Biol Res. 49:e57172016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
McManus MT: MicroRNAs and cancer. Semin
Cancer Biol. 13:253–258. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wu L and Belasco JG: Micro-RNA regulation
of the mammalian lin-28 gene during neuronal differentiation of
embryonal carcinoma cells. Mol Cell Biol. 25:9198–9208. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu ZF, Lee MS, Wong CS, Lu CH, Huang YS,
Lin KT, Lou YS, Lin C, Chang YC and Lai HC: Propofol-based total
intravenous anesthesia is associated with better survival than
desflurane anesthesia in colon cancer surgery. Anesthesiology.
129:932–941. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ikeda T, Kurasako N, Nishitani K, Okada S
and Arai T: Anesthetic management for removal of a giant ovarian
tumor using FloTrac X Vigileo monitoring system. Masui. 63:439–442.
2014.(In Japanese). PubMed/NCBI
|