Introduction
Infantile hemangioma (IH) is one of the most common
vascular tumors that occurs during childhood (1). IH can lead to life-threatening
disease owing to rapid growth and invasion of tumor cells. IHs
consist of two phases: The proliferating phase and the involuting
phase. The proliferating phase is characterized by dysregulated
proliferation of the immature endothelial cells, whereas the
involuting phase is characterized by the appearance of larger and
fewer capillary-like vessels (2).
Although several genes have been reported to be involved during the
development of IH (3,4), the mechanism underlying IH
progression is not yet completely understood.
Increasing evidence suggests that non-coding RNAs
serve important roles during the progression of a number of
diseases (5). Long non-coding RNAs
(lncRNAs) are a type of non-coding RNA that are >200 nucleotides
in length. Recently, a number of lncRNAs have been reported to have
crucial roles during cancer cell proliferation, apoptosis and
metastasis (6,7). Microarray and RNA-sequencing methods
have previously been used to analyze lncRNA expression profiles
during IH, which identified differentially expressed lncRNAs and
microRNAs (miRNAs/miRs), and allowed for competing endogenous RNA
(ceRNA) and lncRNA-mRNA co-expression networks to be constructed
(8,9). Metastasis-associated lung
adenocarcinoma transcript 1 (MALAT1) is upregulated in IH tissues,
and loss of MALAT1 expression has been shown to inhibit IH cell
proliferation, migration and tube formation, and also promotes cell
apoptosis by sponging miR-424 to inactivate the MEKK3/NF-κB
signaling pathway (10).
Furthermore, the lncRNA linc00152 is upregulated in IH tissues, and
loss of lin00152 expression inhibits cell proliferation and induces
cell apoptosis by inactivating the AKT/mTOR and NOTCH1 signaling
pathways (11). The expression of
linc00152 in hemangioma tissues was higher in the proliferating
phase compared with the involuting phase. Depletion of linc00152
inhibited hemangioma tumor progression of by sponging miR-139-5p
and by downregulating its target gene tumor protein D52 (12).
Nuclear paraspeckle assembly transcript 1 (NEAT1) is
an oncogenic lncRNA that confers docetaxel resistance in prostate
cancer cells by sponging miR-204-5p and miR-34a-5p (13). NEAT1 has been shown to enhance
bladder cancer progression by negatively regulating miR-410 and
positively regulating downstream HMGB1 (14). During endometrial cancer, NEAT1
promotes cell proliferation, migration and invasion by sponging
miR-144-3p, leading to the activation of its target gene enhancer,
zeste 2 polycomb repressive complex 2 subunit (15). However, the roles and mechanisms
underlying NEAT1 during IH progression are not yet completely
understood. Hypoxia-inducible factor 1α (HIF1α) is a crucial
regulator during hypoxia-induced angiogenesis, but it is also
involved in the growth of hemangiomas (16,17).
Curcumin-mediated downregulation of HIF1α inhibits the
proliferation of human hemangioma endothelial cells (HemECs)
(18). Furthermore, by using
starBase, NEAT1 was predicted as a ceRNA that regulates HIF1α by
sponging miR-33a-5p (19).
Therefore, the roles of the NEAT1/miR-33a-5p/HIF1α axis during the
tumorigenesis of IH along with the downstream regulatory mechanisms
were assessed in the present study.
Materials and methods
Tissue samples and cell line
Tissue specimens (n=12 patients/group), including
normal subcutaneous tissues and infantile hemangioma tissues in the
involuting stage (9 females and 3 males; median age, 7 months) and
proliferating stage (10 females and 2 males; median age, 6 months),
were obtained from patients at Kunming Children's Hospital
(Kunming, China) between June 2016 and September 2018. Tissue
samples were immediately frozen at −80°C until further analysis.
The present study was approved by the Ethics Committee of Kunming
Children's Hospital (approval no. 2018-005). Written informed
consent was obtained from the parents/legal guardians of each
patient.
HemECs were isolated from infant hemangioma tissues
in the proliferating phase as previously described (20). HemECs were cultured in human
endothelial-serum free medium (Gibco; Thermo Fisher Scientific,
Inc.) containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
with 5% CO2 at 37°C.
Cell transfection
miR-33a-5p mimic, mimic negative control (NC) and
small interfering (si)RNAs targeted against NEAT1 and HIF1α were
purchased from Shanghai GenePharma Co., Ltd. The sequences were as
follows: miR-33a-5p mimic, 5′-GTGCATTGTAGTTGCATTGCA-3′; mimic NC,
5′-TTCTCCGAACGTGTCACGTT-3′; NEAT1 si1, 5′-GCCATCAGCTTTGAATAAATT-3′;
NEAT1 si2, 5′-TGGCTAGCTCAGGGCTTCAG-3′; HIF1α si1,
5′-GCAAGACGTTGTTTGAAATTT-3′; HIF1α si2, 5′-ACACACTGTGTCCAGTTAG-3′;
NC siRNA, 5′-ACTGTTCTATGACTTGTCGTGAATA-3′. HemECs at 50–60%
confluency were transfected with siRNA (50 nM) or miR-33a-5p mimic
(50 nM) using Lipofectamine® 3000 (Gibco; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Following incubation for 48 h at 37°C, reverse
transcription-quantitative PCR (RT-qPCR) and western blotting were
performed.
Cell proliferation
At 24 h post-transfection, HemECs (1×104
cells/well) were seeded into 96-well plates and cultured for 24, 72
or 120 h at 37°C. Subsequently, 10 µl Cell Counting Kit-8 (CCK-8)
solution (Dojindo Molecular Technologies, Inc.) was added to each
well and incubated at 37°C for 1 h, according to the manufacturer's
protocol. The absorbance of each well was measured at a wavelength
of 450 nm using a Bio-Rad 680 microplate reader (Bio-Rad
Laboratories, Inc.). The relative proliferation was calculated by
normalizing the absorbance of the NEAT1 si1- or si2-transfected
cells to that of the NC group at the same time point.
Migration and invasion assays
To assess invasion, the Transwell membranes were
coated with Matrigel® (BD Biosciences) for 1 h at 37°C.
Subsequently, the membranes were hydrated in 100% FBS for 2 h at
37°C. A total of 1×105 cells in serum-free culture
medium were seeded into the upper chamber. Culture medium with 20%
FBS was plated into the lower chambers. Following incubation at
37°C for 36 h, the Transwell membranes were stained with 0.1%
crystal violet solution for 10 min at room temperature. The average
number of invasive cells per field was assessed by counting 10
random fields under a light microscope (magnification, ×40). To
assess migration, the aforementioned protocol was performed;
however, the Transwell membranes were not pre-coated with
Matrigel®.
RT-qPCR
Total RNA was extracted from tissues and cells using
the RNeasy Mini kit (Qiagen GmbH), according to the manufacturer's
protocol. Total RNA was reverse-transcribed and miR-33a-5p
expression was detected using the Hairpin-it™ qRT-PCR Primer Set
for miR-33a-5p (Shanghai GenePharma Co., Ltd.). The other mRNAs
were reverse-transcribed into cDNA using M-MLV reverse
transcriptase (BioTeke Corporation) in the presence of oligo(dT)
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. The expression levels of NEAT1, HIF1α and
c-myc were determined by qPCR using the Power SYBR™ Green PCR
master mix (Thermo Fisher Scientific, Inc.), according to the
manufacturer's instruction. The PCR procedure was as follows: 95°C
for 10 min; followed by 40 cycles of 95°C for 15 sec and 60°C for 1
min. The following primer pairs were used for qPCR: NEAT1 forward,
5′-GTGGCTGTTGGAGTCGGTAT-3′, reverse 5′-TAACAAACCACGGTCCATGA-3′;
c-myc, forward 5′-GGCTCCTGGCAAAAGGTCA-3′, reverse
5′-CTGCGTAGTTGTGCTGATGT-3′; HIF1α, forward
5′-GAACGTCGAAAAGAAAAGTCTCG-3′, reverse
5′-CCTTATCAAGATGCGAACTCACA-3′; GAPDH, forward
5′-CCAGGTGGTCTCCTCTGA-3′, reverse 5′-GCTGTAGCCAAATCGTTGT-3′; U6,
forward 5′-GCTTCGGCAGCACATATACTAAAAT-3′, reverse
5′-CGCTTCACGAATTTGCGTGTCAT-3′. mRNA and miRNA expression levels
were quantified using the 2−ΔΔCq method (21), and normalized to the internal
reference genes GAPDH and U6, respectively.
Western blotting
Total protein was extracted using RIPA protein
extraction buffer (Beyotime Institute of Biotechnology) and
quantified using the BCA Protein assay kit (Beyotime Institute of
Biotechnology). Equal amounts of proteins (7–10 µg) were separated
by 10% SDS-PAGE and transferred to a PVDF membrane (EMD Millipore).
After blocking with 5% skimmed milk for 1 h at room temperature,
the membrane was incubated at 4°C overnight with the following
primary antibodies: Anti-HIF1α (cat. no. ab51608; 1:1,000; Abcam),
anti-NF-κB (p65; cat. no. ab16502; 1:1,000; Abcam),
anti-phosphorylated (p)-NF-κB (p-p65; cat. no. ab86299; 1:1,000;
Abcam) and anti-β-actin (cat. no. 66009-1-IG; 1:20,000; ProteinTech
Group, Inc.). Subsequently, the membrane was incubated with
horseradish peroxidase-conjugated goat anti-rabbit IgG (1:5,000;
cat. no. 5210-0174) and goat anti-mouse IgG (1:5,000; cat. no.
5210-0185; KPL, Inc.) secondary antibodies. Protein bands were
visualized by Immobilon Enhanced Chemiluminescence (EMD Millipore).
β-actin was used as the loading control. Optical density values of
the protein bands were semi-quantified and analyzed using a gel
image processing system Image Lab software (version 3.0; Bio-Rad
Laboratories, Inc.).
Dual-luciferase reporter assay
A Dual-Luciferase Reporter assay system (Promega
Corporation) was used to detect the binding between miR-33a-5p and
lncRNA NEAT1 or 3′-untranslated region (UTR) of HIF1α according to
the manufacturer's protocol. HemECs (5×105 cells/well)
were added to a 6-well plate and cultured with 5% CO2 at
37°C. After 24 h, cells were co-transfected with the pGL3-NEAT1 WT,
or pGL3-NEAT1 Mut (pGL3-HIF1α 3′UTR WT or pGL3-HIF1α 3′UTR Mut) and
miR-33a-5p mimic/mimic NC using Lipofectamine® 3000
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. Following incubation for 48 h at 37°C, luciferase
activities were measured. The firefly luciferase enzyme activity
was normalized to Renilla luciferase enzyme activity.
StarBase database
The starBase v2.0 database (http://starbase.sysu.edu.cn/) is a powerful database
to study non-coding RNAs, such as lncRNA, miRNA and circRNA. The
starBase database was used to predict the binding between NEAT1 and
miR-33a-5p, and between miR-33a-5p and the 3′UTR of HIF1α.
Statistical analysis
Statistical analyses were performed using GraphPad
Prism software (version 6; GraphPad Software, Inc.). Data are
presented as the mean ± SD. Student's t-test and ANOVA (followed by
Tukey's post hoc test for multiple comparisons) were used to
analyze the differences. Correlations were analyzed by Pearson's
correlation coefficient. P<0.05 was considered to indicate a
statistically significant difference.
Results
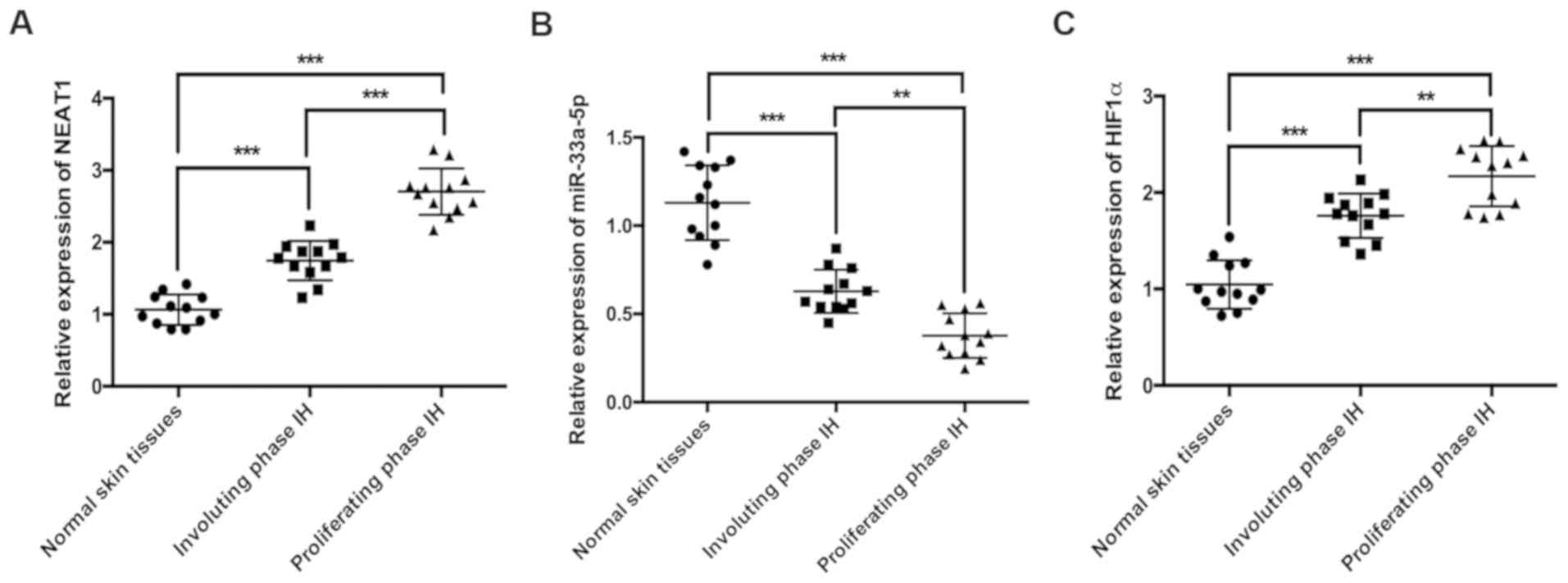
NEAT1 and HIF1α expression levels are
increased and miR-33a-5p expression is decreased in IH tissues
To investigate the associations between NEAT1, HIF1α
and miR-33a-5p during IH, the expression levels of the three
molecules were detected in normal skin, proliferating phase IH and
involuting phase IH tissues by RT-qPCR. NEAT1 was significantly
upregulated in IH tissues compared with normal skin tissues
(Fig. 1A), and its expression was
also significantly higher in proliferating phase IH tissues
compared with involuting phase IH tissues (Fig. 1A). By contrast, the expression of
miR-33a-5p was significantly lower in IH tissues compared with
normal skin tissues (Fig. 1B), and
its expression in proliferating phase IH tissues was significantly
lower compared with involuting phase IH tissues (Fig. 1B). HIF1α expression was
significantly increased in IH tissues compared with normal skin
tissues (Fig. 1C), and the
expression of HIF1α in proliferating phase IH tissues was also
significantly higher compared with involuting phase tissues
(Fig. 1C).
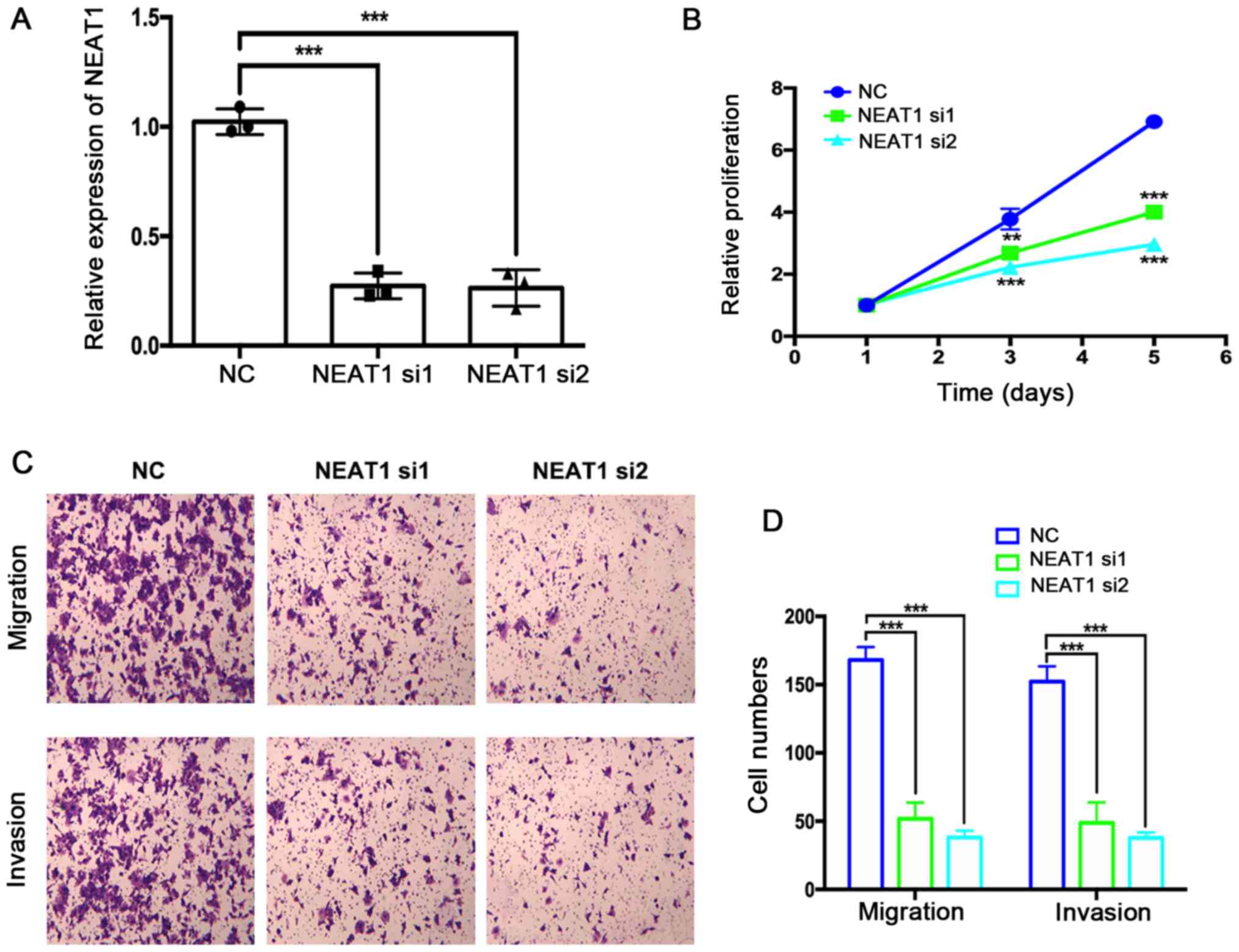
Depletion of NEAT1 mRNA suppresses
HemEC proliferation, migration and invasion
The effects of NEAT1 siRNA transfection on HemEC
proliferation, migration and invasion were assessed using the
CCK-8, Transwell and Matrigel assays, respectively. First, the
efficiency of NEAT1 mRNA depletion by two NEAT1-targeting siRNAs
was verified by RT-qPCR (Fig. 2A).
NEAT1-depleted HemECs exhibited significantly decreased
proliferation compared with the NC group (Fig. 2B). The results of the Transwell and
Matrigel assays suggested that the migration and invasion of HemECs
were also markedly suppressed by depletion of NEAT1 mRNA expression
levels (Fig. 2C and D).
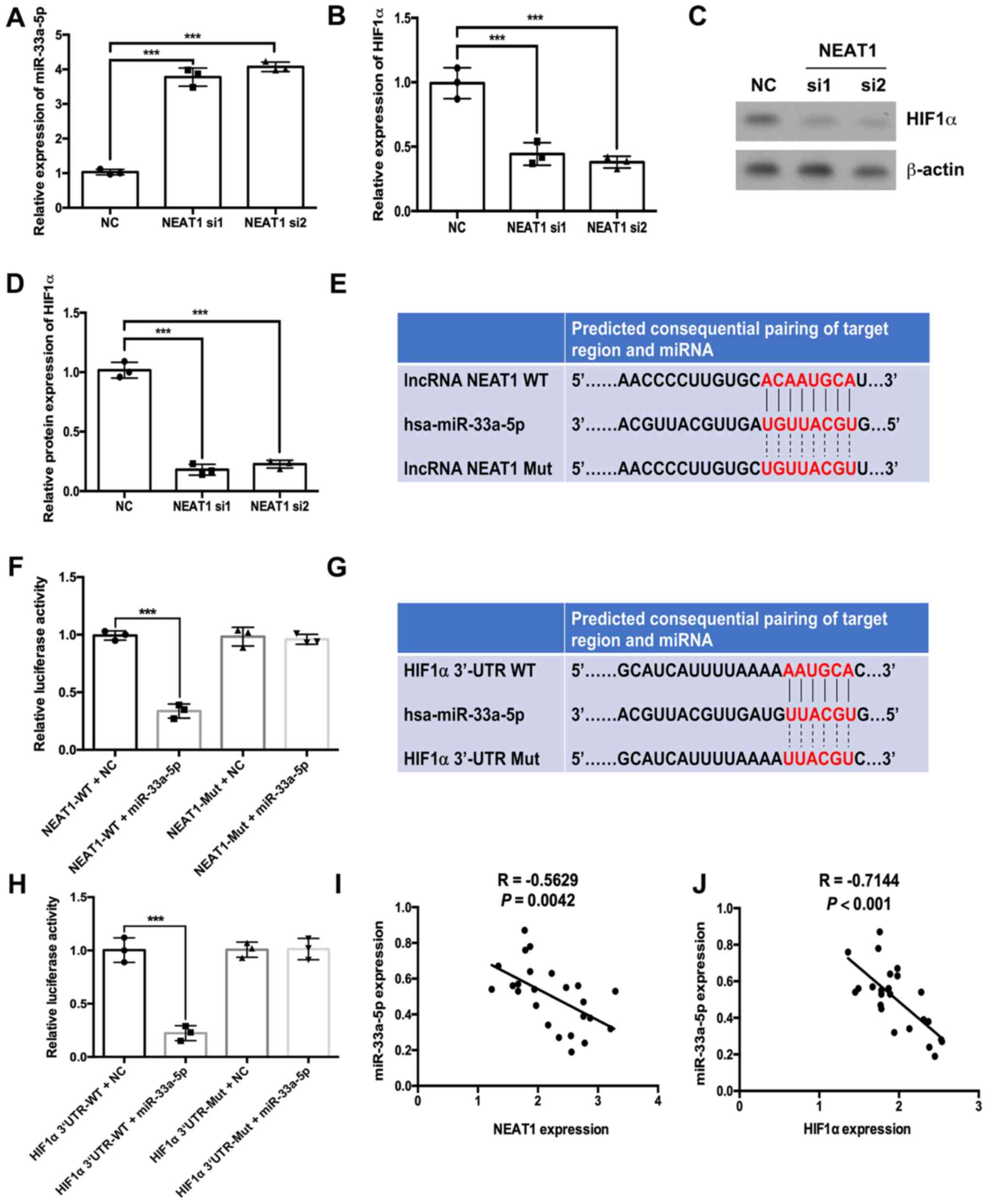
NEAT1 regulates HIF1α expression by
sponging miR-33a-5p in HemECs
RT-qPCR results indicated that depletion of NEAT1
significantly increased the expression levels of miR-33a-5p and
significantly decreased the mRNA expression of HIF1α (Fig. 3A and B, respectively). Western
blotting results further demonstrated that depletion of NEAT1
decreased HIF1α protein expression levels (Fig. 3C and D). Subsequently, a
dual-luciferase assay was performed to evaluate the associations
between NEAT1, miR-33a-5p and HIF1α. Bioinformatics analysis
conducted using the StarBase database suggested that miR-33a-5p
could bind to NEAT1 (Fig. 3E).
Results from the dual-luciferase assay suggested that the
luciferase activities of wild-type NEAT1 were decreased by
miR-33a-5p mimic compared with the miRNA NC group (Fig. 3F). However, in the mutated NEAT1
group, the luciferase activities were not significantly altered in
the miR-33a-5p mimic group compared with the NC group (Fig. 3F). The starBase database also
predicted binding sites between miR-33a-5p and the 3′UTR of HIF1α
(Fig. 3G). The luciferase activity
of the HIF1α 3′UTR-WT + miR-33a-5p group was lower compared with
the HIF1α 3′UTR-WT + NC group (Fig.
3H). However, in the HIF1α 3′UTR-Mut group, the luciferase
activities of the miR-33a-5p mimic group were not significantly
different compared with the NC group (Fig. 3H). Notably, the expression of
miR-33a-5p was negatively correlated with NEAT1 and HIF1α
expressions in IH tissues (Fig. 3I and
J, respectively).
Overexpression of miR-33a-5p or
depletion of HIF1α suppresses HemEC proliferation, migration and
invasion
The effects of miR-33a-5p overexpression and HIF1α
depletion on HemEC proliferation, migration and invasion were
assessed. RT-qPCR results indicated that miR-33a-5p mimic
transfection significantly increased the expression of miR-33a-5p
in HemECs (Fig. 4A), which
significantly inhibited HemEC proliferation, migration and invasion
(Fig. 4B-D, respectively). HIF1α
siRNA transfection resulted in decreased HIF1α mRNA and protein
expression levels compared with the NC group (Fig. 3E-G). HIF1α depletion also
significantly suppressed HemEC proliferation, migration and
invasion (Fig. 4H-J). Furthermore,
miR-33a-5p overexpression downregulated the mRNA and protein
expression levels of HIF1α (Fig. 4K
and L).
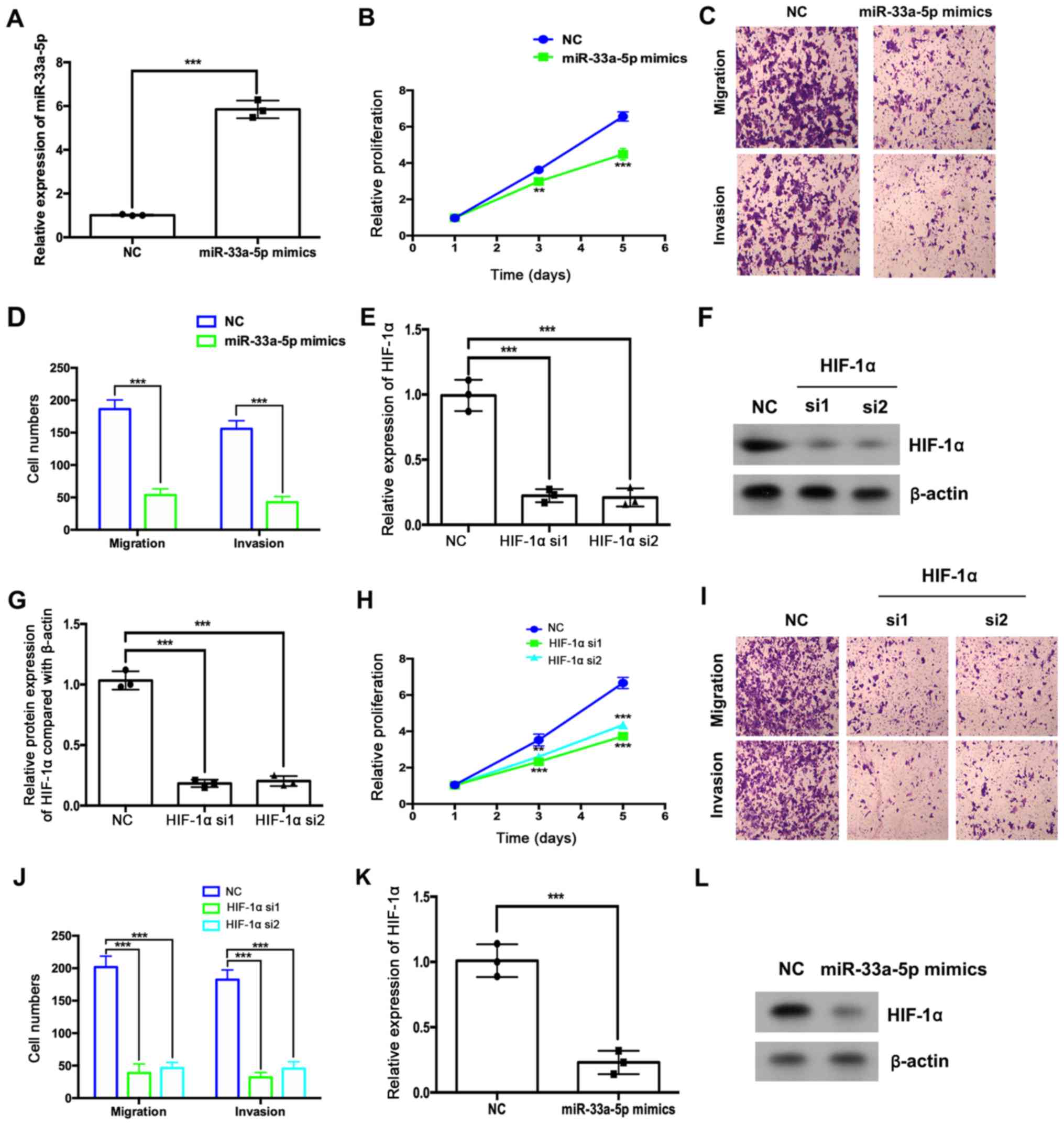 | Figure 4.miR-33a-5p overexpression or HIF1α
depletion suppresses HemEC proliferation, migration and invasion.
(A-C) HemECs were transfected with miR-33a-5p mimics or miRNA NC.
(A) The expression levels of miR-33a-5p were analyzed by RT-qPCR.
(B) The relative proliferation was analyzed using a CCK-8 assay.
(C) Migratory and (D) invasive abilities were analyzed by Transwell
and Matrigel assays, respectively. (F-J) HemECs were transfected
with HIF1α or NC siRNAs. (E) The expression levels of HIF1α were
analyzed by RT-qPCR. (F and G) The protein expression levels of
HIF1α were analyzed by western blotting. (H) The relative
proliferation of HemECs was analyzed using a CCK-8 assay. (I)
Migratory and (J) invasive abilities of HemECs were analyzed by
Transwell and Matrigel assays, respectively. HemECs were
transfected with miR-33a-5p mimics or miRNA NC and the expression
levels of HIF1α (K) mRNA were analyzed by RT-qPCR and (L) protein
was analyzed by western blotting. **P<0.01 and ***P<0.001 vs.
NC group at the same time point or as indicated in figure. CCK-8,
Cell Counting Kit-8; HemEC, hemangioma endothelial cells; HIF1α,
hypoxia-inducible factor 1α; miR, microRNA; NC, negative control;
RT-qPCR, reverse transcription-quantitative PCR; si, small
interfering RNA. |
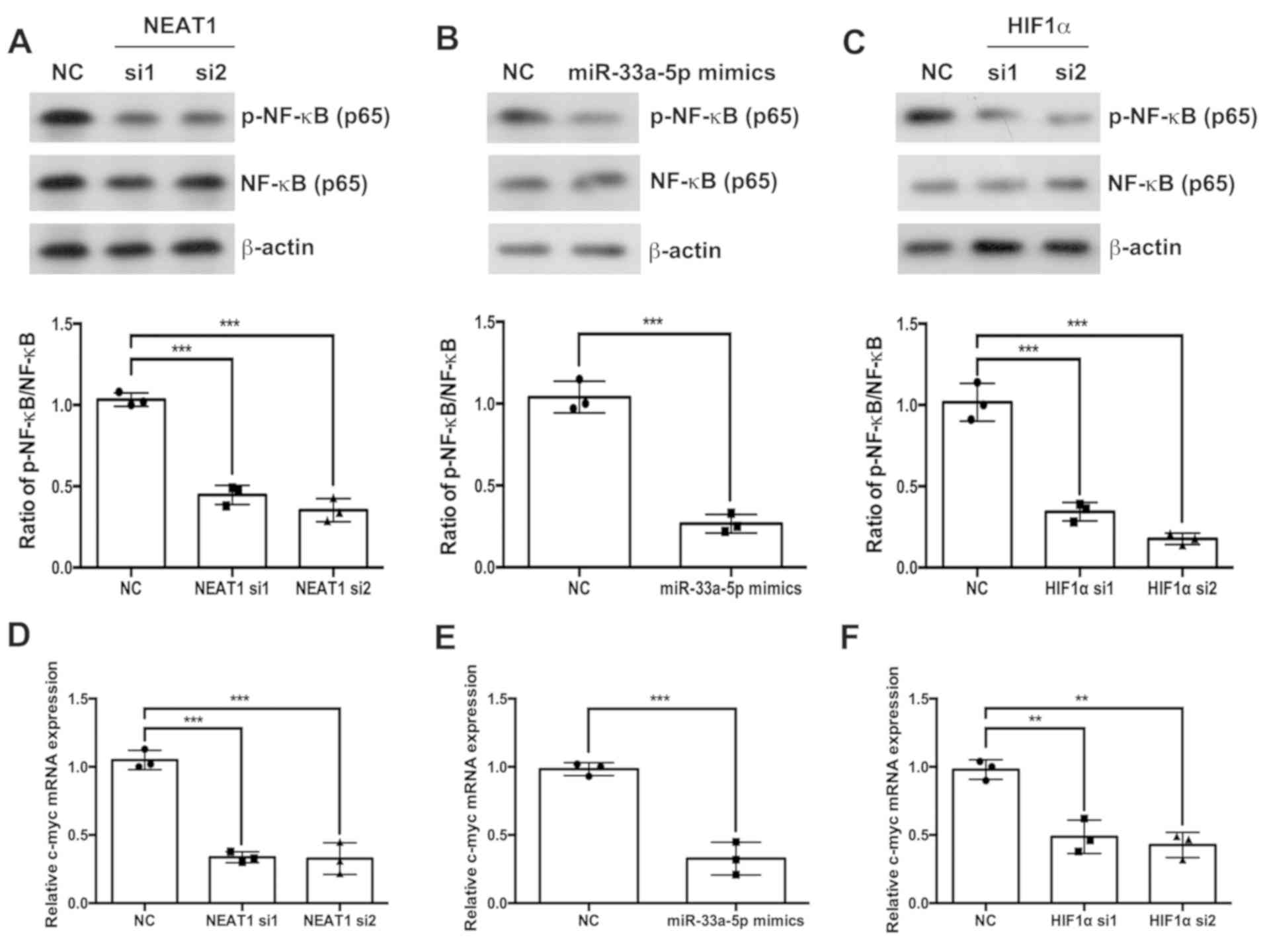
NEAT1/miR-33a-5p/HIF1α axis regulates
the NF-κB signaling pathway
The effects of the NEAT1/miR-33a-5p/HIF1α axis on
the NF-κB signaling pathway were also investigated. Western
blotting results indicated that the depletion of NEAT1 or HIF1α
(Fig. 5A and C, respectively) and
the overexpression of miR-33a-5p (Fig.
5B) significantly inhibited the phosphorylation of p65 NF-κB
compared with the respective NC groups. Similarly, the depletion of
NEAT1 or HIF1α and the overexpression of miR-33a-5p decreased the
mRNA expression levels of c-myc (Fig.
5D-F).
Discussion
A number of lncRNAs have been reported to display
important roles during IH progression. For example, the lncRNA
linc00342 enhances HemEC proliferation and suppresses HemEC
apoptosis by sponging miR-3619-5p and increasing the expression of
its target gene hepatoma-derived growth factor (22). Higher expression levels of lncRNA
urothelial cancer-associated 1 (UCA1) have previously been detected
in proliferating phase hemangioma samples compared with involuting
phase hemangioma samples (23).
UCA1 depletion decreased EOMA mouse hemangioendothelioma
endothelial cell viability, migration and invasion, and also
promoted EOMA cell apoptosis by regulating miR-200c (23). However, the roles and mechanisms
underlying NEAT1 during IH are not yet completely understood.
The results of the present study indicated that the
expression of NEAT1 was higher in IH tissues compared with normal
skin tissues, in particular in proliferating phase samples compared
with involuting phase samples. In vitro studies indicated
that NEAT1 depletion inhibited HemEC proliferation, migration and
invasion. The starBase database predicted that NEAT1 may function
as a sponge for miR-33a-5p and further regulate downstream HIF1α.
The results of the present study further suggested the binding
between miR-33a-5p and NEAT1, as well as between miR-33a-5p and the
3′UTR of HIF1α.
miR-33a-5p expression is downregulated in prostate
cancer tissues, and low expression levels are positively associated
with poor prognosis and bone metastasis-free survival (24). miR-33a-5p, which was
transcriptionally suppressed by zinc-finger E-box-binding homeobox
1 overexpression in prostate cancer, affected migration, invasion
and the epithelial-to-mesenchymal transition of cancer cells
(24). During colorectal cancer,
miR-33a-5p is also downregulated, and its overexpression
significantly suppressed colorectal cancer cell proliferation,
colony formation, G1/S progression and migration
(25). To the best of our
knowledge, the present study suggested for the first time that
miR-33a-5p was downregulated in IH tissues and inactivated by
binding to an abundant amount of lncRNA NEAT1.
HIF1α plays important roles during the development
of IH. HIF1α is significantly upregulated in proliferating IH
tissues compared with involuting IH tissues, and an in vitro
study indicated that hypoxia upregulated HIF1α expression in
hemangioma stem cells (26).
Curcumin-mediated HIF1α downregulation inhibits cell proliferation
and induces infantile hemangioma endothelial cell apoptosis
(18). The results of the present
study suggested that HIF1α was highly expressed in IH tissues, and
the expression levels of HIF1α in proliferating IH tissues were
higher compared with involuting IH tissues. The results also
further suggested that depletion of NEAT1 mRNA downregulated HIF1α,
and this may be through sponging miR-33a-5p during IH, although
further studies are needed to confirm this. A previous study
reported that NEAT1 bound and inactivated miR-186-5p thereby
upregulating HIF1α and promoting cell proliferation and invasion
during osteosarcoma (21).
Collectively, the aforementioned studies suggested that NEAT1
regulated HIF1α through different mechanisms in different types of
cancer.
The NF-κB signaling pathway serves important roles
during cancer progression (27,28).
The expression levels of NF-κB (p65), p-NF-κB inhibitor-α (IκBα)
and p-NF-κB inhibitor-β (IKKβ) were higher in proliferative
hemangioma compared with involutional hemangioma (29). Propranolol treatment could decrease
the expression of NF-κB (p65), and inhibit the phosphorylation of
IκBα and IKKβ in vitro and in vivo during IH
(30). Inactivating the NF-κB
signaling pathway further decreased vascular endothelial growth
factor A expression levels in IH-derived stem cells (31). The results of the present study
indicated that the NF-κB signaling pathway could be regulated by
lncRNA NEAT1 during IH.
The present study suggested that lncRNA NEAT1
depletion suppressed the tumorigenesis of infantile hemangioma by
competitively binding miR-33a-5p to stimulate the HIF1α/NF-κB
signaling pathway. Therefore, blocking the NEAT1/miR-33a-5p/HIF1α
signaling pathway may represent a potential antitumor therapeutic
strategy.
Acknowledgements
Not applicable.
Funding
The present study was funded by The Gathering Plan
of Scientific and Technological Innovation Elements (grant no.
2017-1-S-16631).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LY and HS designed the study. LY, HS, NZ, LX, ML, LL
and YX performed the experiments. ZZ, LZ and YX analyzed the data.
LY and HS wrote the paper.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Kunming Children's Hospital (approval no. 2018-005).
Written informed consent was obtained from the parents/legal
guardians of each patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Acevedo LM and Cheresh DA: Suppressing
NFAT increases VEGF signaling in hemangiomas. Cancer Cell.
14:3358–430. 2008. View Article : Google Scholar
|
|
2
|
Leaute-Labreze C, Prey S and Ezzedine K:
Infantile haemangioma: Part I. Pathophysiology, epidemiology,
clinical features, life cycle and associated structural
abnormalities. J Eur Acad Dermatol Venereol. 25:1245–1253. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yang L, Dai J, Li F, Cheng H, Yan D and
Ruan Q: The expression and function of miR-424 in infantile skin
hemangioma and its mechanism. Sci Rep. 7:118462017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Calicchio ML, Collins T and Kozakewich HP:
Identification of signaling systems in proliferating and involuting
phase infantile hemangiomas by genome-wide transcriptional
profiling. Am J Pathol. 174:1638–1649. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu ZZ, Tian YF, Wu H, Ouyang SY and Kuang
WL: LncRNA H19 promotes glioma angiogenesis through
miR-138/HIF-1α/VEGF axis. Neoplasma. 67:111–118. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang J, Qiu Q, Qian X, Yi J, Jiao Y, Yu M,
Li X, Li J, Mi C, Zhang J, et al: Long noncoding RNA LCAT1
functions as a ceRNA to regulate RAC1 function by sponging
miR-4715-5p in lung cancer. Mol Cancer. 18:1712019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu S, Qiu J, He G, Liang Y, Wang L, Liu C
and Pan H: LncRNA MALAT1 acts as a miR-125a-3p sponge to regulate
FOXM1 expression and promote hepatocellular carcinoma progression.
J Cancer. 10:6649–6659. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu X, Lv R, Zhang L, Xu G, Bi J, Gao F,
Zhang J, Xue F, Wang F, Wu Y, et al: Long noncoding RNA expression
profile of infantile hemangioma identified by microarray analysis.
Tumour Biol. Oct 5–2016.doi: 10.1007/s13277-016-5434-y (Epub ahead
of print). View Article : Google Scholar
|
|
9
|
Li J, Li Q, Chen L, Gao Y, Zhou B and Li
J: Competitive endogenous RNA networks: Integrated analysis of
non-coding RNA and mRNA expression profiles in infantile
hemangioma. Oncotarget. 9:11948–11963. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li MM, Dong CX, Sun B, Lei HZ, Wang YL,
Gong YB, Sun LL and Sun ZW: LncRNA-MALAT1 promotes tumorogenesis of
infantile hemangioma by competitively binding miR-424 to stimulate
MEKK3/NF-κB pathway. Life Sci. 239:1169462019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Li M, Dong C, Ma Y, Xiao L, Zuo S,
Gong Y, Ren T and Sun B: Linc00152 knockdown inactivates the
Akt/mTOR and Notch1 pathways to exert its anti-hemangioma effect.
Life Sci. 223:22–28. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang SJ, Li YJ, Gao B, Li XL, Li YT and He
HY: Long non-coding RNA 00152 slicing represses the growth and
aggressiveness of hemangioma cell by modulating miR-139-5p. Biomed
Pharmacother. 120:1093852019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang X, Guo S, Zhang Y, Zhao Y, Li X, Jia
Y, Xu Y and Ma B: LncRNA NEAT1 promotes docetaxel resistance in
prostate cancer by regulating ACSL4 via sponging miR-34a-5p and
miR-204-5p. Cell Signal. 65:1094222020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shan G, Tang T, Xia Y and Qian HJ: Long
non-coding RNA NEAT1 promotes bladder progression through
regulating miR-410 mediated HMGB1. Biomed Pharmacother.
121:1092482019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang W, Ge L, Xu XJ, Yang T, Yuan Y, Ma XL
and Zhang XH: LncRNA NEAT1 promotes endometrial cancer cell
proliferation, migration and invasion by regulating the
miR-144-3p/EZH2 axis. Radiol Oncol. 53:434–442. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhong H, De Marzo AM, Laughner E, Lim M,
Hilton DA, Zagzag D, Buechler P, Isaacs WB, Semenza GL and Simons
JW: Overexpression of hypoxia-inducible factor 1alpha in common
human cancers and their metastases. Cancer Res. 59:5830–5835.
1999.PubMed/NCBI
|
|
17
|
Kleinman ME, Greives MR, Churgin SS,
Blechman KM, Chang EI, Ceradini DJ, Tepper OM and Gurtner GC:
Hypoxia-induced mediators of stem/progenitor cell trafficking are
increased in children with hemangioma. Arterioscler Thromb Vasc
Biol. 27:2664–2670. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lou S, Wang Y, Yu Z, Guan K and Kan Q:
Curcumin induces apoptosis and inhibits proliferation in infantile
hemangioma endothelial cells via downregulation of MCL-1 and
HIF-1α. Medicine (Baltimore). 97:e95622018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42((Database Issue)): D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Khan ZA, Melero-Martin JM, Wu X, Paruchuri
S, Boscolo E, Mulliken JB and Bischoff J: Endothelial progenitor
cells from infantile hemangioma and umbilical cord blood display
unique cellular responses to endostatin. Blood. 108:915–921. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu Z, Kang Z, Dai Y, Zheng H and Wang Y:
Long noncoding RNA LINC00342 promotes growth of infantile
hemangioma by sponging miR-3619-5p from HDGF. Am J Physiol Heart
Circ Physiol. 317:H830–H839. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang J and Zhang C: Silence of long
non-coding RNA UCA1 inhibits hemangioma cells growth, migration and
invasion by up-regulation of miR-200c. Life Sci. 226:33–46. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dai Y, Wu Z, Lang C, Zhang X, He S, Yang
Q, Guo W, Lai Y, Du H, Peng X and Ren D: Copy number gain of ZEB1
mediates a double-negative feedback loop with miR-33a-5p that
regulates EMT and bone metastasis of prostate cancer dependent on
TGF-β signaling. Theranostics. 9:6063–6079. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yan Y, Zhang D, Lei T, Zhao C, Han J, Cui
J and Wang Y: MicroRNA-33a-5p suppresses colorectal cancer cell
growth by inhibiting MTHFD2. Clin Exp Pharmacol Physiol.
46:928–936. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xia HF, Zhu JY, Wang JN, Ren JG, Cai Y,
Wang FQ, Zhang W, Chen G, Zhao YF and Zhao JH: Association of ATF4
expression with tissue hypoxia and M2 macrophage infiltration in
infantile hemangioma. J Histochem Cytochem. 65:285–294. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen L, De Menna M, Groenewoud A, Thalmann
GN, Kruithof-de Julio M and Snaar-Jagalska BE: A NF-kB-Activin A
signaling axis enhances prostate cancer metastasis. Oncogene.
39:1634–1651. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xiu Y, Dong Q, Fu L, Bossler A, Tang X,
Boyce B, Borcherding N, Leidinger M, Sardina JL, Xue HH, et al:
Coactivation of NF-κB and Notch signaling is sufficient to induce B
cell transformation and enables B-myeloid conversion. Blood.
135:108–120. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang Y, Xu W, Li S, Zhong Z, Li Y, Wang W
and Sun C: Expression and significance of the proteins in TSP-1 and
NF-κB signal pathways of infantile capillary hemangioma. Zhonghua
Zheng Xing Wai Ke Za Zhi. 32:441–446. 2016.(In Chinese). PubMed/NCBI
|
|
30
|
Xu W, Li S, Yu F, Zhang Y, Yang X, An W,
Wang W and Sun C: Role of Thrombospondin-1 and nuclear Factor-κB
signaling pathways in antiangiogenesis of infantile hemangioma.
Plast Reconstr Surg. 142:310e–321e. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Greenberger S, Adini I, Boscolo E,
Mulliken JB and Bischoff J: Targeting NF-κB in infantile
hemangioma-derived stem cells reduces VEGF-A expression.
Angiogenesis. 13:327–335. 2010. View Article : Google Scholar : PubMed/NCBI
|