Introduction
The genome profile stability serves a crucial role
in the maintenance of human health. Emerging evidence has indicated
that the dysregulation of gene expression occurs in multiple
diseases, such as cardiovascular disease (1), Parkinson's disease (2) and malignant tumors (3,4).
Colorectal cancer (CRC) is a common malignant disease of the
gastrointestinal tract; the incidence and mortality of CRC has been
steadily declining for the past two decades (5), with an exception of younger adults
(<50 years), which is possibly related to an increase in cancer
screening and improved therapeutic modalities (5). Currently, the treatment of CRC mainly
includes surgical resection, radiotherapy and chemotherapy
(6). Another modern aspect of CRC
treatment is immunotherapy with programmed cell death protein 1
(PD-1) inhibitors nivolumab and pembrolizumab, which currently
constitute the standard of care for the treatment of
chemotherapy-resistant microsatellite instability-high/mismatch
repair-deficiency CRC (7).
Patients with early stage CRC usually have favorable prognoses
after surgery, yet the prognoses of most patients at a late stage
are poor due to the tumor invasion and metastasis (8). Furthermore, epithelial-mesenchymal
transition (EMT) is a key cellular event for various biological
processes including cell division, fetal development and wound
healing (9,10). In malignant diseases, EMT is
usually considered as an important factor for promoting cell
invasion, migration and drug resistance (10). The expression levels of N-cadherin
and E-cadherin are important biomarkers of EMT progress (11). Previous studies have reported that
EMT participates during colon cancer carcinogenesis and development
(12); therefore, identifying
novel therapeutic targets to prevent EMT may be valuable for
CRC.
MicroRNAs (miRNAs) are a type of short RNA sequences
without the capability of encoding proteins, usually 18–25
nucleotides long (13). miRNAs
post-transcriptionally inhibit target mRNA expression by binding to
their 3′untranslated regions (3′UTRs) (14). miRNAs participate in the regulation
of various biological events, including tumor growth, invasion,
metastasis and angiogenesis (15).
miRNA (miR)-199a-5p belongs to the miR-199a family, which has been
demonstrated to participate in the progression of multiple diseases
such as acute myeloid leukemia (16) and cervical cancer (17). Dai et al (18) reported that miR-199a-5p prevented
hepatocyte damage induced by bile acid. Furthermore, miR-199a-5p
was downregulated in non-small cell lung cancer (19). miR-199a-5p is associated with CRC
cell proliferation via targeting ROCK1 (20) and FZD6 (21); however, the impact of miR-199a-5p
on other signaling pathways during CRC is not completely
understood.
Integrins are a class of transmembrane receptors,
and their main function is to participate in the regulation of
cellular adhesion (22). Integrins
are dimers consisting of α (120–185 kDa) and β (90–110 kDa)
subunits; they can receive mechanical stimuli the from
extracellular matrix and transit these stimuli into downstream
signals to regulate cell proliferation, adhesion and migration
(23,24). Integrin α3β1 (ITGA3) has been
reported to be upregulated and correlated with poor prognosis in
oral squamous cell carcinoma and pancreatic cancer (25,26).
Moreover, ITGA3 promotes EMT and cancer stemness during breast
cancer (27); therefore, ITGA3 may
be a potential target for anticancer therapy.
The present study aimed to identify the role of
miR-199aa-5p during CRC carcinogenesis and progression. Using
bioinformatics analysis, several potential targets of miR-199a-5p
were identified, among them, ITGA3 was further investigated.
Materials and methods
Samples
A total of 101 pairs of tumor and para-tumor tissue
(2 cm away from the tumor margin) specimens obtained from 49 male
and 52 female patients (age range, 38–79 years) were collected from
Jinhua People's Hospital between March 2018 and October 2019. This
research did not cause any extra medical expenses or pain. The
samples were grouped according to the TNM phasing: 37 cases of T1,
43 cases of T2, 11 cases of T3 and 10 cases of T4 tumors. All
samples were collected with the written informed consent of
patients, and the study was approved by the Ethics Committee of
Jinhua People's Hospital. All tissues were stored at −80°C.
Cell culture
Human CRC cell lines HCT-116 and Caco-2 and the
normal colon epithelial cell line NCM460 were obtained from the
Cell Bank of Chinese Academy of Sciences (Shanghai, China). NCM460,
HCT-116 and Caco-2 cells were cultured in McCoy's 5A medium (Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco: Thermo
Fisher Scientific, Inc.) and 1% penicillin/streptomycin in a
humidified atmosphere containing 5% CO2 at 37°C.
Cell transfection
Cells (1×105/well) were plated into
6-well plates and cultured for 24 h, and subsequently transfected
with miR-199a-5p mimics negative control (miR-199a-5p NC; sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′), miR-199a-5p mimics (sense,
5′-CCCAGUGUUCAGACUACCUGUUC-3′ and antisense,
5′-ACAGGUAGUCUGAACACUGGGUU-3′), pcDNA-NC (empty), pcDNA-ITGA3
(Shanghai GenePharma Co., Ltd.), short hairpin RNA (sh)-NC or
sh-ITGA3 (pLVX-sh-ITGA3 plasmids supplied by Shanghai GenePharma
Co., Ltd.) using Lipofectamine® 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's
instructions. Briefly, Firstly, 1 µg nucleic acid fragment and 2 µl
Lipofectamine 2000 were mixed with 0.5 ml serum-free McCoy's 5A
medium. After incubation for 15 min at room temperature, the
mixture was added to each well with 4 ml serum-free medium.
Following incubation for 24 h, the culture medium was replaced with
McCoy's 5A medium containing 10% FBS. After 24–72 h, cells were
used for subsequent experiments. pcDNA-ITGA3 transfection
efficiency is presented in Figs.
S1 and S2.
MTT assay
HCT-116, Caco-2 and NCM460 cells were seeded in
96-well plates at the density of 1×104 cells/well. The
cells were incubated at 37°C for 24 h after the cells were
attached. Subsequently, the medium was removed, and MTT (0.5 mg/ml)
diluted in medium was added to the wells. After 3 h, the medium
containing MTT was removed and 200 µl DMSO was added. Finally, the
absorbance was detected on a microplate reader (Thermo Fisher
Scientific, Inc.) at 595 nm. The experiments were performed
independently at last three times.
Wound healing assay
Wound healing assay was used to investigate the cell
migration in the different groups. Following treatment with
miR-199a-5p mimics, mimics NC, sh-NC or sh-ITGA3, HCT-116 cells
were collected and seeded (1×106/well) in 6-well plates
for 24 h, followed by scratching with a 10-µl pipette tip, and then
images of the scratches were captured under a light microscope.
Following 24 h incubation with McCoy's 5A medium containing 1% FBS,
the migration of the HCT-116 cells were calculated using the
following formula: (S0 h-S24 h)/S0
h × 100%, where S represents the width of the wound.
Transwell assay
The Transwell assay was used to investigate cell
invasion after treatment with miR-199a-5p mimics, mimics NC, sh-NC
or sh-ITGA3. The upper chambers of the Transwell plates were
pre-coated with 50 µl 1:1 mixture of Matrigel® and
McCoy's 5A medium at 37 °C for 1 h. Briefly, HCT-116 cells were
collected and seeded in serum-free McCoy's 5A medium, and then
re-seeded (2×104 cells/well) into the upper chambers,
while McCoy's 5A medium containing 20% FBS was placed in the lower
chambers. Following incubation at 37°C for 24 h, non-invasive cells
were removed using a sterile cotton swab and the invading cells
were fixed with 4% paraformaldehyde at room temperature for 10 min.
Subsequently, invading cells were stained using 0.1% crystal violet
at room temperature for 20 min and observed using a light
microscope (magnification, ×400).
Total RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
miR-199a-5p and ITGA3 mRNA expression levels were
detected by RT-qPCR. Total RNA in the different groups was
extracted using the TRIzol® Reagent kit (Invitrogen;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. Total RNA was reverse transcribed into cDNA using the
RT Reagent kit (TransGen Biotech) or the miRNA First-Strand cDNA
Synthesis SuperMix kit (TransGen Biotech). The following
thermocyclcing conditions were used for reverse transcription: 42°C
for 15 min, followed by 5 sec at 85°C and storage at 4°C until
further analysis. Subsequently, qPCR was preformed using an ABI
Detection System (Applied Biosystems; Thermo Fisher Scientific,
Inc.) and SYBR® Green Premix Ex Taq™ (Takara Bio, Inc.).
The following thermocycling conditions were used for qPCR: Initial
denaturation for 30 sec at 95°C; followed by 40 cycles of 5 sec at
95°C and 30 sec at 60°C; and dissociation at 95°C for 15 sec, 60°C
for 30 sec and 95°C for 15 sec. The primer sequences used for qPCR
were as follows: ITGA3 forward, 5′-TCAACCTGGATACCCGATTCC-3′ and
reverse, 5′-GCTCTGTCTGCCGATGGAG-3′; miR-199a-5p forward:,
5′-TCAAGAGCAATAACGAAAAATGT-3′ and reverse,
5′-GCTGTCAACGATACGCTACGT-3′; U6 forward,
5′-CGCTTCGGCAGCACATATACTA-3′ and reverse,
5′-CGCTTCACGAATTTGCGTGTCA-3′; GAPDH forward,
5′-AGGTCGGTGTGAACGGATTTG-3′ and reverse, 5′-GGGGTCGTTGATGGCAACA-3′.
mRNA and miRNA expression levels were quantified using the
2−∆∆Cq method and normalized to the internal reference
genes GAPDH and U6, respectively (28).
Total protein extraction and western
blotting
After treatment with miR-199a-5p mimics, mimics NC,
sh-ITGA3 or sh-NC, HCT-116 cells were lysed using RIPA buffer
(Nanjing KeyGen Biotech Co., Ltd.). Subsequently, protein samples
(25 µg per lane) were quantified using a Bicinchoninic Acid Assay
kit (Beyotime Institute of Biotechnology), separated via 10%
SDS-PAGE and transferred to PVDF membranes. The membranes were
blocked with 5% skim milk at room temperature for 1–2 h.
Subsequently, the membranes were incubated with an anti-ITGA3
antibody (cat. no. ab131055; 1:500; Abcam) and an anti-GAPDH
antibody (cat. no. 10494-1-AP; 1:2,000; ProteinTech Group, Inc.) at
4°C overnight. Following primary incubation, the membranes were
incubated with a horseradish peroxidase-conjugated Affinipure Goat
Anti-Rat IgG (H+L) secondary antibody (cat. no. SA00001-15;
1:2,000; ProteinTech Group, Inc.) at 37°C for 1 h. Protein bands
were visualized using Western Bright™ ECL (cat. no. K-12045-D50;
Advasta, Inc.) and analyzed using ImageJ software (version 1.8.0;
National Institutes of Health) with GAPDH as the loading
control.
Immunofluorescence assay
The expression levels of E-cadherin and N-cadherin
were evaluated by an immunofluorescence assay. Following
co-transfection with pcDNA-ITGA3 and miR-199a-5p mimics or mimics
NC, HCT-116 cells were fixed with 4% paraformaldehyde at 4°C for 24
h. Subsequently, cells were incubated with the following primary
antibodies at 4°C overnight: Anti-E-cadherin (cat. no. ab40772;
1:1,000; Abcam) and anti-N-cadherin (cat. no. ab18203; 1:1,000;
Abcam). Following primary incubation, cells were incubated with an
anti-Rabbit IgG H&L horseradish peroxidase-conjugated secondary
antibody (cat. no. ab6721; 1:1,000; Abcam) at 37°C for 30 min.
Subsequently, cells were stained with DAPI at room temperature for
5 min and observed using a fluorescence microscope (magnification,
×400).
Dual luciferase reporter assay
The relationship between the 3′UTR of ITGA3 and
miR-199a-5p was investigated using a dual luciferase reporter
assay. HCT-116 cells were transfected with 300 ng/well miR-199a-5p
mimics or mimics NC, 300 ng/well mutant (MUT) or wild-type (WT)
3′UTR ITGA3 and 100 ng Renilla luciferase plasmid (pRL-TK;
Promega Corporation) using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). HCT-116 cells
transfected with pRL-TK alone served as the negative control group.
After co-transfection, the cells were cultured at 37°C for 24 h.
Subsequently, the luciferase activities in the different groups
were analyzed using a Dual-Luciferase Reporter assay system
(Promega Corporation), according to the manufacturer's protocol.
Firefly luciferase activities were normalized to Renilla
luciferase activities.
Immunohistochemical analysis
(IHC)
The expression of ITGA3 in CRC tissues and normal
tissues was detected by IHC staining. The paraffin sections in
different groups were dewaxed in xylene and hydrated using an
ascending alcohol gradient. The tissues were boiled in citric acid
buffer (pH 6.0) for 10 min, followed by cooling to room
temperature. The sections were blocked using goat serum (Beijing
Solarbio Science & Technology Co., Ltd.) for 30 min at room
temperature, and incubated using an anti-ITGA3 antibody (cat. no.
ab131055; 1:500; Abcam) at 4°C overnight. Following primary
incubation, the sections were incubated with a goat anti-rabbit IgG
H&L horseradish peroxidase-conjugated secondary antibody (cat.
no. ab6721; 1:500; Abcam) at 37°C for 20 min. Subsequently, the
sections were stained with DAB (OriGene Technologies, Inc.) at room
temperature for 5 min and observed under a light microscope
(magnification, ×400).
Statistical analysis
Statistical analyses were performed using SPSS
software (version 23.0; IBM Corp.). The differences between two
groups were analyzed using the paired Student's t-test. The
differences among multiple groups were analyzed by one-way ANOVA
followed by Tukey's post-hoc test at the 95% confidence interval.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-199a-5p and ITGA3 expression is
dysregulated in CRC tissues and cells
To determine the expression levels of miR-199a-5p
and ITGA3 in CRC, total RNA was extracted from CRC and adjacent
non-cancerous tissues. RT-qPCR was applied to evaluate the
expression of miR-199a-5p and ITGA3 at the mRNA level. As presented
in Fig. 1A and D, miR-199a-5p was
significantly downregulated in CRC tissues and cancer cells
compared with normal tissues. The expression of miR-199a-5p was
significantly decreased in T2, T3 and T4 tumors compared with
normal tissues, although no statistically significant differences
were observed between normal tissues and T1 (Fig. 1A).
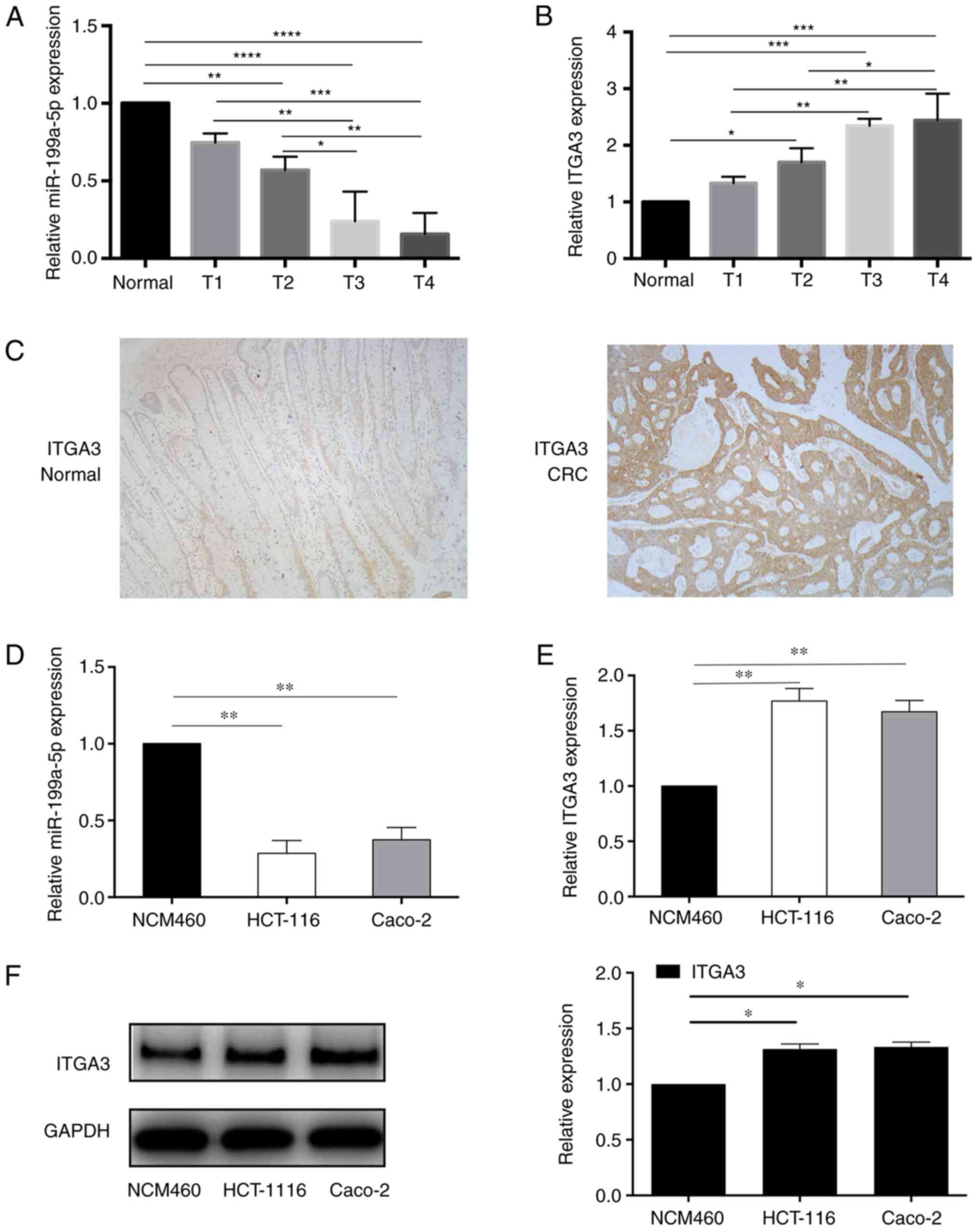 | Figure 1.miR-199a-5p and ITGA3 levels are
dysregulated in CRC tissues and cells. (A) The expression of
miR-199a-5p was significantly decreased in CRC tissues, and the
level was decreased in T2, T3 and T4 tumors compared with normal
tissues. No statistically significant difference was observed
between T1 and normal tissues. (B) RT-qPCR revealed that ITGA3
expression was upregulated in CRC tissues. ITGA3 was significantly
increased in T2, T3 and T4 tumors compared with normal tissues, No
significant difference was observed between T1 and normal tissues.
(C) Immunohistochemical staining results indicated upregulated
ITGA3 protein levels in CRC tissues compared with normal tissues.
(D) RT-qPCR demonstrated that miR-199a-5p was downregulated in
cancer cell lines HCT-116 and Caco-2. (E) RT-qPCR revealed that
ITGA3 was upregulated in cancer cell lines HCT-116 and Caco-2
compared with the normal colon epithelial cell line NCM460. (F)
Western blotting demonstrated that ITGA3 was upregulated in cancer
cell lines HCT-116 and Caco-2 compared with NCM460 cells.
*P<0.05, **P<0.01, ***P<0.001 and ****P<0.0001. CRC,
colorectal cancer; RT-qPCR, reverse transcription-quantitative PCR;
ITGA3, integrin α3β1; miR, microRNA; NC, negative control. |
IHC staining indicated the upregulation of ITGA3 at
the protein level in cancer tissues compared with normal tissues
(Fig. 1C), which was consistent
with the results detected in the cell lines (Fig. 1E and F). ITGA3 was significantly
increased in T2, T3 and T4 tumors compared with normal tissues,
although the differences between normal tissues and T1 were not
statistically significant (Fig.
1B).
These results suggested that miR-199a-5p expression
was downregulated, whereas ITGA3 was upregulated in CRC tissues and
cell lines, and their expression was partially associated with
tumor stages.
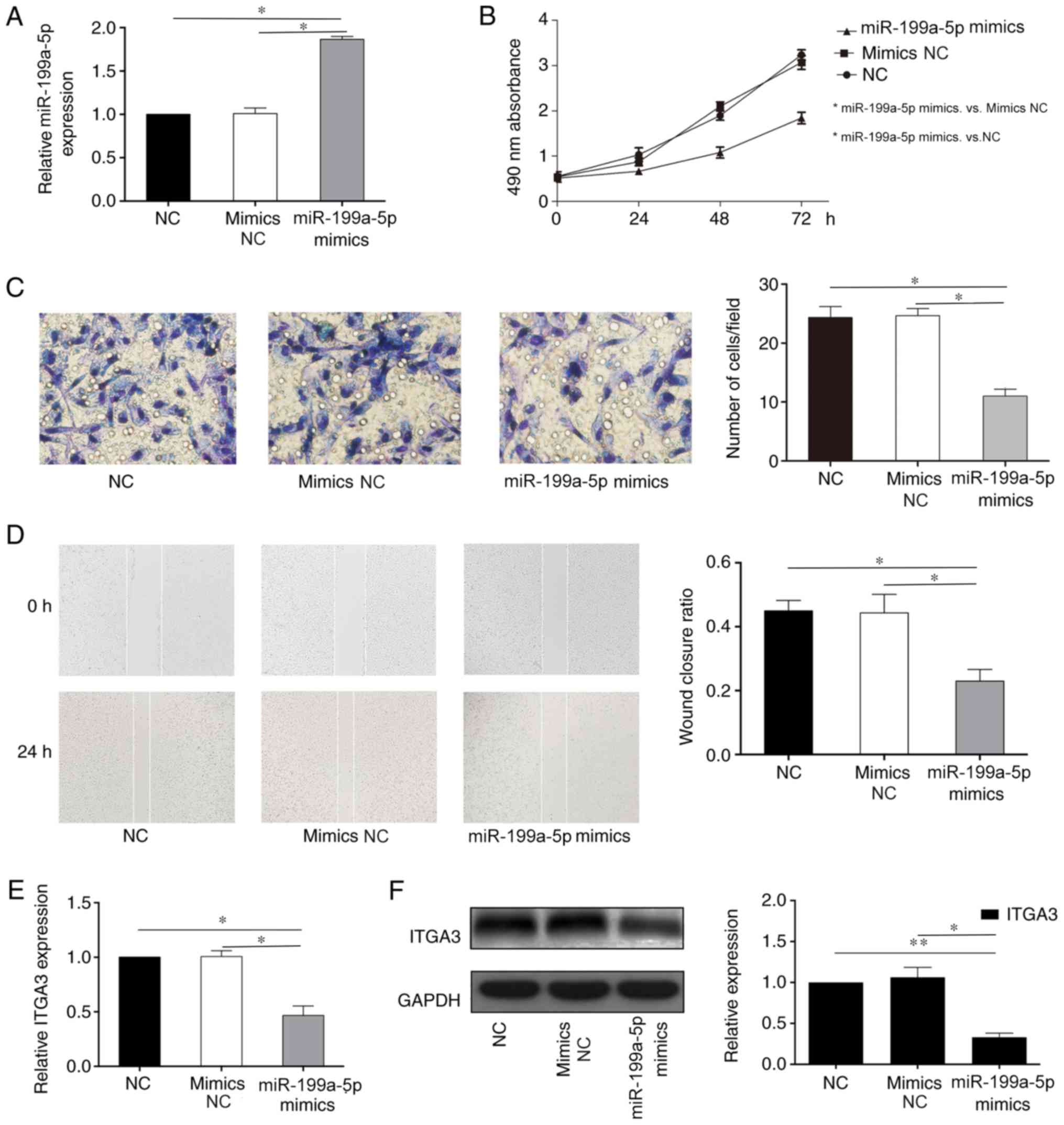
Overexpression of miR-199a-5p
suppresses the proliferation, migration and invasion of HCT116
cells
To assess the biological mechanism of miR-199a-5p in
CRC progression, HCT-116 cells were transfected with miR-199a-5p
mimics or mimics NC. RT-qPCR results suggested that miR-199a-5p
expression was significantly increased in the miR-199a-5p mimics
group compared with that in the mimics NC group (Fig. 2A). MTT, Transwell and wound healing
assays were performed to evaluate the HCT-116 cell proliferation,
invasion and migration. The MTT results demonstrated that
miR-199a-5p mimics suppressed cell proliferation compared with the
mimics NC group (Fig. 2B).
Transwell assay results indicated the inhibition on tumor cell
invasiveness by miR-199a-5p compared with the mimics NC group
(Fig. 2C). Furthermore, wound
healing assay revealed that miR-199a-5p mimics suppressed the
migratory ability of HCT116 cells compared with the mimics NC group
(Fig. 2D). These results
demonstrated that miR-199a-5p suppressed the cell proliferation,
migration and invasion in a CRC cell line. In addition,
downregulated expression of ITGA3 was observed in the miR-199a-5p
mimics group compared with the mimics NC at the mRNA and protein
levels (Fig. 2E and F), which
indicated that ITGA3 may be a direct or indirect target of
miR-199a-5p.
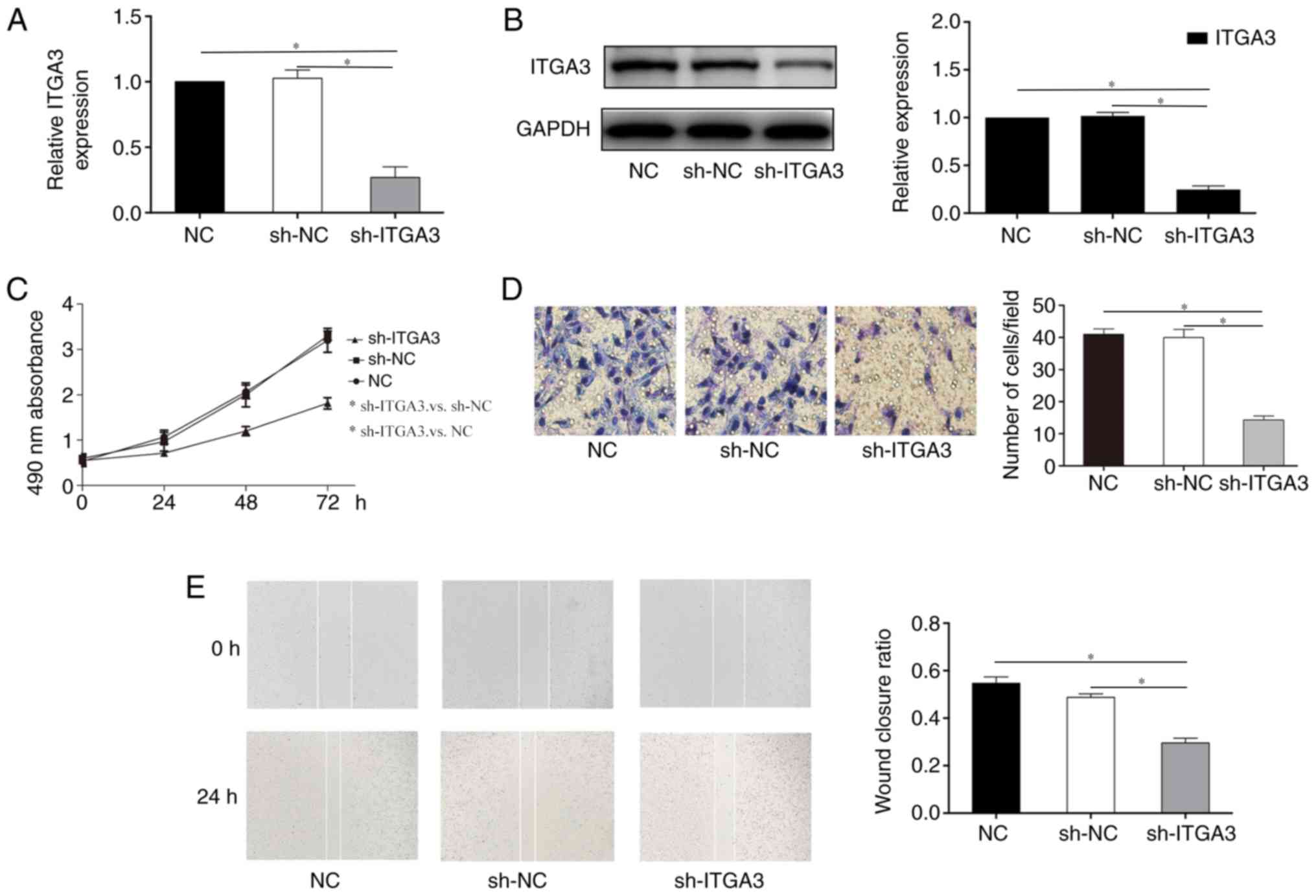
ITGA3 knockdown inhibits the
proliferation, migration and invasion in HCT-116 cells
To further investigate the potential role of ITGA3
in HCT-116 cells, cells were transfected with sh-ITGA3 or sh-NC.
RT-qPCR and western blotting demonstrated that the level of ITGA3
was significantly downregulated in cells transfected with sh-ITGA3
compared with those transfected with sh-NC (Fig. 3A and B). The MTT assay was
performed to detect cell proliferation; as presented in Fig. 3C, the proliferative ability was
remarkably decreased in cells transfected with sh-ITGA3 compared
with sh-NC. The Transwell (Fig.
3D) and wound healing (Fig.
3E) assay results revealed that the invasive and migratory
abilities were significantly decreased in the sh-ITGA3 group
compared with the sh-NC groups. These data indicated that knockdown
of ITGA3 inhibited the proliferation, migration and invasion of
HCT-116 cells.
ITGA3 3′UTR is a direct target of
miR-199a-5p
As a negative association was observed between
miR-199a-5p and ITGA3 in CRC tissues and cell lines, bioinformatics
analysis was used to predict whether miR-199a-5p bound to 3′UTR of
ITGA3 (Fig. 4A). To further
determine the connection between miR-199a-5p and ITGA3, the dual
luciferase reporter assay was performed. As presented in Fig. 4B, the luciferase activity in cells
transfected with ITGA3-WT was significantly suppressed compared
with the ITGA3-MUT group, which suggested that miR-199a-5p bound to
the 3′UTR of ITGA3 to negatively regulate ITGA3 expression.
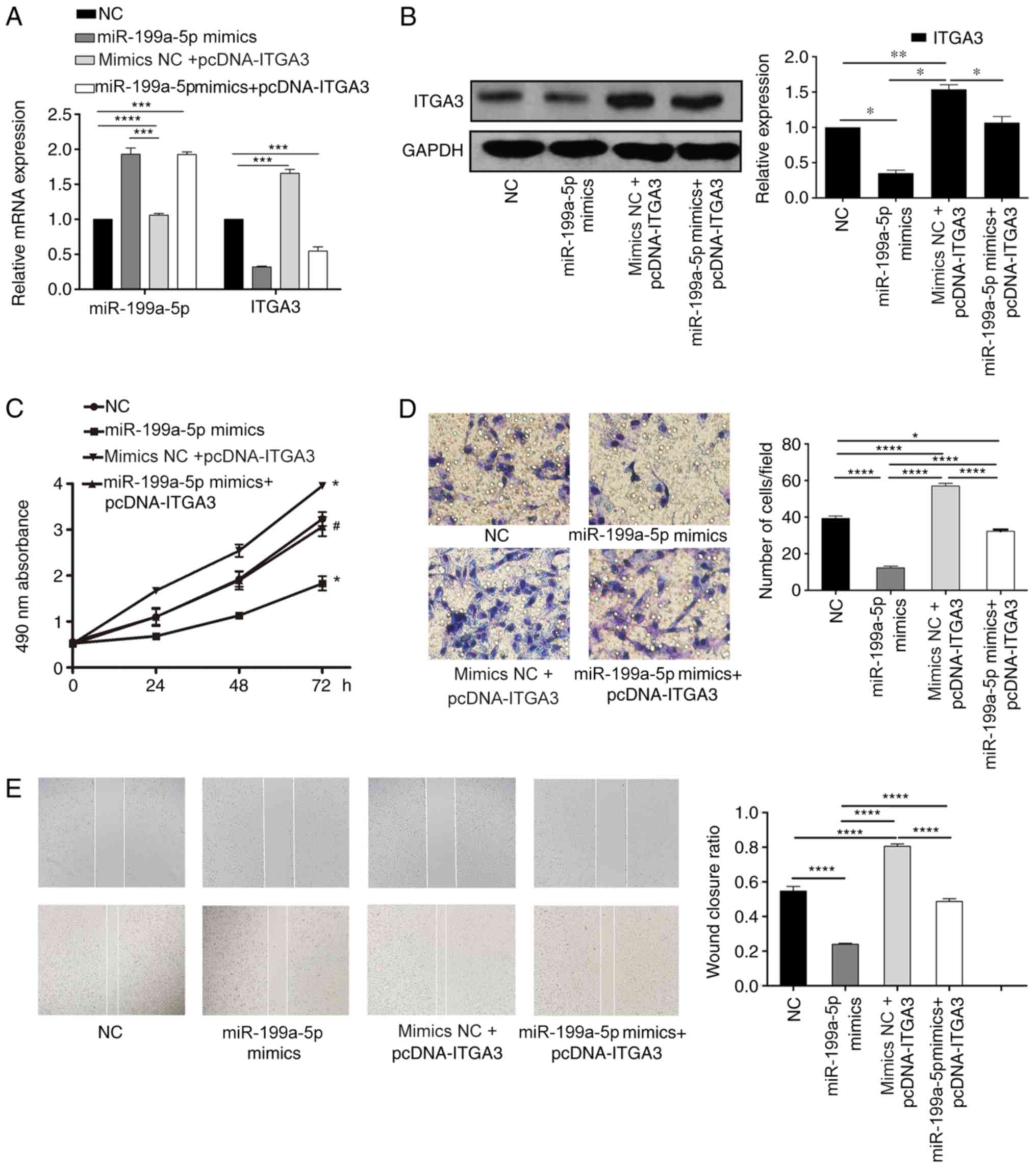
miR-199a-5p suppresses cell
proliferative, migratory and invasive abilities in CRC cells by
negatively regulating ITGA3 expression
To further confirm the proliferative, migratory and
invasive functions of the miR-199a-5p/ITGA3 axis in CRC, mimics NC
+ pcDNA ITGA3 or miR-199a-5p mimics + pcDNA ITGA3 were
co-transfected into HCT-116 cells. As presented in Fig. 5A and B, ITGA3 expression levels
were decreased in the miR-199a-5p mimics group compared with the
miR-199a-5p mimics NC + pcDNA ITGA3 group. The MTT results
demonstrated that overexpression of miR-199a-5p and ITGA3 together
increased the cell proliferation compared with that in cells
transfected with the miR-199a-5p mimics alone (Fig. 5C). The results of the Transwell and
wound healing assay also demonstrated that overexpression of ITGA3
attenuated the effects of the miR-199a-5p mimics on cell invasion
and migration, respectively (Fig. 5D
and E).
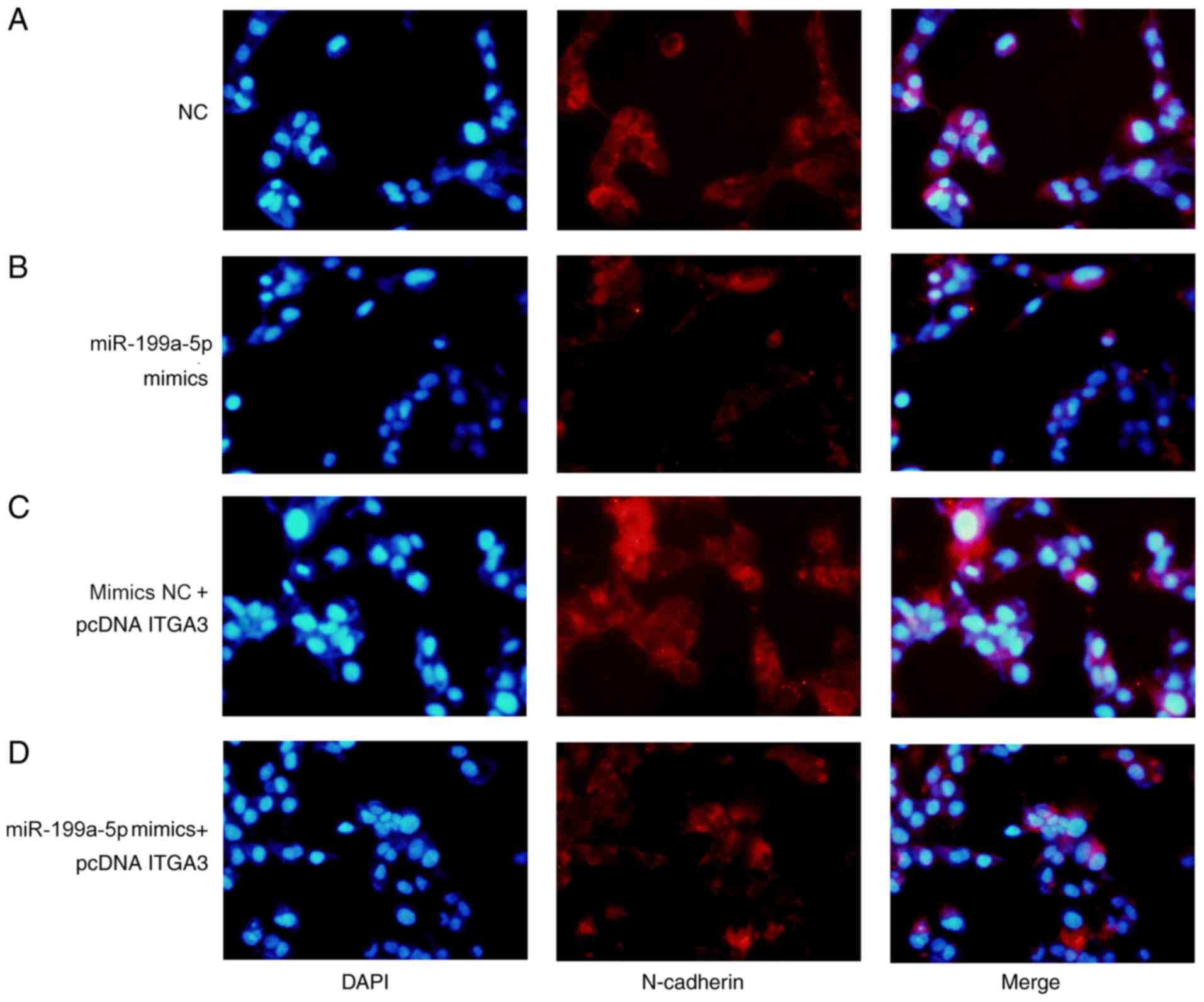
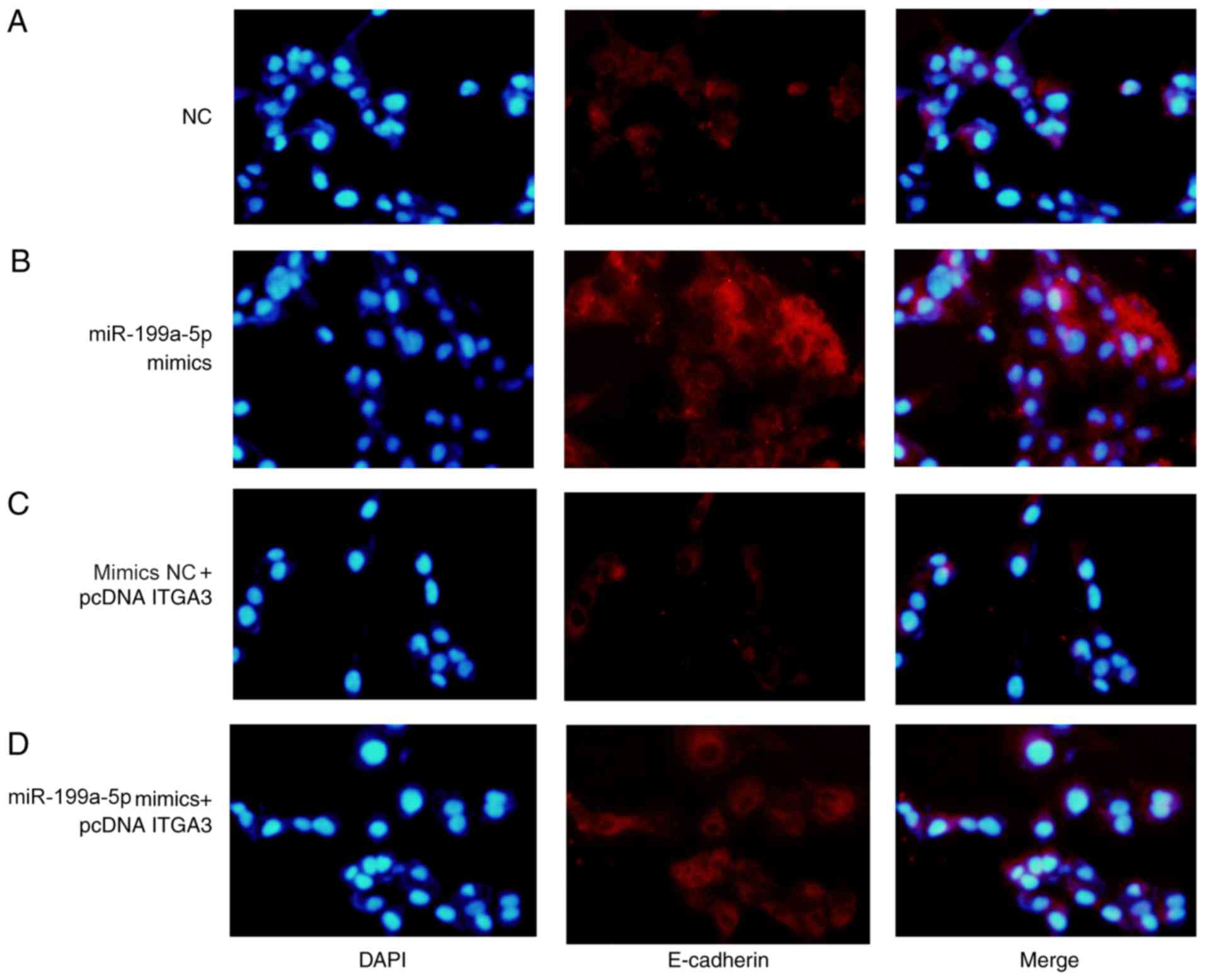
The expression levels of E-cadherin and N-cadherin
were analyzed by immunofluorescence assay. The expression of
N-cadherin increased in the miR-199a-3p mimics + pcDNA-ITGA3 group
compared with the miR-199a-3p mimics group (Fig. 6B and C), whereas the expression of
E-cadherin decreased in the miR-199a-3p mimics + pcDNA-ITGA3 group
compared with the miR-199a-3p mimics group (Fig. 7B and C). The expression levels of
E-cadherin decreased and the expression levels of N-cadherin
increased in the miR-199a-5p mimics NC + pcDNA ITGA3 group compared
with the NC group. The results suggested that ITGA3 overexpression
rescued miR-199a-3p-mediated inhibition of EMT, indicating that the
tumor suppressive activity of miR-199a-5p during CRC was
established by inhibiting the expression of ITGA3.
Discussion
Although great progress has been achieved in tumor
diagnosis and treatment, the mortality rates of various types of
cancer remain high worldwide, especially those of CRC (29). The surgical treatment of early
stage CRC can effectively increase the survival period, whereas the
prognosis for patients with late stage CRC remains in need of
further improvement (30).
Accumulating evidence has suggested that the invasion and migration
are crucial reasons for poor prognosis at the late stage of cancers
(31,32). Thus, it is crucial to investigate
specific and effective prognostic biomarkers for CRC. The
biological functions of miRNAs in the development of tumors have
attracted the attention of researchers (33). The present study focused on whether
miR-199a-5p, a rarely reported miRNA in CRC, may function as a
tumor suppressor in CRC. The results demonstrated that the
expression of miR-199a-5p was downregulated, whereas ITGA3
expression was upregulated in CRC tissues, which was consistent
with the results in CRC cells. The 3′UTR of ITGA3 was predicted to
be a binding target for miR-199a-5p via bioinformatics analysis.
Based on the above results, it was hypothesized that miR-199a-5p
may regulate the progression of colon cancer by regulating the
expression of ITGA3.
Despite the definition of miRNAs as non-coding RNAs,
they have been demonstrated to be involved in >60% of human gene
expression by participating in post-transcriptional regulation
(34). Dysregulation in miRNAs
results in a variety of diseases, including malignant tumors
(3,35). Previous studies have demonstrated
that miRNA dysregulation is involved in cell proliferation,
invasion and tumor metastasis (36,37).
Yuan et al (38) reported
that elevated miR-199a-5p promoted the apoptosis and suppressed the
proliferation in oral submucous fibrocytes, which was consistent
with the results of the present study indicating cell proliferation
inhibition. Furthermore, a previous study reported that upregulated
miR-199a-5p expression alleviated sustained endoplasmic reticulum
stress, and thus protected hepatocytes from bile acid-induced cell
death (18). The results of the
present study demonstrated that miR-199a-5p was downregulated in
CRC, and miR-199a-5p mimics suppressed CRC cell invasion, migration
and proliferation. These results were in accordance with a previous
study by Chao et al (39),
which demonstrated that downregulated miR-199a-5p was involved in
the migration and invasion of the CRC cell line DLD-1. In addition,
the present study demonstrated that ITGA3 expression levels
decreased when HCT-116 cells were transfected with miR-199a-5p
mimics, which suggested that miR-199a-5p may negatively regulate
ITGA3 expression.
As a member of the integrin family, ITGA3
participates in the Wnt/TGF-β signaling pathway among cells and
extracellular matrix. It has been reported that ITGA3 may be
involved in cell proliferation, migration and invasion. Previously,
studies have reported the association between overexpressed ITGA3
and invasion in gastric carcinomas (40,41).
Furthermore, ITGA3 has been considered as a biomarker for tongue
squamous cell carcinoma cervical lymph node metastasis (42). The results of the present study
demonstrated an increase of ITGA3 expression in CRC tissues and
cell lines, and knockdown of ITGA3 inhibited CRC cell
proliferation, invasion and migration. Furthermore, luciferase
reporter assay confirmed that ITGA3 was a direct target of
miR-199a-5p. In addition, the expression of EMT biomarkers was
investigated in the different groups, and the results suggested
that miR-199a-5p mimics significantly inhibited the EMT process of
CRC cells, whereas overexpression of ITGA3 reversed the effects of
miR-199a-5p on the EMT process. These results also supported the
tumor-suppressive role of miR-199a-5p by downregulating ITGA3.
In conclusion, the present study demonstrated that
miR-199-5p suppressed ITGA3 expression by directly binding to its
3′UTR, and thus may exert a tumor-suppressive role in CRC,
indicating that miR-199a-5p may be a potential candidate for gene
regulation-based tumor therapy. In the present study, miR-199a-5p
and IGTA3 were detected in tumor tissues and cell lines; in our
future work, the expression of miR-199a-5p will be further studied
in the blood, and the feasibility of using it as a biomarker of CRC
will be evaluated.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LT and MC performed the majority of the experiments,
wrote the manuscript and analyzed the data. QH performed the
western blotting experiments. QY and CZ designed the study and
edited the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Blomme B, Deroanne C, Hulin A, Lambert C,
Defraigne JO, Nusgens B, Radermecker M and Colige A: Mechanical
strain induces a pro-fibrotic phenotype in human mitral valvular
interstitial cells through RhoC/ROCK/MRTF-A and Erk1/2 signaling
pathways. J Mol Cell Cardiol. 135:149–159. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zhao L and Wang Z: MicroRNAs: Game
changers in the regulation of α-synuclein in Parkinson's disease.
Parkinsons Dis. 2019:17431832019.PubMed/NCBI
|
|
3
|
Zhao X, Chen GQ and Cao GM: Abnormal
expression and mechanism of miR-330-3p/BTG1 axis in hepatocellular
carcinoma. Eur Rev Med Pharmacol Sci. 23:6888–6898. 2019.PubMed/NCBI
|
|
4
|
Yang L, Hong Q, Xu SG, Kuang XY, Di GH,
Liu GY, Wu J, Shao ZM and Yu SJ: Downregulation of transgelin 2
promotes breast cancer metastasis by activating the reactive oxygen
species/nuclear factor-κB signaling pathway. Mol Med Rep.
20:4045–4258. 2019.PubMed/NCBI
|
|
5
|
Lin JS, Piper MA, Perdue LA, Rutter C,
Webber EM, O'Connor E, Smith N and Whitlock EP: Screening for
colorectal cancer: A systematic review for the U.S. Preventive
Services Task Force. Agency for Healthcare Research and Quality
(US). (Rockville, MD). 2016.
|
|
6
|
Brenner H, Kloor M and Pox CP: Colorectal
cancer. Lancet. 383:1490–1502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Koliarakis I, Psaroulaki A, Nikolouzakis
TK, Kokkinakis M, Sgantzos M, Goulielmos G, Androutsopoulos VP,
Tsatsakis A and Tsiaoussis J: Intestinal microbiota and colorectal
cancer: A new aspect of research. J BUON. 23:1216–1234.
2018.PubMed/NCBI
|
|
8
|
Lin F, Zhang P, Zuo Z, Wang F, Bi R, Shang
W, Wu A, Ye J, Li S, Sun X, et al: Thioredoxin-1 promotes
colorectal cancer invasion and metastasis through crosstalk with
S100P. Cancer Lett. 401:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: EMT: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Derynck R and Weinberg RA: EMT and cancer:
More than meets the eye. Dev Cell. 49:313–316. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu D, Li J, Li RY, Lan T, Xiao C and Gong
P: PD-L1 expression is regulated by NF-κB during EMT signaling in
gastric carcinoma. Onco Targets Ther. 12:10099–10105. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Szeder B, Tárnoki-Zách J, Lakatos D, Vas
V, Kudlik G, Merő B, Koprivanacz K, Bányai L, Hámori L, Róna G, et
al: Absence of the Tks4 scaffold protein induces
epithelial-mesenchymal transition-like changes in human colon
cancer cells. Cells. 8:13432019. View Article : Google Scholar
|
|
13
|
Zhang HD, Jiang LH, Sun DW, Li J and Ji
ZL: The role of miR-130a in cancer. Breast Cancer. 24:521–527.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mellis D and Caporali A: MicroRNA-based
therapeutics in cardiovascular disease: Screening and delivery to
the target. Biochem Soc Trans. 46:11–21. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Di Leva G, Garofalo M and Croce CM:
MicroRNAs in cancer. Annu Rev Pathol. 9:287–314. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Y, Zhang G, Wu B, Yang W and Liu Z:
miR-199a-5p represses protective autophagy and overcomes
chemoresistance by directly targeting DRAM1 in acute myeloid
leukemia. J Oncol. 2019:56134172019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qu D, Yang Y and Huang X: miR-199a-5p
promotes proliferation and metastasis and epithelial-mesenchymal
transition through targeting PIAS3 in cervical carcinoma. J Cell
Biochem. 120:13562–13572. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dai BH, Geng L, Wang Y, Sui CJ, Xie F,
Shen RX, Shen WF and Yang JM: microRNA-199a-5p protects hepatocytes
from bile acid-induced sustained endoplasmic reticulum stress. Cell
Death Dis. 4:e6042013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ahmadi A, Khansarinejad B, Hosseinkhani S,
Ghanei M and Mowla SJ: miR-199a-5p and miR-495 target GRP78 within
UPR pathway of lung cancer. Gene. 620:15–22. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhu QD, Zhou QQ, Dong L, Huang Z, Wu F and
Deng X: MiR-199a-5p inhibits the growth and metastasis of
colorectal cancer cells by targeting ROCK1. Technol Cancer Res
Treat. 17:15330346187755092018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim BK, Yoo HI, Kim I, Park J and Kim YS:
FZD6 expression is negatively regulated by miR-199a-5p in human
colorectal cancer. BMB Rep. 48:360–366. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Weber C, Alon R, Moser B and Springer TA:
Sequential regulation of alpha 4 beta 1 and alpha 5 beta 1 integrin
avidity by CC chemokines in monocytes: Implications for
transendothelial chemotaxis. J Cell Biol. 134:1063–1073. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: Biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun CC, Qu XJ and Gao ZH:
Arginine-glycine-aspartate-binding integrins as therapeutic and
diagnostic targets. Am J Ther. 23:e198–e207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nagata M, Noman AA, Suzuki K, Kurita H,
Ohnishi M, Ohyama T, Kitamura N, Kobayashi T, Uematsu K, Takahashi
K, et al: ITGA3 and ITGB4 expression biomarkers estimate the risks
of locoregional and hematogenous dissemination of oral squamous
cell carcinoma. BMC Cancer. 13:4102013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiao Y, Li Y, Liu S, Chen Q and Liu Y:
ITGA3 serves as a diagnostic and prognostic biomarker for
pancreatic cancer. Onco Targets Ther. 12:4141–4152. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang H, Cui X, Cao A, Li X and Li L:
ITGA3 interacts with VASP to regulate stemness and
epithelial-mesenchymal transition of breast cancer cells. Gene.
734:1443962020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li GF, Li ZB, Zhuang SJ and Li GC:
Inhibition of microRNA-34a protects against propofol
anesthesia-induced neurotoxicity and cognitive dysfunction via the
MAPK/ERK signaling pathway. Neurosci Lett. 675:152–159. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Heo G, Kang D, Park C, Kim SJ, Choo J, Lee
Y, Yoo JW, Jung Y, Lee J, Kim ND, et al: Pro-apoptotic effect of
the novel benzylidene derivative MHY695 in human colon cancer
cells. Oncol Lett. 18:3256–3264. 2019.PubMed/NCBI
|
|
30
|
Fu J, Wu L, Ge C, Xu T, Li D, Fu W, Wang L
and Du J: De-escalating chemotherapy for stage II colon cancer?
Therap Adv Gastroenterol. 12:17562848198675532019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luo W, Gao F, Li S and Liu L: FoxM1
promotes cell proliferation, invasion, and stem cell properties in
nasopharyngeal carcinoma. Front Oncol. 8:4832018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dhamija S and Diederichs S: From junk to
master regulators of invasion: lncRNA functions in migration, EMT
and metastasis. Int J Cancer. 139:269–280. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu TJ, Qiu P, Zhang YB, Yu SY, Xu GM and
Yang W: MiR-148a inhibits the proliferation and migration of
glioblastoma by targeting ITGA9. Hum Cell. 32:548–556. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Norouzinia M, Azodi MZ, Seyfi DN, Kardan
A, Naseh A and Akbari Z: Predication of hub target genes of
differentially expressed microRNAs contributing to infection in
gastric non-cancerous tissue. Gastroenterol Hepatol Bed Bench. 12
(Suppl 1):S44–S50. 2019.PubMed/NCBI
|
|
35
|
Zhang W, Xu J, Wang K, Tang X and He J:
miR1393p suppresses the invasion and migration properties of breast
cancer cells by targeting RAB1A. Oncol Rep. 42:1699–1708. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dai X, Liang Z, Liu L, Guo K, Xu S and
Wang H: Silencing of MALAT1 inhibits migration and invasion by
sponging miR13p in prostate cancer cells. Mol Med Rep.
20:3499–3508. 2019.PubMed/NCBI
|
|
37
|
Ye K, Xu C and Hui T: MiR-34b inhibits the
proliferation and promotes apoptosis in colon cancer cells by
targeting Wnt/β-catenin signaling pathway. Biosci Rep.
39:BSR201917992019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yuan Y, Li N, Zeng L, Shen Z and Jiang C:
Pathogenesis investigation of miR-199-5p in oral submucous fibrosis
based on bioinformatics analysis. Oral Dis. 25:456–465. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chao CC, Wu PH, Huang HC, Chung HY, Chou
YC, Cai BH and Kannagi R: Downregulation of miR-199a/b-5p is
associated with GCNT2 induction upon epithelial-mesenchymal
transition in colon cancer. FEBS Lett. 591:1902–1917. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Song B, Du J, Feng Y, Gao YJ and Zhao JS:
Co-expressed differentially expressed genes and long non-coding
RNAs involved in the celecoxib treatment of gastric cancer: An RNA
sequencing analysis. Exp Ther Med. 12:2455–2468. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ura H, Denno R, Hirata K, Yamaguchi K and
Yasoshima T: Separate functions of alpha2beta1 and alpha3beta1
integrins in the metastatic process of human gastric carcinoma.
Surg Today. 28:1001–1006. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kurokawa A, Nagata M, Kitamura N, Noman
AA, Ohnishi M, Ohyama T, Kobayashi T, Shingaki S, Takagi R; Oral
and Maxillofacial Pathology; Surgery Group, : Diagnostic value of
integrin alpha3, beta4, and beta5 gene expression levels for the
clinical outcome of tongue squamous cell carcinoma. Cancer.
112:1272–1281. 2008. View Article : Google Scholar : PubMed/NCBI
|