Introduction
Vitiligo is an acquired depigmenting disorder
affecting 0.5% of the world population, which demonstrates no sex
or ethnic differences (1). The
most significant progress in the understanding of the disease
etiology has been made in three research areas: Characterization of
the stress responses activated by triggers of vitiligo, delineating
the autoimmune components that promote the progression of the
disease and identifying susceptibility genes (2). However, there are currently no
existing treatments for vitiligo that can effectively promote the
complete re-pigmentation with long-lasting effects, while
preventing recurrence (1).
Nevertheless, narrowband-UVB therapy, and the combined treatment
with systemic therapies (steroids and immunosuppressants) and
topical therapies (corticosteroids and calcineurin inhibitors) are
commonly used to treat vitiligo (1).
Interferons (IFNs), which are widely used for
antiviral and antitumor therapies, are multifunctional proteins
that initiate and regulate various cellular responses, including
antiviral and antiproliferative activity, controlling cell
apoptosis and immune regulation (3–6).
Previously, IFN-γ was discovered to inhibit T lymphocyte
aggregation, which further impeded pigment loss in vitiligo model
transgenic mice (7). Another study
demonstrated that IFN-γ recruited T lymphocytes via the C-X-C motif
ligand (CXCL)10/C-X-C chemokine receptor 3 signaling axis (8). In addition, it has been reported that
the IFN-γ-induced apoptosis of melanocytes induced vitiligo
(9). It was also observed that
increased expression levels of IFN-γ served an important role in
vitiligo-induced depigmentation via the direct induction of
melanocyte apoptosis (9). Vitiligo
is a disease that affects the appearance and leads to low
self-esteem and social stigma (10). It is characterized by localized or
generalized patches of skin depigmentation; therefore, either the
reduction or absence of melanocytes in the local epidermis may be a
prominent cause for the formation of vitiligo leukopluriae
(11–13).
In the canonical IFN-γ signaling pathway, janus
kinase 1 (JAK1) and STAT1 are the major signaling molecules that
regulate the activation of downstream IFN-γ-inducible genes
(14,15). For example, a previous study
reported that IFN-γ was able to inhibit STAT1 signaling, which
induced apoptosis and promoted pancreatic β-cell survival (16). Significant research has been
conducted into the pathogenic role of increased rates of apoptosis
in melanocytes during vitiligo to further understand the
theoretical and experimental basis for the prevention and treatment
of the disease (9); however, to
the best of our knowledge, little is known in terms of its
molecular mechanism and whether JAK/STAT signaling is involved in
the process.
The present study aimed to demonstrate the effect of
IFN-γ on the apoptosis of human melanocytes and to investigate the
underlying mechanism. The results suggested that IFN-γ induction
may activate the JAK1/STAT1 signaling pathway, differentially
regulate the expression levels of Bcl-2, Bax, Bcl-2 homologous
antagonist killer (Bak) and cleaved caspase-3, and consequently
promote the apoptosis of human melanocytes.
Materials and methods
Cell culture
Human epidermal melanocytes (HEMs) from moderately
pigmented skin were purchased from ScienCell Research Laboratories,
Inc. HEMs were cultured at 37°C in a humidified incubator
containing 5% CO2, in Medium 254 (Sigma-Aldrich; Merck
KGaA), supplemented with 1% human melanocyte growth supplement
(ScienCell Research Laboratories, Inc.). HEMs were treated with
100, 200 or 400 ng/ml IFN-γ at 37°C (PeproTech, Inc.).
Cell Counting Kit-8 (CCK-8) assay
A total of 2×103 HEMs/well were plated in
triplicate into 96-well plates in 100 µl growth medium. Following
24, 48 and 72 h of treatment with IFN-γ, the cultured primary HEMs
were collected and the viability was assessed using a CCK-8 assay
(MedChemExpress), according to the manufacturer's protocol.
Subsequently, the absorbance of each well was measured at 450 nm
using a Dynatech MR5000 plate reader (Dynatech Laboratories).
Flow cytometric analysis of
apoptosis
A total of 1×106 HEMs/well were collected
and plated in triplicate into six-well plates in 2 ml growth
medium. Morphological changes were observed using an inverted
microscope (magnification, ×100) before (0 h) and after the
treatment with IFN-γ (48 h) at 37°C. The cells were subsequently
stained using the Annexin V/FITC Apoptosis Detection kit
(eBioscience; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol, 48 h after the treatment with IFN-γ at
37°C. Apoptotic cells were analyzed using a flow cytometer (BD
Accuri C6; BD Biosciences) and analyzed using BD CellQuest™ version
5.1 software (BD Biosciences).
Western blotting
HEMs treated with IFN-γ were collected by
centrifugation (300 × g; 10 min; room temperature) and total
protein was extracted using RIPA lysis buffer (10 mM NaPO4, pH 7.4,
300 mM NaCl, 0.1% SDS, 1% Nonidet P-40, 1% deoxycholic acid and 2
mM EDTA), supplemented with protease inhibitors (Pierce; Thermo
Fisher Scientific, Inc.). Total protein was quantified using a
bicinchoninic acid assay kit (cat. no. BL521A; Biosharp Life
Sciences) and 25 µg protein/lane was separated by 8–10% SDS-PAGE.
The separated proteins were subsequently transferred onto a
polyvinylidene difluoride membrane (EMD Millipore) and blocked with
5% BSA (Beyotime Institute of Biotechnology) for 1 h at room
temperature. The membrane was incubated overnight at 4°C with the
following specific primary antibodies: Anti-Bcl-2 (1:1,000; cat.
no. BS1511; Bioworld Technology, Inc.), anti-Bax (1:1,000; cat. no.
BS2538; Bioworld Technology, Inc.), anti-Bak (1:5,000; cat. no.
ab32371; Abcam), anti-cleaved caspase-3 (1:500; cat. no. ab32042;
Abcam), anti-phosphorylated (p)-JAK1 (1:2,000; cat. no. ab138005;
Abcam), anti-p-JAK2 (1:5,000; cat. no. ab32101; Abcam),
anti-p-STAT1 (1:5,000; cat. no. ab109461; Abcam), anti-GAPDH
(1:5,000; cat. no. AB-P-R001; Goodhere), anti-JAK2 (1:3,000; cat.
no. ab108596; Abcam), anti-JAK1 (1:5,000; cat. no. ab133666; Abcam)
and anti-STAT1 (1:1,000; cat. no. ab92506; Abcam). Following the
primary antibody incubation, the membranes were incubated with a
horseradish peroxidase-conjugated anti-rabbit secondary antibody
(1:3,000; cat. no. sc-2004; Santa Cruz Biotechnology, Inc.) at room
temperature in the dark for 1 h. Protein bands were visualized
using an ECL reagent (Beyotime Institute of Biotechnology) and
expression levels were semi-quantified using ImageJ version 1.51k
software (National Institutes of Health).
Statistical analysis
Statistical analysis was performed using SPSS
version 13.0 software (SPSS, Inc.) and data are presented as the
mean ± SD of three independent experiments. Statistical differences
among multiple groups were determined using a one-way ANOVA,
followed by Bonferroni's post hoc test for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
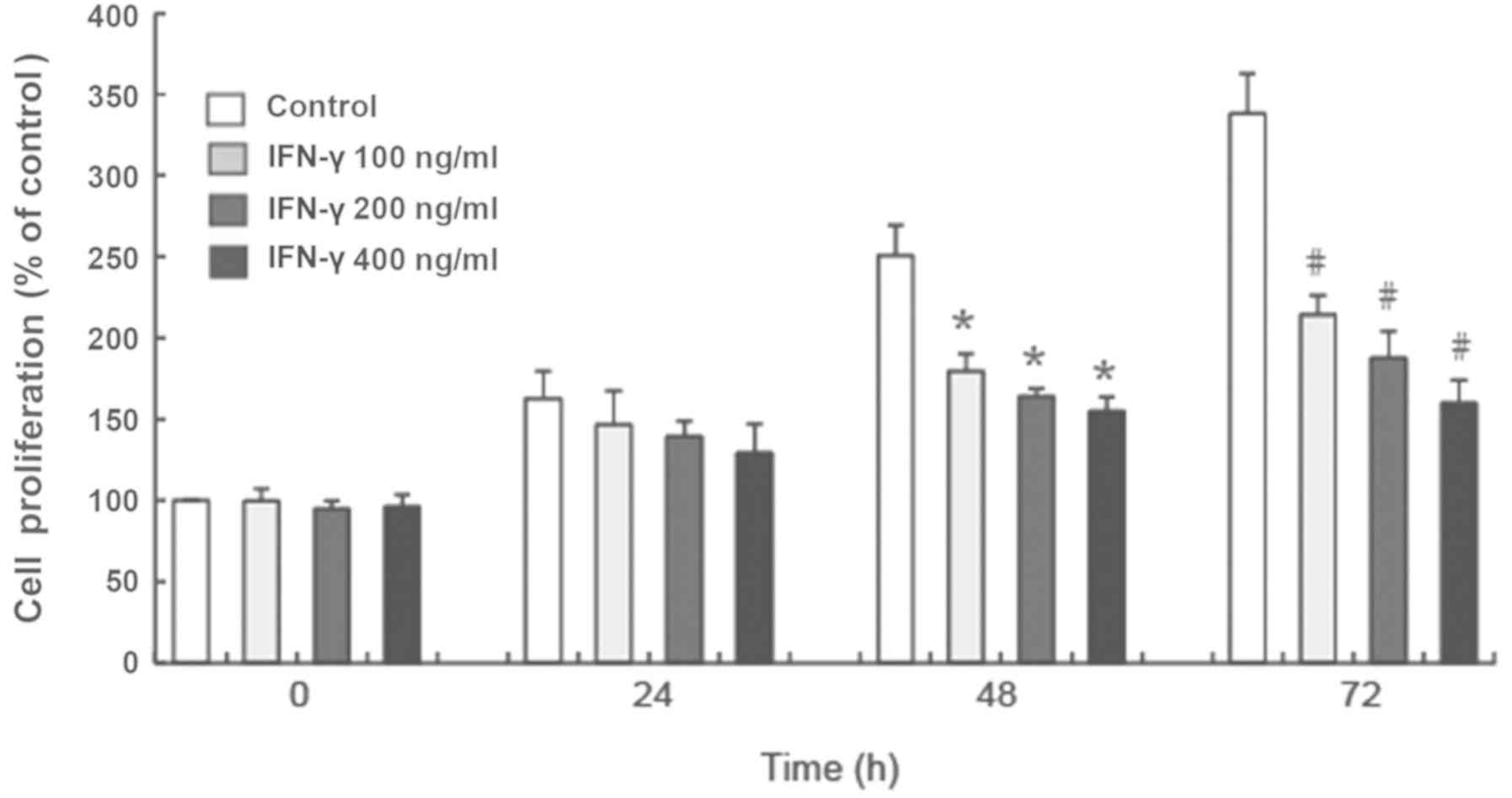
IFN-γ inhibits the proliferation of
HEMs
The results of the CCK-8 assay demonstrated that
100, 200 and 400 ng/ml IFN-γ significantly inhibited the
proliferation of human melanocytes in vitro at 48 and 72 h
compared with the control group (P<0.05; Fig. 1). Additionally, the inhibited
proliferation of HEMs treated with different concentrations of
IFN-γ occurred in a time-dependent manner (Fig. 1).
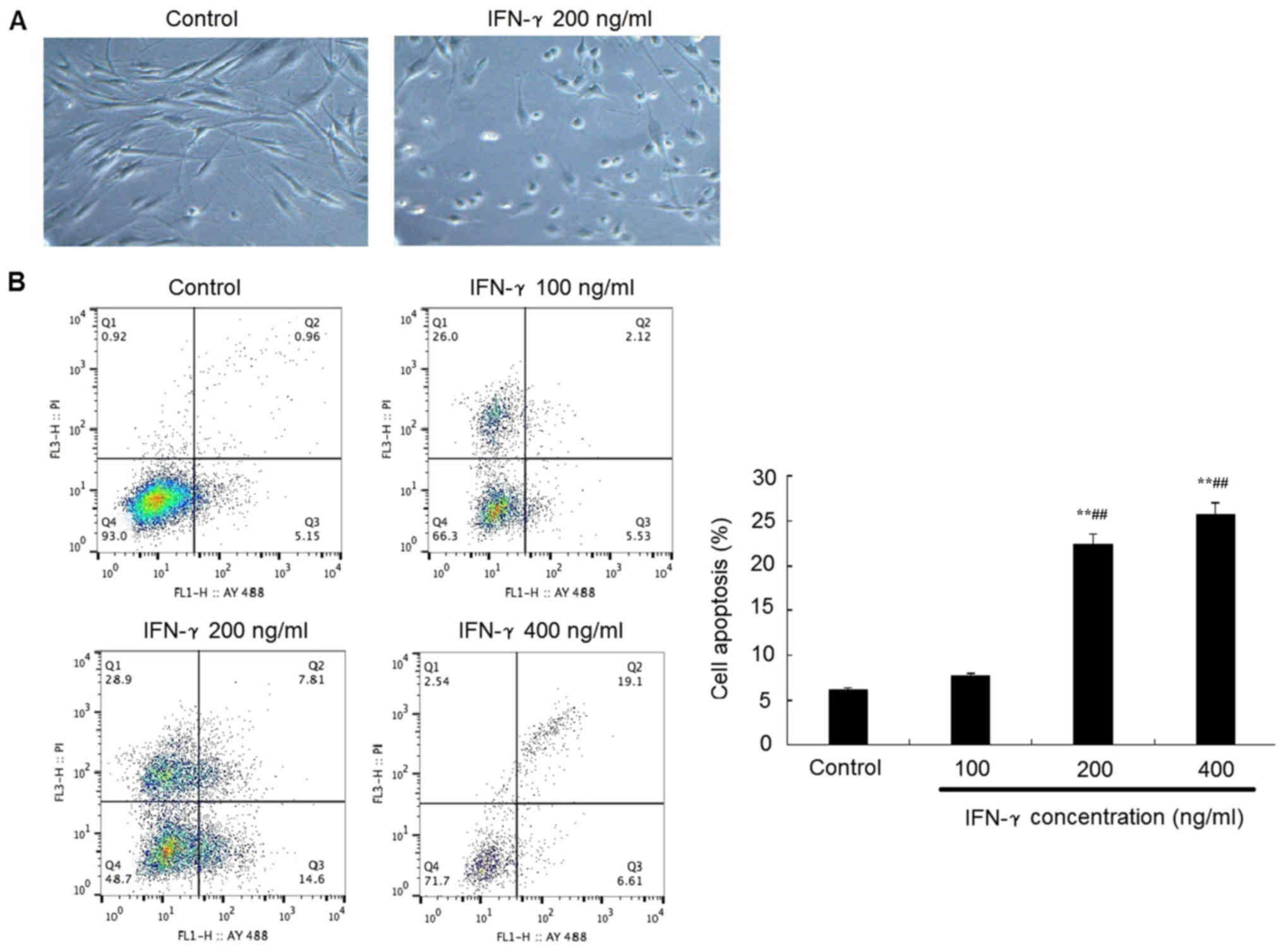
IFN-γ promotes the apoptosis of
HEMs
Representative morphological images revealed that
the treatment with 200 ng/ml IFN-γ for 48 h promoted the HEMs to
conform to a rounder morphology, with an increased number of cells
in the suspension compared with the control group (Fig. 2A). In addition, flow cytometric
analysis of apoptosis revealed that following the treatment with
200 or 400 ng/ml IFN-γ treatment for 48 h, the apoptotic rate of
HEMs was significantly increased compared with the control group
(P<0.05; Fig 2B).
IFN-γ activates the JAK1/STAT1
signaling pathway
It is well established that upon IFN-γ binding to
its receptor, the JAK/STAT signaling axis is activated by
phosphorylation (17). Western
blotting revealed that the treatment with IFN-γ at various
concentrations significantly increased the phosphorylation levels
of JAK1 and STAT1 (Fig. 3A and B);
however, although 100 and 400 ng/ml IFN-γ demonstrated these
effects on the phosphorylation status of JAK2, the 200 ng/ml group
demonstrated no significant different in the phosphorylation status
compared with the control group. Moreover, the difference between
100 and 400 ng/ml IFN-γ treatment was not statistically different.
Meanwhile, the expression levels of the anti-apoptotic protein
Bcl-2 were significantly reduced, whereas the expression levels of
the proapoptotic proteins Bax, Bak and cleaved caspase-3 were
significantly increased in the 100, 200 and 400 ng/ml IFN-γ
treatment groups compared with the control group (P<0.05;
Fig. 3A and B).
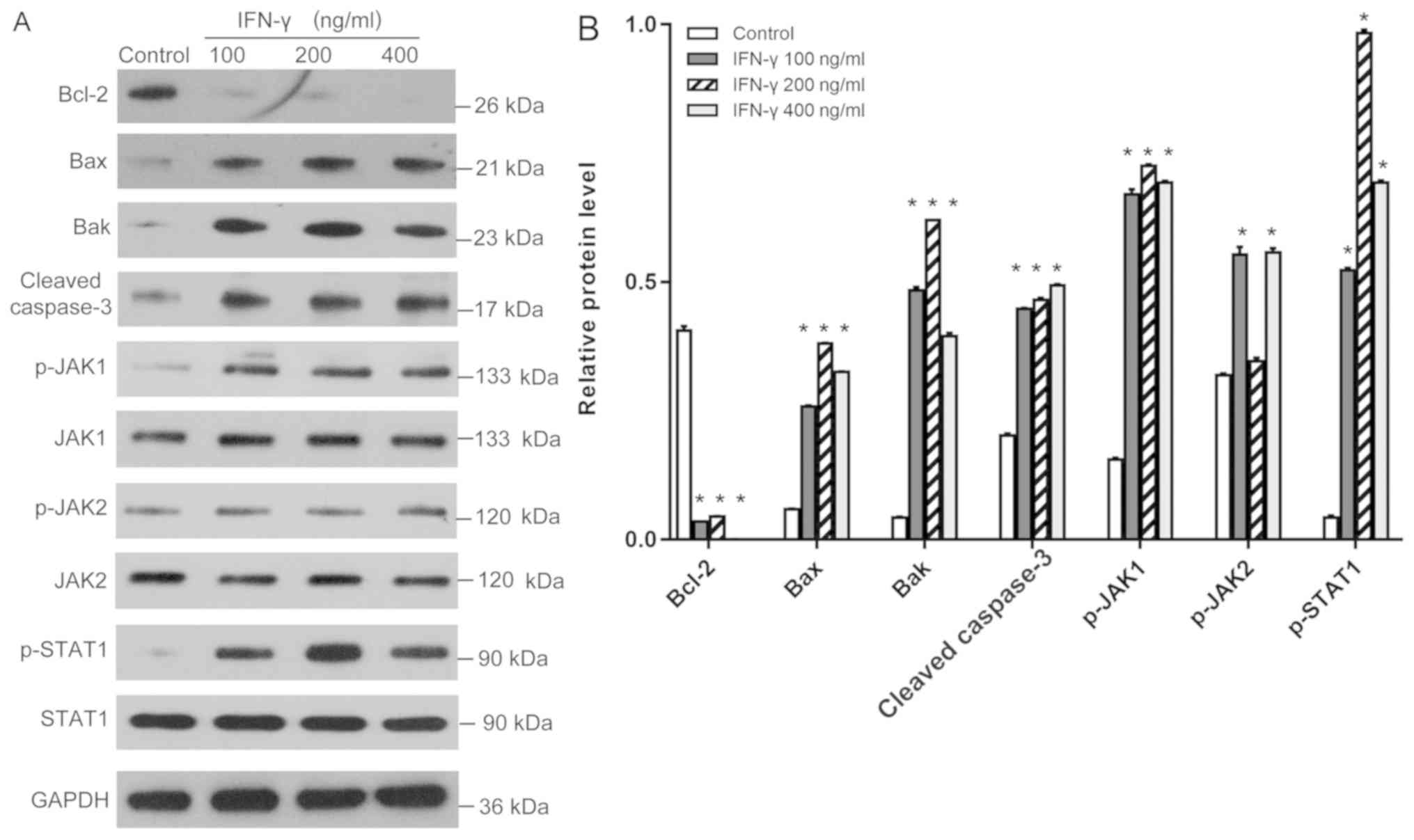 | Figure 3.Effects of IFN-γ on the expression
levels of Bcl-2, Bax, Bak, cleaved caspase-3, p-JAK1, p-JAK2 and
p-STAT1. (A) Western blotting was used to analyze the expression
levels of the proteins in human epidermal melanocytes following the
treatment with 100, 200 or 400 ng/ml IFN-γ. (B) Quantification of
the western blotting bands in part A. Bcl-2, Bax, Bak, cleaved
caspase-3 were normalized to GAPDH. p-JAK1, p-JAK2 and p-STAT1 were
normalized to JAK-1, JAK-2 or STAT1, respectively. *P<0.05 vs.
control. p-, phosphorylated; IFN, interferon; JAK, janus kinase;
Bak, Bcl-2 homologous antagonist killer. |
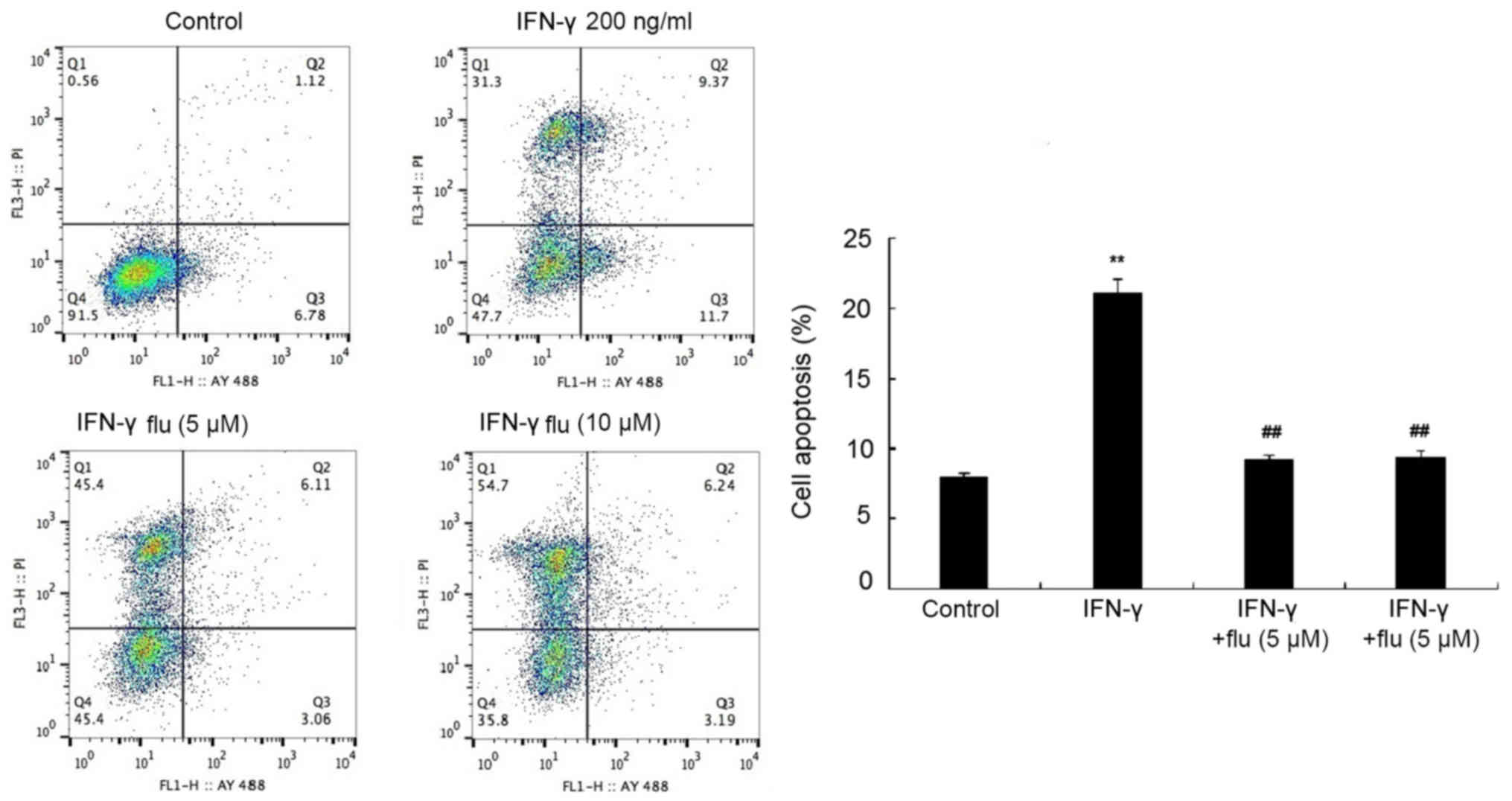
STAT1 inhibitor attenuates the
proapoptotic effect of IFN-γ on melanocytes
HEMs were pretreated with 5 or 10 µM fludarabine
(STAT1 inhibitor) for 1 h and then treated with 200 ng/ml IFN-γ for
48 h. The apoptotic rate was analyzed by flow cytometry following
Annexin V-FITC/PI double staining and it was revealed that
fludarabine treatment significantly reduced the proapoptotic
effects of IFN-γ on HEMs (P<0.01; Fig. 4).
Discussion
IFN-γ, a type III interferon, is a soluble
glycoprotein with antiviral, antitumor, anti-parasite and
immunoregulatory activities (2).
It has been hypothesized that the IFN-γ present in the skin lesions
during vitiligo is secreted by CD8+ cytotoxic T
lymphocytes, which can induce the apoptosis of melanocytes
(18,19); however, the mechanism of
IFN-γ-induced melanocyte apoptosis remains unclear. In the present
study, primary HEMs were used to investigate the toxicity of IFN-γ.
The results demonstrated that 200 and 400 ng/ml IFN-γ inhibited the
proliferation of HEMs in a time-dependent manner. In addition, an
increased apoptotic rate of HEMs was also observed following the
treatment with 200 ng/ml IFN-γ for 48 h. These results indicated
that the cytotoxic effect of IFN-γ on HEMs may be predominantly
achieved by inducing apoptosis. Notably, these results were
consistent with previous findings (9,14).
Further evidence has revealed that by binding with specific
receptors, IFN-γ induced cell apoptosis as well as promoting cell
proliferation. The effects of IFN-γ on cell proliferation and
apoptosis have been discovered to depend on the differential
expression levels of IFN-γ receptors (IFNGRs), with increased
expression levels of IFNGR2 more likely to induce apoptosis
(2); however, further
investigations are required to determine whether IFNGR2 interferes
with IFN-γ-induced melanocyte apoptosis.
The JAK/STAT signaling pathway, which widely exists
in vivo and participates in multiple cellular processes, is
known to serve an important role in signal transduction and the
transcriptional activation of STAT1 (20–22).
The present study results demonstrated an decrease in the
proliferative rate in IFN-γ-treated groups following 48 and 72 h,
with the difference being even more pronounced following 72 h
compared with the initial treatment. However, neither the treatment
for 0 nor 24 h resulted in a decrease in the proliferative rate
between the IFN-γ groups and controls. These findings may be
explained by the increased expression levels of p-JAK1 and p-STAT1
observed following 48 h of treatment. A previous study reported
that the JAK/STAT signaling pathway served both antiproliferative
and proapoptotic effects (23).
Furthermore, the expression levels of p-JAK1and p-STAT1 were the
highest in the 200 ng/ml IFN-γ group compared with the 100 and 400
ng/ml IFN-γ treatment groups. These findings provided evidence to
suggest that the activation of the JAK1/STAT1 signaling pathway may
contribute to the apoptosis of IFN-γ-induced HEMs.
Furthermore, the activation of STAT1 was previously
found to be involved in the regulation of apoptosis by regulating
the downstream Bcl-2 family members, Bcl-2 and Bax (24). The ratio between the anti-apoptotic
protein Bcl-2 and the proapoptotic protein Bax has been revealed to
be inversely correlated with the rate of apoptosis (25,26).
Caspase-3 is an apoptosis-executing protein in the caspase family
and functional cleaved caspase-3 is known to promote apoptosis
together with Bcl-2 (27,28). The present study results discovered
that melanocytes induced with IFN-γ for 48 h exhibited reduced
expression levels of Bcl-2 protein and increased expression levels
of Bax, Bak, cleaved caspase-3, p-JAK1 and p-STAT1; however, no
significant difference was observed in the expression levels of
p-JAK2 following 200 ng/ml IFN-γ treatment compared with the
control. To understand whether the apoptosis was associated with
the activation of the JAK/STAT1 signaling pathway in IFN-γ induced
human melanocytes, the specific STAT1 inhibitor fludarabine was
used in the present study; a significantly reduced apoptotic rate
was discovered in HEMs co-treated with the inhibitor and IFN-γ
compared with IFN-γ alone, suggesting that IFN-γ-induced human
melanocyte apoptosis may be associated with the activation of the
JAK/STAT1 signaling pathway, and STAT1 may regulate the expression
levels of Bax, Bak and cleaved caspase-3.
IFN-γ was previously discovered to be critical for
the progression of vitiligo by recruiting autoreactive
CD8+ T lymphocytes to the skin through the induction of
CXCL10 (28,29). The present study identified that
IFN-γ could also induce the apoptosis of human melanocytes through
activating the JAK1/STAT1 signaling pathway, therefore potentially
serving as an initiating factor that destroys melanocytes. Future
studies using animal models will be helpful in determining how
important IFN-γ-induced apoptosis is compared with other mechanisms
of cytotoxicity during the progression of vitiligo in vivo
(29,30). These findings may have therapeutic
implications, as inhibiting IFN-γ or its downstream signaling may
provide a novel, effective and long-lasting therapy for
vitiligo.
In conclusion, the findings of the present study
suggested that IFN-γ may induce the apoptosis of human melanocytes.
These proapoptotic effects of IFN-γ on HEMs may be mediated through
the activation of the JAK1/STAT1 signaling pathway, increasing the
expression levels of Bax, Bak and cleaved caspase-3, and decreasing
the expression levels of Bcl-2. Overall, the intervention of the
JAK1/STAT1 signaling pathway may be an effective method to reduce
the IFN-γ-induced apoptosis of HEMs, which is important for the
prevention and treatment of vitiligo.
Acknowledgements
Not applicable.
Funding
The present study was supported by the General
Subject of Medical Science and Technology Development of Nanjing
Municipal Health and Family Planning Commission (grant no.
YKK17275) and the Fundamental Research Funds for the Central
Universities (grant no. 2242019K49253).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
QS and FW conceived and designed the study; QS and
ZD conducted the experiments; MC and RC analyzed the data; QS, ZD,
MC and RC drafted the manuscript; and FW revised the manuscript.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Taieb A, Alomar A, Böhm M, Dell'anna ML,
De Pase A, Eleftheriadou V, Ezzedine K, Gauthier Y, Gawkrodger DJ,
Jouary T, et al Vitiligo European Task Force (VETF); European
Academy of Dermatology: Venereology (EADV); Union Europe´enne des
Me´decins Spe´cialistes (UEMS), : Guidelines for the management of
vitiligo: The European Dermatology Forum consensus. Br J Dermatol.
168:3111–19. 2013. View Article : Google Scholar
|
|
2
|
Manga P, Elbuluk N and Orlow SJ: Recent
advances in understanding vitiligo. F1000 Res. 5:52016. View Article : Google Scholar
|
|
3
|
Arya V, Bansal M, Girard L, Arya S and
Valluri A: Vitiligo at Injection Site of PEG-IFN-α 2a in Two
Patients with Chronic Hepatitis C: Case Report and Literature
Review. Case Rep Dermatol. 2:156–164. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hamadah I, Binamer Y, Sanai FM, Abdo AA
and Alajlan A: Interferon-induced vitiligo in hepatitis C patients:
A case series. Int J Dermatol. 49:829–833. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rozera C, Cappellini GA, D'Agostino G,
Santodonato L, Castiello L, Urbani F, Macchia I, Aricò E, Casorelli
I, Sestili P, et al: Intratumoral injection of IFN-alpha dendritic
cells after dacarbazine activates anti-tumor immunity: Results from
a phase I trial in advanced melanoma. J Transl Med. 13:1392015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang S, Liu D, Ning W and Xu A: Cytosolic
dsDNA triggers apoptosis and pro-inflammatory cytokine production
in normal human melanocytes. Exp Dermatol. 24:298–300. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Harris JE, Harris TH, Weninger W, Wherry
EJ, Hunter CA and Turka LA: A mouse model of vitiligo with focused
epidermal depigmentation requires IFN-γ for autoreactive
CD8+ T-cell accumulation in the skin. J Invest Dermatol.
132:1869–1876. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rashighi M and Harris JE: Interfering with
the IFN-γ/CXCL10 pathway to develop new targeted treatments for
vitiligo. Ann Transl Med. 3:3432015.PubMed/NCBI
|
|
9
|
Yang L, Wei Y, Sun Y, Shi W, Yang J, Zhu L
and Li M: Interferon-gamma Inhibits Melanogenesis and Induces
Apoptosis in Melanocytes: A Pivotal Role of CD8+
Cytotoxic T lymphocytes in Vitiligo. Acta Derm Venereol.
95:664–670. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee JH, Kwon HS, Jung HM, Lee H, Kim GM,
Yim HW and Bae JM: Treatment Outcomes of Topical Calcineurin
Inhibitor Therapy for Patients With Vitiligo: A Systematic Review
and Meta-analysis. JAMA Dermatol. 155:9292019. View Article : Google Scholar
|
|
11
|
Nahhas AF, Mohammad TF and Hamzavi IH:
Vitiligo Surgery: Shuffling Melanocytes. J Investig Dermatol Symp
Proc. 18:S34–S37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ma J, Li S, Zhu L, Guo S, Yi X, Cui T, He
Y, Chang Y, Liu B, Li C, et al: Baicalein protects human vitiligo
melanocytes from oxidative stress through activation of
NF-E2-related factor2 (Nrf2) signaling pathway. Free Radic Biol
Med. 129:492–503. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang K, Xiong X, Pallavi G, Ling Y, Ding
F, Duan W, Sun W, Ding G, Gong Q, Zhu W, et al: The early
repigmentation pattern of vitiligo is related to the source of
melanocytes and by the choice of therapy: A retrospective cohort
study. Int J Dermatol. 57:324–331. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jia H, Song L, Cong Q, Wang J, Xu H, Chu
Y, Li Q, Zhang Y, Zou X, Zhang C, et al: The LIM protein AJUBA
promotes colorectal cancer cell survival through suppression of
JAK1/STAT1/IFIT2 network. Oncogene. 36:2655–2666. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chou DH, Vetere A, Choudhary A, Scully SS,
Schenone M, Tang A, Gomez R, Burns SM, Lundh M, Vital T, et al:
Kinase-Independent Small-Molecule Inhibition of JAK-STAT Signaling.
J Am Chem Soc. 137:7929–7934. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kaplan DH, Greenlund AC, Tanner JW, Shaw
AS and Schreiber RD: Identification of an interferon-gamma receptor
alpha chain sequence required for JAK-1 binding. J Biol Chem.
271:9–12. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schroder K, Hertzog PJ, Ravasi T and Hume
DA: Interferon-gamma: An overview of signals, mechanisms and
functions. J Leukoc Biol. 75:163–189. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Strassner JP and Harris JE: Understanding
mechanisms of autoimmunity through translational research in
vitiligo. Curr Opin Immunol. 43:81–88. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Richmond JM, Bangari DS, Essien KI,
Currimbhoy SD, Groom JR, Pandya AG, Youd ME, Luster AD and Harris
JE: Keratinocyte-Derived Chemokines Orchestrate T-Cell Positioning
in the Epidermis during Vitiligo and May Serve as Biomarkers of
Disease. J Invest Dermatol. 137:350–358. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lu B, Antoine DJ, Kwan K, Lundbäck P,
Wähämaa H, Schierbeck H, Robinson M, Van Zoelen MA, Yang H, Li J,
et al: JAK/STAT1 signaling promotes HMGB1 hyperacetylation and
nuclear translocation. Proc Natl Acad Sci USA. 111:3068–3073. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ahn J, Lee J and Kim S: Interferon-gamma
inhibits the neuronal differentiation of neural progenitor cells by
inhibiting the expression of Neurogenin2 via the JAK/STAT1 pathway.
Biochem Biophys Res Commun. 466:52–59. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang S, Zheng G, Zhao L, Xu F and Qian J:
Shp-2 contributes to anti-RSV activity in human pulmonary alveolar
epithelial cells by interfering with the IFN-α-induced Jak/Stat1
pathway. J Cell Mol Med. 19:2432–2440. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li T, Dong ZR, Guo ZY, Wang CH, Tang ZY,
Qu SF, Chen ZT, Li XW and Zhi XT: Aspirin enhances IFN-α-induced
growth inhibition and apoptosis of hepatocellular carcinoma via
JAK1/STAT1 pathway. Cancer Gene Ther. 20:366–374. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cao ZH, Zheng QY, Li GQ, Hu XB, Feng SL,
Xu GL and Zhang KQ: STAT1-mediated down-regulation of Bcl-2
expression is involved in IFN-γ/TNF-α-induced apoptosis in NIT-1
cells. PLoS One. 10:e01209212015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu L, Hao J, Cheng M, Zhang C, Huo C, Liu
Y, Du W and Zhang X: Hyperglycemia-induced Bcl-2/Bax-mediated
apoptosis of Schwann cells via mTORC1/S6K1 inhibition in diabetic
peripheral neuropathy. Exp Cell Res. 367:186–195. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Toscano ECB, Vieira ÉLM, Portela ACDC,
Reis JLJ, Caliari MV, Giannetti AV, Gonçalves AP, Siqueira JM,
Suemoto CK, Leite REP, et al: Bcl-2/Bax ratio increase does not
prevent apoptosis of glia and granular neurons in patients with
temporal lobe epilepsy. Neuropathology. 39:348–357. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang J, Xia Y, Xu Z and Deng X: Propofol
Suppressed Hypoxia/Reoxygenation-Induced Apoptosis in HBVSMC by
Regulation of the Expression of Bcl-2, Bax, Caspase3, Kir6.1, and
p-JNK. Oxid Med Cell Longev. 2016:15187382016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhao L, Zhu Z, Yao C, Huang Y, Zhi E, Chen
H, Tian R, Li P, Yuan Q, Xue Y, et al: VEGFC/VEGFR3 Signaling
Regulates Mouse Spermatogonial Cell Proliferation via the
Activation of AKT/MAPK and Cyclin D1 Pathway and Mediates the
Apoptosis by affecting Caspase 3/9 and Bcl-2. Cell Cycle.
17:225–239. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rashighi M, Agarwal P, Richmond JM, Harris
TH, Dresser K, Su MW, Zhou Y, Deng A, Hunter CA, Luster AD, et al:
CXCL10 is critical for the progression and maintenance of
depigmentation in a mouse model of vitiligo. Sci Transl Med.
6:223ra232014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Harris JE: IFN-γ in Vitiligo, Is It the
Fuel or the Fire? Acta Derm Venereol. 95:643–644. 2015.PubMed/NCBI
|