Introduction
Oral inflammation, including periodontitis and
peri-implant disease, has become a common challenge encountered in
the dental clinic. It directly leads to bone resorption around the
tooth or implant, subsequently causing loosening of the tooth and
implant failure (1,2). There is a high prevalence of
periodontitis and peri-implant disease in the United States
(3,4). The National Health and Nutrition
Examination Survey conducted between 2009 and 2014 reporterd that
42.2% of US dentate adults aged ≥30 years had periodontitis
(5). Derks et al (6) found that 45% of 588 patients who had
received implant-supported oral rehablitation therapy 9 years
earlier presented with peri-implant disease. The principal
etiological factor for these inflammatory diseases is plaque
accumulation (7,8). Among the vast array of oral bacteria,
Porphyromonas gingivalis serves a particularly important
role in both periodontitis and peri-implant diseases, resulting
from the expression of various virulence factors, the most
recognizable of which is lipopolysaccharide (LPS) (9,10).
LPS, also known as endotoxin, induces a variety of cell types, such
as osteoblasts (11), human
umbilical vein endothelial cells (12) and gingival fibroblasts (13), to overexpress related inflammatory
factors, including interleukin (IL)-1β, IL-6 and tumor necrosis
factor α (TNF-α). Recently, it was demonstrated that LPS could
induce the overexpression of TNF-α, IL-1β and IL-6 in bone marrow
mesenchymal stem cells (BMSCs) (14). To identify the molecular mechanism
underlying LPS-mediated inflammation, the majority of studies have
focused on the vital role served by nuclear factor (NF)-κB, as LPS
can mediate the phosphorylation of p65, which is critical for the
transactivation of NF-κB (15).
By contrast, less attention has been paid to the
nucleotide-binding oligomerization domain-like receptor protein 3
(NLRP3) inflammasome in studies on LPS-induced inflammation. LPS is
widely known to stimulate the NLRP3 inflammasome in different cell
types (16–18). Additionally, the NLRP3 inflammasome
serves a significant role in LPS-mediated inflammation, as its
upregulation is conducive to the secretion of IL-1β and the
excessive release of IL-1β contributes to the cellular inflammatory
response (19–21). Thus, there is an urgent need for a
novel therapeutic approach that blocks activation of the NLRP3
inflammasome and controls inflammation triggered by LPS, thereby
eradicating the infection that occurs in both soft and hard tissue
and eventually facilitating the development of bone around the
tooth or implant.
N-acetyl cysteine (NAC), a precursor of glutathione,
is a typical oxygen-free radical scavenger and anti-inflammatory
agent (22). It is used as a tool
for inhibiting the release of inflammatory factors and reducing the
inflammation triggered by LPS (11), as well as in the treatment of
different diseases (23–25), including cardiovascular diseases,
respiratory diseases and cancer. In our previous studies (10,11,26),
human umbilical vein endothelial cells, osteoblasts and periodontal
ligament fibroblasts were pretreated with NAC prior to their
stimulation with LPS. The results demonstrated that this
pretreatment led to the inhibition of IL-1β, IL-6 and TNF-α, and
the inflammatory response initiated by LPS.
Based on these findings, it was hypothesized that
NAC may inhibit the inflammatory response in LPS-induced BMSCs. To
test this hypothesis and to clarify the molecular mechanism
underlying the anti-inflammatory action of NAC, the effect of NAC
on inflammation stimulated by LPS was evaluated. Whether NAC could
control the inflammatory response in LPS-mediated BMSCs was also
assessed and the associated anti-inflammatory mechanisms were
determined.
Materials and methods
Cell culture
BMSCs (cat. no. BNCC340947; BeNa Culture Collection;
Beijing Beina Chunglian Biotechnology Research Institute) were
maintained in Dulbecco's modified Eagle's medium (DMEM; HyClone;
Cytiva) supplemented with 1% (v/v) penicillin-streptomycin
(Beyotime Institute of Biotechnology) and 10% (v/v) fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.) and then cultured at
37°C with 5% CO2. BMSCs were used from the fifth to
eighth generations.
Four experimental groups were established (Table I): i) Control; ii) NAC + LPS; iii)
resveratrol (Res, 50 µM; Sigma-Aldrich; Merck KGaA) + LPS; and iv)
LPS. Cells in the NAC + LPS group were incubated with 1 mM NAC
(Sigma-Aldrich; Merck KGaA) for 1 h, while cells in the Res + LPS
group were cultured for 2 h in the dark. Next, all groups,
excluding the control group, were stimulated with LPS (1 µg/ml;
Sigma-Aldrich; Merck KGaA) for 24 h. All experiments were performed
at 37°C with 5% CO2.
 | Table I.Experimental groups. |
Table I.
Experimental groups.
| Groups | Control | NAC + LPS | Res + LPS | LPS |
|---|
| NAC | – | + | – | – |
| LPS | – | + | + | + |
| Res | – | – | + | – |
Cell proliferation assay
The effects of NAC on BMSC proliferation were
determined by a Cell Counting Kit-8 (CCK-8) assay. First,
1×105/cm2 BMSCs were seeded into a 96-well
plate and incubated for 24 h at 37°C. Next, BMSCs were pretreated
with NAC at concentrations of 0, 0.1, 0.5, 1 and 2 mM for 1 h prior
to stimulation with LPS at 37°C. The cells were then cultured for
24 h at 37°C. Subsequently, the original medium was carefully
removed prior to the addition of 10 µl CCK-8 reagent (Beyotime
Institute of Biotechnology) and 100 µl DMEM to each well. Following
incubation at 37°C for 1 h, a microplate reader (Thermo Fisher
Scientific, Inc.) was used to detect the absorbance of each well at
450 nm. The appropriate concentration of NAC for cell proliferation
was selected with three replicates and the assay was conducted
according to the manufacturer's protocols.
Reverse transcription-quantitative
(RT-q) PCR
BMSCs (1×105/cm2) were seeded
into four plates. Subsequently, the cells were collected and total
RNA of apoptosis-associated speck-like protein containing a CARD
(ASC), NLRP3, caspase-1, thioredoxin-interacting protein (TXNIP)
and thioredoxin (TRX) was isolated from the BMSCs using
TRIzol® reagent (Thermo Fisher Scientific, Inc.). The
SYBR-Green Real-Time PCR Master Mix (KAPA Biosystems; Roche
Diagnostics) was used for qPCR, and cDNA synthesis was performed
using the PrimeScript RT Reagent kit (Takara Bio, Inc.) in a
reaction that included 1 µl cDNA, 0.5 µl forward primer, 0.5 µl
reverse primer, 10 µl SYBR FAST qPCR Master Mix and 8 µl
ddH2O to a total volume of 20 µl. All reactions were
performed according to the manufacturer's protocols. The PCR
protocol was as follows: 95°C for 3 min, followed by 40 cycles of
95°C for 5 sec, 56°C for 10 sec, and 72°C for 25 sec. Assays were
conducted in triplicate and β-actin served as the reference gene.
The primers are shown in Table
II. Finally, the values of 2−ΔΔCq reflected the mRNA
abundance (27).
| Table II.Primers for reverse
transcription-quantitative PCR. |
Table II.
Primers for reverse
transcription-quantitative PCR.
| Gene name | Primer sequence
(5′→3′) | Amplicon length
(bp) |
|---|
| ASC | F:
AGCATCCAGCAAACCA | 259 |
|
| R:
GGACCCCATAGACCTCA |
|
| NLRP3 | F:
CATCTTAGTCCTGCCAA | 94 |
|
| R:
CAACAGACGCTACACCC |
|
|
Caspase-1 | F:
TTGAAGAGCAGAAAGCA | 105 |
|
| R:
CAGTAGGAAACTCCGAAG |
|
| TXNIP | F:
CAAGGTAAGTGTGCCG | 105 |
|
| R:
GATTCTGTGAAGGTGATGA |
|
| TRX | F:
CCAACCTTTTGACCCTTT | 143 |
|
| R:
CCCTTCTTTCATTCCCTC |
|
| β-actin | F:
TAGGAGCCAGGGCAGTA | 110 |
|
| R:
CGTTGACATCCGTAAAGAC |
|
Western blot analysis
Western blot analysis was conducted to calculate the
levels of related proteins, including ASC, NLRP3, caspase-1, TXNIP
and TRX, in the BMSCs. The BMSCs were treated as aforementioned,
and the total protein was collected with radioimmunoprecipitation
assay buffer (Thermo Fisher Scientific, Inc.). The total protein
concentration was normalized following quantification with the BCA
Protein Assay (Beijing Solarbio Science & Technology Co., Ltd).
The proteins (20 µg) were then separated via 12% SDS-PAGE
(Sigma-Aldrich; Merck KGaA) and electrotransferred to
polyvinylidene fluoride membranes. The membranes were blocked in 5%
non-fat milk at room temperature for 2 h and then incubated with
the following rabbit primary antibodies for ≥8 h at 4°C: ASC
(1:1,000; cat. no. PAB30696; Bioswamp), caspase-1 (1:1,000; cat.
no. PAB36756; Bioswamp), TXNIP (1:1,000; cat. no. PAB43948;
Bioswamp), TRX (1:1,000; cat. no. PAB32168; Bioswamp), NLRP3
(1:1,000; cat. no. PAB37930; Bioswamp) and β-actin (1:1,000; cat.
no. PAB36265; Bioswamp). Subsequently, the membranes were treated
at room temperature for 1 h using goat anti-rabbit IgG (1:20,000;
cat. no. SAB43714; Bioswamp) labeled with horseradish peroxidase.
The bands were detected by enhanced chemiluminescence (Analytik
Jena AG). Tanon GIS software version 4.2 (Tanon Science &
Technology Co., Ltd.) was used to determine the band intensity.
Enzyme-linked immunoassay (ELISA)
The BMSCs were treated as aforementioned, then the
supernatants were collected, which were centrifuged at 1,000 × g at
room temperature for 10 min. The expression of inflammatory
mediators, including IL-1β, IL-6 and TNF-α, was evaluated. All
steps were in accordance with the protocols of rat IL-1β (cat. no.
RLB00), IL-6 (cat. no. R6000B) and TNF-α (cat. no. RTA00) ELISA
kits (R&D Systems, Inc.). The absorbance of each group was
evaluated at 450 nm with a microplate reader. Finally, the
concentrations of the inflammatory mediators were measured
according to a standard curve.
Statistics analysis
All data are expressed as the mean ± standard
deviation. Data analysis was performed using SPSS 17.0 (SPSS,
Inc.). For multiple comparisons, the differences in variables among
groups were analyzed using one-way ANOVA followed by a Tukey's post
hoc test. For comparisons between two groups, one-way ANOVA was
used. P<0.05 was considered to indicate a statistically
significant difference.
Results
Effect of NAC on the proliferation
rate of LPS-mediated BMSCs
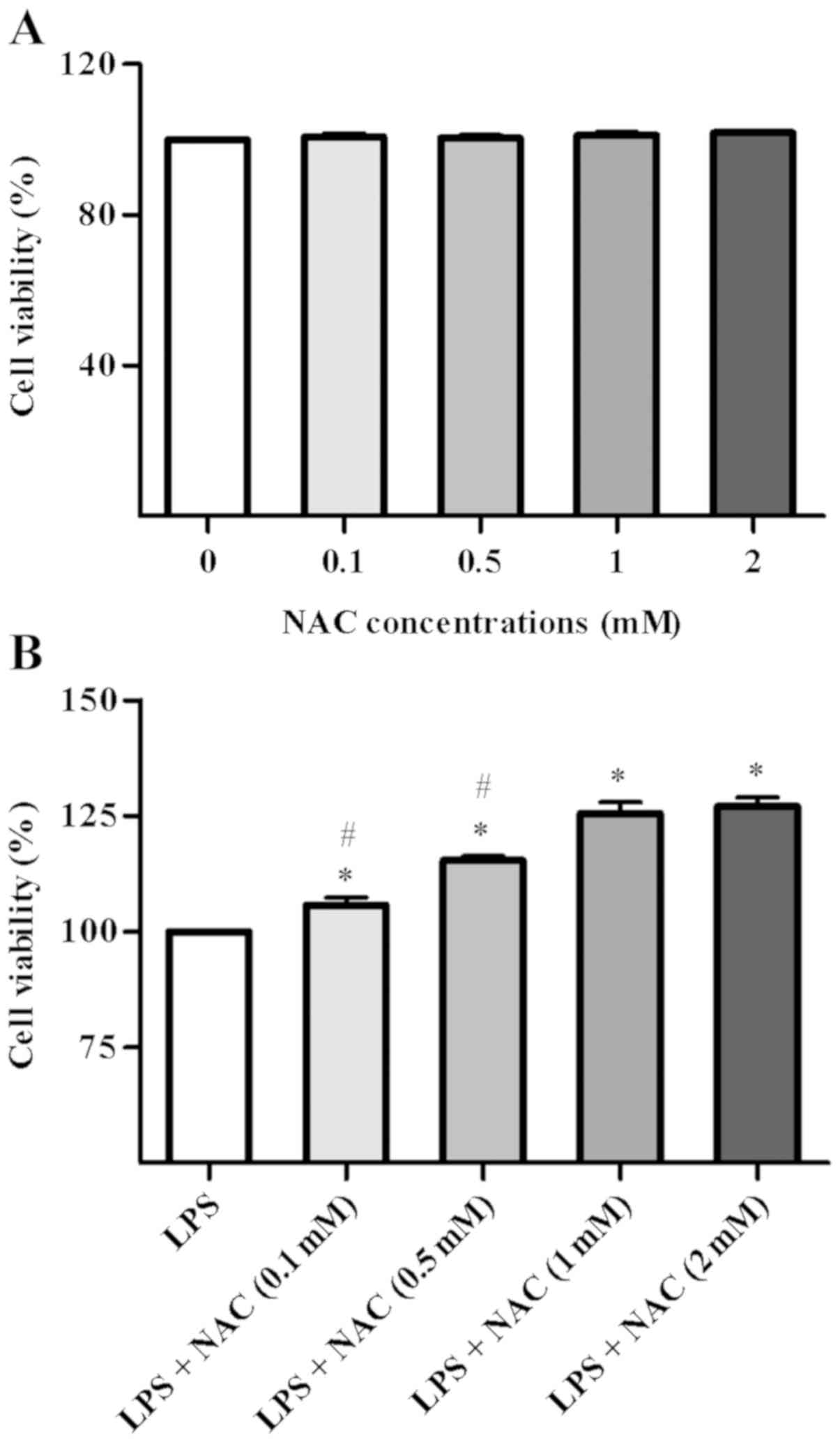
The cytotoxicity of NAC is shown in Fig. 1A. The results demonstrated no
significant changes in the viability of BMSCs following
pretreatment with NAC alone at various concentrations (0, 0.1, 0.5,
1 or 2 mM) for 24 h. In order to determine the most suitable
concentration for BMSC proliferation to use in subsequent
experiments, BMSCs were pretreated with NAC (0, 0.1, 0.5, 1, 2 mM)
and incubated for 1 h prior to stimulation with LPS for 24 h. The
results are shown in Fig. 1B. An
NAC concentration of 1 mM had the strongest regulatory effect on
the proliferation rate of BMSCs compared with no treatment
(P<0.05 vs. control group); an increase in concentration from 1
to 2 mM NAC did not lead to a significant difference [LPS + NAC (1
mM) group vs. LPS + NAC (2 mM) group, P>0.05, P=0.957]. Thus, 1
mM NAC was selected for the subsequent assays.
NAC and Res decrease the levels of
inflammatory mediators in LPS-stimulated BMSCs
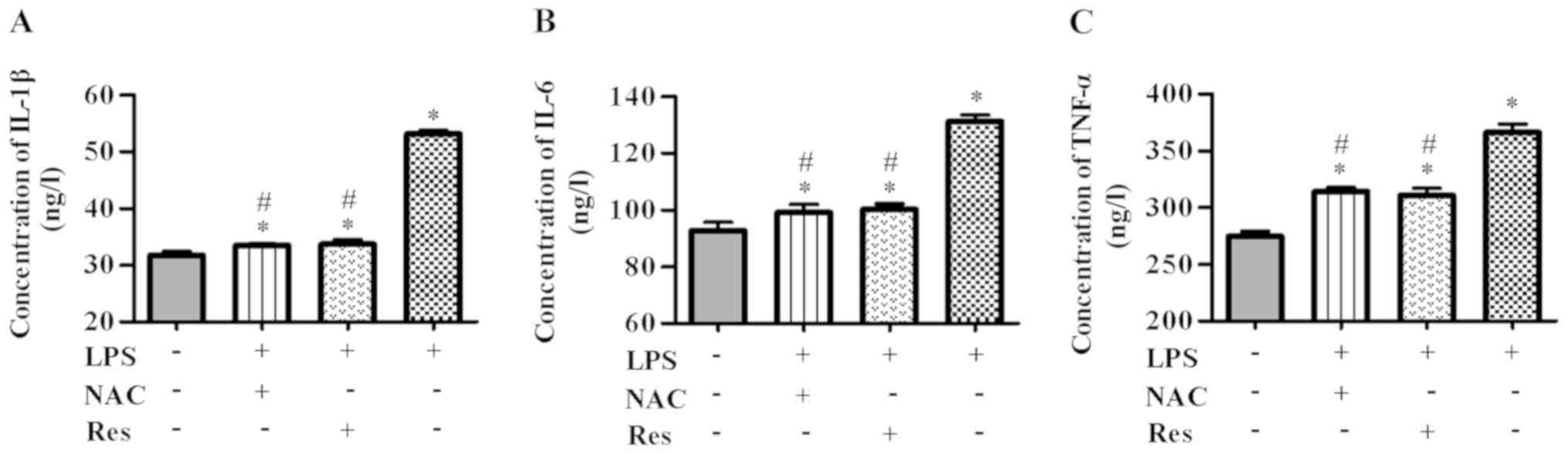
To investigate whether NAC could inhibit
inflammation in LPS-induced BMSCs, inflammatory mediators in
response to NAC or Res, which is a signaling pathway inhibitor,
were detected by ELISA kits. As shown in Fig. 2, expression of IL-1β, IL-6 and
TNF-α was higher in BMSCs in the LPS group compared with the
control group (P<0.05). At the same time, pretreatment with NAC
for 1 h led to a significant decrease in inflammatory mediators
(P<0.05). Similar results were observed in the Res + LPS and NAC
+ LPS groups (NAC + LPS vs. Res + LPS groups, P>0.05; IL-β,
P=0.980; IL-6, P=0.961; TNF-α, P=0.876). Together, the results
demonstrated that LPS could induce an inflammatory response,
whereas NAC or Res pretreatment clearly suppressed LPS-triggered
inflammation.
TXNIP/NLRP3 mediates the regulatory
effects of NAC on inflammation in LPS-treated BMSCs
To further identify the potential molecular
mechanism underlying the regulatory effects of NAC on LPS-mediated
BMSCs, RT-qPCR and western blotting were conducted to evaluate the
expression of mRNA and proteins, respectively, in LPS-treated
BMSCs. According to the RT-qPCR results (Fig. 3), compared with the other groups,
the LPS group (P<0.05) had the highest expression of ASC, NLRP3,
caspase-1 and TXNIP, but the lowest expression of TRX. However, the
effects of LPS on BMSCs were reversed with NAC pretreatment
(P<0.05). Specifically, NAC pretreatment downregulated the
expression of mRNAs increased by LPS and only upregulated the
expression of TRX. In addition, when BMSCs were pretreated with
Res, the same result as treatment with NAC was observed (NAC + LPS
vs. Res + LPS, P>0.05; NLRP3, P=0.892; ASC, P=1.000; caspase-1,
P=0.999; TXNIP, P=1.000; TRX, P=0.927). As shown in Fig. 4, the western blotting results were
consistent with the mRNA data (P<0.05 vs. LPS group) (NAC + LPS
vs. Res + LPS, P>0.05; NLRP3, P=0.394; ASC, P=0.998; caspase-1,
P=0.954; TXNIP, P=0.912; TRX, P=1.000) (control vs. NAC + LPS,
P>0.05; NLRP3, P=0.932; ASC, P=0.986; caspase-1, P=0.913; TXNIP,
P=1.000; TRX, P=0.945) (control vs. Res + LPS, P>0.05; NLRP3,
P=0.708; ASC, P=0.998; caspase-1, P=0.666; TXNIP, P=0.898; TRX,
P=0.922). Briefly, pretreatment with NAC or Res reduced the
expression of ASC, NLRP3, caspase-1 and TXNIP, but increased
expression of TRX compared with cells only treated with LPS.
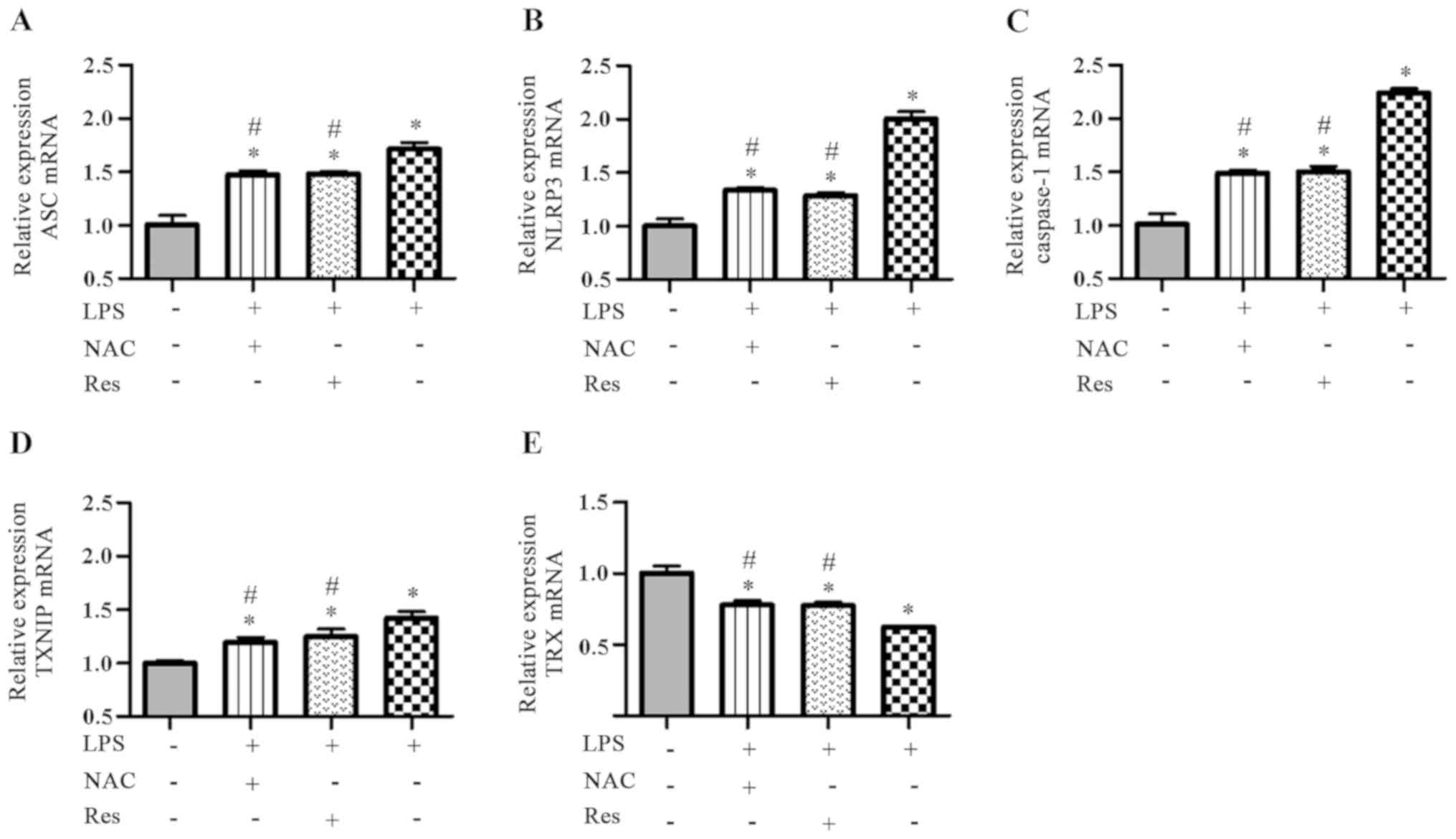 | Figure 3.Effects of NAC and Res on the mRNA
expression of ASC, NLRP3, caspase-1, TXNIP and TRX in
LPS-stimulated BMSCs. The levels were determined by reverse
transcription-quantitative PCR. The relative expression of (A) ASC,
(B) NLRP3, (C) caspase-1, (D) TXNIP and (E) TRX. *P<0.05 vs.
control group; #P<0.05 vs. LPS group. NAC, N-acetyl
cysteine; Res, resveratrol; ASC, apoptosis-associated speck-like
protein containing a CARD; NLRP3, nucleotide-binding
oligomerization domain-like receptor protein 3; TXNIP,
thioredoxin-interacting protein; TRX, thioredoxin; LPS,
lipopolysaccharide; BMSCs, bone marrow mesenchymal stem cells. |
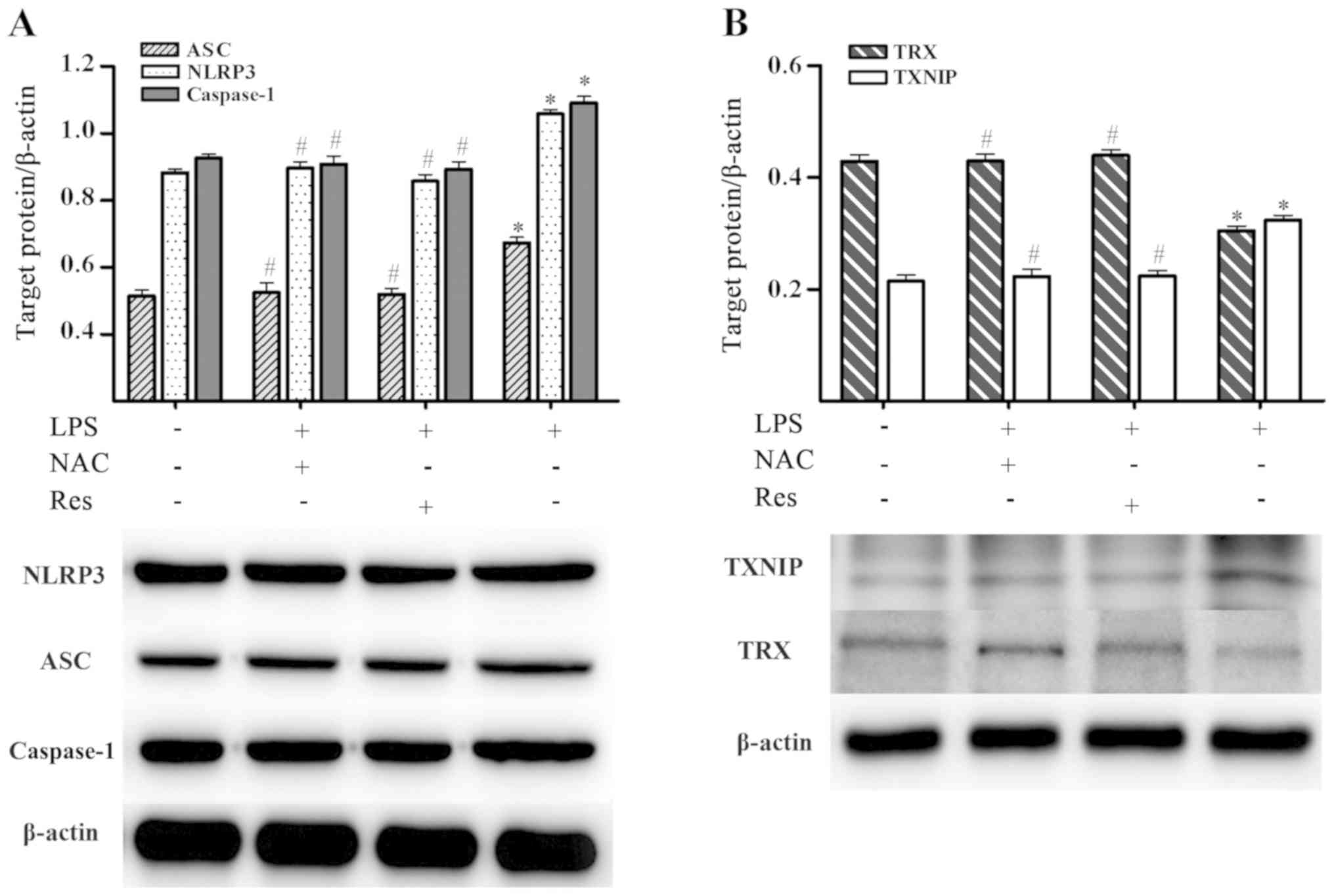 | Figure 4.Effects of NAC and Res on the protein
expression of ASC, NLRP3, caspase-1, TXNIP and TRX in
LPS-stimulated BMSCs. Levels of proteins were measured by western
blot analysis. (A) Protein expression of ASC, NLRP3 and caspase-1.
(B) Protein expression of TXNIP and TRX. *P<0.05 vs. control
group; #P<0.05 vs. LPS group. NAC, N-acetyl cysteine;
Res, resveratrol; ASC, apoptosis-associated speck-like protein
containing a CARD; NLRP3, nucleotide-binding oligomerization
domain-like receptor protein 3; TXNIP, thioredoxin-interacting
protein; TRX, thioredoxin; LPS, lipopolysaccharide; BMSCs, bone
marrow mesenchymal stem cells. |
Discussion
The results of the present study demonstrated that
NAC inhibited the LPS-induced inflammatory response in BMSCs by
significantly mediating the downregulation of IL-1β, IL-6 and
TNF-α, which were upregulated in response to LPS treatment. In
addition, it was identified that following treatment with LPS,
NLRP3, ASC, caspase-1 and TXNIP were upregulated, but TRX was
downregulated in BMSCs. It was also identified that NAC
downregulated the expression of ASC, NLRP3, caspase-1 and TXNIP,
but enhanced the expression of TRX in LPS-induced BMSCs, which was
similar to the effect of Res. Consequently, the results suggested
that LPS induced inflammatory responses in BMSCs, which could be
reversed by the anti-inflammatory activity of NAC via inhibition of
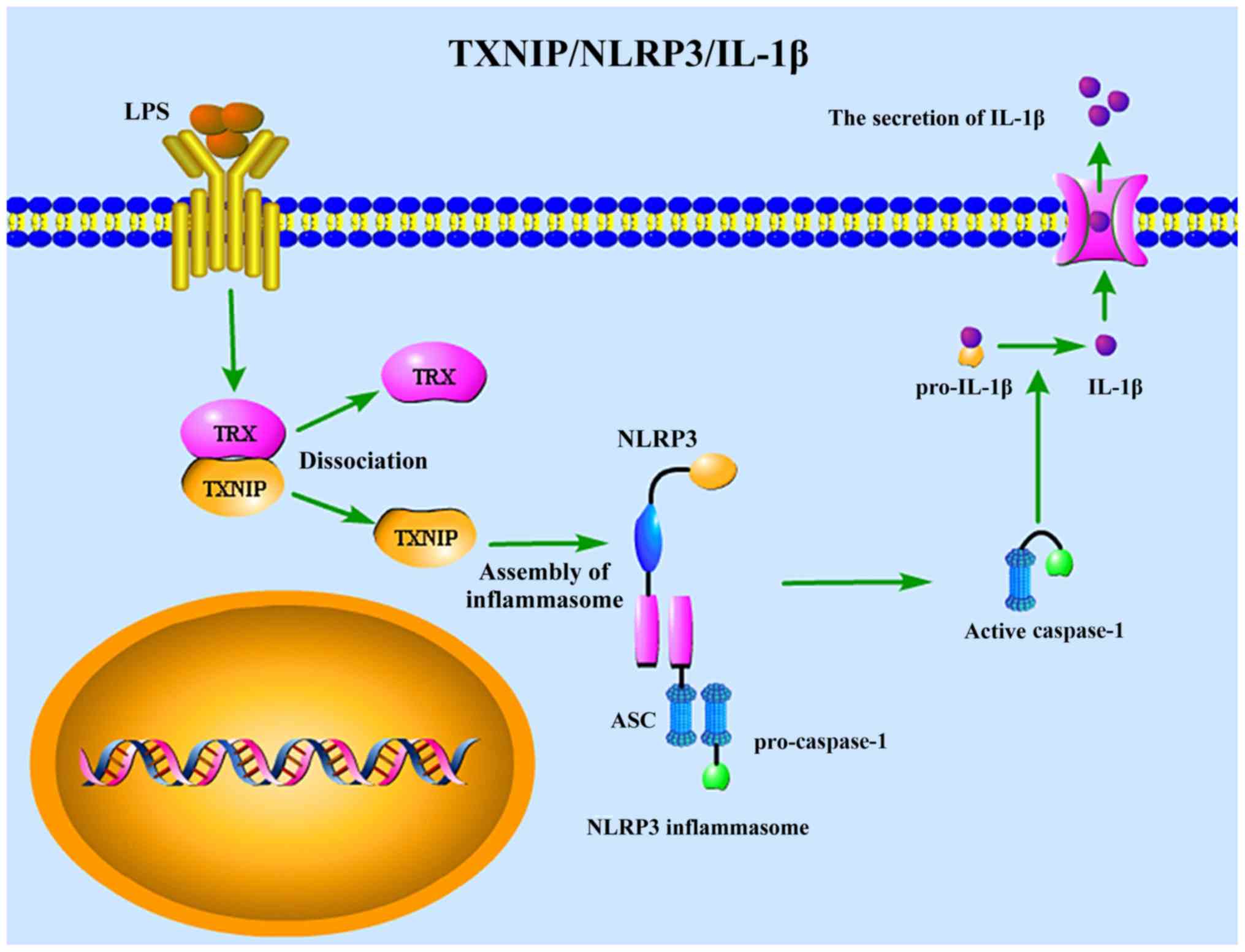
the TXNIP/NLRP3/IL-1β signaling pathway (Fig. 5).
Recently, BMSCs have attracted increasing attention,
due to their self-renewal ability and multidirectional
differentiation potential (28).
BMSCs are located in alveolar bone and are strongly associated with
bone formation and resorption (29). It has been confirmed that the
majority of osteoblasts colonizing the implant surface are
generated from BMSCs, contributing significantly to
osseointegration (30,31). Meanwhile, several studies have
reported that BMSCs can be effectively used for the treatment of
periodontitis (14,32,33).
However, the viability and osteogenic differentiation of BMSCs can
be downregulated in an inflammatory environment, which directly
results in treatment failure (34). Thereby, controlling inflammation is
essential for diseases associated with BMSCs.
A number of studies have demonstrated that
LPS-induced overproduction of IL-1β, IL-6 and TNF-α contributes to
the majority of inflammatory responses that are involved in the
pathogenesis of inflammatory-stimulated oral bone diseases,
including periodontitis (35),
peri-implantitis (36) and apical
periodontitis (37). By contrast,
our previous studies identified that NAC inhibited LPS-mediated
synthesis of IL-1β, IL-6 and TNF-α (10,26).
Thus the present study focused on the secretion of IL-1β, IL-6 and
TNF-α. It was demonstrated that in BMSCs, IL-1β, IL-6 and TNF-α
were overexpressed following administration of LPS, leading to
generation of inflammatory responses. Pretreatment with NAC reduced
the secretion of IL-1β, IL-6 and TNF-α, and inhibited the
inflammatory response. These findings revealed that NAC could be
used to manage the inflammatory response.
The NLRP3 inflammasome is critical for secretion of
the proinflammatory cytokine IL-1β (38). The present study demonstrated that
NAC can reduce the secretion of IL-1β. Subsequent experiments
determined whether NAC reduced IL-1β levels by inhibiting the NLRP3
inflammasome. The NLRP3 inflammasome, which contains NLRP3, ASC and
caspase-1 (39), can initiate
inflammation in response to infection of microbial products
(40,41). It is widely recognized that LPS
results in activation of the NLRP3 inflammasome (42–44).
Once activated, NLRP3 interacts with ASC, inducing the reciprocal
identification of NLRP3 and pro-caspase-1, thereby leading to
activation of caspase-1 (45,46).
Immediately, caspase-1, also termed inflammatory caspase, mediates
the maturation and secretion of IL-1β (47,48),
which serves a significant role in inflammation (49,50).
In the present study, protein and mRNA expression levels indicated
that LPS mediated overexpression of the NLRP3 inflammasome in
BMSCs. However, pretreatment with NAC attenuated the expression of
mRNA and target proteins. These results clearly demonstrated that
NAC downregulated the NLRP3 inflammasome and inhibited inflammatory
responses caused by LPS.
The specific molecular mechanism by which NAC acts
on the NLRP3 inflammasome and exhibits an anti-inflammatory effect
remains to be elucidated. TXNIP is an essential multi-functional
protein, which interferes with the function of TRX (51). The overexpression of TXNIP
activates inflammatory pathways, in part since TRX-mediated
inhibition of inflammation is reversed by TXNIP (51). Additionally, TXNIP directly
activates the NLRP3 inflammasome, further stimulating inflammation
(52–54). Previous studies have suggested
that, in response to stimulation, TXNIP dissociates from TRX and
then directly combines with a specific region of NLRP3 (55,56).
In addition, the incorporation of TXNIP and NLRP3 accelerates
inflammasome aggregation and oligomerization, inducing
transformation of caspase-1 and maturation of IL-1β (57). In the present study, LPS treatment
induced the overproduction of TXNIP and decreased expression of TRX
at the mRNA and protein levels. However, the expression of TXNIP
was downregulated and that of TRX was upregulated by pretreatment
with NAC. These observations further indicated that TXNIP and TRX
are involved in the anti-inflammatory effect of NAC on LPS-induced
BMSCs.
To clarify the underlying signaling pathway, Res was
used as a pathway blocker. Previous studies have indicated that
Res, a natural plant polyphenolic compound, can restrain NLRP3
inflammasome activation, which prevents the over-release of
inflammatory cytokines (58,59).
It was reported that Res can significantly inhibit the TXNIP/TRX
cascade, restrain the expression of TXNIP, inhibit the
incorporation of TXNIP and NLRP3 and finally block the inflammasome
aggregation and oligomerization (60). As expected, in the present study,
the effect of NAC was identified to be consistent with that of Res.
Therefore, it was concluded that the inflammatory regulatory
effects of NAC on LPS-induced BMSCs were closely associated with
the TXNIP/NLRP3/IL-1β pathway.
On one hand, the results of the present study
provided novel evidence that NAC can inhibit the inflammatory
response in BMSCs stimulated by LPS; on the other hand, it
demonstrated that one of the molecular mechanisms underlying the
inflammatory effects of NAC on LPS-induced BMSCs was closely
associated with the TXNIP/NLRP3/IL-1β pathway. These findings not
only broadened our view of how NAC affects LPS-induced BMSCs but
also provided an improved understanding of the underlying molecular
mechanism that may promote the development of anti-inflammation
strategies. However, in vivo experiments are required to
validate these findings so we plan to establish animal models to
further explore the anti-inflammatory effects of NAC in future
research.
Acknowledgements
Not applicable.
Funding
This study was funded by a grant from
Luzhou-Southwest Medical University (grant no. 2016LZXNYD-J21).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW and LG were primarily responsible for the
research protocol design. XW and MJ performed the experiments. XW,
XH, BZ and WP analyzed and interpreted the data. XW drafted the
manuscript. XW, LG and XH revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Berglundh T, Armitage G, Araujo MG,
Avila-Ortiz G, Blanco J, Camargo PM, Chen S, Cochran D, Derks J,
Figuero E, et al: Peri-implant diseases and conditions: Consensus
report of workgroup 4 of the 2017 world workshop on the
classification of periodontal and peri-implant diseases and
conditions. J Clin Periodontol. 45 (Suppl 20):3299–S291. 2018.
View Article : Google Scholar
|
|
2
|
Kinane DF, Stathopoulou PG and Papapanou
PN: Periodontal diseases. Nat Rev Dis Primers. 3:170382017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Eke PI, Borgnakke WS and Genco RJ: Recent
epidemiologic trends in periodontitis in the USA. Periodontol 2000.
82:257–267. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dreyer H, Grischke J, Tiede C, Eberhard J,
Schweitzer A, Toikkanen SE, Glöckner S, Krause G and Stiesch M:
Epidemiology and risk factors of peri-implantitis: A systematic
review. J Periodontal Res. 53:657–681. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eke PI, Thornton-Evans GO, Wei L,
Borgnakke WS, Dye BA and Genco RJ: Periodontitis in US adults:
National health and nutrition examination survey 2009–2014. J Am
Dent Assoc. 149:576–588.e6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Derks J, Schaller D, Hakansson J,
Wennstrom JL, Tomasi C and Berglundh T: Effectiveness of implant
therapy analyzed in a Swedish population: Prevalence of
peri-implantitis. J Dent Res. 95:43–49. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Monje A, Insua A and Wang HL:
Understanding peri-implantitis as a plaque-associated and
site-specific entity: On the local predisposing factors. J Clin
Med. 8:2792019. View Article : Google Scholar
|
|
8
|
Lamont RJ, Koo H and Hajishengallis G: The
oral microbiota: Dynamic communities and host interactions. Nat Rev
Microbiol. 16:745–759. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Singhrao SK and Olsen I: Assessing the
role of Porphyromonas gingivalis in periodontitis to
determine a causative relationship with Alzheimer's disease. J Oral
Microbiol. 11:15634052019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang L, Yang Y, Xiong X, Yu T, Wang X,
Meng W, Wang H, Luo G and Ge L: Oral lichen-planus-associated
fibroblasts acquire myofibroblast characteristics and secrete
pro-inflammatory cytokines in response to Porphyromonas
gingivalis lipopolysaccharide stimulation. BMC Oral Health.
18:1972018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo L, Zhang H, Li W, Zhan D and Wang M:
N-acetyl cysteine inhibits lipopolysaccharide-mediated induction of
interleukin-6 synthesis in MC3T3-E1 cells through the NF-κB
signaling pathway. Arch Oral Biol. 93:149–154. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang ZZ, Xiong T, Zheng R, Huang JL and
Guo L: N-acetyl cysteine protects HUVECs against
lipopolysaccharide-mediated inflammatory reaction by blocking the
NF-κB signaling pathway. Mol Med Rep. 20:4349–4357. 2019.PubMed/NCBI
|
|
13
|
Huang JL, Xiong T, Zhang ZZ, Tan YJ and
Guo L: Inhibition of the receptor for advanced glycation inhibits
lipopolysaccharide-mediated High mobility group protein B1 and
Interleukin-6 synthesis in human gingival fibroblasts through the
NF-κB signaling pathway. Arch Oral Biol. 105:81–87. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yu X, Quan J, Long W, Chen H, Wang R, Guo
J, Lin X and Mai S: LL-37 inhibits LPS-induced inflammation and
stimulates the osteogenic differentiation of BMSCs via P2X7
receptor and MAPK signaling pathway. Exp Cell Res. 372:178–187.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Čebatariūnienė A, Kriaučiūnaitė K,
Prunskaitė J, Tunaitis V and Pivoriūnas A: Extracellular vesicles
suppress basal and lipopolysaccharide-induced NFκB activity in
human periodontal ligament stem cells. Stem Cells Dev.
28:1037–1049. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lian D, Dai L, Xie Z, Zhou X, Liu X, Zhang
Y, Huang Y and Chen Y: Periodontal ligament fibroblasts migration
injury via ROS/TXNIP/Nlrp3 inflammasome pathway with
Porphyromonas gingivalis lipopolysaccharide. Mol Immunol.
103:209–219. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Qiu Z, He YH, Ming H, Lei SQ, Leng Y and
Xia ZY: Lipopolysaccharide (LPS) aggravates high glucose- and
hypoxia/reoxygenation-induced injury through activating
ROS-dependent NLRP3 inflammasome-mediated pyroptosis in H9C2
cardiomyocytes. J Diabetes Res. 2019:81518362019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang YS, Zhao YH, Zhong Y, Li XZ, Pu JX,
Luo YC and Zhou QL: Neferine inhibits LPS-ATP-induced endothelial
cell pyroptosis via regulation of ROS/NLRP3/caspase-1 signaling
pathway. Inflamm Res. 68:727–738. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Raudales JLM, Yoshimura A, Ziauddin SM,
Kaneko T, Ozaki Y, Ukai T, Miyazaki T, Latz E and Hara Y: Dental
calculus stimulates interleukin-1β secretion by activating NLRP3
inflammasome in human and mouse phagocytes. PLoS One.
11:e01628652016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shibata K: Historical aspects of studies
on roles of the inflammasome in the pathogenesis of periodontal
diseases. Mol Oral Microbiol. 33:203–211. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li P, Allen H, Banerjee S, Franklin S,
Herzog L, Johnston C, McDowell J, Paskind M, Rodman L, Salfeld J,
et al: Mice deficient in IL-l beta-converting enzyme are defective
in production of mature IL-l beta and resistant to endotoxic shock.
Cell. 80:401–411. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Neuwelt EA, Pagel MA, Hasler BP,
Deloughery TG and Muldoon LL: Therapeutic efficacy of aortic
administration of N-acetylcysteine as a chemoprotectant against
bone marrow toxicity after intracarotid administration of
alkylators, with or without glutathione depletion in a rat model.
Cancer Res. 61:7868–7874. 2001.PubMed/NCBI
|
|
23
|
Rosic G, Selakovic D, Joksimovic J,
Srejovic I, Zivkovic V, Tatalović N, Orescanin-Dusic Z, Mitrovic S,
Ilic M and Jakovljevic V: The effects of N-acetylcysteine on
cisplatin-induced changes of cardiodynamic parameters within
coronary autoregulation range in isolated rat hearts. Toxicol Lett.
242:34–46. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kelly GS: Clinical applications of
N-acetylcysteine. Altern Med Rev. 3:114–127. 1998.PubMed/NCBI
|
|
25
|
Shih WL, Chang CD, Chen HT and Fan KK:
Antioxidant activity and leukemia initiation prevention in vitro
and in vivo by N-acetyl-L-cysteine. Oncol Lett. 16:2046–2052.
2018.PubMed/NCBI
|
|
26
|
Zheng R, Tan YJ, Gu MQ, Kang T, Zhang H
and Guo L: N-acetyl cysteine inhibits lipopolysaccharide-mediated
synthesis of interleukin-1β and tumor necrosis factor-α in human
periodontal ligament fibroblast cells through nuclear factor-kappa
B signaling. Medicine (Baltimore). 98:e171262019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang C, Geng J and Jiang S: MicroRNAs in
regulation of osteogenic differentiation of mesenchymal stem cells.
Cell Tissue Res. 368:229–238. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Arvidson K, Abdallah BM, Applegate LA,
Baldini N, Cenni E, Gomez-Barrena E, Granchi D, Kassem M, Konttinen
YT, Mustafa K, et al: Bone regeneration and stem cells. J Cell Mol
Med. 15:718–746. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu D, Li K, Xie Y, Pan H, Zhao J, Huang L
and Zheng X: The combined effects of nanotopography and Sr ion for
enhanced osteogenic activity of bone marrow mesenchymal stem cells
(BMSCs). J Biomater Appl. 31:1135–1147. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Heng BC, Cao T, Stanton LW, Robson P and
Olsen B: Strategies for directing the differentiation of stem cells
into the osteogenic lineage in vitro. J Bone Miner Res.
19:1379–1394. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu L, Liu Y, Zhang X and Lin J: The
therapeutic role of bone marrow stem cell local injection in rat
experimental periodontitis. J Oral Rehabil. Jun 20–2019.(Epub ahead
of print). doi: 10.1111/joor.12843. View Article : Google Scholar
|
|
33
|
Han N, Zhang F, Li G, Zhang X, Lin X, Yang
H, Wang L, Cao Y, Du J and Fan Z: Local application of IGFBP5
protein enhanced periodontal tissue regeneration via increasing the
migration, cell proliferation and osteo/dentinogenic
differentiation of mesenchymal stem cells in an inflammatory niche.
Stem Cell Res Ther. 8:2102017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu L, Zhang G, Guo C, Zhao X, Shen D and
Yang N: MiR-128-3p mediates TNF-α-induced inflammatory responses by
regulating Sirt1 expression in bone marrow mesenchymal stem cells.
Biochem Biophys Res Commun. 521:98–105. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang L and Deng S: Effects of
astragaloside IV on inflammation and immunity in rats with
experimental periodontitis. Braz Oral Res. 33:e0322019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li H, Chen Z, Zhong X, Li J and Li W:
Mangiferin alleviates experimental peri-implantitis via suppressing
interleukin-6 production and toll-like receptor 2 signaling
pathway. J Orthop Surg Res. 14:3252019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Cosme-Silva L, Dal-Fabbro R, Cintra LTA,
dos Santos VR, Duque C, Ervolino E, Bomfim SM and Gomes-Filho JE:
Systemic administration of probiotics reduces the severity of
apical periodontitis. Int Endod J. 52:1738–1749. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
He Y, Hara H and Núñez G: Mechanism and
regulation of NLRP3 inflammasome activation. Trends Biochem Sci.
41:1012–1021. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Guo H, Callaway JB and Ting JP:
Inflammasomes: Mechanism of action, role in disease, and
therapeutics. Nat Med. 21:677–687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shin S and Brodsky IE: The inflammasome:
Learning from bacterial evasion strategies. Semin Immunol.
27:102–110. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Place DE and Kanneganti TD: Recent
advances in inflammasome biology. Curr Opin Immunol. 50:32–38.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Mangan MSJ, Olhava EJ, Roush WR, Seidel
HM, Glick GD and Latz E: Targeting the NLRP3 inflammasome in
inflammatory diseases. Nat Rev Drug Discov. 17:588–606. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Baker PJ, Boucher D, Bierschenk D, Tebartz
C, Whitney PG, D'Silva DB, Tanzer MC, Monteleone M, Robertson AAB,
Cooper MA, et al: NLRP3 inflammasome activation downstream of
cytoplasmic LPS recognition by both caspase-4 and caspase-5. Eur J
Immunol. 45:2918–2926. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Feng H, Gu J, Gou F, Huang W, Gao C, Chen
G, Long Y, Zhou X, Yang M, Liu S, et al: High glucose and
lipopolysaccharide prime NLRP3 inflammasome via ROS/TXNIP pathway
in mesangial cells. J Diabetes Res. 2016:69731752016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Xiaoyu H, Si H, Li S, Wang W, Guo J, Li Y,
Cao Y, Fu Y and Zhang N: Induction of heme oxygenas-1 attenuates
NLRP3 inflammasome activation in lipopolysaccharide-induced
mastitis in mice. Int Immunopharmacol. 52:185–190. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bolívar BE, Vogel TP and Bouchier-Hayes L:
Inflammatory caspase regulation: Maintaining balance between
inflammation and cell death in health and disease. FEBS J.
286:2628–2644. 2019.PubMed/NCBI
|
|
47
|
Martinon F, Burns K and Tschopp Jr: The
inflammasome: A molecular platform triggering activation of
inflammatory caspases and processing of proIL-beta. Mol Cell.
10:417–426. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lu WL, Song DZ, Yue JL, Wang TT, Zhou XD,
Zhang P, Zhang L and Huang DM: NLRP3 inflammasome may regulate
inflammatory response of human periodontal ligament fibroblasts in
an apoptosis-associated speck-like protein containing a CARD
(ASC)-dependent manner. Int Endod J. 50:967–975. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Dinarello CA: A clinical perspective of
IL-1β as the gatekeeper of inflammation. Eur J Immunol.
41:1203–1217. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mantovani A, Dinarello CA, Molgora M and
Garlanda C: Interleukin-1 and related cytokines in the regulation
of inflammation and immunity. Immunity. 50:778–795. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Devi TS, Lee I, Hüttemann M, Kumar A,
Nantwi KD and Singh LP: TXNIP links innate host defense mechanisms
to oxidative stress and inflammation in retinal Muller glia under
chronic hyperglycemia: Implications for diabetic retinopathy. Exp
Diabetes Res. 2012:1–19. 2012. View Article : Google Scholar
|
|
52
|
Wang CY, Xu Y, Wang X, Guo C, Wang T and
Wang ZY: Dl-3-n-Butylphthalide inhibits NLRP3 inflammasome and
mitigates Alzheimer's-like pathology via Nrf2-TXNIP-TrX axis.
Antioxid Redox Signal. 30:1411–1431. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hou Y, Wang Y, He Q, Li L, Xie H, Zhao Y
and Zhao J: Nrf2 inhibits NLRP3 inflammasome activation through
regulating Trx1/TXNIP complex in cerebral ischemia reperfusion
injury. Behav Brain Res. 336:32–39. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Chen W, Zhao MJ, Zhao SZ, Lu QY, Ni LS,
Zou C, Lu L, Xu X, Guan HJ, Zheng Z and Qiu QH: Activation of the
TXNIP/NLRP3 inflammasome pathway contributes to inflammation in
diabetic retinopathy: A novel inhibitory effect of minocycline.
Inflamm Res. 66:157–166. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Liu Y, Lian K, Zhang L, Wang R, Yi F, Gao
C, Xin C, Zhu D, Li Y, Yan W, et al: TXNIP mediates NLRP3
inflammasome activation in cardiac microvascular endothelial cells
as a novel mechanism in myocardial ischemia/reperfusion injury.
Basic Res Cardiol. 109:4152014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang X, Zhang JH, Chen XY, Hu QH, Wang
MX, Jin R, Zhang QY, Wang W, Wang R, Kang LL, et al: Reactive
oxygen species-induced TXNIP drives fructose-mediated hepatic
inflammation and lipid accumulation through NLRP3 inflammasome
activation. Antioxid Redox Signal. 22:848–870. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ma Z and Damania B: Editorial: NLRP3:
Immune activator or modulator? J Leukoc Biol. 99:641–643. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Jiang L, Zhang L, Kang K, Fei D, Gong R,
Cao Y, Pan S and Zhao M and Zhao M: Resveratrol ameliorates
LPS-induced acute lung injury via NLRP3 inflammasome modulation.
Biomed Pharmacother. 84:130–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chang YP, Ka SM, Hsu WH, Chen A, Chao LK,
Lin CC, Hsieh CC, Chen MC, Chiu HW, Ho CL, et al: Resveratrol
inhibits NLRP3 inflammasome activation by preserving mitochondrial
integrity and augmenting autophagy. J Cell Physiol. 230:1567–1579.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Feng L and Zhang L: Resveratrol suppresses
Aβ-induced microglial activation through the TXNIP/TRX/NLRP3
signaling pathway. DNA Cell Biol. 38:874–879. 2019. View Article : Google Scholar : PubMed/NCBI
|