Introduction
Breast cancer is the most common type of cancer in
women worldwide, and is the second leading cause of
cancer-associated mortality (1–3).
Triple negative breast cancer (TNBC) is a breast cancer subtype
associated with high rates of metastasis, heterogeneity, drug
resistance and a poor prognosis. The absence of estrogen and
progesterone receptors, and low expression of Her-2/neu
characterizes TNBC. Moreover, the incidence of TNBC is continuously
increasing in young women (4–6).
Extracellular vesicles (EVs) are vesicles derived
from membranes that are secreted by healthy and cancer cells, and
mediate a variety of biological functions. EVs are classified into
two groups according to their origin and size: Exosomes (30–100 nm)
and microvesicles (100-1,000 nm) (7). Exosomes are a homogeneous group of
vesicles that originate from multivesicular bodies, while
microvesicles are a heterogeneous population derived from the
plasma membrane during membrane blebbing (8). Furthermore, EVs derived from tumor
cells express phosphatidylserine (PS) on their membrane surfaces,
and these EVs express higher levels of PS compared with EVs from
healthy cells (9–11). Therefore, the quantification of PS
in tumor-derived EVs is proposed as a biomarker of tumor
progression (12,13).
In cancer, cells are able to transfer nucleic acids,
chemokine receptors, growth factor receptors and functional
transcription factors via the secretion and uptake of EVs (14). Moreover, cells associated with
stroma and tumor cells release EVs that mediate a variety of
processes related to cancer progression, including migration,
invasion, angiogenesis, evasion of immune response, resistance to
drugs and extracellular matrix (ECM) degradation (14–16).
EVs are present in human fluids, including lymph, urine, breast
milk, saliva, amniotic fluid, blood and malignant ascites. In
addition, in patients with gastric or breast cancer, the number of
EVs circulating is higher compared with healthy controls, and these
have been associated with poor prognosis (8,17–19).
Matrix metalloproteinases (MMPs) are a family of
proteases that are categorized into different types based on their
substrate specificity and sequence characteristics (20,21).
Furthermore, MMPs are endopeptidases, which are zinc-dependent and
are able to degrade all ECM components. In cancer, MMPs participate
in tumor progression by mediating expansion, angiogenesis and
invasion via the basement membrane (BM) and interstitial matrices
(20). Moreover, cancer
progression is associated with upregulation and secretion of MMP-2
(gelatinase A) and MMP-9 (gelatinase B), as malignant tumors
overexpress these MMPs, which degrade type IV collagen, which is
the most abundant component of BM (20,22,23).
Focal adhesions are the structures on which integrin
receptors mediate the adhesion between the actin cytoskeleton and
ECM, which are important for a variety of cellular components,
including scaffolding proteins, GTPases, phosphatases and kinases
(24,25). Moreover, one component of focal
adhesions is the focal adhesion kinase (FAK), which is a 125 kDa
protein tyrosine kinase that is activated by a variety of agonists,
including free fatty acids (26–28).
FAK has been implicated in the regulation of cell spreading,
differentiation, proliferation, apoptosis, migration, invasion,
survival and angiogenesis (29–31).
Furthermore, activation of FAK occurs via its autophosphorylation
at tyrosine (Tyr)-397, which creates a binding site for the Src
kinase and other downstream effectors. Moreover, FAK-Src complex
formation is mediated via the Src Homology 2 (SH2) domain of Src
and its formation leads to Src activation. Subsequently, Src
phosphorylates FAK at Tyr-576 and Tyr-577, which are localized in
the activation domain, and their phosphorylation induces maximal
FAK kinase activity (31,32).
The present results demonstrated that EVs isolated
from the plasma of Mexican women with breast cancer promoted MMP-2
and MMP-9 secretion, Src and FAK activation, assembly of focal
adhesions, and the migration and invasion of TNBC MDA-MB-231
cells
Materials and methods
Materials
BD Matrigel™ was purchased from BD Biosciences.
Hoechst dye was obtained from Santa Cruz Biotechnology, Inc.
CellMask™ Orange plasma membrane stain was obtained from Invitrogen
(Thermo Fisher Scientific, Inc.). Tetramethylrhodamine
(TRITC)-conjugated phalloidin, mitomycin C, Annexin V and Triton™
X-100 were from Sigma-Aldrich (Merck KGaA). PP2 and PP3 were from
Merck KGaA.
Cell culture
MDA-MB-231 and MCF-7 breast cancer cells were
obtained from the American Type Culture Collection (ATCC) and
cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.), with 5%
FBS (ByProductos), 3.7 g/l sodium bicarbonate and antibiotics under
a humidified atmosphere with 5% CO2 and 95% air at
37°C.
Human mammary non-tumorigenic epithelial cells
(MCF12A) were obtained from ATCC and cultured in DMEM/F12 medium
(Gibco; Thermo Fisher Scientific, Inc.) with 5% FBS, 0.5 µg/ml
hydrocortisone (Sigma-Aldrich, Merck KGaA), 20 ng/ml epidermal
growth factor (EGF; Sigma-Aldrich, Merck KGaA), 10 µg/ml insulin
(Sigma-Aldrich, Merck KGaA) and antibiotics under a humidified
atmosphere with 5% CO2 and 95% air at 37°C.
MDA-MB-231 and MCF-7 cells were serum-starved in
DMEM for 24 h, and MCF12A cells were serum-starved in DMEM/F12 for
18 h before treatment with inhibitors and/or EVs.
Patients
The patients enrolled between August 2014 and
December 2018 consisted of 32 unrelated women who resided in Mexico
City (median age, 57.7 years; age range, 38–80 years) from the
First October Regional Hospital-ISSSTE (Mexico) with
biopsy-diagnosed breast cancer at clinical stages II and III, and
without receiving therapy. The control group consisted of 20
healthy women (median age, 42.7 years; age range, 16–86 years) who
did not have any family history of breast cancer. All study
participants provided signed informed consent, and the protocol was
approved by The Committee of Research, Ethics and Biosafety of the
First October Regional Hospital-ISSSTE, and was conducted in
accordance with the Declaration of Helsinki. The pathological and
clinical information of patients with breast cancer is shown in
Table I.
 | Table I.Patient characteristics. |
Table I.
Patient characteristics.
|
Characteristics | Patients, n |
|---|
| Tumor type |
|
| In
situ ductal carcinoma | 0 |
| In
situ lobular carcinoma | 0 |
|
Invasive ductal carcinoma | 32 |
| Primary tumor
size |
|
| T1 | 1 |
| T2 | 20 |
| T3 | 8 |
| T4 | 3 |
| Stage of breast
cancer |
|
| In
situ | 0 |
| I | 0 |
| II | 20 |
|
III | 12 |
| IV | 0 |
| Lymph node
status |
|
|
Negative | 0 |
|
Positive | 32 |
| Estrogen receptor
status |
|
|
Negative | 11 |
|
Positive | 21 |
| Her2/Neu
status |
|
|
Negative | 17 |
|
Positive | 15 |
| Progesterone
receptor status |
|
|
Negative | 16 |
|
Positive | 16 |
| Age, years |
|
|
Median | 57.7 |
|
Range | 38-80 |
Preparation of human plasma and
isolation of EVs
In total, 4 ml peripheral blood was collected into
polypropylene tubes containing sodium citrate (Vacutainer System;
BD Biosciences). Whole blood samples were centrifuged at 1,500 × g
for 15 min at 4°C, and plasma samples were obtained. Isolation of
EVs was performed as described previously (17). Then, one volume of 1 ml plasma was
centrifuged at 3,000 × g for 30 min at 4°C to remove platelets.
Plasma was obtained and centrifuged at 10,000 × g for 30 min at 4°C
to remove apoptotic bodies and platelet EVs, and supernatants were
aliquoted and frozen at −80°C until further analysis. Frozen
samples were thawed on ice and centrifuged at 110,000 × g for 70
min at 4°C and pellets were reconstituted in PBS or DMEM (EV
fractions). The absolute number of EVs was determined by flow
cytometry using BD Trucount™ tubes (BD Biosciences) (18).
Transmission electron microscopy
(TEM)
TEM of EV fractions was performed as described
previously (33). EV fractions
were adsorbed for 5 min on carbon-coated copper grids with mesh
formvar (0.3%) at room temperature. The grids were stained for 30
sec at room temperature with 2% uranyl acetate solution and excess
fluid was removed. Grids were air-dried and analyzed using a
JEM-1400 TEM (JEOL, Ltd.), operated at 80 kV and coupled with a
digital camera Veleta (Olympus SIS).
Nanoparticle tracking analysis
(NTA)
NTA was used to assess the size distribution of EV
fractions. EV fractions were diluted in 10 ml filtered PBS and
analyzed with a NanoSight NS300 (Malvern Instruments, Ltd.), which
was equipped with a 488 nm laser and a sCMOS camera. Then, three
videos of each sample were captured at a duration of 60 sec and
data were analyzed with NTA v3.0 software (Malvern Instruments,
Ltd.).
Stimulation of MDA-MB-231 cells with
EVs
Cultures of MDA-MB-231 cells (1.5×106
cells/dish) were serum-starved for 24 h, washed twice with PBS and
then stimulated at different time periods with EV fractions (20,000
EVs/1.5×106 cells/experimental condition) from patients
with breast cancer and healthy women. Scratch-wound assays were
performed for 48 h; invasion assays for 72 h; proliferation assays
for 48; and zymography assays for 3, 6, 9, 12 and 24 h.
Western blotting
Cells and EVs were solubilized in 0.1 ml of ice-cold
RIPA buffer (50 mM HEPES pH 7.4, 150 mM NaCl, 1 mM EGT4, 1 mM
sodium orthovanadate, 100 mM NaF, 10 mM sodium pyrophosphate, 10%
glycerol, 1% Triton X-100, 1% sodium deoxycholate, 1.5 mM
MgCl2, 0.1% SDS and 1 mM PMSF). Protein concentration of
each sample was determined by using the Bradford protein assay
(Bio-Rad Laboratories, Inc.). Equal amounts of protein (30 µg/lane)
were separated by SDS-PAGE on a 10% gel and proteins were
transferred to nitrocellulose membranes. Nitrocellulose membranes
were blocked for 2 h with 5% non-fat dried milk in PBS (pH 7.2/0.1%
Tween 20; wash buffer) at room temperature. The membranes were
incubated with the following primary antibodies at 4°C overnight:
Anti-Flotillin-2 (Flot-2) antibody (Ab; Mouse monoclonal; cat. no.
610383; 1:1,000; BD Biosciences), anti-CD9 Ab C-4 (Mouse
monoclonal; cat. no. sc-13118; 1:300; Santa Cruz Biotechnology,
Inc.), anti-FAK Ab D-1 (Mouse monoclonal; cat. no. sc-271126;
1:1,000; Santa Cruz Biotechnology, Inc.), anti-c-Src Ab 17AT28
(Mouse monoclonal; cat. no. sc-130124; 1:500; Santa Cruz
Biotechnology, Inc.), anti-CD81 Ab EPR4244 (Rabbit monoclonal; cat.
no. ab109201; 1:1,000; Abcam), anti-phosphorylated-specific Ab to
Tyr-397 of FAK (anti-p-FAK Ab; Rabbit polyclonal; cat. no. 44-624G;
IF 1:250; WB 1:1,000; Thermo Fisher Scientific, Inc.),
anti-phosphorylated-specific Ab to Tyr-418 of Src (anti-p-Src Ab;
rabbit polyclonal IgG; cat. no. AF2685; 1:1,000; R&D Systems,
Inc.), anti-vinculin Ab (Rabbit polyclonal; cat. no. V4139; IF
1:250; WB 1:500; Sigma-Aldrich; Merck KGaA) and anti-actin Ab
(Mouse polyclonal; 1:1,000) was provided by Dr Manuel Hernandez
(Cinvestav-IPN). Subsequently, membranes were washed three times
with wash buffer and incubated with horseradish
peroxidase-conjugated secondary Ab (cat. nos. G21040 and G21234;
1:5,000; Thermo Fisher Scientific, Inc.) for 2 h at room
temperature, and washed again with wash buffer three times. ECL
detection reagent (cat. no. sc-2048; Santa Cruz Biotechnology,
Inc.) and images were used for visualization of immunoreactive
bands. Bands were analyzed by using the ImageJ software v1.52e
(National Institute of Health).
EV uptake assays using flow
cytometry
EV uptake assays were performed as described
previously (34,35). EV fractions (20,000 EVs/condition)
were stained via incubation for 30 min at 4°C with CellMask Orange
dye solution (2.5 µg/ml). EV fractions were then washed with PBS,
centrifuged at 110,000 × g for 70 min at 4°C and reconstituted in
100 µl DMEM. MDA-MB-231 cells (1.5×106 cells/condition)
were incubated for 4 h at 37°C with stained EVs, and after
incubation, cells were washed twice with PBS and fixed for 20 min
at room temperature with a solution of 4% paraformaldehyde. Next,
cells were incubated for 10 min at room temperature with a solution
of trypsin (0.1%), washed twice with PBS and re-suspended in PBS-1%
BSA (Santa Cruz Biotechnology, Inc.). Controls of inhibition of EVs
uptake were included, and were obtained by treatment of EVs with
200 ng/ml Annexin V for 30 min at 4°C, before staining of EVs with
CellMask Orange dye solution for 30 min at 4°C. Cells were analyzed
using a BD FACSCalibur™ flow cytometer (BD Biosciences) and data
analysis was performed with Summit v4.3 software (Beckman Coulter,
Inc.).
EV uptake assays using confocal
microscopy
MDA-MB-231 cells (1.5×105
cells/condition) were incubated with stained EVs (20,000/condition)
for 4 h at 37°C. After incubation, cells were washed twice with
PBS, fixed for 20 min at room temperature with a solution of
paraformaldehyde (4%) and washed twice again with PBS. Next, cells
were counterstained with Hoechst for 20 min at room temperature and
mounted on glass slides using Vectashield. Preparations were
analyzed by confocal microscopy (Model TCS SP2; Leica Microsystems,
Inc.).
Scratch-wound assays
MDA-MB-231 cells (1.5×106 cells) were
cultured until they reached confluency and pretreated for 2 h at
37°C with 12 µM mitomycin C to inhibit proliferation during the
experiment. Cell cultures were scratched, washed with PBS and
supplemented with serum-free DMEM with or without inhibitors and/or
EV fractions (20,000 EVs/condition) at 37°C. After treatment for 48
h, the cultures were imaged using an inverted microscope coupled to
a camera (FSX100; Olympus Corporation; magnification, ×100). Images
from ≥3 fields per experimental condition were acquired and
analyzed using ImageJ software v1.52e (National Institutes of
Health). In total, one control of cell migration was included (5%
FBS).
Invasion assays
Inserts of 24-well plates (Costar; Corning, Inc.)
were covered with 50 µl Matrigel (3 mg/ml) and incubated at 37°C
for 2 h. Then, 1.2×105 MDA-MB-231 cells in serum-free
DMEM were plated on Matrigel of each insert in the upper chamber,
and the lower chamber contained EV fractions (20,000 EVs/condition)
in 600 µl DMEM. The plates with the inserts were incubated at 37°C
for 72 h under a humidified atmosphere with 5% CO2 and
95% air. After incubation, cells and Matrigel on the upper surface
of the membrane were removed, and cells on the lower surface of the
membrane were washed with PBS followed by fixation for 12 min at
room temperature with 4% paraformaldehyde. After fixation, invaded
cells were imaged using an inverted microscope couple to a camera
(FSX100, Olympus Corporation; magnification, ×400). Quantification
of invaded cells was calculated by staining membranes with 0.5%
crystal violet for 15 min at room temperature and elution of the
dye with 300 µl 30% acetic acid. Absorbance of the collected
solution was measured at 600 nm. Cells treated with 5% FBS were
included as an invasion control.
Zymography
Conditioned media from TNBC MDA-MB-231 cells
(1.5×106 cells) treated with EV fractions (20,000
EVs/condition) at different time points (3, 6, 9, 12 and 24 h) at
37°C from women with breast cancer and healthy controls were
concentrated using 5,000 Da Centricon® filters (EMD
Millipore). An equal volume (12 µl) of non-heated conditioned
medium and sample buffer (2% sucrose, 2.5% SDS, 4 µg/ml phenol red)
was mixed and the samples were loaded onto 8% acrylamide gels
copolymerized with gelatin (1 mg/ml). Next, gels were rinsed three
times for 30 min with Triton X-100 (2.5%) and incubated in assay
buffer (50 mM Tris-HCl pH 7.4, 5 mM CaCl2) for 48 h at
37°C. After incubation, gels were stained for 1 h at room
temperature with a solution of Coomassie Brilliant Blue G-250
(0.25%) dissolved in acetic acid (10%) and methanol (30%).
Proteolytic activity was identified as white zones on a blue
background. Controls of MMP-2 and MMP-9 secretions were included,
which were obtained by treatment of MDA-MB-231 cells with 400 mg/dl
ethanol and 100 ng/ml phorbol-12,13-dibutyrate for 24 h at 37°C, as
treatment of cells with these compounds induces the expression and
secretion of MMP-2 and MMP-9, respectively (36,37).
Immunofluorescence confocal
microscopy
MDA-MB-231 cells (1.5×105 cells) were
grown on coverslips, washed with PBS, equilibrated in DMEM and
treated with EV fractions (20,000 EVs/condition) for 30 min at
37°C. Cells were fixed with paraformaldehyde (4%) for 20 min at
room temperature, permeabilized with 0.5% Triton X-100 and then
blocked for 30 min at room temperature with FBS (10%) dissolved in
PBS. Next, cells were incubated overnight at 4°C with anti-p-FAK Ab
(1:250) followed by FITC-labeled anti-mouse secondary Ab (cat. no.
115-095-003; Jackson ImmunoResearch, Inc) for 2 h at 4°C. Staining
of focal contacts was performed by incubation of cells with
anti-vinculin Ab for 12 h at 4°C, while staining of cells for
fibrillar actin was performed by incubation for 2 h at 4°C with
TRITC-conjugated phalloidin (cat. no. R415; Thermo Fisher
Scientific, Inc.) Cells then were analyzed by confocal microscopy
(Model TCS SP2; Leica Microsystems, Inc.).
Cell proliferation assay
MDA-MB-231 cells (30,000 cells per well) were
treated for 48 h at 37°C with EV fractions (20,000 EVs/condition).
Next, 10 µl WST-1 reagent (cat. no. ab65473; Abcam) was added to
the cells in each well and microplates were incubated at 37°C for 2
h. The absorbance of each well was measured at 450 nm, and one
control for proliferation was included, which was prepared by
treatment of cells with 5% FBS for 48 h at 37°C.
Statistical analysis
Data are presented as the mean ± SD of ≥3
independent experiments. Data were analyzed by one-way ANOVA
followed by Tukeys post hoc test for ≥3 groups, while Students
t-test was used to analyze groups of two. P<0.05 was considered
to indicate a statically significant difference.
Results
EVs from plasma of patients with
breast cancer induce the migration of MDA-MB-231 cells
The present study characterized EV fractions using
NTA, TEM and western blotting against Flot-2, CD9 and CD81, which
are molecular markers associated with EVs (7). NTA and TEM results identified a
population of spherical vesicles with sizes between 30–300 nm in
healthy women, while women with breast cancer had vesicles between
50–600 nm (Fig. 1A and B).
Moreover, NTA showed that the number of EVs was significantly
higher in women with breast cancer (1.542×1011 EVs/ml)
than in healthy women (2.14×1010 EVs/ml). Western
blotting results demonstrated the presence of Flot-2, CD9 and CD81
protein expression in EV fractions from healthy women (Ctrl EVs)
and EV fractions from women with breast cancer (BC EVs; Fig. 1C).
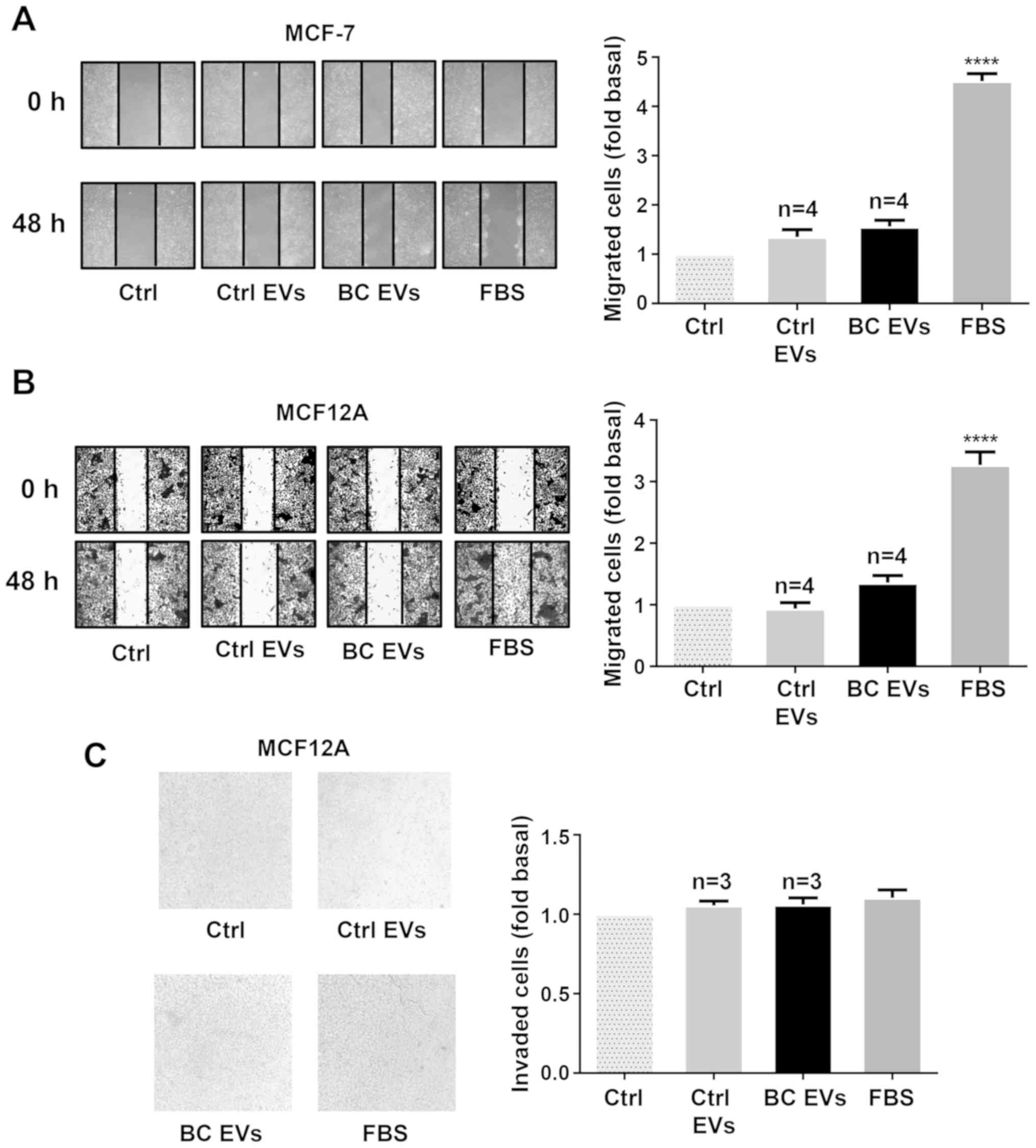
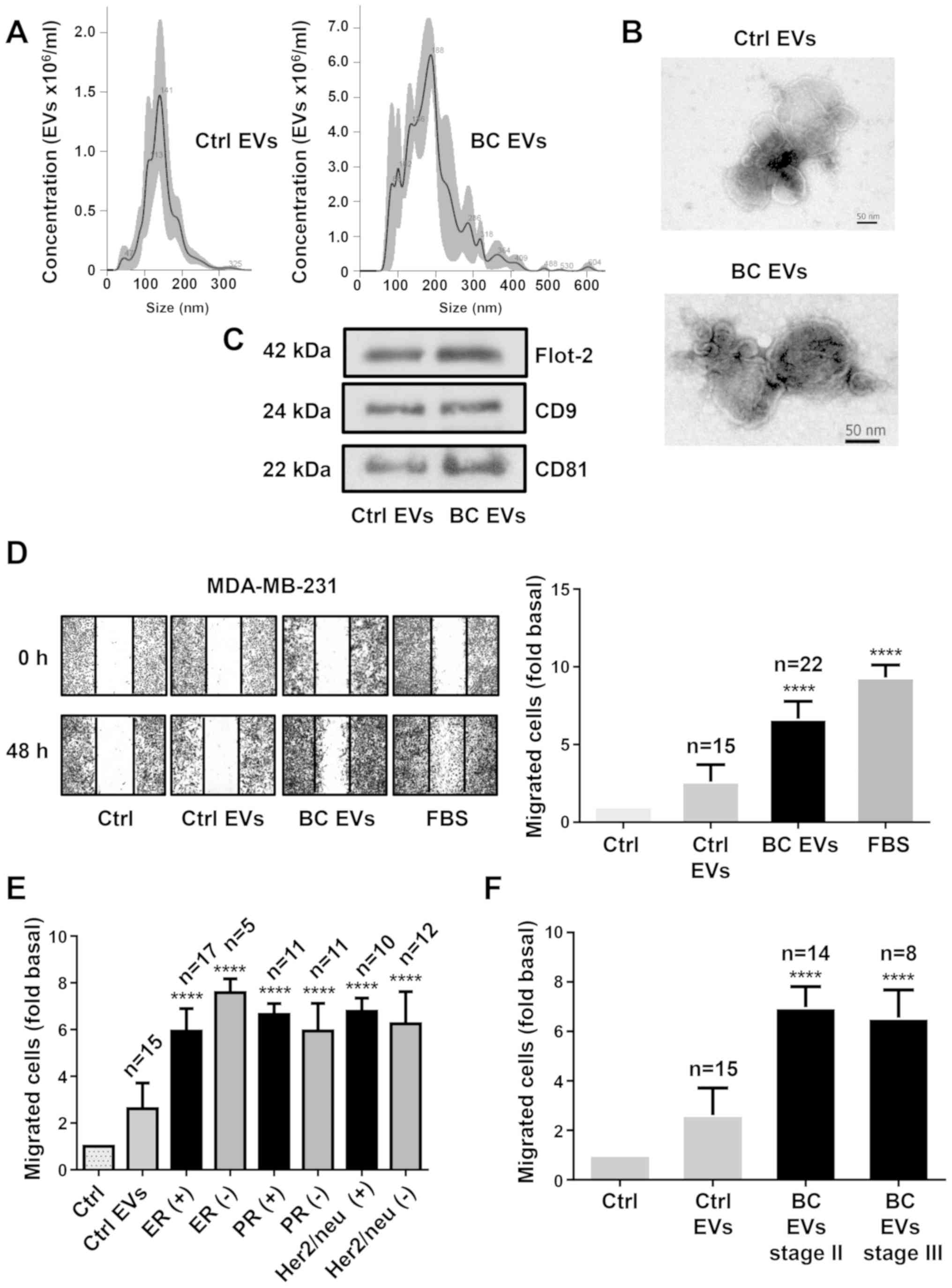 | Figure 1.EVs from plasma of patients with
breast cancer enhance migration in MDA-MB-231 cells. (A) Ctrl EVs
and BC EVs were analyzed by nanoparticle tracking analysis. The
calculated size distribution of particles was depicted as the mean
(black line) with standard errors (gray shaded area). (B) Ctrl EVs
and BC EVs were visualized by transmission electron microscopy. (C)
Ctrl EVs and BC EVs were analyzed by western blotting with
anti-Flot-2 Ab, anti-CD9 Ab and anti-CD81 Ab. Images are
representative of three independent experiments. (D) Cultures of
MDA-MB-231 cells were scratch-wounded and treated for 48 h with
Ctrl EVs and BC EVs; one control of FBS was included. (E) Analysis
of migration induced by BC EVs in relation to expression of ER, PR
and Her2/neu overexpression. (F) Analysis of migration induced by
BC EVs in relation to clinical stage of patients. Magnification,
100×. Data are presented as the mean ± SD, and indicate the fold of
migration above Ctrl. ****P<0.0001 vs. Ctrl. Ctrl, control; Ctrl
EVs, EV fractions obtained from healthy women; BC EVs, EV fractions
obtained from women with breast cancer; Ab, antibody; ER, estrogen
receptor; PR, progesterone receptor; EVs, extracellular vesicles;
Flot-2, flotillin-2. |
Next, whether BC EVs induced migration in TNBC
MDA-MB-231 cells was investigated. Cell migration assays were
performed using scratch-wound assays with MDA-MB-231 cells treated
with EVs from 22 patients with breast cancer and from 15 healthy
women. The results indicated that BC EVs induced increased
migration compared with Ctrl EVs in MDA-MB-231 cells (Fig. 1D). In addition, the association
between the migration induced by BC EVs in MDA-MB-231 cells and the
expression levels of estrogen, progesterone and Her2/neu receptors
in the mammary tumors of the women from where the plasma EVs were
obtained was analyzed. It was found that the migration induced by
BC EVs was not related with the expression levels of estrogen,
progesterone and Her2/neu receptors in the tumors of women with
breast cancer (Figs. 1E and
S1A). Furthermore, migration of
MDA-MB-231 cells induced by BC EVs was not related with the stages
II and III of the women with breast cancer (Figs. 1F and S1B).
To further examine these findings, whether BC EVs
induced migration and/or invasion was determined in another breast
cancer cell line (MCF-7) and mammary non-tumorigenic epithelial
cells (MCF12A). The results showed that treatment with BC EVs did
not induce migration in MCF-7 cells, and it did not induce
migration and invasion in MCF12A cells (Fig. 2A-C).
EVs from healthy women and patients
with breast cancer are taken up by MDA-MB-231 cells
It was studied whether Ctrl EVs and BC EVs were
taken up by MDA-MB-231 cells. Ctrl EVs and BC EVs were labeled with
CellMask orange dye, MDA-MB-231 cells were incubated with unstained
EVs and stained EVs, and the fluorescence intensity was analyzed by
flow cytometry. It was identified that fluorescence intensity was
higher in MDA-MB-231 cells treated with stained Ctrl EVs and
stained BC EVs compared with MDA-MB-231 cells treated with
unstained EVs (Figs. 3A and B and
S2). In addition, the comparison
of fluorescence intensity was not significantly different between
the value obtained from MDA-MB-231 cells treated with stained BC
EVs and the value obtained from MDA-MB-231 cells treated with
stained Ctrl EVs (Fig. 3C).
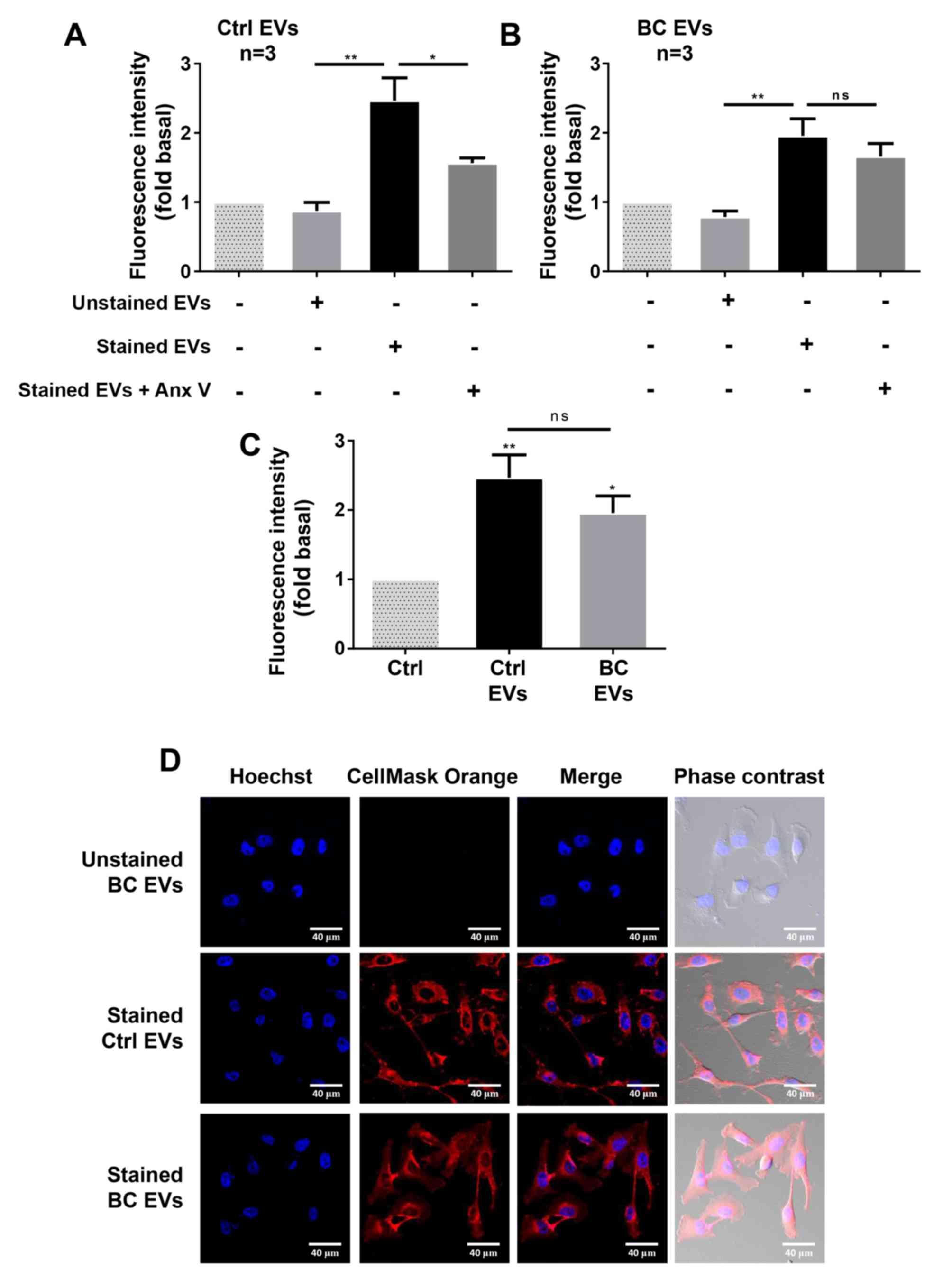 | Figure 3.EVs from plasma of healthy and breast
cancer groups are taken up by MDA-MB-231 cells. (A and B) Flow
cytometry analysis of MDA-MB-231 cells incubated with unstained or
stained Ctrl EVs and unstained or stained BC EVs. Controls of cells
without treatment with EVs and uptake inhibition (Annexin V) were
included. (C) Comparison of stained EVs uptake between MDA-MB-231
cells treated with stained BC EVs and stained Ctrl EVs. (D)
Confocal microscopy analysis of MDA-MB-231 cells incubated with
unstained BC EVs, stained Ctrl EVs, stained BC EVs and nucleus
stained with Hoechst dye. Scale bar, 40 µm. Data are presented as
the mean fluorescence intensities ± SD, and indicate the fold of
fluorescence intensity above Ctrl and Ctrl EVs. *P<0.05,
**P<0.01 vs. Ctrl or as indicated. ns, not significant; Ctrl,
control; EVs, extracellular vesicles; Ctrl EVs, EV fractions
obtained from healthy women; BC EVs, EV fractions obtained from
women with breast cancer. |
EVs express PS on their surface, and Annexin V has a
strong and specify affinity for PS (38). Controls of uptake inhibition were
included, and were obtained by treatment of EVs with 200 ng/ml
Annexin V, before staining of EVs with CellMask Orange dye. The
results indicated that treatment of stained Ctrl EVs with Annexin V
inhibited the increase of fluorescence intensity induced by stained
Ctrl EVs (Fig. 3A). However,
treatment of BC EVs with Annexin V did not inhibit the increased
fluorescence intensity induced by stained BC EVs (Fig. 3B).
To further examine whether EVs are taken up by
breast cancer cells, MDA-MB-231 cells treated with stained Ctrl
EVs, stained BC EVs and unstained BC EVs were analyzed by confocal
microscopy. It was found that MDA-MB-231 cells treated with stained
BC EVs and stained Ctrl EVs showed a red staining, while cells
treated with unstained BC EVs did not show any color (Fig. 3D).
EVs from patients with breast cancer
induce secretion of gelatinases
It was determined whether treatment of MDA-MB-231
cells with BC EVs induced gelatinase secretion. MDA-MB-231 cells
were stimulated for 3, 6, 9, 12 and 24 h with EVs from three
patients with breast cancer, and stimulated for 24 h with EVs from
three healthy women. Conditioned media were obtained, concentrated
and analyzed by gelatin zymography. It was identified that
stimulation of MDA-MB-231 cells with BC EVs induced increased
secretion of MMP-2 and MMP-9 at 12 and 24 h of treatment (Fig. 4A).
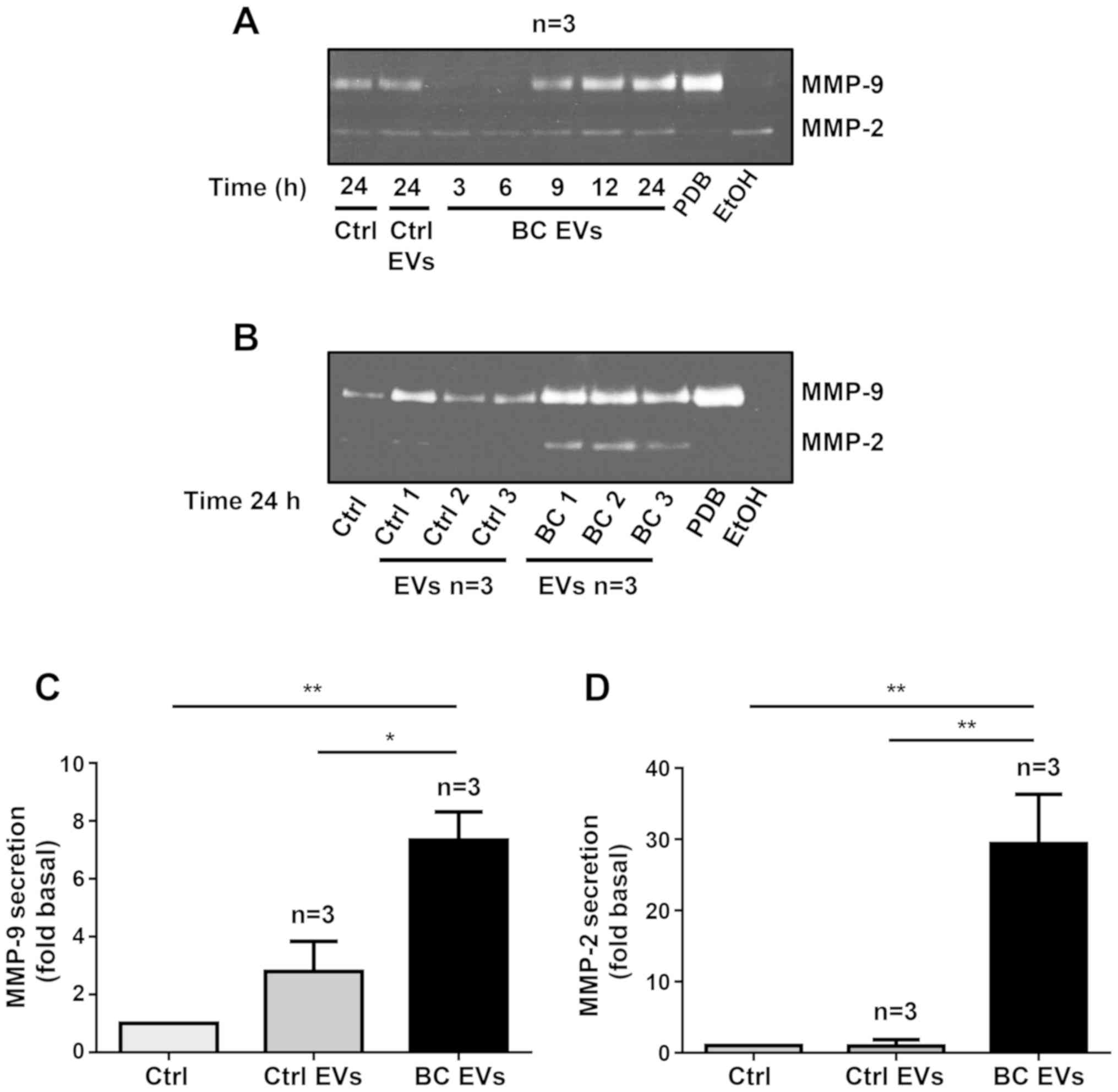 | Figure 4.EVs isolated from patients with
breast cancer mediate secretion of gelatinases. (A) MDA-MB-231
cells were incubated for various times with EVs from three healthy
women and EVs from three patients with breast cancer, and
conditioned media were collected. (B) MDA-MB-231 cells were
incubated for 24 h with EVs from three healthy women and three
patients with breast cancer. Gelatinase secretion was analyzed by
gelatin-substrate gels, and positive controls of MMP-2 (EtOH) and
MMP-9 (PDB) secretions were included. (C and D) Densitometric
analysis of MMP-9 and MMP-2 secretion. Data are presented as the
mean ± SD, and are expressed as fold of MMP-2 or MMP-9 secretion
above Ctrl and Ctrl EVs. *P<0.05, **P<0.01 as indicated.
Ctrl, control; EVs, extracellular vesicles; Ctrl EVs, EV fractions
obtained from healthy women; BC EVs, EV fractions obtained from
women with breast cancer; MMP, matrix metalloproteinase; EtOH,
ethanol; PBD, phorbol-12,13-dibutyrate. |
Since treatment of MDA-MB-231 cells with BC EVs for
24 h induced MMP-2 and MMP-9 secretion, MDA-MB-231 cells were
treated for 24 h with EVs from three patients with breast cancer
and EVs from three healthy women, and supernatants were analyzed by
gelatin zymography. The results indicated that treatment of
MDA-MB-231 cells with Ctrl EVs induced a small increase in MMP-9
secretion, but stimulation with BC EVs induced a significant
increase in both MMP-9 and MMP-2 secretion (Fig. 4B-D).
EVs from patients with breast cancer
induce Src activation
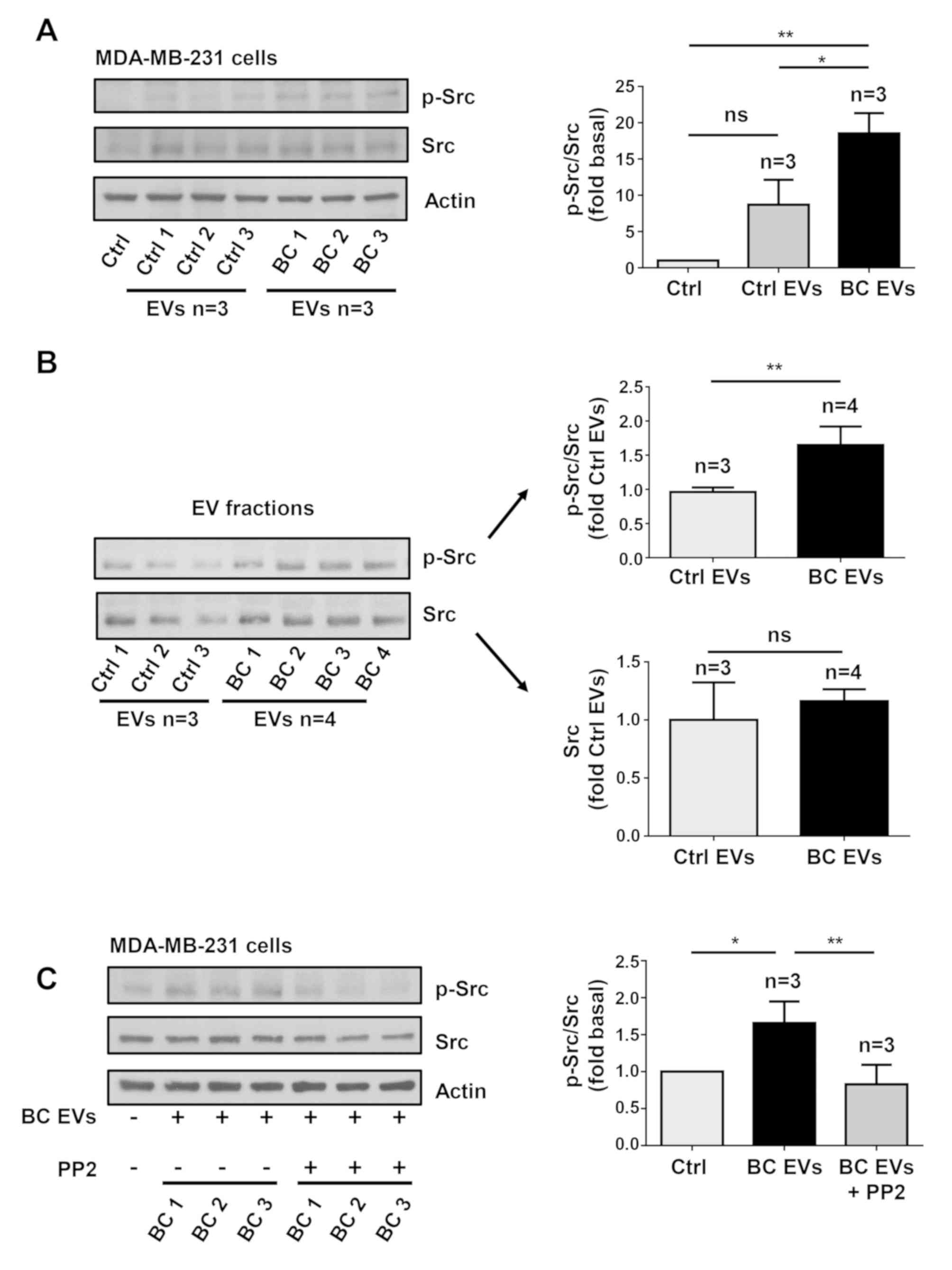
In order to determine whether treatment of
MDA-MB-231 cells with BC EVs induced Src activation, which is
initiated by its phosphorylation at Tyr-418 (p-Src), MDA-MB-231
cells were treated for 20 min with three Ctrl EV samples and three
BC EV samples, and cell lysates were analyzed by western blotting
with anti-p-Src Ab. It was identified that treatment of MDA-MB-231
cells with Ctrl EVs induced a small level of phosphorylation of Src
at Tyr-418, while treatment with BC EVs induced a strong
phosphorylation of Src at Tyr-418 (Fig. 5A).
Next, whether Ctrl EVs and BC EVs contain p-Src and
Src kinase was determined by western blotting with p-Src Ab and Src
Ab. The results showed that Ctrl EVs contained a low amount of
p-Src, while BC EVs contained a significantly larger amount of
p-Src (Fig. 5B). However, it was
demonstrated that Ctrl EVs and BC EVs contained a similar amount of
Src kinase (Fig. 5B).
EVs from patients with breast cancer
induce FAK activation via Src activity
The present study determined whether BC EVs induced
FAK activation, which is induced by its phosphorylation at Tyr-397
(p-FAK). Lysates of MDA-MB-231 cells treated for 20 min with three
Ctrl EV samples and three BC EV samples were analyzed by western
blotting with anti-p-FAK Ab. It was identified that treatment with
Ctrl EVs induced low levels of phosphorylation of FAK at Tyr-397,
whereas treatment with BC EVs induced a significantly higher level
of phosphorylation of FAK at Tyr-397 (Fig. 6A).
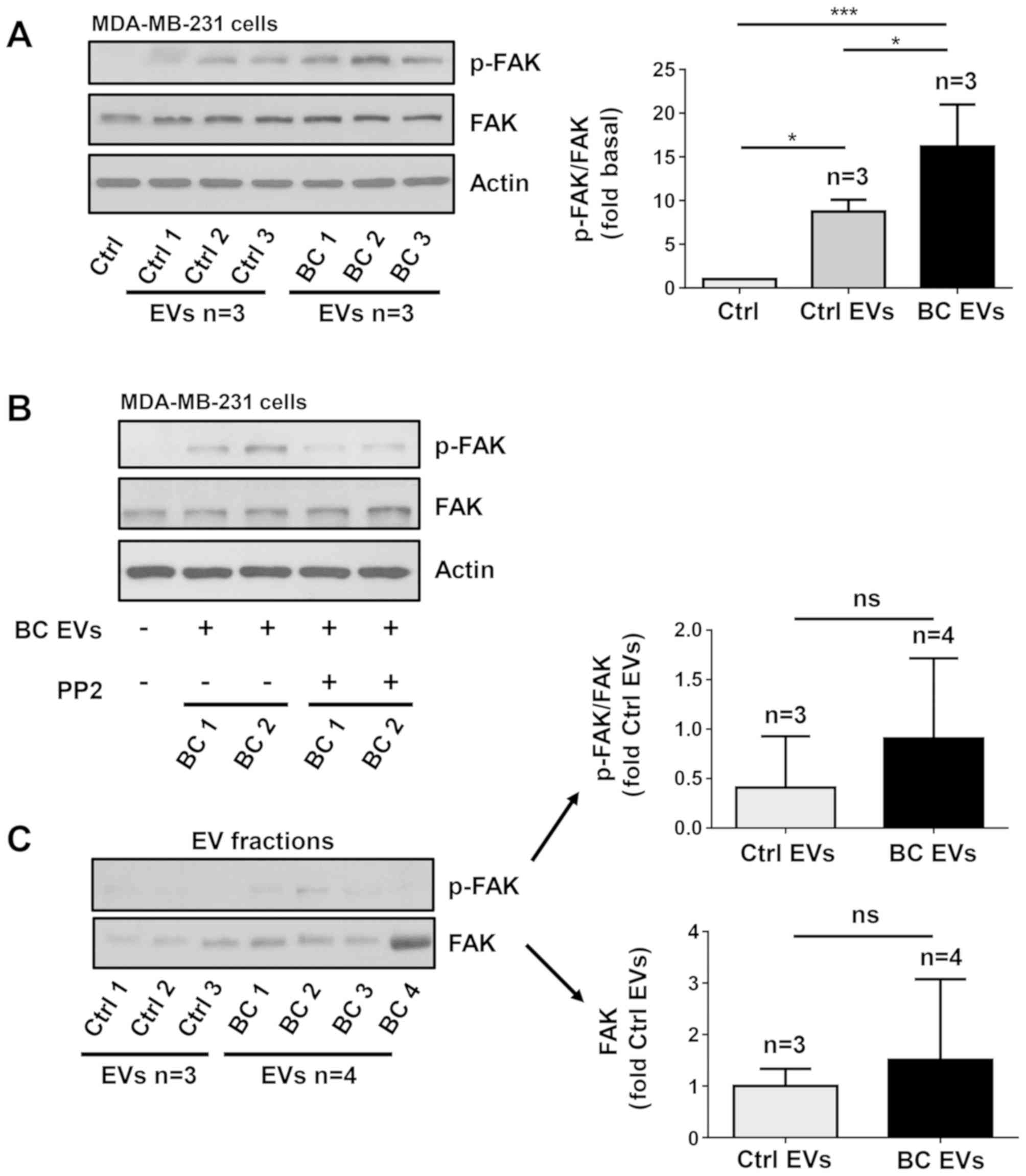 | Figure 6.EVs isolated from patients with
breast cancer induce FAK activation via a Src-dependent pathway.
(A) Lysates from MDA-MB-231 cells treated for 20 min with three
Ctrl EVs and three BC EVs were analyzed by western blotting with
anti-p-FAK Ab. Membranes were further analyzed by western blotting
with anti-FAK Ab and anti-actin Ab as loading controls. (B)
MDA-MB-231 cells were untreated and treated for 1 h with 10 µM PP2
and stimulated for 20 min with two BC EVs and lysed. Cell lysates
were analyzed by western blotting with anti-p-FAK Ab. Membranes
were analyzed further with anti-FAK Ab and anti-actin Ab as loading
controls. (C) Three Ctrl EVs and four BC EVs were analyzed by
western blotting with anti-p-FAK Ab and anti-FAK Ab. Data are
presented as the mean ± SD, and indicate the fold of p-FAK or FAK
above Ctrl and Ctrl EVs. *P<0.05, ***P<0.001 vs. Ctrl and
Ctrl EVs. ns, not significant; Ctrl, control; Ab, antibody; EVs,
extracellular vesicles; Ctrl EVs, EV fractions obtained from
healthy women; BC EVs, EV fractions obtained from women with breast
cancer; p-, phosphorylated; FAK, focal adhesion kinase. |
Since Src is able to induce maximal FAK activation
mediated by G-protein-coupled receptors (GPCRs) and tyrosine kinase
receptors (27), the role of Src
in FAK activation was investigated. The role of Src was examined
using PP2, which is a specific inhibitor of Src family members
(39). To determine whether PP2
inhibited the activity of Src, MDA-MB-231 cells were untreated and
treated for 1 h with 10 µM PP2 and then treated for 20 min with
three BC EVs, and lysed. Cell lysates were analyzed by western
blotting with anti-p-Src Ab, and it was found that PP2 inhibited
the increase of p-Src induced by BC EVs (Fig. 5C). Next, MDA-MB-231 cells were
untreated and treated for 1 h with 10 µM PP2 and stimulated for 20
min with BC EVs. It was identified that treatment with BC EVs
induced an increase of p-FAK via a Src-dependent pathway in
MDA-MB-231 cells (Fig. 6B).
The present study also assessed whether p-FAK and
FAK were localized in EV fractions, and three Ctrl EVs and four BC
EVs were analyzed by western blotting with p-FAK Ab and FAK Ab. The
results showed that Ctrl EVs and BC EVs expressed variable levels
of p-FAK and FAK, and there was no significant difference in p-FAK
and FAK expression levels between Ctrl EVs and BC EVs (Fig. 6C)
EVs from patients with breast cancer
induce redistribution of p-FAK and focal adhesions assembly
It was examined whether BC EVs induced a
redistribution of p-FAK, as well as the assembly of focal adhesions
and the role of Src in the assembly of focal adhesions. MDA-MB-231
cells cultured on coverslips were untreated or treated for 1 h with
10 µM PP2 and stimulated with Ctrl EVs and BC EVs. The
redistribution of p-FAK was analyzed by immunofluorescence with
anti-p-FAK Ab. Moreover, the number of focal adhesions was analyzed
by immunofluorescence with anti-vinculin Ab, as vinculin is a
cytoplasmic actin binding protein enriched in focal adhesions
(40). It was demonstrated that BC
EVs induced the redistribution of p-FAK at the edges of cells, and
increased the number of focal adhesions (Fig. 7A and B). Furthermore, focal
adhesion assembly induced by BC EVs was dependent on Src activity
(Fig. 7B). In addition, the
present study determined whether vinculin was localized in EV
fractions, and three Ctrl EVs and four BC EVs were analyzed by
western blotting with anti-vinculin Ab; the results indicated that
Ctrl EVs and BC EVs did not express vinculin (Fig. 7C). One control of vinculin
expression was included (Fig.
7D).
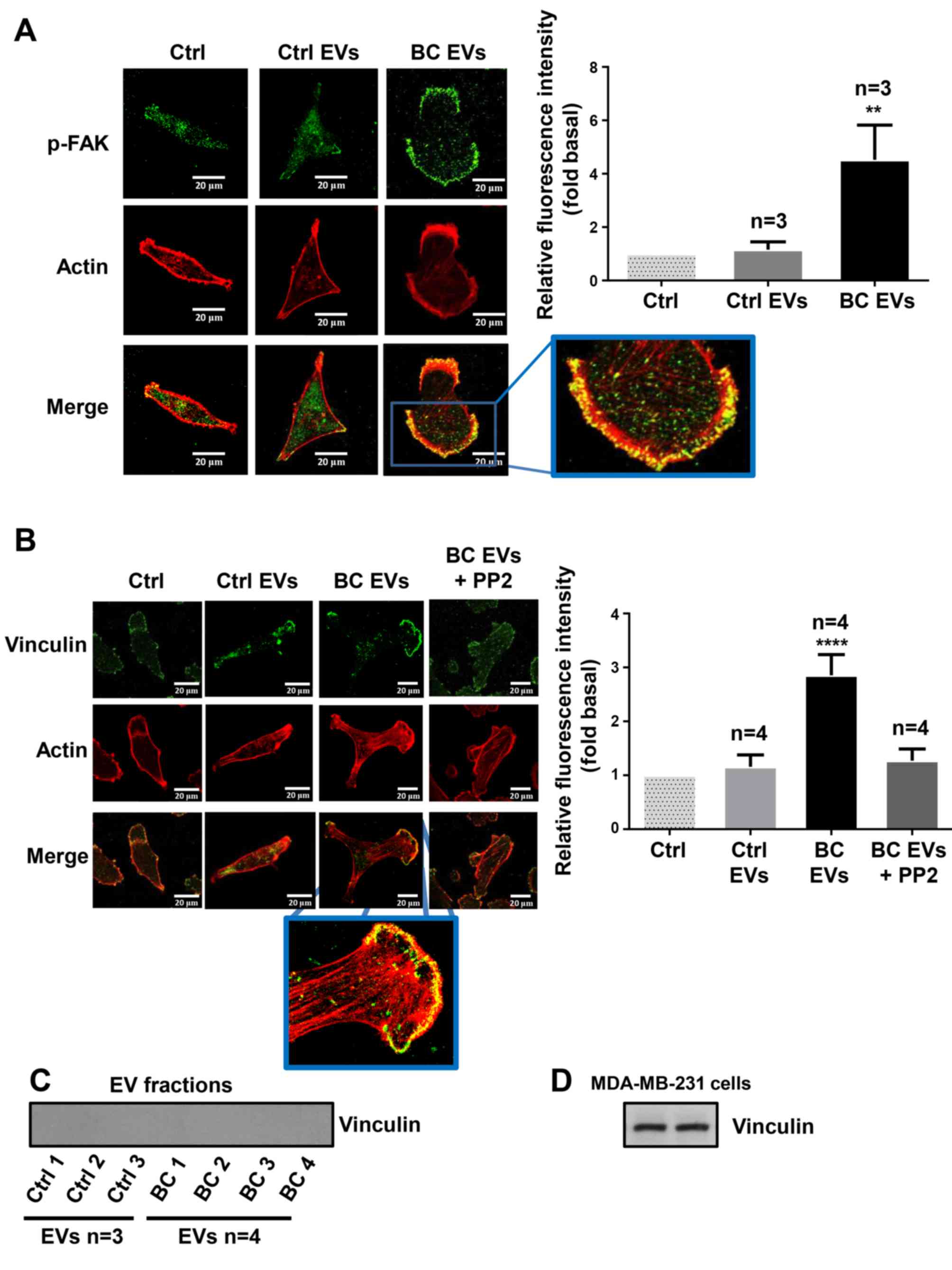 | Figure 7.EVs from patients with breast cancer
induce redistribution of p-FAK and focal adhesions assembly. (A and
B) MDA-MB-231 cells cultured on coverslips were treated for 1 h
with or without 10 µM of PP2, and stimulated for 30 min with Ctrl
EVs and BC EVs. Cells were incubated with Abs against p-FAK,
vinculin and tetramethylrhodamine-conjugated phalloidin, and were
analyzed by confocal microscopy. (C) Analysis of vinculin in Ctrl
EVs and BC EVs by western blotting. (D) Whole cell lysates of
MDA-MB-231 cell were included as a control of vinculin expression.
Data are presented as the mean ± SD of fluorescent intensities of
p-FAK and vinculin, and are expressed as fold above Ctrl.
**P<0.01, ****P<0.0001 vs. Ctrl. ns, not significant; Ctrl,
control; Ab, antibody; EVs, extracellular vesicles; Ctrl EVs, EV
fractions obtained from healthy women; BC EVs, EV fractions
obtained from women with breast cancer; p-, phosphorylated; FAK,
focal adhesion kinase. |
Role of Src in migration and invasion
mediated by EVs from patients with breast cancer
The role of Src in migration was examined using PP2
and its inactive analog (PP3) (39). Cultures of MDA-MB-231 cells were
treated for 1 h with 10 µM PP2 or 10 µM PP3, scratch-wounded and
stimulated with Ctrl EVs and BC EVs. It was demonstrated that
migration induced by BC EVs was dependent on Src activity in
MDA-MB-231 cells (Fig. 8A).
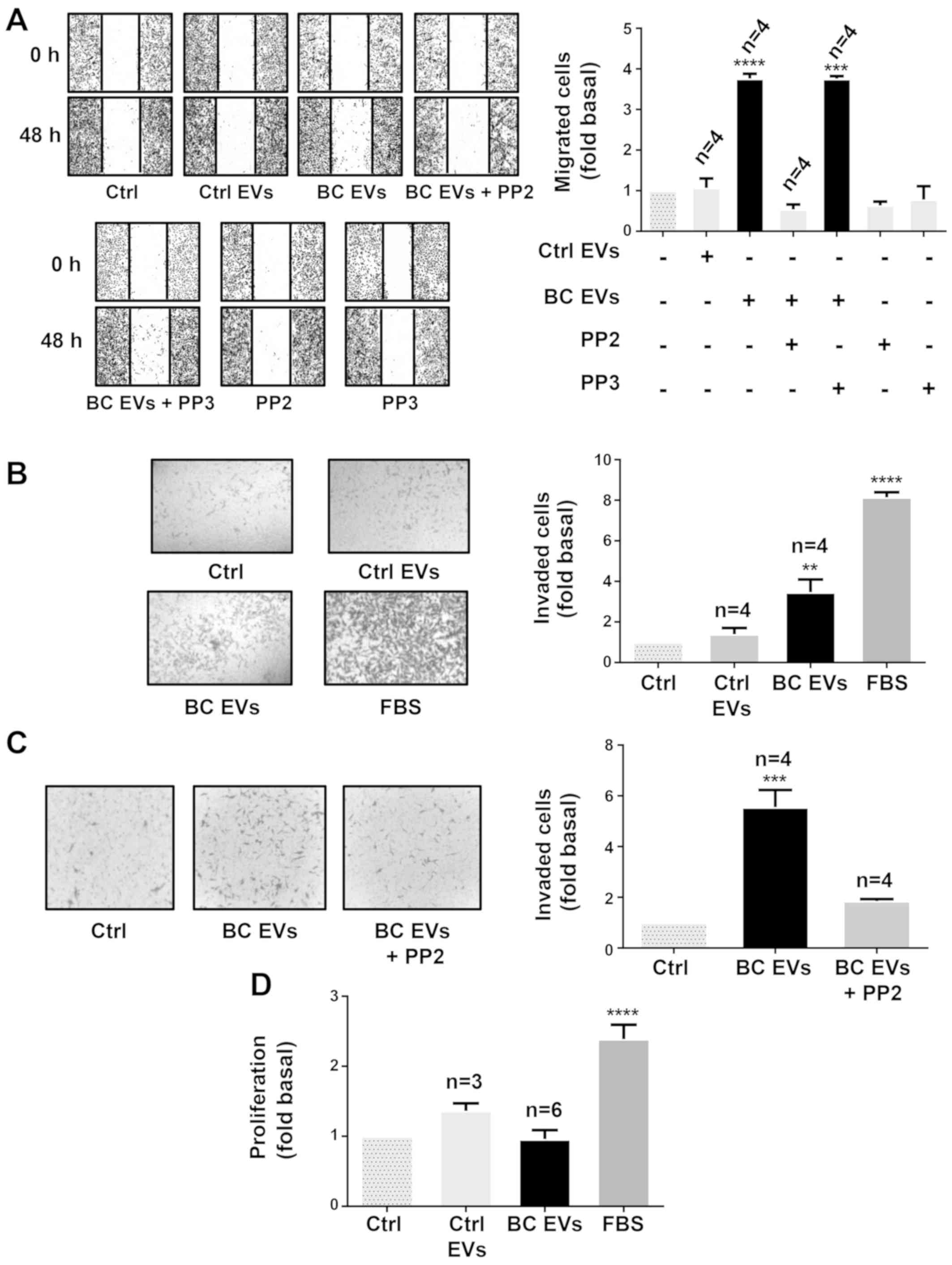 | Figure 8.EVs from patients with breast cancer
induce migration and invasion via a Src-dependent pathway. (A)
Migration assays of MDA-MB-231 cells treated for 1 h with or
without 10 µM PP2 or 10 µM PP3, and stimulated for 48 h with Ctrl
EVs and BC EVs. Magnification, ×100. (B and C) Invasion assays of
MDA-MB-231 cells treated with or without 10 µM PP2 and stimulated
for 48 h with Ctrl EVs and BC EVs. Magnification, ×400. (D)
Proliferation assay of MDA-MB-231 cells incubated with Ctrl EVs and
BC EVs. FBS was included as the control. Data are presented as the
mean ± SD, and are expressed as fold of migration, invasion or
proliferation above Ctrl. **P<0.01, ***P<0.001,
****P<0.0001 vs. Ctrl. Ctrl, control; EVs, extracellular
vesicles; Ctrl EVs, EV fractions obtained from healthy women; BC
EVs, EV fractions obtained from women with breast cancer. |
Next, it was studied whether BC EVs induced
invasion, and the role of Src was assessed using invasion assays
with MDA-MB-231 cells treated with Ctrl EVs and BC EVs. It was
identified that BC EVs induced invasion of MDA-MB-231 cells
(Fig. 8B). Moreover, invasion
assays performed in the presence of PP2 demonstrated that invasion
required Src activity (Fig.
8C).
In addition, whether BC EVs induce proliferation in
MDA-MB-231 cells was examined, and proliferation assay results
found that BC EVs and Ctrl EVs did not induce proliferation in
MDA-MB-231 cells (Fig. 8D).
Discussion
EVs are comprised of exosomes and microvesicles,
which constitute a variety of vesicles between 30–1,000 nm
(7). The present study isolated
EVs from plasma samples as described previously by Baran et
al (17), as this method was
reported to isolate EVs via the depletion of EVs from platelets. In
plasma, EVs from platelets constitute ~80% of total EVs (17,41).
The present results demonstrated that isolated EV fractions are
comprised of vesicles with sizes between 30–300 nm in healthy
women, while women with breast cancer showed EVs from 50–600 nm.
Furthermore, both Ctrl EVs and BC EVs expressed molecular markers
associated with EVs. Therefore, it was speculated that isolated EV
fractions from plasma samples corresponded to exosomes and
microvesicles, which are not contaminated with cell debris and
apoptotic bodies, and were free of platelet-derived EVs. Therefore,
it was proposed that cell processes studied may be mediated by
exosomes and/or microvesicles. The contribution of microvesicles
and exosomes to the cell processes analyzed remains to be
investigated.
Moreover, the present results demonstrated that the
number of EVs in plasma is higher in women with breast cancer than
in healthy women; however, the number of EVs in the present study
were found to be higher than the number of EVs reported in a
previous study (18). A different
number of EVs was found in the present study because the number of
EVs was determined using NTA, while in the previous study the
number of EVs was determined by flow cytometry. NTA has a higher
sensitivity for determining the number of EVs than flow cytometry.
However, both studies demonstrated that the number of EVs is higher
in women with breast cancer than in healthy women.
Cancer metastasis consists of several sequential
steps, including detachment of cells, migration, invasion to
surrounding tissues, intravasation, survival in circulation,
extravasation and colonization. Moreover, invasion of cancer cells
to other tissues involves cell migration as single cells
(mesenchymal type) or epithelial sheets (42). EVs are implicated in intercellular
communication in the tumor microenvironment, as they mediate
crosstalk between cancer and stromal cells (43). In addition, EVs support cancer
development, adaptation to hypoxic conditions, deprivation of
nutrients, escape of apoptosis, immune evasion and cancer
progression (43–45). Furthermore, exosomes released from
cancer-associated fibroblasts (CAFs) induce the formation of
protrusions and motility in MDA-MB-231 cells, while mesenchymal
stem cells secrete exosomes that promote motility and invasiveness
in breast cancer cells (46,47).
It has been shown that Hs578T cells and their more invasive variant
Hs578T(i)8 secrete EVs that promote proliferation,
migration and invasion in breast cancer cells (48). The present results showed that EVs
from women with breast cancer stages II and III induced cell
migration and this was dependent on Src activity in MDA-MB-231
cells. However, EVs from healthy women did not induce migration in
MDA-MB-231 cells. Moreover, migration induced by EVs from patients
with breast cancer was independent of the expression levels of
estrogen, progesterone and Her-2/neu receptors in the tumors of
patients. In contrast, it was identified that BC EVs did not induce
migration in MCF-7 cells, and did not induce migration and invasion
in MCF12A mammary epithelial cells. However, in contrast to the
present results, it has been previously reported that exosomes from
healthy women stimulate migration and invasion in MDA-MB-231 cells
(49). Thus, it was speculated
that BC EVs consist of subpopulations of exosomes and microvesicles
secreted from cancer cells (tumor) and stromal cells, such as
tumor-associated macrophages, mesenchymal stem cells and CAFs.
Therefore, BC EVs have a larger capacity for the induction of cell
migration and invasion compared with Ctrl EVs in MDA-MB-231 cells.
Thus, this may be the reason for the lack of the migration and
invasion mediated by stimulation with EVs from healthy women.
Furthermore, it was speculated that only BC EVs
contain molecules that induce the activation of specific signal
transduction pathways, including Src activation, which mediate a
variety of cell processes, including migration in breast cancer
cells. It was identified that BC EVs do not induce migration and/or
invasion in MCF12A mammary epithelial cells and MCF-7 breast cancer
cells, which express progesterone and estrogen receptors.
Furthermore, the present results suggested that EVs from women with
breast cancer stages II and III can induce migration in breast
cancer cells that do not express these receptors, and that this was
a specific process in TNBC cells. Moreover, it was speculated that
EVs in patients with breast cancer play an important role in cancer
progression, as they are able to induce migration and invasion. In
line with the present results, it has been reported that treatment
of HMLE human mammary epithelial cells with exosomes from
MDA-MB-231 cells transfected with microRNA-1246 increases its
ability for drug resistance, growth and invasion (50). In addition, K562 myeloid leukemia
cells release exosomes that promote angiogenesis via Src
activation, while exosomes from C4-2B, PC3 and DUI45 prostate
cancer cells contain Src, insulin-like growth factor 1 (IGF-1)
receptor and FAK, which are molecules involved in cell migration
and invasion (51,52).
Tumor cells communicate with surrounding cells,
including CAFs, endothelial cells and mesenchymal cells, via the
secretion and uptake of EVs, which mediate biological processes,
such as migration, angiogenesis and invasion, and can also modulate
the tumor microenvironment and metastasis (34,43,53).
The present results indicated that MDA-MB-231 cells are able to
take up BC EVs and Ctrl EVs, however only BC EVs induce migration.
Thus, BC EVs may have cargo molecules that promote activation of
specific signal transduction pathways, which increases migration
and invasion in breast cancer cells.
EVs express PS on their membrane surface, which
mediates the fusion of EVs with target cells. Therefore, treatment
of EVs with Annexin V inhibits the fusion of EVs with target cells,
as Annexin V binds to PS (43,54,55).
In the present study, it was identified that treatment of Ctrl EVs
with Annexin V inhibited the increase of fluorescence intensity
mediated by Ctrl EVs. However, treatment of BC EVs with Annexin V
did not inhibit the increased fluorescence intensity mediated by BC
EVs. Thus, it was speculated that BC EVs do not express, or express
very low levels, of PS on their membrane surface. It has been
reported that EV fractions obtained from females with type I
diabetes and controls without disease contain both EVs expressing
PS and EVs without expression of PS (56), which is consistent with the present
results. Moreover, the percentage of PS in EVs from patients is
31%, while the percentage of PS in controls without disease is 44%
(56).
Src family kinases mediate a variety of cellular
processes, such as cell cycle progression, proliferation, survival
and migration (57). Furthermore,
it has been reported that breast cancer tumors and cell lines have
an increase in Src activity (58).
In addition, linoleic acid induces FAK and Src activation, and cell
migration via a Src-dependent pathway in MDA-MB-231 cells (28). The present results demonstrated
that stimulation with BC EVs induced a stronger activation of FAK
and Src compared with treatment with Ctrl EVs, and also increased
the number of focal adhesions in MDA-MB-231 cells. As FAK and Src
activation and focal adhesion assembly mediate migration and
invasion, it was speculated that BC EVs induced the activation of
signal transduction pathways, including the activation of FAK and
Src, which mediated migration and invasion in MDA-MB-231 cells. In
line with the present results, mouse embryonic fibroblasts
expressing onco-Dbl release microvesicles containing FAK and
stimulation of fibroblasts with these vesicles promotes
proliferation, which is independent of anchorage and survival in
fibroblasts (59).
A variety of phosphoproteins, including Src, have
been revealed in EVs from human plasma (60). Moreover, EVs from plasma of women
with breast cancer contain FAK and EGFR kinases, and the level of
these kinases are increased in specific stages of breast cancer in
comparison with the control group (18). Furthermore, the present results
indicated that Ctrl EVs and BC EVs contained similar expression
levels of p-FAK, FAK and Src, but BC EVs contained larger amounts
of p-Src compared with Ctrl EVs. Thus, it was demonstrated, that BC
EVs induced FAK and Src activation; however, the contribution of
p-FAK, FAK, p-Src and Src expressed in EVs to the activation of
these kinases in the target MDA-MB-231 cells requires further
investigation.
A catalytic reciprocal activation model of FAK and
Src has been previously reported, where Src associates with FAK and
phosphorylates FAK at Tyr-576 and Tyr-577, which induces maximal
kinase activity of FAK (32,61,62).
In addition, it has been revealed that maximal kinase activity of
FAK promotes intermolecular phosphorylation between FAK molecules
at Tyr-397, which induces signal amplification (32,61,62).
In line with this model, the present results demonstrated that BC
EVs induced activation of FAK, which was dependent on Src kinase
activity. Linoleic acid, oleic acid and arachidonic acid induce
activation of FAK via GPCRs and a mechanism of reciprocal catalytic
activation of FAK (28,63–65).
Therefore, it was speculated that EVs mediated activation of FAK
and Src, as well as migration and invasion via activation of
receptors.
Tumors release EVs that play a pivotal role in the
invasion and metastasis processes, EVs from patients with breast
cancer are associated with metastasis and relapse (14,43).
In the present study, it was identified that EVs from patients with
breast cancer induced migration and invasion via Src activity in
MDA-MB-231 cells. Moreover, in line with the present results, it
has been shown that exosomes from patients with TNBC induce
invasion in SKBR3 cells (48). In
addition, it was found that treatment of non-tumorigenic mammary
epithelial cells MCF12A with BC EVs does not induce migration. It
is proposed that BC EVs may mediate progression processes in breast
cancer via the transfer of molecules and activation of signal
transduction pathways. Supporting this hypothesis, different tumors
release EVs with a variety of integrin expression patterns, which
are able to determine organ specific metastasis (14,66).
Moreover, EVs expressing α6β4 and α6β1 integrins are associated
with lung metastasis, while EVs expressing αvβ5 are associated with
liver metastasis (14,66).
ECM degradation is required for tumor growth and
metastasis (67). In addition, EVs
contain proteases that mediate degradation of ECM (68,69).
It has been reported that human fibrosarcoma cells HT1080 release
EVs expressing gelatinases in an active form, while breast cancer
cells 8701-BC secrete EVs containing MMP-9 (68,69).
The present results demonstrated that BC EVs induced an increase in
MMP-9 and MMP-2 secretion in MDA-MB-231 cells. Therefore, EVs may
participate in the progression of breast cancer.
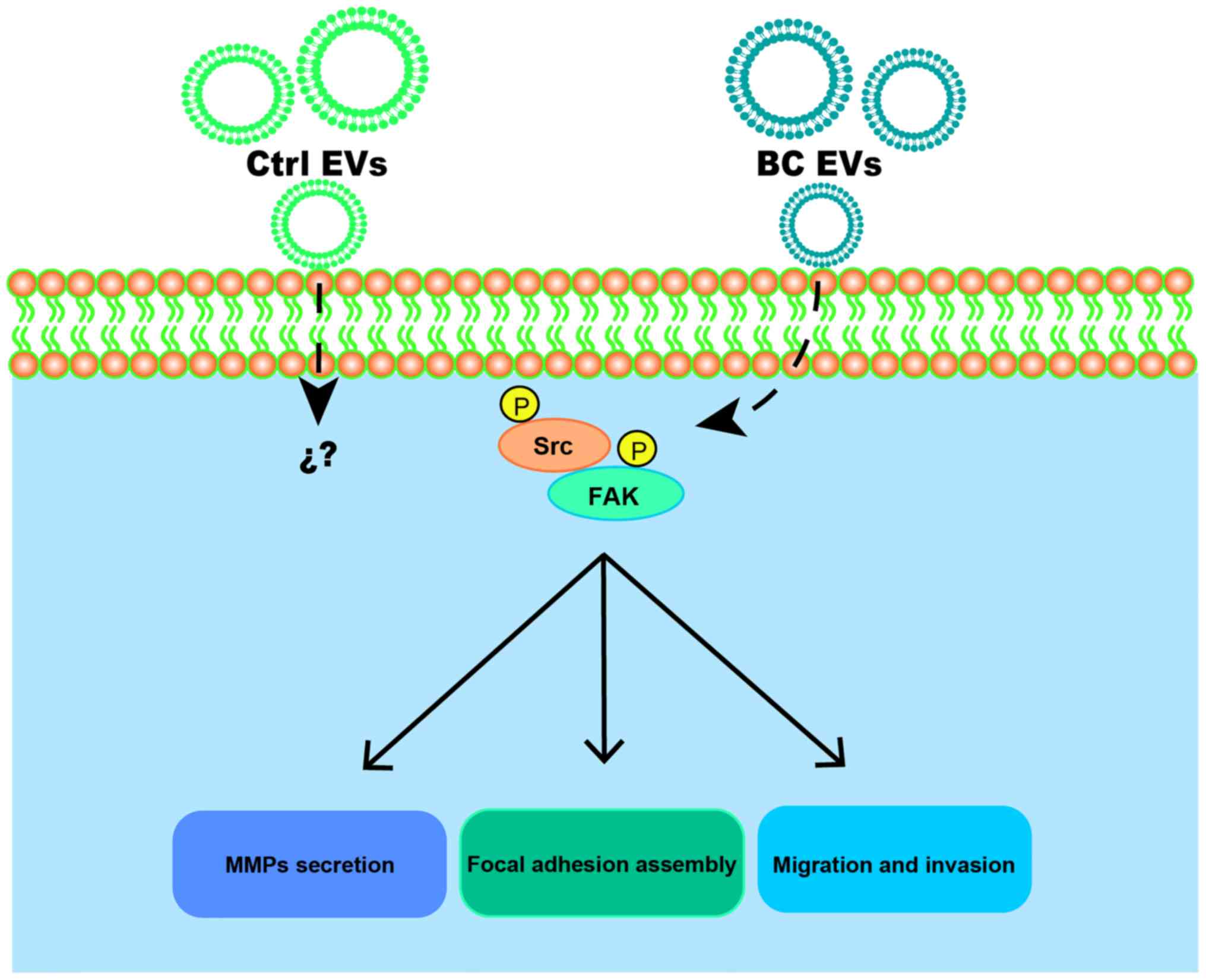
In conclusion, it was demonstrated that BC EVs
promoted migration, invasion and MMP-2 and MMP-9 secretion in
MDA-MB-231 cells (Fig. 9).
Moreover, BC EVs enhanced migration and invasion via a
Src-dependent pathway. Therefore, the present results suggested
that EVs may participate in breast cancer progression.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Ms. Nora Ruiz and
Ms. María de Lourdes-Rojas (LaNSE, Cinvestav-IPN) for their
technical assistance.
Funding
The research was funded by CONACYT (grant no.
255429) and CONACYT-FOSISS (grant no. 261637), Mexico. Grants from
CONACYT supported the present study.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors contributions
JRR performed the majority of experiments. EPS, JRR
and RTB drafted the paper. ELO and PCR helped with experiments and
reviewed/edited the manuscript. EPS, RTB, EMB, COM, FBM, AHT and
IEA analyzed and validated data, and reviewed part of the
manuscript. EPS and RTB coordinated the study and wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All studied participants provided signed informed
consent, and the protocol was approved by the ethics committee of
the First October Regional Hospital-ISSSTE (approval no. 121.2016),
and was conducted in accordance with the Declaration of
Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Samavat H and Kurzer MS: Estrogen
metabolism and breast cancer. Cancer Lett 356 (2 Pt A). 231–243.
2015. View Article : Google Scholar
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ruiz R, Herrero C, Strasser-Weippl K,
Touya D, St Louis J, Bukowski A and Goss PE: Epidemiology and
pathophysiology of pregnancy-associated breast cancer: A review.
Breast. 35:136–141. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nishimura R and Arima N: Is triple
negative a prognostic factor in breast cancer? Breast Cancer.
15:303–308. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Perou CM, Sørlie T, Eisen MB, van de Rijn
M, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA,
et al: Molecular portraits of human breast tumours. Nature.
406:747–752. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sørlie T, Perou CM, Tibshirani R, Aas T,
Geisler S, Johnsen H, Hastie T, Eisen MB, van de Rijn M, Jeffrey
SS, et al: Gene expression patterns of breast carcinomas
distinguish tumor subclasses with clinical implications. Proc Natl
Acad Sci USA. 98:10869–10874. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Raposo G and Stoorvogel W: Extracellular
vesicles: Exosomes, microvesicles, and friends. J Cell Biol.
200:373–383. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Théry C, Zitvogel L and Amigorena S:
Exosomes: Composition, biogenesis and function. Nat Rev Immunol.
2:569–579. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Skotland T, Sandvig K and Llorente A:
Lipids in exosomes: Current knowledge and the way forward. Prog
Lipid Res. 66:30–41. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kelleher RJ Jr, Balu-Iyer S, Loyall J,
Sacca AJ, Shenoy GN, Peng P, Iyer V, Fathallah AM, Berenson CS,
Wallace PK, et al: Extracellular vesicles present in human ovarian
tumor microenvironments induce a phosphatidylserine-dependent
arrest in the T-cell signaling cascade. Cancer Immunol Res.
3:1269–1278. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lima LG, Chammas R, Monteiro RQ, Moreira
ME and Barcinski MA: Tumor-derived microvesicles modulate the
establishment of metastatic melanoma in a
phosphatidylserine-dependent manner. Cancer Lett. 283:168–175.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lea J, Sharma R, Yang F, Zhu H, Ward ES
and Schroit AJ: Detection of phosphatidylserine-positive exosomes
as a diagnostic marker for ovarian malignancies: A proof of concept
study. Oncotarget. 8:14395–14407. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sharma R, Huang X, Brekken RA and Schroit
AJ: Detection of phosphatidylserine-positive exosomes for the
diagnosis of early-stage malignancies. Br J Cancer. 117:545–552.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kanada M, Bachmann MH and Contag CH:
Signaling by extracellular vesicles advances cancer hallmarks.
Trends Cancer. 2:84–94. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dourado MR, Korvala J, Åström P, De
Oliveira CE, Cervigne NK, Mofatto LS, Campanella Bastos D, Pereira
Messetti AC, Graner E, Paes Leme AF, et al: Extracellular vesicles
derived from cancer-associated fibroblasts induce the migration and
invasion of oral squamous cell carcinoma. J Extracell Vesicles.
8:15785252019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Au Yeung CL, Co NN, Tsuruga T, Yeung TL,
Kwan SY, Leung CS, Li Y, Lu ES, Kwan K, Wong KK, et al: Exosomal
transfer of stroma-derived miR21 confers paclitaxel resistance in
ovarian cancer cells through targeting APAF1. Nat Commun.
7:111502016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baran J, Baj-Krzyworzeka M, Weglarczyk K,
Szatanek R and Zembala M, Barbasz J, Czupryna A, Szczepanik A and
Zembala M: Circulating tumour-derived microvesicles in plasma of
gastric cancer patients. Cancer Immunol Immunother. 59:841–850.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Galindo-Hernandez O, Villegas-Comonfort S,
Candanedo F, González-Vázquez MC, Chavez-Ocaña S,
Jimenez-Villanueva X, Sierra-Martinez M and Salazar EP: Elevated
concentration of microvesicles isolated from peripheral blood in
breast cancer patients. Arch Med Res. 44:208–214. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhao Z, Fan J, Hsu YS, Lyon CJ, Ning B and
Hu TY: Extracellular vesicles as cancer liquid biopsies: From
discovery, validation, to clinical application. Lab Chip.
19:1114–1140. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sternlicht MD and Werb Z: How matrix
metalloproteinases regulate cell behavior. Annu Rev Cell Dev Biol.
17:463–516. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Coussens LM, Fingleton B and Matrisian LM:
Matrix metalloproteinase inhibitors and cancer: Trials and
tribulations. Science. 295:2387–2392. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
McCawley LJ and Matrisian LM: Matrix
metalloproteinases: Multifunctional contributors to tumor
progression. Mol Med Today. 6:149–156. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
DeMali KA, Wennerberg K and Burridge K:
Integrin signaling to the actin cytoskeleton. Curr Opin Cell Biol.
15:572–582. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wozniak MA, Modzelewska K, Kwong L and
Keely PJ: Focal adhesion regulation of cell behavior. Biochim
Biophys Acta. 1692:103–119. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Parsons JT, Martin KH, Slack JK, Taylor JM
and Weed SA: Focal adhesion kinase: A regulator of focal adhesion
dynamics and cell movement. Oncogene. 19:5606–5613. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schlaepfer DD, Hauck CR and Sieg DJ:
Signaling through focal adhesion kinase. Prog Biophys Mol Biol.
71:435–478. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Serna-Marquez N, Villegas-Comonfort S,
Galindo-Hernandez O, Navarro-Tito N, Millan A and Salazar EP: Role
of LOXs and COX-2 on FAK activation and cell migration induced by
linoleic acid in MDA-MB-231 breast cancer cells. Cell Oncol
(Dordr). 36:65–77. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Schaller MD: Biochemical signals and
biological responses elicited by the focal adhesion kinase. Biochim
Biophys Acta. 1540:1–21. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao J and Guan JL: Signal transduction by
focal adhesion kinase in cancer. Cancer Metastasis Rev. 28:35–49.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Parsons JT: Focal adhesion kinase: The
first ten years. J Cell Sci. 116:1409–1416. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Schaller MD, Hildebrand JD and Parsons JT:
Complex formation with focal adhesion kinase: A mechanism to
regulate activity and subcellular localization of Src kinases. Mol
Biol Cell. 10:3489–3505. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Thery C, Amigorena S, Raposo G and Clayton
A: Isolation and characterization of exosomes from cell culture
supernatants and biological fluids. Curr Protoc Cell Biol.
30:3.22.1–3.22.29. 2006. View Article : Google Scholar
|
|
34
|
Galindo-Hernandez O, Gonzales-Vazquez C,
Cortes-Reynosa P, Reyes-Uribe E, Chavez-Ocaña S, Reyes-Hernandez O,
Sierra-Martinez M and Salazar EP: Extracellular vesicles from women
with breast cancer promote an epithelial-mesenchymal
transition-like process in mammary epithelial cells MCF10A. Tumour
Biol. 36:9649–9659. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kawamoto T, Ohga N, Akiyama K, Hirata N,
Kitahara S, Maishi N, Osawa T, Yamamoto K, Kondoh M, Shindoh M, et
al: Tumor-derived microvesicles induce proangiogenic phenotype in
endothelial cells via endocytosis. PLoS One. 7:e340452012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ke Z, Lin H, Fan Z, Cai TQ, Kaplan RA, Ma
C, Bower KA, Shi X and Luo J: MMP-2 mediates ethanol-induced
invasion of mammary epithelial cells over-expressing ErbB2. Int J
Cancer. 119:8–16. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Park MJ, Park IC, Hur JH, Rhee CH, Choe
TB, Yi DH, Hong SI and Lee SH: Protein kinase C activation by
phorbol ester increases in vitro invasion through regulation of
matrix metalloproteinases/tissue inhibitors of metalloproteinases
system in D54 human glioblastoma cells. Neurosci Lett. 290:201–204.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Muralidharan-Chari V, Clancy JW, Sedgwick
A and DSouza-Schorey C: Microvesicles: Mediators of extracellular
communication during cancer progression. J Cell Sci. 123:1603–1611.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hanke JH, Gardner JP, Dow RL, Changelian
PS, Brissette WH, Weringer EJ, Pollok BA and Connelly PA: Discovery
of a novel, potent, and Src family-selective tyrosine kinase
inhibitor. Study of Lck- and FynT-dependent T cell activation. J
Biol Chem. 271:695–701. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bays JL and DeMali KA: Vinculin in
cell-cell and cell-matrix adhesions. Cell Mol Life Sci.
74:2999–3009. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Diamant M, Nieuwland R, Pablo RF, Sturk A,
Smit JW and Radder JK: Elevated numbers of tissue-factor exposing
microparticles correlate with components of the metabolic syndrome
in uncomplicated type 2 diabetes mellitus. Circulation.
106:2442–2447. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gupta GP and Massagué J: Cancer
metastasis: Building a framework. Cell. 127:679–695. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Becker A, Thakur BK, Weiss JM, Kim HS,
Peinado H and Lyden D: Extracellular vesicles in cancer:
Cell-to-cell mediators of metastasis. Cancer Cell. 30:836–848.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yáñez-Mó M, Siljander PR, Andreu Z, Zavec
AB, Borràs FE, Buzas EI, Buzas K, Casal E, Cappello F, Carvalho J,
et al: Biological properties of extracellular vesicles and their
physiological functions. J Extracell Vesicles. 4:270662015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Maas SLN, Breakefield XO and Weaver AM:
Extracellular vesicles: Unique intercellular delivery vehicles.
Trends Cell Biol. 27:172–188. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lin R, Wang S and Zhao RC: Exosomes from
human adipose-derived mesenchymal stem cells promote migration
through Wnt signaling pathway in a breast cancer cell model. Mol
Cell Biochem. 383:13–20. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Luga V and Wrana JL: Tumor-stroma
interaction: Revealing fibroblast-secreted exosomes as potent
regulators of Wnt-planar cell polarity signaling in cancer
metastasis. Cancer Res. 73:6843–6847. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
OBrien K, Rani S, Corcoran C, Wallace R,
Hughes L, Friel AM, McDonnell S, Crown J, Radomski MW and ODriscoll
L: Exosomes from triple-negative breast cancer cells can transfer
phenotypic traits representing their cells of origin to secondary
cells. Eur J Cancer. 49:1845–1859. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shtam T, Naryzhny S, Samsonov R, Karasik
D, Mizgirev I, Kopylov A, Petrenko E, Zabrodskaya Y, Kamyshinsky R,
Nikitin D, et al: Plasma exosomes stimulate breast cancer
metastasis through surface interactions and activation of FAK
signaling. Breast Cancer Res Treat. 174:129–141. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li XJ, Ren ZJ, Tang JH and Yu Q: Exosomal
microRNA miR-1246 promotes cell proliferation, invasion and drug
resistance by targeting CCNG2 in breast cancer. Cell Physiol
Biochem. 44:1741–1748. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
DeRita RM, Zerlanko B, Singh A, Lu H,
Iozzo RV, Benovic JL and Languino LR: c-Src, insulin-like growth
factor I receptor, G-protein-coupled receptor kinases and focal
adhesion kinase are enriched into prostate cancer cell exosomes. J
Cell Biochem. 118:66–73. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mineo M, Garfield SH, Taverna S, Flugy A,
De Leo G, Alessandro R and Kohn EC: Exosomes released by K562
chronic myeloid leukemia cells promote angiogenesis in a
Src-dependent fashion. Angiogenesis. 15:33–45. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Naito Y, Yoshioka Y, Yamamoto Y and Ochiya
T: How cancer cells dictate their microenvironment: Present roles
of extracellular vesicles. Cell Mol Life Sci. 74:697–713. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Meckes DG Jr, Shair KH, Marquitz AR, Kung
CP, Edwards RH and Raab-Traub N: Human tumor virus utilizes
exosomes for intercellular communication. Proc Natl Acad Sci USA.
107:20370–20375. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Keller S, König AK, Marmé F, Runz S,
Wolterink S, Koensgen D, Mustea A, Sehouli J and Altevogt P:
Systemic presence and tumor-growth promoting effect of ovarian
carcinoma released exosomes. Cancer Lett. 278:73–81. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bergen K, Mobarrez F, Jörneskog G, Wallén
H and Tehrani S: Phosphatidylserine expressing microvesicles in
relation to microvascular complications in type 1 diabetes. Thromb
Res. 172:158–164. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Parsons JT and Parsons SJ: Src family
protein tyrosine kinases: Cooperating with growth factor and
adhesion signaling pathways. Curr Opin Cell Biol. 9:187–192. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Egan C, Pang A, Durda D, Cheng HC, Wang JH
and Fujita DJ: Activation of Src in human breast tumor cell lines:
Elevated levels of phosphotyrosine phosphatase activity that
preferentially recognizes the Src carboxy terminal negative
regulatory tyrosine 530. Oncogene. 18:1227–1237. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kreger BT, Dougherty AL, Greene KS,
Cerione RA and Antonyak MA: Microvesicle cargo and function changes
upon induction of cellular transformation. J Biol Chem.
291:19774–19785. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen IH, Xue L, Hsu CC, Paez JS, Pan L,
Andaluz H, Wendt MK, Iliuk AB, Zhu JK and Tao WA: Phosphoproteins
in extracellular vesicles as candidate markers for breast cancer.
Proc Natl Acad Sci USA. 114:3175–3180. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Owen JD, Ruest PJ, Fry DW and Hanks SK:
Induced focal adhesion kinase (FAK) expression in FAK-null cells
enhances cell spreading and migration requiring both auto- and
activation loop phosphorylation sites and inhibits
adhesion-dependent tyrosine phosphorylation of Pyk2. Mol Cell Biol.
19:4806–4818. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Salazar EP and Rozengurt E: Src family
kinases are required for integrin-mediated but not for G
protein-coupled receptor stimulation of focal adhesion kinase
autophosphorylation at Tyr-397. J Biol Chem. 276:17788–17795. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Navarro-Tito N, Robledo T and Salazar EP:
Arachidonic acid promotes FAK activation and migration in
MDA-MB-231 breast cancer cells. Exp Cell Res. 314:3340–3355. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Navarro-Tito N, Soto-Guzman A,
Castro-Sanchez L, Martinez-Orozco R and Salazar EP: Oleic acid
promotes migration on MDA-MB-231 breast cancer cells through an
arachidonic acid-dependent pathway. Int J Biochem Cell Biol.
42:306–317. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Soto-Guzman A, Robledo T, Lopez-Perez M
and Salazar EP: Oleic acid induces ERK1/2 activation and AP-1 DNA
binding activity through a mechanism involving Src kinase and EGFR
transactivation in breast cancer cells. Mol Cell Endocrinol.
294:81–91. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hoshino A, Costa-Silva B, Shen TL,
Rodrigues G, Hashimoto A, Tesic Mark M, Molina H, Kohsaka S, Di
Giannatale A, Ceder S, et al: Tumour exosome integrins determine
organotropic metastasis. Nature. 527:329–335. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hotary K, Li XY, Allen E, Stevens SL and
Weiss SJ: A cancer cell metalloprotease triad regulates the
basement membrane transmigration program. Genes Dev. 20:2673–2686.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Dolo V, Ginestra A, Cassarà D, Violini S,
Lucania G, Torrisi MR, Nagase H, Canevari S, Pavan A and Vittorelli
ML: Selective localization of matrix metalloproteinase 9, beta1
integrins, and human lymphocyte antigen class I molecules on
membrane vesicles shed by 8701-BC breast carcinoma cells. Cancer
Res. 58:4468–4474. 1998.PubMed/NCBI
|
|
69
|
Ginestra A, Monea S, Seghezzi G, Dolo V,
Nagase H, Mignatti P and Vittorelli ML: Urokinase plasminogen
activator and gelatinases are associated with membrane vesicles
shed by human HT1080 fibrosarcoma cells. J Biol Chem.
272:17216–17222. 1997. View Article : Google Scholar : PubMed/NCBI
|