Introduction
Cardiovascular diseases pose a threat to public
health; recent estimates show that the incidence of hypertension
and coronary heart disease is rising (1). Although reperfusion therapy for
patients with myocardial ischemia is considered an effective
treatment, reperfusion itself may induce adverse effects on the
heart muscle (2,3). However, suppression of
apoptosis-associated pathways may improve cardiac dysfunction
(4). The morphology and
mitochondrial structure of HL-1 and H9c2 cells are similar to
primary myocardial cells in terms of biochemistry and bioenergy,
and therefore these cells can be used to simulate cardiac
ischemia-reperfusion injury in vitro (5). It is important to investigate
myocardial protection strategies and molecular mechanisms during
reperfusion.
Previous studies have shown that therapeutic
hypothermia (TH) may protect the nervous system of patients with
neonatal hypoxic ischemic encephalopathy or cardiac arrest
(6,7). Several studies have demonstrated that
TH was effective in protecting the myocardium against
hypoxia/reoxygenation (H/R)-induced damage by reducing the infarct
size (8,9). Application of TH to individuals with
acute myocardial infarction at temperatures <35°C prior to
reperfusion significantly reduced the infarct size and the release
of adverse cardiac biochemical markers (10,11).
To maximize the cardioprotective effect of TH, it is important to
understand the protective mechanisms of hypothermia, in order to
provide guidelines for managing patients with myocardial
infarction.
Post-translational modification (PTM) is crucial in
regulating protein functions and stability under physiological and
pathological conditions. Small ubiquitin-like modifier
(SUMO)ylation is a PTM process that occurs by covalently and
reversibly conjugating SUMO proteins with their target proteins
(12). Previous studies have
demonstrated the effects of SUMOylation on the control of several
cellular functions, including cell cycle control, apoptosis,
epigenetic management and transcription (13–15).
Similarly, an imbalance in the SUMOylation/de-SUMOylation cycle has
been implicated in several diseases (16). Previous studies have reported that
multiple signaling pathways serve a role in protein SUMOylation,
which mostly affect cardiac function and development (17,18).
SUMOylation is involved in cancer, abnormal cardiac development and
cerebral ischemia (16), and
promotes adaptation of the heart to pathological stress (19,20),
suggesting that it is a valuable target for treating cardiovascular
disorders (21,22). Thus, SUMOylation may have a role in
controlling the response of the heart to hypoxic-ischemic stress.
However, the protective roles of SUMOylation in H/R-induced
myocardial cell injury have not been fully investigated. Therefore,
it is imperative to examine the mechanisms of myocardial cell fate
regulated by SUMOylation under physiological and pathological
conditions.
Previous studies have described the link between
hypothermia and SUMOylation in the brain, and indicated that
hypothermia may regulate SUMOylation to provide protection to the
nervous system during hypoxic ischemia (23,24).
However, it is unknown whether hypothermia alters SUMOylation
modifications in the heart. A previous study revealed that
SUMO-conjugated protein levels rose to varying degrees in organs in
rats subjected to extracorporeal circulation at 18°C (25). Currently, there is no systematic
understanding of how moderate hypothermia contributes to
SUMOylation modifications in the heart. Furthermore, several
aspects of cellular mechanisms responsible for the cardioprotective
effects of hypothermia are relatively unknown. To further
understand the mechanisms underlying hypothermia-induced
cardioprotection, it is necessary to identify whether
hypothermia-induced SUMOylation occurs in the heart.
The present study established an in vitro
myocardial H/R model and detected the protein expression levels of
SUMO1, SUMO2/3, caspase-3 and Bcl-2. In addition, changes in
reactive oxygen species (ROS) production and mitochondrial membrane
potential were evaluated. The present results suggested that
moderate hypothermia enhanced SUMOylation in cardiomyocytes and
antagonized H/R-induced apoptosis. Thus, the present study
indicated the treatment possibilities of moderate hypothermia for
H/R injury.
Materials and methods
Cell culture
HL-1 cardiomyocytes from a murine atrial tumor were
provided by American Type Culture Collection (ATCC). The cells were
grown at 37°C with 5% CO2 in Claycomb medium
(Sigma-Aldrich; Merck KGaA) containing 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.), 100 U/ml penicillin, 100 µg/ml
streptomycin, 2 mM L-glutamine and 0.1 mM norepinephrine. H9c2
embryonic rat heart cells were purchased from ATCC and grown in
DMEM (Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS at
37°C in a 5% CO2−95% air atmosphere. Cells were passaged
to 80–90% confluence at ratios of 1:3 or 1:4 using trypsin with
0.05% EDTA.
Small interfering RNA (siRNA)
transfection
H9c2 and HL-1 cells were cultured in complete medium
for 12 h. Transfections were performed using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) by incubating cells (5.6×104
cells/well) with the respective siRNA (10 µM; developed and
prepared by Shanghai GenePharma Co., Ltd.) for 48 h. SUMO1 siRNA
sequences were as follows: SUMO1 siRNA-1
(5′-TGACAACACATCTCAAGAA-3′), SUMO1 siRNA-2
(5′-GACAGGGTGTTCCAATGAA-3′), SUMO1 siRNA-3
(5′-TCTTTGAGGGTCAGAGGA7-3′) and negative control siRNA,
5′-UUCUCCGACCGUGUCACGUTT-3′. In addition, SUMO1 full-length cDNA
was subcloned into the mammalian expression plasmid pcDNA3.1
(Thermo Fisher Scientific, Inc.) with a Flag tag at the C-terminus.
The Flag-SUMO1 plasmid and the empty vector (0.8 µg) were
transiently transfected into HL-1 and H9c2 cells
(0.5–2×105 cells/well) using Lipofectamine®
2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's instructions. All experiments were conducted
48 h post-transfection.
H/R
H9c2 and HL-1 cells were cultured with low-glucose
DMEM and subjected to chemical hypoxia using different
concentrations of CoCl2 (50, 100, 300 and 600 µM;
HEBIO). After 12 h of CoCl2-induced hypoxia (37°C),
reoxygenation was achieved in complete medium with 10% FBS for 12 h
(37°C). Cells were subjected to 12 h of hypoxia and 12 h of
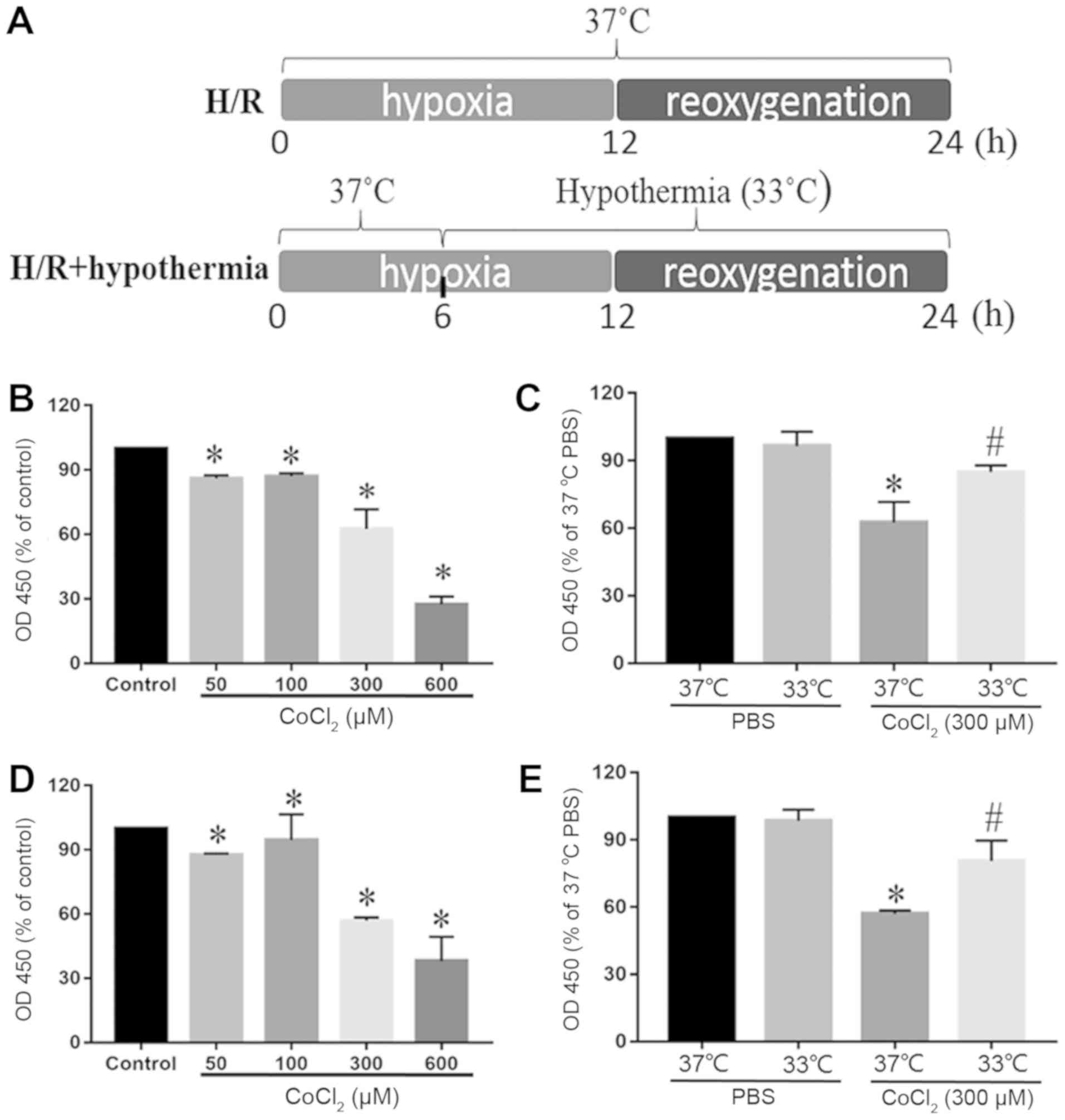
reoxygenation (H/R). The schematic illustration of the H/R
experimental protocol is shown in Fig.
1A. Cells were cultured under moderate hypothermia (33°C) for 6
h before reoxygenation. Control cells were cultured in DMEM
supplemented with 10% FBS at 37°C in a 5% CO2−95% air
atmosphere. All media were supplemented with 100 µM/ml
penicillin/streptomycin and 2 mM L-glutamine.
Moderate hypothermia protocol
The European Resuscitation Council for cardiac
arrest survivors guidelines (26)
were used in the present study. After 6 h of hypoxia, the
temperature of the CO2 incubator was adjusted and
maintained at 33°C during simulated reperfusion in the TH group of
cells. During 12 h of reoxygenation, H9c2 and HL-1 cells were
incubated in DMEM containing 10% FBS at 33°C with 5%
CO2−95% air. The temperature of control group cells was
maintained at 37°C. After 24 h the samples were analyzed. The
schematic illustration of the H/R and hypothermia experimental
protocol is shown in Fig. 1A.
Cell viability assay
Cell viability was examined using a Cell Counting
Kit-8 (CCK-8) (Beijing Solarbio Science & Technology Co., Ltd.)
assay. H9c2 and HL-1 cells were cultured in 96-well plates
(2×103 cells/well) and were treated as aforementioned.
Subsequently, 10 µl CCK-8 solution was added to each well and
incubated at 37°C for 2 h. A microplate reader (VersaMax Absorbance
microplate reader; Molecular Devices) was used to analyze the
absorbance at 450 nm. To increase the reliability of measures, each
experiment was performed three times.
Determination of intracellular ROS
levels
ROS was detected using the fluorescent probe
2′7′-dichlorodihydrofluorescein diacetate (DCFH-DA; Beijing
Solarbio Science & Technology Co., Ltd.). DCFH-DA has no
fluorescence and can freely penetrate the cell membrane. In the
presence of ROS, DCFH is oxidized into fluorescent DCF. Thus, the
fluorescence intensity and the level of intracellular ROS are
directly proportional. After H/R, 10 µM DCFH-DA was added to cells
at 37°C for 30 min. The cells were then washed three times with
serum-free cell culture medium. The fluorescence of DCF was
detected using a microplate reader (Gemini XPS microplate reader;
Molecular Devices) at excitation and emission wavelengths of 488
and 525 nm, respectively, to measure intracellular ROS levels.
Detection of changes in the
mitochondrial membrane potential
The fluorescent probe
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzi-midazolylcarbocyanine
iodide (JC-1; Beijing Solarbio Science & Technology Co., Ltd.)
was used to measure the mitochondrial membrane potential. After
incubation at 37°C for 20 min in a cell incubator with 1 ml JC-1
staining solution, the cells were washed two times using JC-1
staining buffer according to the manufacturer's instructions.
Changes in laser-induced fluorescence of JC-1 were detected by
confocal microscopy (Olympus Corporation).
Western blot analysis
H9c2 and HL-1 cells were washed twice with cold PBS.
To avoid de-SUMOylation of proteins during sample preparation,
cells were lysed for 30 min with 120 µl ice-cold RIPA lysis buffer
(Beijing Solarbio Science & Technology Co., Ltd.) with 100:1
phenylmethanesulfonyl fluoride and 10:1 N-ethlmaleimide. The
protein supernatant was collected after centrifugation at 10,000 ×
g at 4°C for 15 min. Protein concentration was measured using a
bicinchoninic acid protein assay (Beijing Solarbio Science &
Technology Co., Ltd.). A mixture of the samples and 4X SDS-PAGE
loading buffer was incubated for 5 min at 95°C, then 40 µg/lane
samples underwent SDS-PAGE on 12% gels. Subsequently, proteins were
transferred to a PVDF membrane at 110 mV for 2 h. The membrane was
blocked for 1 h at room temperature using Tris-HCl containing 0.1%
Tween-20 and 5% skim milk powder (Thermo Fisher Scientific, Inc.),
followed by overnight incubation at 4°C with anti-SUMO1 antibody
(cat. no. ab11672; 1:1,000; Abcam), anti-SUMO2/3 antibody (cat. no.
ab3742; 1:1,000; Abcam), anti-caspase-3 antibody (cat. no. ab13847;
1:500; Abcam), anti-Bcl-2 antibody (cat. no. ab692; 1:500; Abcam)
and anti-GAPDH antibody (cat. no. ab8245; 1:1,000; Abcam). The
membranes were incubated at room temperature for 1 h with goat
anti-rabbit IgG (cat. no. 111-035-003; 1:2,000; Jackson
ImmunoResearch Laboratories, Inc.) or goat anti-mouse IgG (cat. no.
115-035-003; 1:2,000; Jackson ImmunoResearch Laboratories, Inc.) as
the secondary antibody. Subsequently, blotted membranes were
visualized with enhanced chemiluminescence reagents (EMD
Millipore). Protein bands were detected using a C-DiGit Blot
Scanner (LI-COR Biosciences) and ImageJ software (version
1.8.0.112; National Institutes of Health) was used to normalize the
data.
Statistical analysis
All HL-1 and H9c2 cell culture dishes were randomly
assigned to different experimental groups (n=5). Data are presented
as the mean ± SD. Statistical analysis was performed using GraphPad
Prism 6.0 software (GraphPad Software, Inc.). Statistical
significance was determined using a one-way ANOVA followed by
Tukey's test or two-way ANOVA followed by Bonferroni's test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of different concentrations of
CoCl2 on cell viability
CCK-8 assay was used to analyze how different
concentrations of CoCl2 (50, 100, 300 and 600 µM)
affected the cytotoxicity of myocardial cells. The viability of
HL-1 and H9c2 cells was reduced significantly with CoCl2
doses >300 µM. Moreover, with 50 and 100 µM CoCl2,
cell viability was decreased slightly compared with the control
group (Fig. 1B and D). In
addition, the cytotoxic damage was greatest at 600 µM. It was found
that at 300 µM, moderate TH (33°C) antagonized H/R-induced
cytotoxicity of cardiomyocytes (Fig.
1C and E).
Moderate TH antagonizes the decrease
in mitochondrial membrane potential and ROS production triggered by
H/R in cardiomyocytes
In both the brain and heart, the pathological
mechanisms of H/R-induced injury are homoplastic, and involve
numerous processes such as mitochondrial dysfunction, intracellular
acid-base disturbance and disrupted ion homeostasis (27,28).
During hypoxic ischemia, mitochondrial dysfunction can cause
insufficient ATP production and produce excessive ROS, which impair
the homeostasis of ions in cardiac cells and excitability of the
cytomembrane (29). To establish
whether TH has a protective function in mitochondria, the present
study established a H/R model in vitro. ROS production was
quantified by flow cytometry (Fig.
2A). It was revealed that after exposure to moderate TH, the
production of mitochondrial ROS was significantly reduced compared
with in the H/R group (Fig. 2B).
Changes in laser-induced fluorescence of JC-1 were detected by
confocal microscopy to assess the early variation of mitochondrial
membrane potential (Fig. 2C). It
was demonstrated that exposure of cells to moderate TH
significantly increased the mitochondrial membrane potential
compared with in the H/R group (Fig.
2D). Thus, the present results suggested that moderate TH
improved mitochondrial functions.
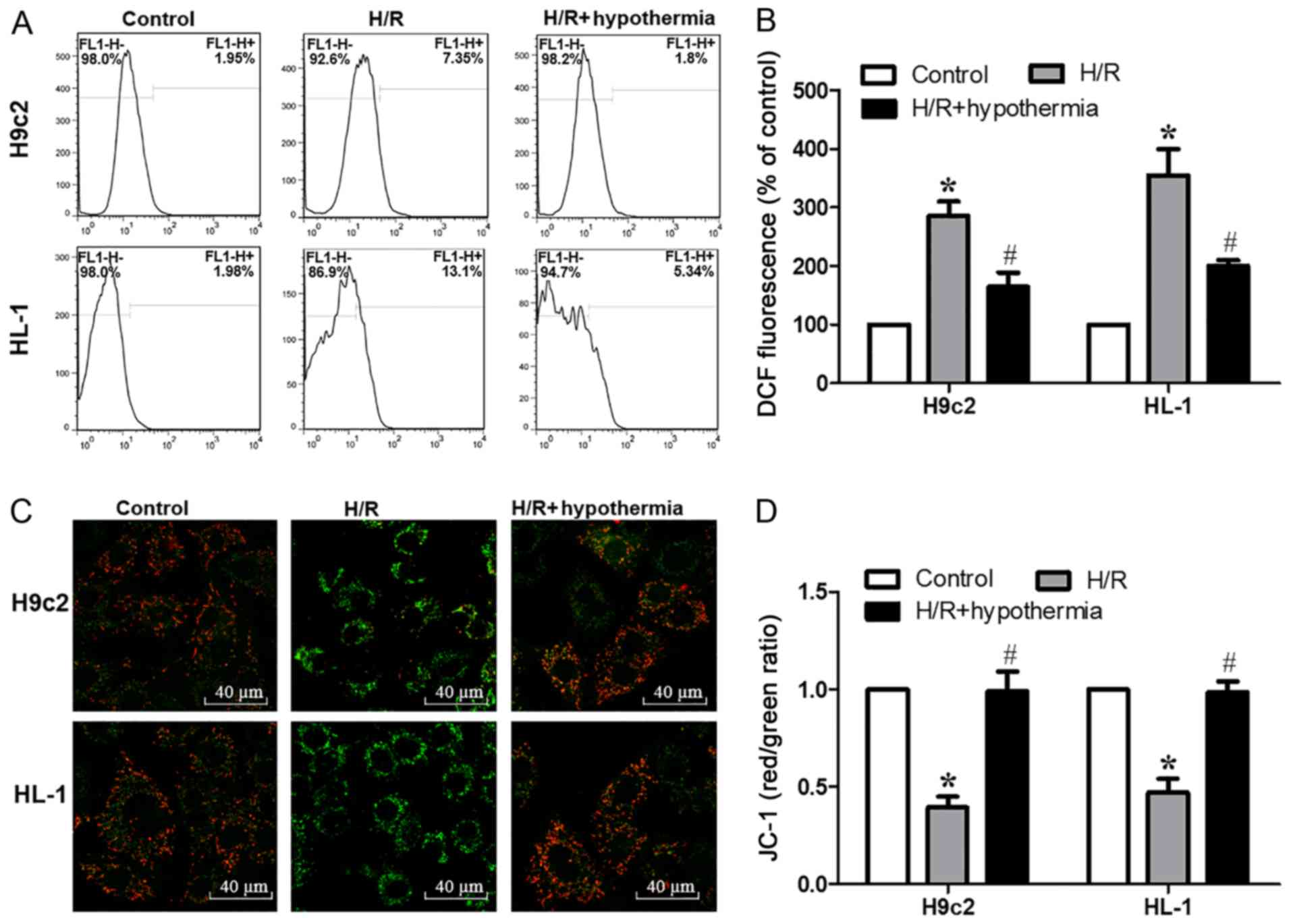 | Figure 2.Moderate hypothermia antagonizes the
decrease in mitochondrial membrane potential and the production of
ROS triggered by H/R in cardiomyocytes. (A) Flow cytometric
analysis of ROS generation. Data are presented as the mean ± SE,
n=5. (B) Summarized data of DCF fluorescence in HL-1 and H9c2 cells
expressed as % of baseline. (C) Confocal fluorescence images
(magnification, ×60) of H9c2 and HL-1 cells loaded with JC-1. Scale
bar, 40 µm. (D) Quantified data showing the JC-1 ratio. *P<0.05
vs. control; #P<0.05 vs. H/R. DCF,
dichlorofluorescein; H/R, hypoxia/reoxygenation; JC-1,
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzi-midazolylcarbocyanine
iodide; ROS, reactive oxygen species. |
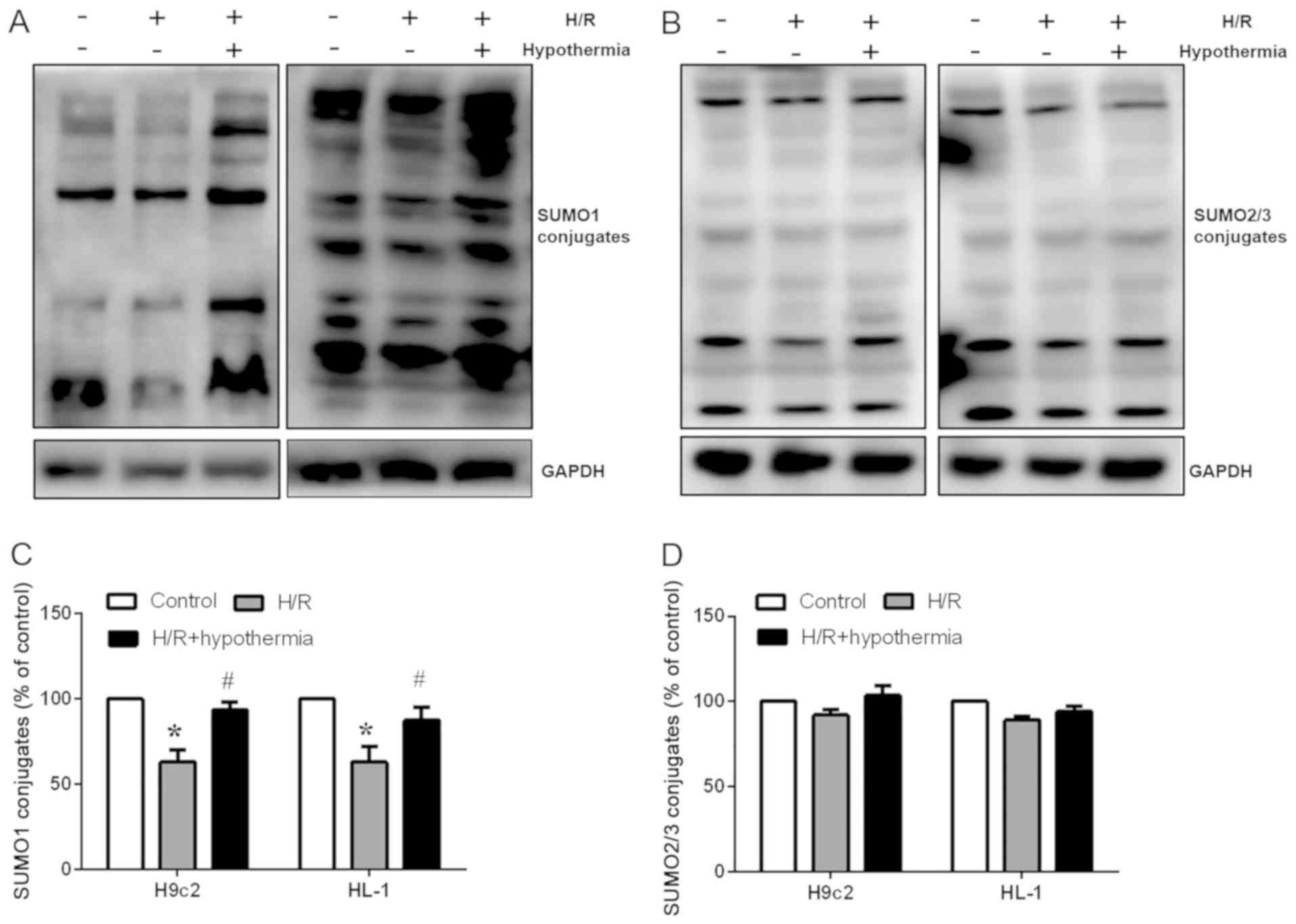
Moderate TH affects protein
SUMOylation after myocardial H/R injury
Further experiments were performed in the present
study to investigate the signaling pathways involved in protein
SUMOylation. SUMOylation is an important cellular process. Previous
studies have investigated the relationship between moderate
hypothermia and SUMOylation, and have suggested that moderate
hypothermia can trigger SUMOylation, which is an active process
that regulates protein function in vitro (23,30).
Therefore, the aim of the present study was to determine how
moderate TH affected protein SUMOylation in the heart. Western
blotting (Fig. 3A and B) was used
to detect the expression of SUMO1 and SUMO2/3 proteins, and
SUMOylation of cardiomyocytes was evaluated in vitro. It was
revealed that exposure of cells to H/R decreased the protein
expression levels of SUMO1 compared with in the control group
(Fig. 3A and B). TH during
reoxygenation partly reversed the reduction in SUMO1 protein
expression in the H/R group (Fig.
3C). However, the protein expression levels of SUMO2/3 were not
significantly altered (Fig. 3D).
The present results indicated how hypothermia may regulate
SUMOylation levels and suggested that, following moderate TH,
production of new SUMO1 conjugates may have cytoprotective
effects.
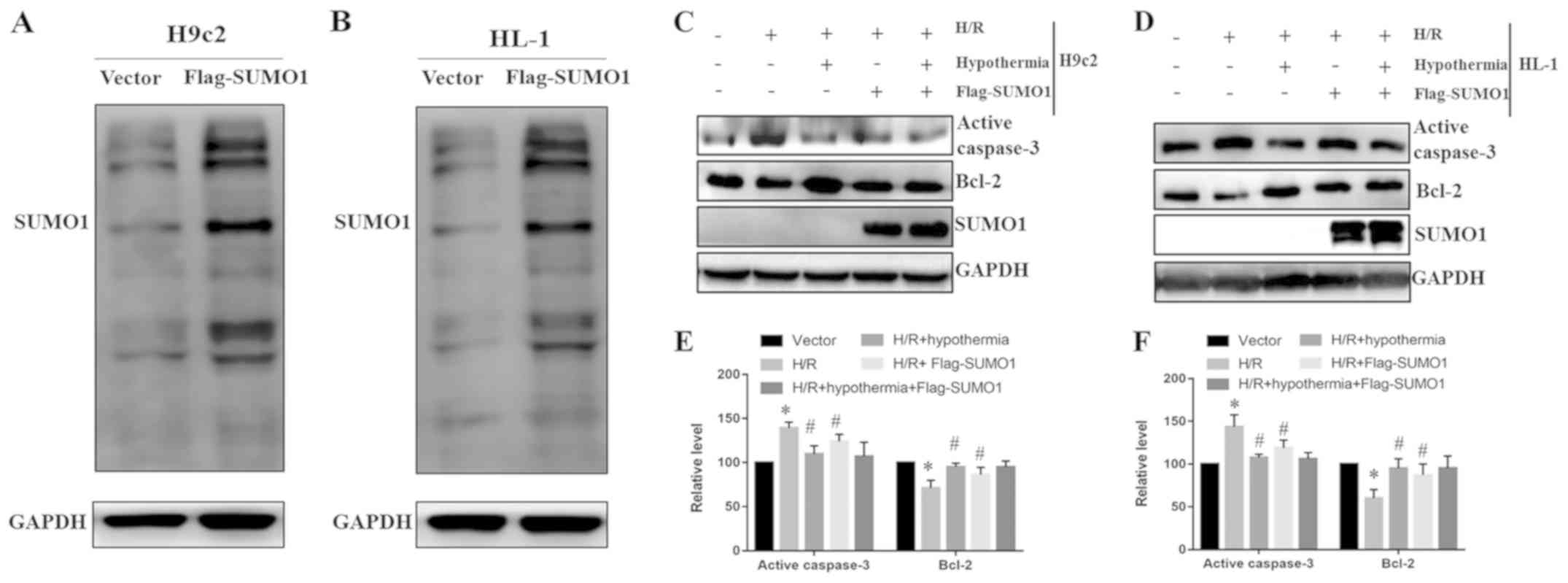
Overexpression of SUMO1 strengthens
the protective effect of TH on cardiomyocytes under H/R
conditions
The present study transfected H9c2 and HL-1 cells
with Flag-SUMO1 to induce overexpression of SUMO1 under normal
conditions. Western blotting results indicated that SUMO1
overexpression was effective (Fig. 4A
and B). Western blotting was also used to determine the protein
expression levels of caspase-3 and Bcl-2 in H9c2 (Fig. 4C) and HL-1 (Fig. 4D) cells. The present results
suggested that SUMO1 overexpression in the H/R + Flag-SUMO1 group
reduced the expression levels of caspase-3 and increased the
expression levels of Bcl-2 compared with the H/R group (Fig. 4E and F). In addition, the similar
effect of moderate TH treatment and overexpression of SUMO1 reduced
the expression levels of caspase-3 and increased the expression
levels of Bcl-2 compared with H/R group of H9c2 and HL-1 cells
(Fig. 4E and F). Therefore, the
present results suggested that the protective effects of moderate
TH may be potentiated through SUMO1 overexpression.
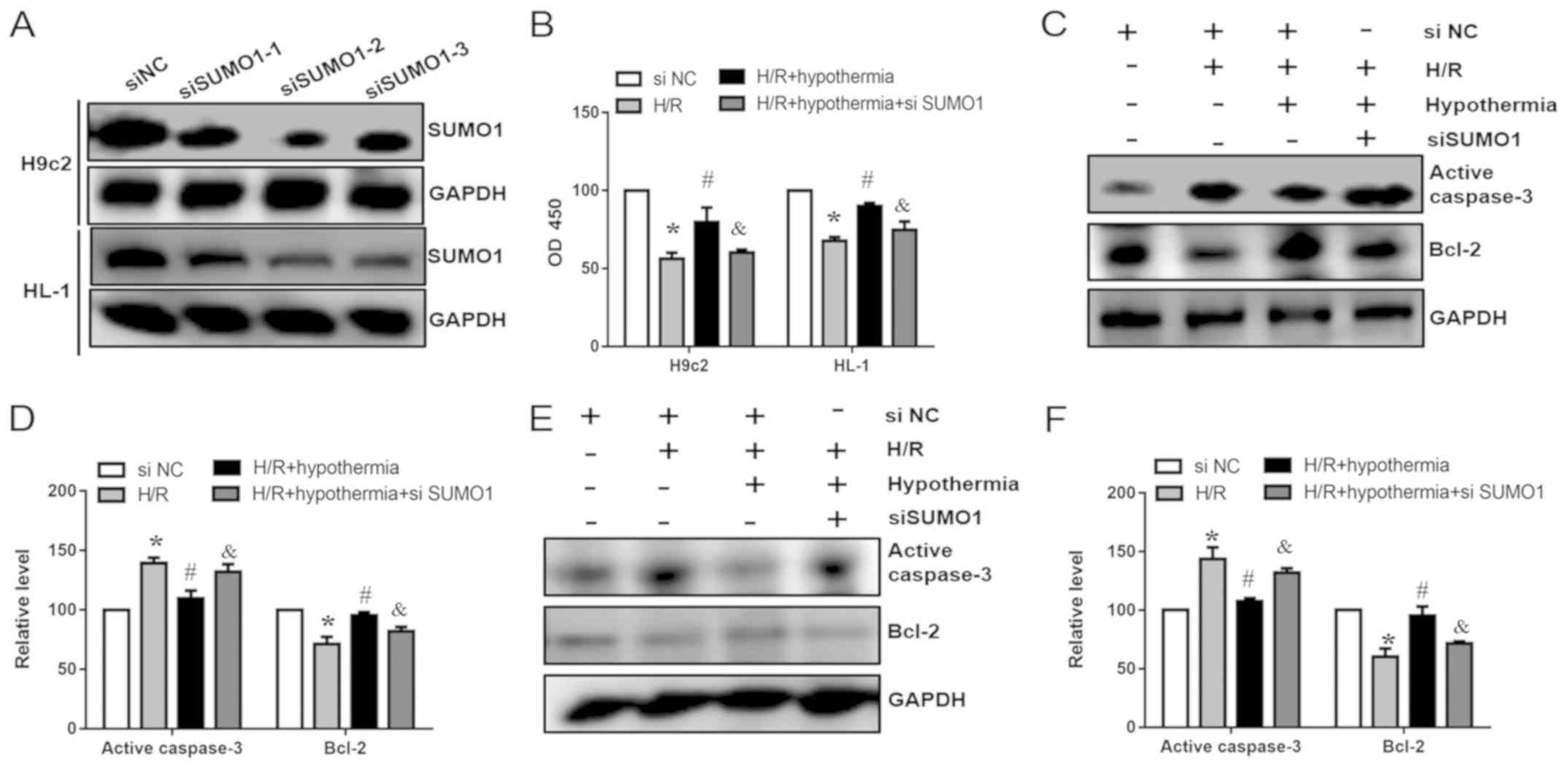
Knockdown of SUMO1 weakens the
protective effect of TH in cardiomyocytes under H/R conditions
To further investigate whether overexpression of
SUMO1 can reduce cardiomyocyte apoptosis, siRNAs (si-SUMO1-1,
si-SUMO1-2 and si-SUMO1-3) were used to knock down the expression
of SUMO1 in HL-1 and H9c2 cells. Post-transfection with si-SUMO1,
the protein expression levels of SUMO1 were decreased compared with
in the si-NC groups (Fig. 5A). In
addition, cell viability was significantly decreased compared with
in the H/R + hypothermia groups (Fig.
5B). As SUMO1 siRNA-2 had the highest efficiency, HL-1 and H9c2
cells were transfected with SUMO1 siRNA-2 and then subjected to TH.
Western blotting was performed to assess the protein expression
levels of caspase-3 and Bcl-2. In the present study, it was
demonstrated that SUMO1 knockdown and hypothermia increased the
expression levels of caspase-3 and reduced the expression levels of
Bcl-2 compared with in the hypothermia group without siRNA
transfection (Fig. 5C-F).
Collectively, the present results suggested that SUMO1 affected
apoptosis of H9c2 and HL-1 cells following exposure to moderate
TH.
Under moderate TH, SUMO1 knockdown
reduces the mitochondrial membrane potential and enhances ROS
production in cardiomyocytes
To further demonstrate that TH antagonizes
cardiomyocyte apoptosis by enhancing SUMOylation, SUMO1 was knocked
down, and the mitochondrial membrane potential (Fig. 6A) and ROS in cardiomyocytes were
measured. It was demonstrated that SUMO1 knockdown significantly
increased mitochondrial ROS levels (Fig. 6B) and significantly decreased the
mitochondrial membrane potential (Fig.
6C) compared with in the H/R + hypothermia group. Therefore,
the present results suggested that SUMO1 knockdown reversed the
protective effect of TH on cardiomyocytes.
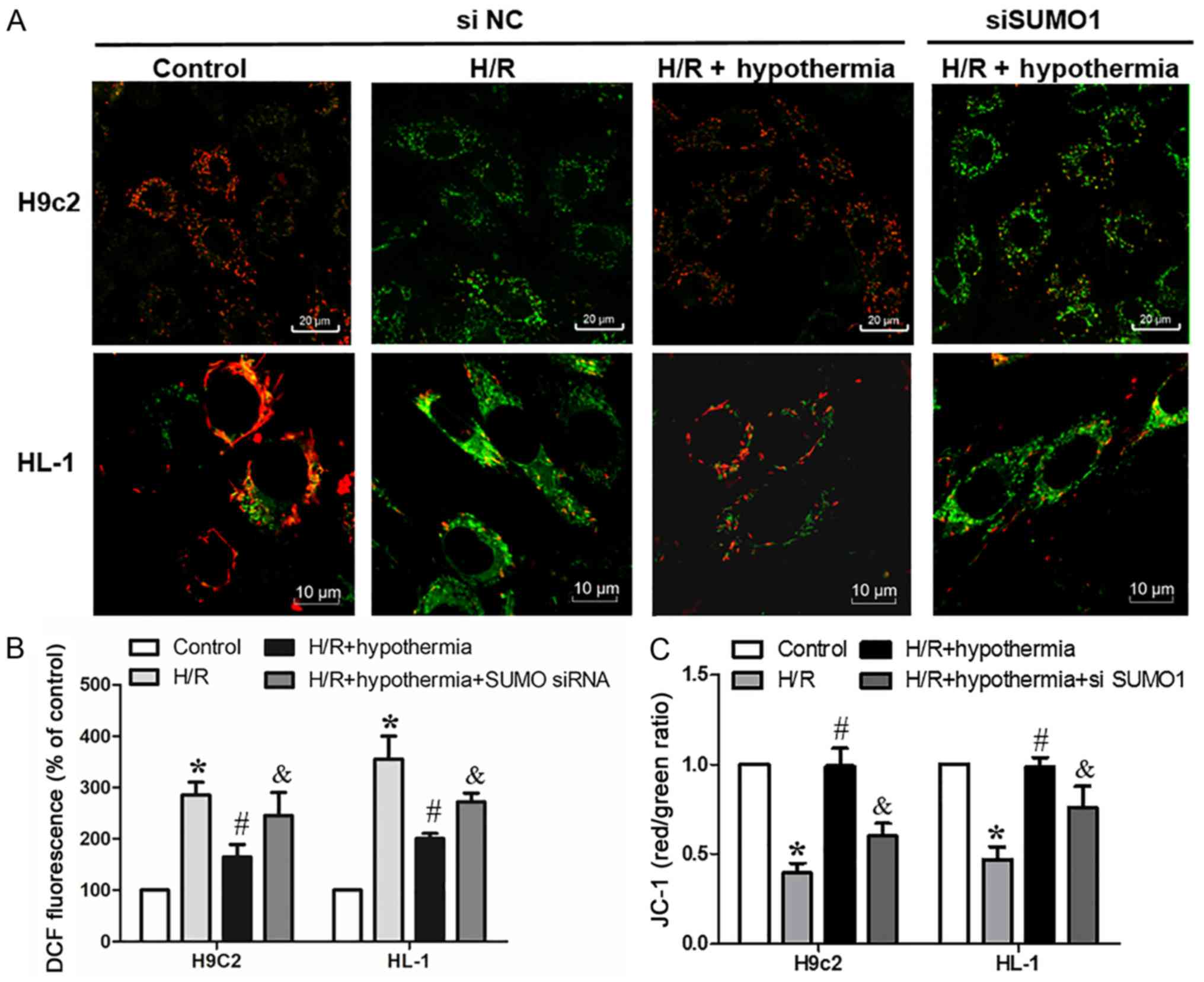 | Figure 6.Under moderate hypothermia
conditions, SUMO1 knockdown decreases mitochondrial membrane
potential and enhances the production of reactive oxygen species in
cardiomyocytes. (A) Confocal fluorescence images of HL-1
(magnification, ×100) and H9c2 (magnification, ×60) cells loaded
with JC-1. Scale bars, 20 and 10 µm. (B) Fluorescence of H9c2 and
HL-1 cells loaded with DCF. (C) Quantified data showing the JC-1
ratio. *P<0.05 vs. control (NC siRNA); #P<0.05 vs.
H/R (NC siRNA); &P<0.05 vs. H/R + hypothermia (NC
siRNA). DCF, dichlorofluorescein; H/R, hypoxia/reoxygenation; NC,
negative control; JC-1,
5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzi-midazolylcarbocyanine
iodide; siRNA, small interfering RNA; SUMO, small ubiquitin-like
modifier. |
Discussion
While previous studies have demonstrated the
importance of applying moderate hypothermia in simulated hypoxic
ischemia to attenuate apoptosis of cardiomyocytes, the mechanisms
by which moderate hypothermia reduces cell death has not been fully
elucidated (29). In the hypoxic
ischemia model of HL-1 and H9c2 cardiomyocytes, the application of
hypothermia treatment can effectively maintain the integrity and
function of the mitochondrial membrane of cells (31,32).
In the present study, it was demonstrated that hypothermia
treatment during reoxygenation reduced ROS release and enhanced
mitochondrial membrane potential, as indicated by flow cytometric
measurement of the intensity of DCF fluorescence and JC-1
detection. Furthermore, these indicators were not significantly
different between HL-1 and H9c2 myocardial cells, which is
inconsistent with results that HL-1 cells may be more resistant to
hypoxia than H9c2 cells (5),
possibly because these two cell lines have a similar tolerance to
H/R injury. Therefore, the present results support the hypothesis
that moderate hypothermia attenuates mitochondrial dysfunction and
preserves the integrity of the mitochondrial membrane, which is
important for inhibiting apoptosis following H/R injury.
A number of signaling pathways are influenced by
SUMOylation (13). SUMOylation
plays a role in the progression of several human diseases, such as
heart failure and abnormal heart development (16). Previous studies demonstrated that
congenital heart defects and retarded cardiac development occurred
in mice with SUMO1 knockdown and in mice with SUMO-specific
peptidase 2 overexpression (18,33).
However, the kinetics of SUMO2/3 and SUMO1 changes during
myocardial injury are not fully understood. SUMO1 and SUMO2/3 are
coupled by common enzymes and their modification effects on some
proteins overlap; these proteins can also specifically target
certain proteins and exhibit diverse kinetics (17,34,35).
Almost all SUMO1 binds to proteins under physiological conditions
(36). Under stress conditions,
the dynamic conjugation of SUMO2/3 is increased, which prevents its
interaction with target proteins (37). Moreover, in the brain, a strong
relationship between SUMO2/3 modifications and neuroprotection has
been identified (38,39). In addition, it has been reported
that SUMO1 and SUMO2/3 protein modifications are increased during
the early stage of myocardial I/R injury (40). However, to the best of our
knowledge, no previous study has reported the detailed molecular
mechanism of SUMO1 and SUMO2/3 modifications during myocardial
H/R-induced injury. In the present study, an increase in SUMO1
protein modification in the early stage of hypothermia was observed
when myocardial H/R injury was initiated. This is similar to
previous studies, in which the effect of SUMO1 on preventing
harmful ventricular remodeling and abnormal myocardial cell
functional response was reported (41,42).
However, no differences were found in SUMO2/3 protein modifications
in the present study, which is inconsistent with previous studies
that showed an early increase in SUMO2/3 protein expression
following myocardial H/R injury (43). The cause of this discrepancy is
unclear but may be due to the binding of different types of
proteins to SUMOs in different tissues and organs, or due to
differences in degrees of binding between target proteins and SUMOs
during hypothermia.
It has been reported that TH as an adjunctive
therapy to treat patients with acute myocardial infarction had no
significant clinical advantage (44). The hypothesis that moderate
hypothermia can stop the apoptosis of cardiomyocytes in response to
H/R-induced injury via multiple pathways has been questioned based
on results from conflicting studies. Previous work demonstrated
that moderate hypothermia provided cardioprotection after injury
triggered by H/R in HL-1 cardiomyocytes by reducing the activation
of caspase-3 (45). In addition,
hypothermia may maintain integrity of the mitochondrial membrane,
thus decreasing the release of cytochrome c and apoptosis-inducing
factor, which minimizes caspase-dependent and caspase-independent
cell death (29). Bcl-2 proteins
largely control mitochondria-associated release of apoptotic
factors, but data on the expression of Bcl-2 in response to
cryogenic regulation is limited. It was previously reported that
moderate hypothermia after ischemia significantly decreased Bax
protein expression, and increased Bcl-2 expression levels in a
human umbilical vein endothelial cell model (46). In addition, cardiac arrest liquid
cooling to 20°C for 20 min in H9c2 cardiac muscle cells increased
Bcl-2 expression levels (31).
Moreover, in vitro rabbit heart and in vivo studies
have shown that as temperature decreases to 30°C, the expression
levels of Bcl-2 family proteins may increase (47). Thus, the integrity of the
mitochondrial membrane may be upheld by increasing Bcl-2 expression
levels via TH (32). Apoptosis can
be assessed by measuring Bcl-2 expression levels in H9c2
cardiomyocytes (48). Myocardial
energy metabolism is a highly controlled process. The mitochondria
forms 40% of myocardial intracellular volume, and the continuous
need for ATP is achieved via aerobic metabolism (49). Therefore, the stability of heart
function is highly dependent on normal energy metabolism in the
mitochondria. If hypoxic ischemia is succeeded by reoxygenation,
reintroduction of nutrients and oxygen contributes to
overproduction of ROS, which decreases mitochondrial membrane
potential and subsequent apoptosis (5). However, the use of TH as a treatment
for patients with severe myocardial infarction has demonstrated
beneficial effect in reducing reperfusion injury (44,50).
The present study investigated the therapeutic roles of hypothermia
in H/R, and identified that the expression levels of SUMO1 were
increased during hypothermia therapy. The present results suggested
that TH during reperfusion conferred myocardial protection, rather
than hypothermia intervention after ischemia and hypoxia.
To further investigate the role of SUMO1, the
effects of moderate hypothermia on cell apoptosis, mitochondrial
membrane potential and ROS generation were examined. Using a SUMO1
knockdown model, it was demonstrated that SUMO1 protected
cardiomyocytes against H/R injury. In addition, ROS generation was
decreased and mitochondrial membrane potential was increased during
reperfusion as a result of moderate hypothermia treatment. The
present study assessed the expression levels of SUMO1 during H/R
with moderate hypothermia treatment. It was revealed that SUMO1
knockdown blocked the binding of SUMO1 to its target proteins,
which in turn decreased the expression levels of Bcl-2. The present
results suggested that the protein expression levels of
apoptosis-related caspase-3 were significantly increased. Moreover,
siSUMO1 enhanced ROS generation and the mitochondrial membrane
potential was decreased. Therefore, the present results suggested
that SUMOylation may have a role in the cardioprotective mechanisms
of moderate hypothermia. In the present study, overexpression of
SUMO1 was sufficient to inhibit the expression of
apoptosis-associated proteins, thereby indicating better protection
in conjunction with moderate hypothermia. The present results
indicated that knockdown of SUMO1 protein expression during
moderate hypothermia treatment exacerbated H/R-induced
cardiomyocyte injury.
There are still mechanisms that require further
investigation; in particular, it is not clear whether moderate
hypothermia enhanced the protective effect of myocardial cells via
other signaling pathways. A previous study demonstrated that
hypothermia reduced the effects of ischemia/reperfusion injury on
myocardial cell apoptosis by minimizing autophagy (51). However, the effect of moderate
hypothermia on autophagy is a controversial topic, and whether
other target proteins (such as the dynamin-related protein-1) bound
to SUMO are present in the mitochondria of cardiomyocytes is not
fully understood. Due to practical constraints, in the present
study the protective mechanisms of hypothermia were studied using
in vitro models, which does not accurately simulate the
systemic responses to myocardial H/R injury. Moreover, the present
study did not conduct a detailed analysis on side effects related
to arrhythmia and changes in myocardial contractility. In addition,
the present study investigated H/R-induced mitochondrial damage but
did not examine alternative sources of myocardial dysfunction after
H/R injury, including myocardial injury and microvascular
obstruction. Götberg et al (8) demonstrated that low temperature after
ischemia did not alter the myocardial infarct area of pigs, but did
significantly reduce microvascular obstruction. Hence, further
research is required to better understand the functional
implications of the association between the specific target
proteins that conjugate with SUMO1. In vivo models need to
be established to investigate the feasibility of moderate
hypothermia treatment, while minimizing adverse reactions.
In conclusion, moderate hypothermia significantly
enhanced the ability of SUMO1 to bind to target proteins in
cardiomyocytes, which suggests an important role in antagonizing
myocardial apoptosis and preserving mitochondrial function
following hypoxic ischemia/reoxygenation. The present study
provides novel evidence on the molecular mechanisms underlying the
protective effects of moderate hypothermia. Further research will
continue to examine the theoretical mechanisms demonstrated in the
present study to facilitate the development of new drugs for
improved patient management.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81471175) and Binhai New
Area Health and Family Planning Commission Science and Technology
Project (grant nos. 2015BWKY001 and 2014BWKZ006).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL, YB and XD conceived and designed the study. JC,
XB and LL analyzed the data. JC, XB, YL, XX, CW and YY conducted
the experiments. JC wrote the manuscript. The final manuscript was
read and approved by all the authors.
Ethics approval and consent to
participate
The study protocol was approved by the Ethics
Committee of Tianjin Fifth Central Hospital.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nowbar AN, Howard JP, Finegold JA, Asaria
P and Francis DP: 2014 global geographic analysis of mortality from
ischaemic heart disease by country, age and income: Statistics from
World Health Organisation and United Nations. Int J Cardiol.
174:2617–298. 2014. View Article : Google Scholar
|
|
2
|
Joiner ML, Koval OM, Li J, He BJ,
Allamargot C, Gao Z, Luczak ED, Hall DD, Fink BD, Chen B, et al:
CaMKII determines mitochondrial stress responses in heart. Nature.
491:269–273. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Inoue T: Ischemia-reperfusion injury is
still a big hurdle to overcome for treatment of acute myocardial
infarction. J Cardiol. 67:305–306. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mandl A, Toth A and Erhardt P: Analysis of
apoptosis in isolated primary cardiac myocytes. Methods Mol Biol.
559:293–311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kuznetsov AV, Javadov S, Sickinger S,
Frotschnig S and Grimm M: H9c2 and HL-1 cells demonstrate distinct
features of energy metabolism, mitochondrial function and
sensitivity to hypoxia-reoxygenation. Biochim Biophys Acta.
1853:276–284. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Azzopardi DV, Strohm B, Edwards AD, Dyet
L, Halliday HL, Juszczak E, Kapellou O, Levene M, Marlow N, Porter
E, et al TOBY Study Group, : Moderate hypothermia to treat
perinatal asphyxial encephalopathy. N Engl J Med. 361:1349–1358.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bernard S, Buist M, Monteiro O and Smith
K: Induced hypothermia using large volume, ice-cold intravenous
fluid in comatose survivors of out-of-hospital cardiac arrest: A
preliminary report. Resuscitation. 56:9–13. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Götberg M, Olivecrona GK, Engblom H,
Ugander M, van der Pals J, Heiberg E, Arheden H and Erlinge D:
Rapid short-duration hypothermia with cold saline and endovascular
cooling before reperfusion reduces microvascular obstruction and
myocardial infarct size. BMC Cardiovasc Disord. 8:72008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tissier R, Couvreur N, Ghaleh B, Bruneval
P, Lidouren F, Morin D, Zini R, Bize A, Chenoune M, Belair MF, et
al: Rapid cooling preserves the ischaemic myocardium against
mitochondrial damage and left ventricular dysfunction. Cardiovasc
Res. 83:345–353. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Erlinge D, Götberg M, Lang I, Holzer M,
Noc M, Clemmensen P, Jensen U, Metzler B, James S, Bötker HE, et
al: Rapid endovascular catheter core cooling combined with cold
saline as an adjunct to percutaneous coronary intervention for the
treatment of acute myocardial infarction. The CHILL-MI trial: A
randomized controlled study of the use of central venous catheter
core cooling combined with cold saline as an adjunct to
percutaneous coronary intervention for the treatment of acute
myocardial infarction. J Am Coll Cardiol. 63:1857–1865. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Götberg M, Olivecrona GK, Koul S, Carlsson
M, Engblom H, Ugander M, van der Pals J, Algotsson L, Arheden H and
Erlinge D: A pilot study of rapid cooling by cold saline and
endovascular cooling before reperfusion in patients with
ST-elevation myocardial infarction. Circ Cardiovasc Interv.
3:400–407. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhong N, Kim CY, Rizzu P, Geula C, Porter
DR, Pothos EN, Squitieri F, Heutink P and Xu J: DJ-1
transcriptionally up-regulates the human tyrosine hydroxylase by
inhibiting the sumoylation of pyrimidine tract-binding
protein-associated splicing factor. J Biol Chem. 281:20940–20948.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gareau JR and Lima CD: The SUMO pathway:
Emerging mechanisms that shape specificity, conjugation and
recognition. Nat Rev Mol Cell Biol. 11:861–871. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Prudden J, Perry JJP, Arvai AS, Tainer JA
and Boddy MN: Molecular mimicry of SUMO promotes DNA repair. Nat
Struct Mol Biol. 16:509–516. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Z and Prelich G: Quality control of a
transcriptional regulator by SUMO-targeted degradation. Mol Cell
Biol. 29:1694–1706. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Flotho A and Melchior F: Sumoylation: A
regulatory protein modification in health and disease. Annu Rev
Biochem. 82:357–385. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim EY, Zhang Y, Beketaev I, Segura AM, Yu
W, Xi Y, Chang J and Wang J: SENP5, a SUMO isopeptidase, induces
apoptosis and cardiomyopathy. J Mol Cell Cardiol. 78:154–164. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim EY, Zhang Y, Ye B, Segura AM, Beketaev
I, Xi Y, Yu W, Chang J, Li F and Wang J: Involvement of activated
SUMO-2 conjugation in cardiomyopathy. Biochim Biophys Acta.
1852:1388–1399. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cai R, Gu J, Sun H, Liu X, Mei W, Qi Y,
Xue S, Ren S, Rabinowitz JE, Wang Y, et al: Induction of SENP1 in
myocardium contributes to abnormities of mitochondria and
cardiomyopathy. J Mol Cell Cardiol. 79:115–122. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim EY, Chen L, Ma Y, Yu W, Chang J,
Moskowitz IP and Wang J: Enhanced desumoylation in murine hearts by
overexpressed SENP2 leads to congenital heart defects and cardiac
dysfunction. J Mol Cell Cardiol. 52:638–649. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Montgomery RL, Davis CA, Potthoff MJ,
Haberland M, Fielitz J, Qi X, Hill JA, Richardson JA and Olson EN:
Histone deacetylases 1 and 2 redundantly regulate cardiac
morphogenesis, growth, and contractility. Genes Dev. 21:1790–1802.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Joung H, Kwon S, Kim KH, Lee YG, Shin S,
Kwon DH, Lee YU, Kook T, Choe N, Kim JC, et al: Sumoylation of
histone deacetylase 1 regulates MyoD signaling during myogenesis.
Exp Mol Med. 50:e4272018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lee YJ, Mou Y, Klimanis D, Bernstock JD
and Hallenbeck JM: Global SUMOylation is a molecular mechanism
underlying hypothermia-induced ischemic tolerance. Front Cell
Neurosci. 8:4162014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu X, Ren W, Jiang Z, Su Z, Ma X, Li Y,
Jiang R, Zhang J and Yang X: Hypothermia inhibits the proliferation
of bone marrow-derived mesenchymal stem cells and increases
tolerance to hypoxia by enhancing SUMOylation. Int J Mol Med.
40:1631–1638. 2017.PubMed/NCBI
|
|
25
|
Wei Y and Qing M: G Burkhard M and Wulf P:
Deep hypothermia markedly activates the small ubiquitin-like
modifier conjugation pathway; implications for the fate of cells
exposed to transient deep hypothermic cardiopulmonary bypass. J
Cereb Blood Flow Metab (Nihongoban). 29:886–890. 2009. View Article : Google Scholar
|
|
26
|
Nolan JP, Soar J, Cariou A, Cronberg T,
Moulaert VR, Deakin CD, Bottiger BW, Friberg H, Sunde K and
Sandroni C: European Resuscitation Council and European Society of
Intensive Care Medicine Guidelines for Post-resuscitation Care
2015: Section 5 of the European Resuscitation Council Resuscitation
Guidelines 2015. Resuscitation. 81:1389–1399. 2015.
|
|
27
|
Lesley C, Yi D, Rita K, Karin P and
Sanderson TH: Mitochondrial dynamics: An emerging paradigm in
ischemia-reperfusion injury. Curr Pharm Des. 19:6848–6857. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Requião-Moura LR, Durão Junior MS, Matos
AC and Pacheco-Silva A: Ischemia and reperfusion injury in renal
transplantation: Hemodynamic and immunological paradigms. Einstein
(Sao Paulo). 13:129–135. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang CH, Chiang CY, Pen RH, Tsai MS, Chen
HW, Hsu CY, Wang TD, Ma MH, Chen SC and Chen WJ: Hypothermia
treatment preserves mitochondrial integrity and viability of
cardiomyocytes after ischaemic reperfusion injury. Injury.
46:233–239. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Prudent J, Zunino R, Sugiura A, Mattie S,
Shore GC and McBride HM: MAPL SUMOylation of Drp1 stabilizes an
ER/mitochondrial platform required for cell death. Mol Cell.
59:941–955. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Drescher C, Diestel A, Wollersheim S,
Berger F and Schmitt KR: How does hypothermia protect
cardiomyocytes during cardioplegic ischemia? Eur J Cardiothorac
Surg. 40:352–359. 2011.PubMed/NCBI
|
|
32
|
Krech J, Tong G, Wowro S, Walker C,
Rosenthal LM, Berger F and Schmitt KR: Moderate therapeutic
hypothermia induces multimodal protective effects in oxygen-glucose
deprivation/reperfusion injured cardiomyocytes. Mitochondrion.
35:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ashraf H, Pradhan L, Chang EI, Terada R,
Ryan NJ, Briggs LE, Chowdhury R, Zárate MA, Sugi Y, Nam HJ, et al:
A mouse model of human congenital heart disease: High incidence of
diverse cardiac anomalies and ventricular noncompaction produced by
heterozygous Nkx2-5 homeodomain missense mutation. Circ Cardiovasc
Genet. 7:423–433. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Iribarren PA, Berazategui MA, Bayona JC,
Almeida IC, Cazzulo JJ and Alvarez VE: Different proteomic
strategies to identify genuine Small Ubiquitin-like MOdifier
targets and their modification sites in Trypanosoma brucei
procyclic forms. Cell Microbiol. 17:1413–1422. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jin LZ, Lu JS and Gao JW: Silencing SUMO2
promotes protection against degradation and apoptosis of nucleus
pulposus cells through p53 signaling pathway in intervertebral disc
degeneration. Biosci Rep. 38:BSR201715232018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Hay RT: SUMO-specific proteases: A twist
in the tail. Trends Cell Biol. 17:370–376. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang W, Sheng H, Warner DS and Paschen W:
Transient global cerebral ischemia induces a massive increase in
protein sumoylation. J Cereb Blood Flow Metab. 28:269–279. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Datwyler AL, Lättig-Tünnemann G, Yang W,
Paschen W, Lee SL, Dirnagl U, Endres M and Harms C: SUMO2/3
conjugation is an endogenous neuroprotective mechanism. J Cereb
Blood Flow Metab. 31:2152–2159. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li G, Liu X, Su Z and Zhang D: Hypothermia
exerts early neuroprotective effects involving protein conjugation
of SUMO 2/3 in a rat model of middle cerebral artery occlusion. Mol
Med Rep. 16:3217–3223. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shimizu Y, Lambert JP, Nicholson CK, Kim
JJ, Wolfson DW, Cho HC, Husain A, Naqvi N, Chin LS, Li L, et al:
DJ-1 protects the heart against ischemia-reperfusion injury by
regulating mitochondrial fission. J Mol Cell Cardiol. 97:56–66.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee A, Jeong D, Mitsuyama S, Oh JG, Liang
L, Ikeda Y, Sadoshima J, Hajjar RJ and Kho C: The role of SUMO-1 in
cardiac oxidative stress and hypertrophy. Antioxid Redox Signal.
21:1986–2001. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shao R, Zhang F-P, Tian F, Friberg PA,
Wang X, Sjöland H and Billig H: Increase of SUMO-1 expression in
response to hypoxia: Direct interaction with HIF-1α in adult mouse
brain and heart in vivo. FEBS Lett. 569:293–300. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang L, Ma Q, Yang W, Mackensen GB and
Paschen W: Moderate hypothermia induces marked increase in levels
and nuclear accumulation of SUMO2/3-conjugated proteins in neurons.
J Neurochem. 123:349–359. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Blatt A, Elbaz-Greener GA, Mizrachi A,
J'bara Z, Taraboulos T, Litovchik I, Vered Z and Minha S:
Adjunctive mild hypothermia therapy to primary percutaneous
coronary intervention in patients with ST segment elevation
myocardial infarction complicated with cardiogenic shock: A pilot
feasibility study. Cardiol J. 22:285–289. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tong G, Walker C, Bührer C, Berger F,
Miera O and Schmitt KR: Moderate hypothermia initiated during
oxygen-glucose deprivation preserves HL-1 cardiomyocytes.
Cryobiology. 70:101–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yang D, Guo S, Zhang T and Li H:
Hypothermia attenuates ischemia/reperfusion-induced endothelial
cell apoptosis via alterations in apoptotic pathways and JNK
signaling. FEBS Lett. 583:2500–2506. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ning XH, Chen SH, Xu CS, Li L, Yao LY,
Qian K, Krueger JJ, Hyyti OM and Portman MA: Hypothermic protection
of the ischemic heart via alterations in apoptotic pathways as
assessed by gene array analysis. J Appl Physiol (1985).
92:2200–2207. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang C, Liu X, Miao J, Wang S, Wu L, Yan
D, Li J, Guo W, Wu X and Shen A: Heat shock protein 70 protects
cardiomyocytes through suppressing SUMOylation and nucleus
translocation of phosphorylated eukaryotic elongation factor 2
during myocardial ischemia and reperfusion. Apoptosis. 22:608–625.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Vogel S, Rath D, Lu J, Chatterjee M,
Geisler T and Gawaz M: Elevated mitochondrial membrane potential of
circulating monocyte-platelet aggregates in patients with coronary
heart disease. Int J Cardiol. 181:135–137. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mohammad MA, Noc M, Lang I, Holzer M,
Clemmensen P, Jensen U, Metzler B and Erlinge D: Proteomics in
hypothermia as adjunctive therapy in patients with ST-segment
elevation myocardial infarction: A CHILL-MI substudy. Ther
Hypothermia Temp Manag. 7:152–161. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cheng BC, Huang HS, Chao CM, Hsu CC, Chen
CY and Chang CP: Hypothermia may attenuate
ischemia/reperfusion-induced cardiomyocyte death by reducing
autophagy. Int J Cardiol. 168:2064–2069. 2013. View Article : Google Scholar : PubMed/NCBI
|