Introduction
Neonatal hypoxic-ischemic brain damage (HIBD) is a
common clinical syndrome in newborns caused by anoxia and reduced
cerebral blood flow or temporary severance during the perinatal
period (1–3). It has been reported that 2–6/1,000
newborns experience HIBD (4), and
~25% of infants who have HIBD develop neurologic sequela (5,6).
Hypothermia is the only admitted therapy for treatment of neonatal
HIBD in the clinic; however, the therapeutic window of hypothermia
is confined to 6 h after birth, and even then, >40% of the
infants either died or survive with various impairments following
hypothermic treatment (7–9). Thus, alternative effective and safe
therapies for treatment of HIBD are required.
Globins are hemoproteins that bind O2 and
serve an important role in the animal's respiration and oxidative
energy production (10,11). However, globins may also possess
other functions, such as the decomposition of nitric oxide (NO),
the detoxification of reactive oxygen species (ROS) or
intracellular signaling (12).
Cytoglobin (CYGB), the fourth member of the vertebrate globin
family of hemoproteins, is ubiquitously expressed in various
tissues and organs, including the liver, kidney, brain and retina
(13). CYGB may serve a
cytoprotective role under hypoxic and/or ischemic conditions
(14–17). Previous studies have also reported
that pretreatment of CYGB-overexpression reduces HI injury and
improves long term memory and athletic ability following neonatal
HIBD (18). However, a safe and
effective method of upregulating CYGB expression in animals remains
a challenge that requires further investigation.
Previous studies have revealed that several types of
cells may serve important roles in relieving HIBD (19–21).
Preclinical trials on stem cells in cerebral palsy have been
conducted and have reported significant improvements in acute
hypoxic injury animal models (22). Mesenchymal stem cells (MSCs) are
well-known for their ‘immunosuppressive’ properties, and thus may
be important candidates for allogeneic cell therapy (23). In recent years, human umbilical
cord-derived MSCs (HuMSCs) have become an alternate source of MSCs.
Moreover, studies have observed that transplantation of HuMSCs at
an early stage following HIBD can reduce HI injury in rats and
decrease gliosis (24,25). Stem cells can also act as a gene
transporter in gene therapy (26).
The use of stem cells as transgenic strategies does not require
integration of the therapeutic DNA into the chromosomes of the
patient's cells, and instead, the transferred DNA is stabilized
extrachromosomally (27). However,
it has been identified that there are major risks associated with
the use of integrating vectors, including retroviral vectors,
arising from their potential for insertional mutagenesis, in which
the vector inserts into the DNA of a cell and disrupts a functional
element of that DNA (27); it has
been reported that stem cell transgenic therapy can reduce this
risk (28).
The aim of the present study was to investigate
whether nasal transplantation of CYGB genetically modified HuMSCs
(CYGB-HuMSCs) exhibited higher protective effects in neonatal rats
from HIBD compared with rats treated without CYGB genetically
modified HuMSCs. Additionally, the potential underlying mechanism
was examined.
Materials and methods
Preparation of HuMSCs
Ethical approval was obtained from the Institutional
Review Board of Maternal and Child Health Care Hospital of Shenzhen
University (approval no. SZPSFY2017-06). HuMSCs were prepared as
previously described (29). With
the written consent of the patients, human umbilical cords from 5
patients (age range, 23–34 years old) who underwent full-term
caesarian sections in the Maternal and Child Health Care Hospital
(Pingshan, Shenzhen, China) from February 2018 to March 2018 were
collected immediately into sterilized 50 ml tubes. After washing
with PBS for 30 sec, the human umbilical cord was cut into 2–3 cm
thick sections. The umbilical arteries and veins were removed, and
the remaining tissue, the Wharton's jelly, was dissected into
smaller fragments and transferred to 75 cm2 flasks
containing DMEM/F12 media (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.),
100 µg/ml penicillin/streptomycin (Sangon Biotech Co., Ltd.), 1
g/ml amphotericin B (Gilead Sciences, Inc.), 5 ng/ml epidermal
growth factor (PeproTech, Inc.) and 5 ng/ml basic fibroblast growth
factor (PeproTech, Inc.). Cultures remained undisturbed for 5–7
days at 37°C with 95% air/5% CO2 in a humidified
incubator to allow migration of cells from the explants.
Subsequently, the media was replaced. After three passages, cells
were harvested. Cells were observed under a Zeiss Axio Imager Z1
light inverted microscope (magnification, ×100; Carl Zeiss AG).
Adenovirus-mediated transfection
Adenovirus plasmid carrying CYGB (Ad-CYGB) was
purchased from Guangzhou Forevergen Technology Co., Ltd. Adenovirus
plasmid carrying only enhanced green fluorescent protein (eGFP) was
used as the transfection control. The transfection was performed
using Lipofectamine® 2000 reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). HuMSCs were seeded into 12-well plates at
a concentration of 2×105 cells per well. After culture
for 12 h, Ad-CYGB (2 µl; multiplicity of infection, 10) were added
to the respective wells; the same volume of control adenovirus was
added. On the second day after infection, the virus-containing
medium was aspirated and replaced with fresh complete medium to
continue the culture. The culture medium was replaced every 2 days
thereafter for a total period of 7 days. Follow-up experiments were
performed 48 h after the transfection. Cells were observed under a
Zeiss Axio Imager Z1 fluorescent inverted microscope
(magnification, ×200; Carl Zeiss AG).
Experimental animals
Sprague-Dawley rats (age, 7 days; 64 females; 56
males; weight, 12.5-16.3 g) were obtained from the Experimental
Animal Center of Shantou University Medical College. The present
study was approved by the Institutional Animal Care and Use
Committee of Shenzhen University (approval no. 2017-06) and
strictly adhered to the ARRIVE guidelines (30). All animals were maintained in a
temperature- and humidity-controlled room (temperature, 22–24°C;
humidity, 40–70%) with a 12 h light/dark cycle, and free access to
food and water. Throughout the experiment, all animals were treated
in accordance with the Guide for the Care and Use of Laboratory
Animals in Shenzhen University, and all procedures conformed to
internationally accredited guidelines and ethical regulations on
animal research. Surgery was performed under isoflurane anesthesia,
and all efforts were made to reduce the total number of animals
used and minimize their potential suffering.
Animal groups
A total of 120 Sprague-Dawley rats were used in the
present experiment. The rats were divided into four groups, with 30
rats in each group. Sham group animals received only anesthesia and
exposure of the left common carotid artery, but no ligation and
hypoxia operation. Animals in the HIBD, HuMSCs and CYGB-HuMSCs
groups received ligation of the left common carotid artery combined
with hypoxia. A total of 30 min after HIBD model preparation,
animals in the HuMSCs group and CYGB-HuMSCs group received
intranasal administration of 1×106 HuMSCs or CYGB-HuMSCs
(3 µl), respectively. Animals in the HIBD group received only PBS
(3 µl) via intranasal administration.
Establishment of an animal model of
HIBD and transplantation of HuMSCs
The HIBD neonatal rat model was based on the
classical Rice-Vannucci model (31). The experimental rat was deeply
anesthetized via inhalation of isoflurane (3% to effect). Then, the
left common carotid artery was exposed, followed by double-ligation
with 5-0 silk sutures and the artery between the ligations was cut
off. The wound was then sutured. The total time for surgery in each
animal was ≤3 min. The body temperature of the animal was
maintained at 37°C with a radiant warmer table. After surgery,
animals were allowed 1–2 h to recover from anesthesia with their
mother. After recovery, animals were placed in a container with a
lowered oxygen percentage (8% oxygen balanced with 92% nitrogen)
and placed in a 37°C water bath for 2.5 h to induce systemic
hypoxia. Animals in the sham group only received anesthesia and
exposure of the left common carotid artery. A total of 30 min after
HI, 1×106 cells HuMSCs or CYGB-HuMSCs (3 µl) were
intranasally administered, respectively. As described previously
(32), prior to the administration
of HuMSCs, to increase the permeability of the nasal mucosa,
nostrils were treated with 3 µl hyaluronidase (100 U;
Sigma-Aldrich; Merck KGaA) in PBS. A total of 30 min later, animals
were administered 3 µl HuMSCs, CYGB-HuMSCs (1×106 cells)
or PBS (3 µl) twice in each nostril.
Observation
After modeling for 3, 14 and 29 days, five randomly
selected rats were sacrificed via inhalation anesthesia with
isoflurane (3% to effect) followed by decapitation in each group.
Then, reverse transcription-quantitative (RT-q)PCR and western
blotting were used to detect CYGB expression in brain tissues.
Additionally, ERK, phosphorylated-(p-)ERK, JNK, p-JNK, p38 and
p-p38 protein expression levels were assessed using western
blotting. Nissl staining results of the cortex and hippocampal
Cornu Ammonis 1 (CA1) area of rats in each group were compared
after modeling for 3 days. TUNEL assays and immunofluorescence were
performed on day 3 after modeling; immunofluorescence was also
performed on day 14. Moreover, a Morris-Water maze experiment was
used 29 days after modeling.
Nissl staining
The obtained brain specimens were fixed in 4% (w/v)
formaldehyde at room temperature for 36 h, paraffin-embedded and
sliced into 3-µm thick sections. The paraffin sections were
deparaffinized in xylene and rehydrated in a descending ethanol
series at room temperature, then, the sections were immersed in 1%
toluidine blue stain at 37°C. After 5 min, the sections were washed
with water, differentiated with 1% glacial acetic acid and placed
in xylene for 5 min at room temperature. Then the slices were
removed from the xylene, dried and sealed with neutral gum. Samples
were observed under a light microscope (Olympus BX43; Olympus
Corporation) at ×200 magnification.
TUNEL staining
TUNEL staining was performed on paraffin sections
using an in-situ cell death detection kit (DeadEnd™
Fluorometric TUNEL system; Promega Corporation), according to the
manufacturer's protocol. Paraffin sections were prepared and fixed
as previously described for Nissl staining. Paraffin sections were
dewaxed in xylene and rehydrated in a descending ethanol series at
room temperature, then treated with 20 mg/l DNase proteinase K at
20–37°C for 15 min. Subsequently, the sections were washed with PBS
three times. Sections were then covered with 100 µl equilibration
buffer and incubated at room temperature for 5–10 min. After
removing the balance solution, 50 µl rTdT incubation buffer was
added and the sections covered with plastic coverslips to incubate
at 37°C for 60 min. Sections were then immersed in 2X saline sodium
citrate in a dyeing tank at room temperature for 15 min. To stop
the reactions, sections were washed twice with PBS. Subsequently,
sections were stained with 1X Hoechst nuclear stain at 37°C for 15
min and mounted with neutral gum. Samples were observed under a
Zeiss Axio Imager Z1 fluorescent microscope (magnification, ×200;
Carl Zeiss AG) in three randomly selected fields of view.
Image-Pro-Plus software (version 6.0; Media Cybernetics, Inc.) was
used to perform the semi-quantitative analysis of the apoptotic
cells.
Immunofluorescence
Paraffin sections were prepared and fixed as
previously described for Nissl staining. The paraffin sections were
deparaffinized in xylene and rehydrated using a descending ethanol
series at room temperature. Then, the sections were permeabilized
with 0.1% Triton X-100 at room temperature for 10 min. Sections
were blocked with normal goat blocking serum (Abbkine Scientific
Co., Ltd.) at a volume fraction of 3% at room temperature for 1 h.
After washing three times with PBS for 2 min/time, sections were
treated overnight with primary antibody at 4°C. The primary
antibodies used were glial fibrillary acidic protein (GFAP; Cell
Signaling Technology, Inc.; cat. no. 80788S; 1:50) and
neuron-specific enolase (NSE; ProteinTech Group, Inc.; cat. no.
10149-1-AP; 1:50). The sections were rinsed twice with PBS and then
incubated with the secondary Alexa Fluor 555 antibody (Abcam; cat.
no. ab150074; 1:200) for 1 h at room temperature in the dark.
Samples were observed under a Zeiss Axio Imager Z1 fluorescent
microscope (magnification, ×200; Carl Zeiss AG).
RT-qPCR
After HI modeling for 3, 14 or 29 days, rats from
different groups were anesthetized and decapitated. Samples of
injured brain tissues were collected rapidly and total RNA was
prepared from samples collected using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) as previously
described (33). Total RNA (2 µg)
was reverse transcribed using M-MLV reverse transcriptase (Promega
Corporation). In a sterile RNase-free microcentrifuge tube, 2 µg
total RNA and 1 µg primer in a total volume of 15 µl in water were
added. The tube was heated tube to 70°C for 5 min. Then, the tube
was cooled immediately on ice and centrifuged (500 × g; 4°C; 1 min)
briefly to collect the solution at the bottom of the tube. In
total, 5 µl 5X buffer, 6.25 µl 2 mM dNTP, 1 µl M-MLV and 12.75 µl
RNase-free water were added to the tube, which was mix gently and
incubated for 60 min at 42°C. cDNA expression levels were
determined via qPCR using specific primers and GoTaq®
qPCR Master mix (Promega Corporation) in a StepOne Plus amplifier
(Bio-Rad Laboratories, Inc.). The thermocycling protocol was as
follows: Initial denaturation at 95°C for 120 sec; followed by 40
cycles at 95°C for 15 sec, 60°C for 30 sec and 72°C for 45 sec, and
then a final elongation at 72°C for 10 min. Data were quantified
using the 2−ΔΔCq method (34). The primers used were as follows:
CYGB (NM_134268.5; product length, 152 bp) forward,
5′-TTGCCAGTGACTTCCCACC-3′ and reverse 5′-CCCGAAGAGGGCAGTGTG-3′; and
GAPDH (NM_001289745.3; product length, 185 bp) forward,
5′-GAGTCAACGGATTTGGTCG-3′ and reverse
5′-GAGTCAACGGATTTGGTCGT-3′.
Western blotting
Brain tissues collected simultaneously for the qPCR
experiments were homogenized in ice-cold RIPA lysis buffer
(Beyotime Institute of Biotechnology), and the protein
concentrations were measured using a bicinchoninic acid protein
assay kit (Beyotime Institute of Biotechnology). A total of 40 µg
total protein per lane was loaded on a 12% SDS-gel, resolved using
SDS-PAGE and transferred to a PVDF membrane (EMD Millipore). The
membrane was blocked with TBS-Tween (20 mM Tris; pH 7.6; 135 mM
NaCl; 0.05% Tween) containing 5% non-fat dry milk overnight at 4°C,
washed with 1X TBS-0.1% Tween-20 and then incubated at 4°C with
primary antibodies in blocking solution. The following primary
antibodies were used: GAPDH (1:10,000; cat. no. 10494-1-AP;
ProteinTech Group, Inc.), CYGB (1:1,000; cat. no. 13317-1-AP;
ProteinTech Group, Inc.), p-p38 (1:1,000; cat. no. 4511S; Cell
Signaling Technology, Inc.), p38 (1:1,000; cat. no. 8690S; Cell
Signaling Technology, Inc.), p-ERK (1:1,000; cat. no. 4370; Cell
Signaling Technology, Inc.), ERK (1:1,000; cat. no. 9102; Cell
Signaling Technology, Inc.), p-JNK (1:1,000; cat. no. 4668; Cell
Signaling Technology, Inc.) and JNK (1:1,000; cat. no. 9252; Cell
Signaling Technology, Inc.). After washing the membrane four times
with TBS-Tween, the horseradish peroxidase-labeled secondary goat
anti-rabbit IgG antibody (1:8,000; cat. no. RS0002; Immunoway;
Suzhou Ruiying Biotechnology Co., Ltd.), was added and incubated
for 2 h at room temperature, and then washed again. Protein bands
were visualized using an ECL Plus chemiluminescence kit (Guangzhou
Forevergen Technology Co., Ltd.). ImageJ software version 1.52 h
(National Institutes of Health) was used to perform the
densitometric analysis.
Morris-Water maze test
The water maze was composed of a cylindrical pool
and a platform. The height of the pool was 70 cm and the diameter
was 80 cm. The diameter of the platform was 8 cm. Over the pool,
there was a digital camera connected to the computer. The pool was
filled with water until the surface of the water was ~0.5 cm above
the surface of the platform, the water temperature was controlled
at 22.0±0.5°C. A specific point on the pool was used as the point
of entry for rats. The platform was placed in the third quadrant,
and the position of the platform was unchanged throughout the
entire experimental process. Training twice a day started 4 days
prior to the formal experiment; rats were placed in the water from
the four quadrants. If the animal found the platform within 120
sec, it was left on the platform for 20 sec. If the animal did not
find the platform, it was placed on the platform and left for 20
sec. On the second day after the training finished, experiments
were performed to record the time and the path length (PL) from the
point of entry to the platform in 120 sec.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism version 8 (GraphPad Software, Inc.). Continuous experimental
data are presented as the mean ± standard deviation of ≥3
experimental repeats. A one-way ANOVA followed by Tukey's of
Bonferroni post hoc test was used to compare the differences
between multiple groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
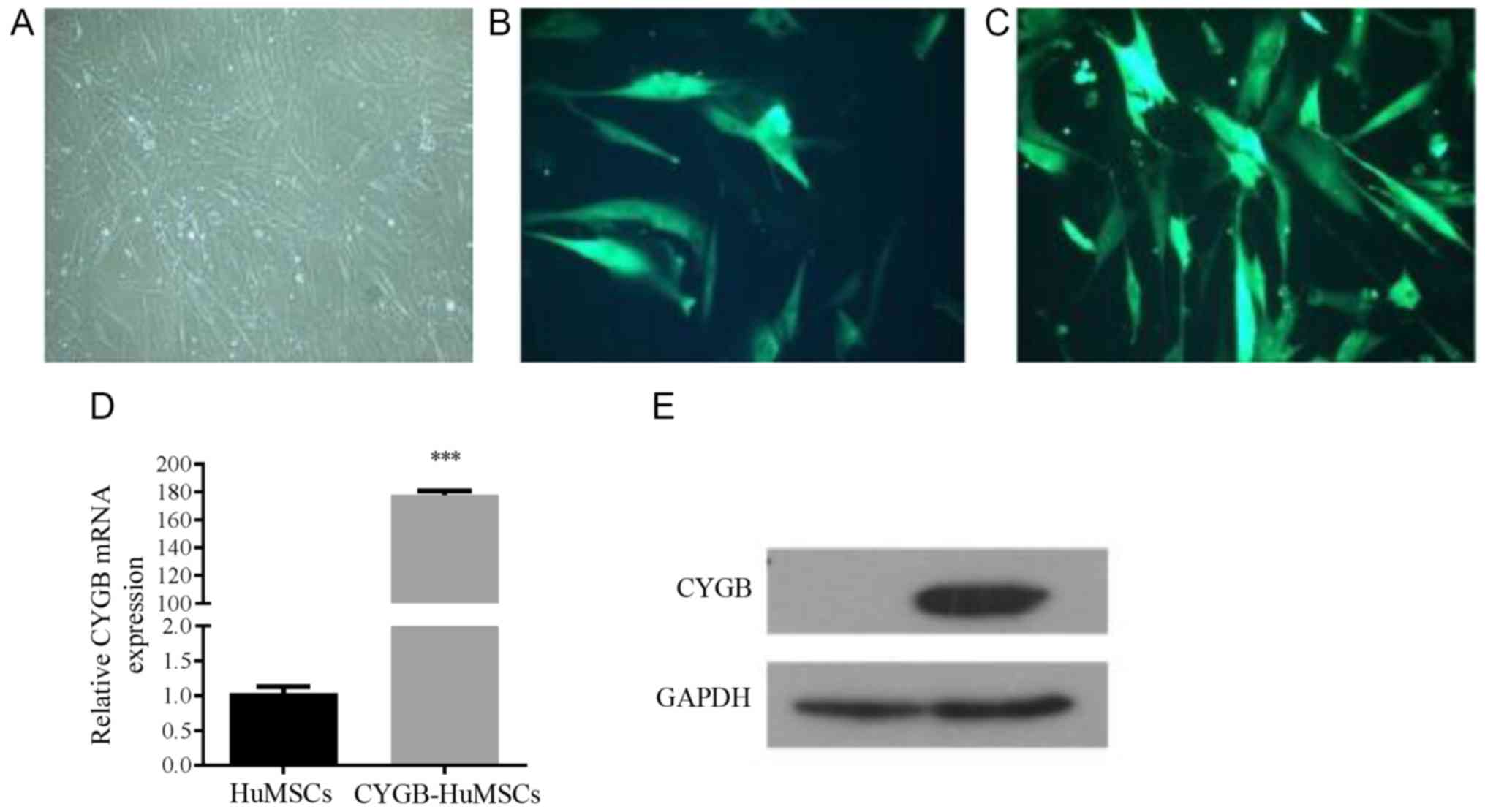
Morphology of cultured HuMSCs and
transfection
After 5–7 days of culture, fibroblast-like cells
migrated out from the surrounding tissues. Primary HuMSCs were
passaged when they were cultured for 10–14 days and the cells
reached 80% confluence. In the 3rd generation, cells had a stable
fibroblast-like morphology (Fig.
1A). The results of eGFP immunofluorescence analysis
demonstrated that the adenovirus plasmid transfection efficiency
was ~80% (Fig. 1B and C).
Furthermore, RT-qPCR results suggested that the transfected CYGB
gene was efficiently expressed in HuMSCs (Fig. 1D and E).
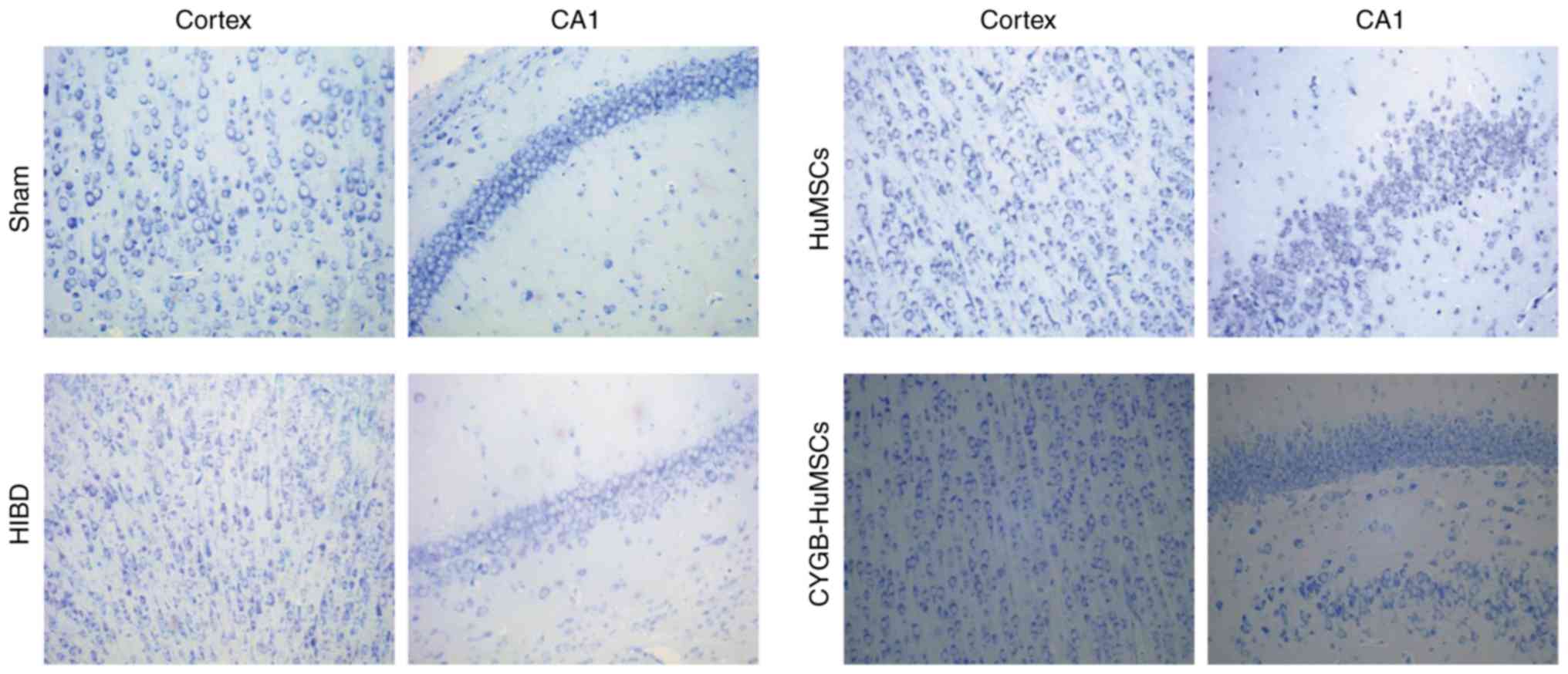
Observation of histological changes in
the brain using Nissl staining after modeling for 3 days
Neuronal cell loss in both the cerebral cortex and
hippocampus was observed via Nissl staining after modeling for 3
days. HI markedly reduced the number of cells and resulted in
irregularly arranged and smaller neurons in the brain 3 days after
HI compared with the sham group (Fig.
2). However, in the HuMSCs group, nerve cells were regularly
arranged and survival numbers were notably increased compared with
the HI group (Fig. 2).
Furthermore, high numbers of Nissl-stained cells were observed in
the CYGB-HuMSCs group compared with the HuMSCs group (Fig. 2).
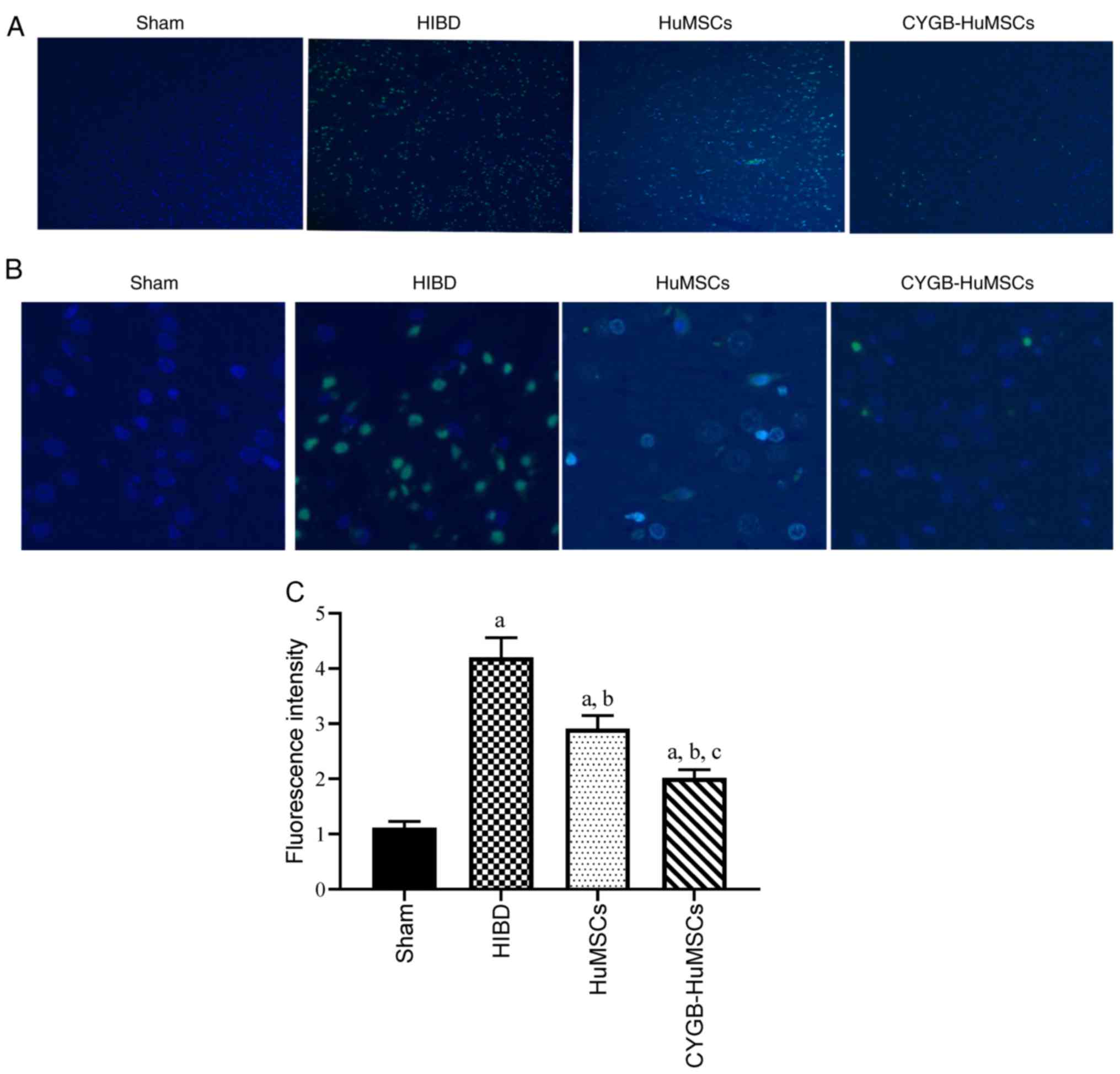
TUNEL staining
To determine whether HuMSCs and CYGB-HuMSCs
transplantation prevented apoptosis at the acute stage of neonatal
HI injury, TUNEL staining of tissue sections was performed after 3
days of modelling (Fig. 3A and B).
Significantly fewer apoptotic cells were observed in the HuMSCs
group compared with the HI groups (P<0.01), and the number of
apoptotic cells was significantly lower in the CYGB-HuMSCs group
compared with the HuMSCs groups (Fig.
3A-C).
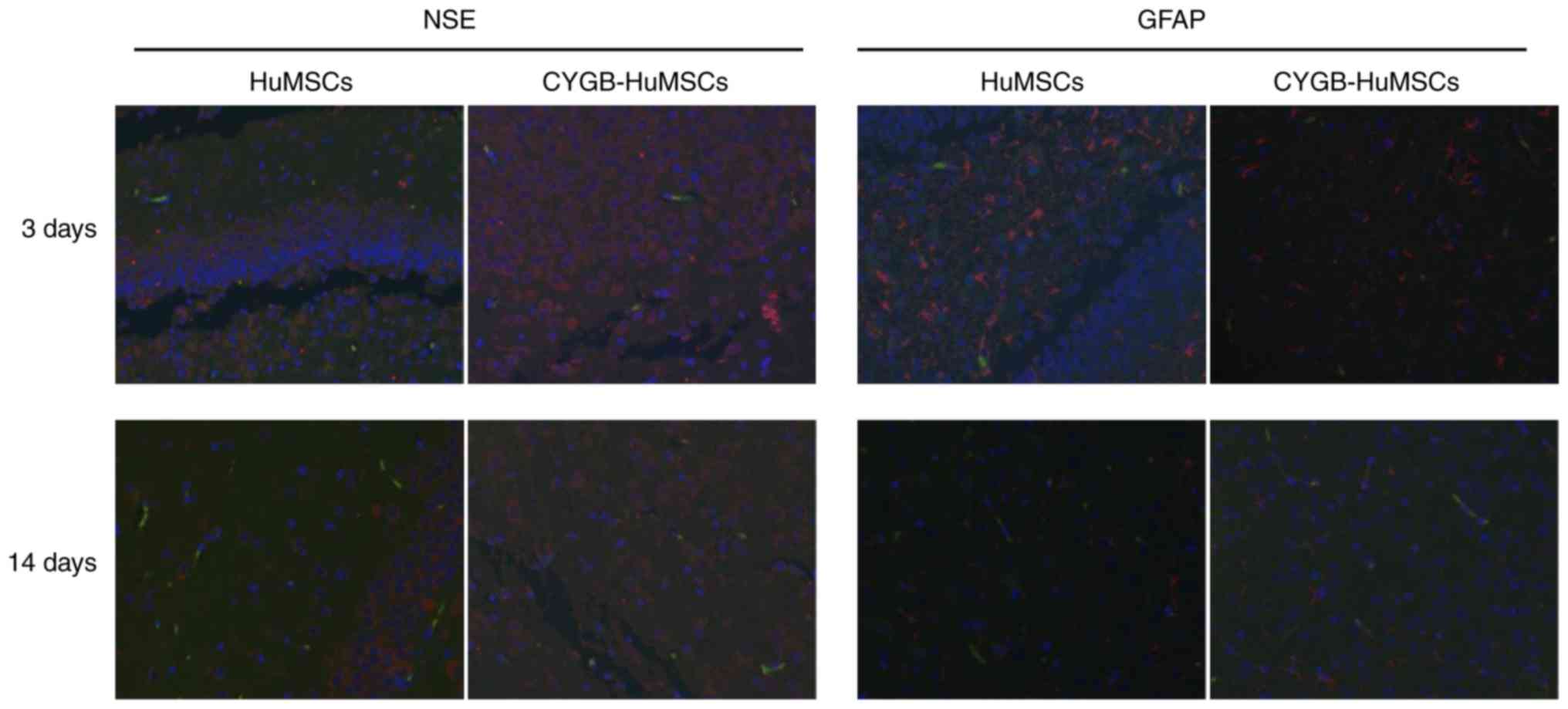
Immunofluorescence
To assess whether HuMSCs or CYGB-HuMSCs underwent
cell replacement via differentiation into neuron-like cells,
immunofluorescence co-localization analysis was performed 3 and 14
days after HI. It was identified that HuMSCs or CYGB-HuMSCs were
present and scattered in the damaged brain tissue (Fig. 4). Moreover, no eGFP and GFAP or
EGFP and NSE double-stained positive cells were observed and no
substantial differences were observed between 3 and 14 days.
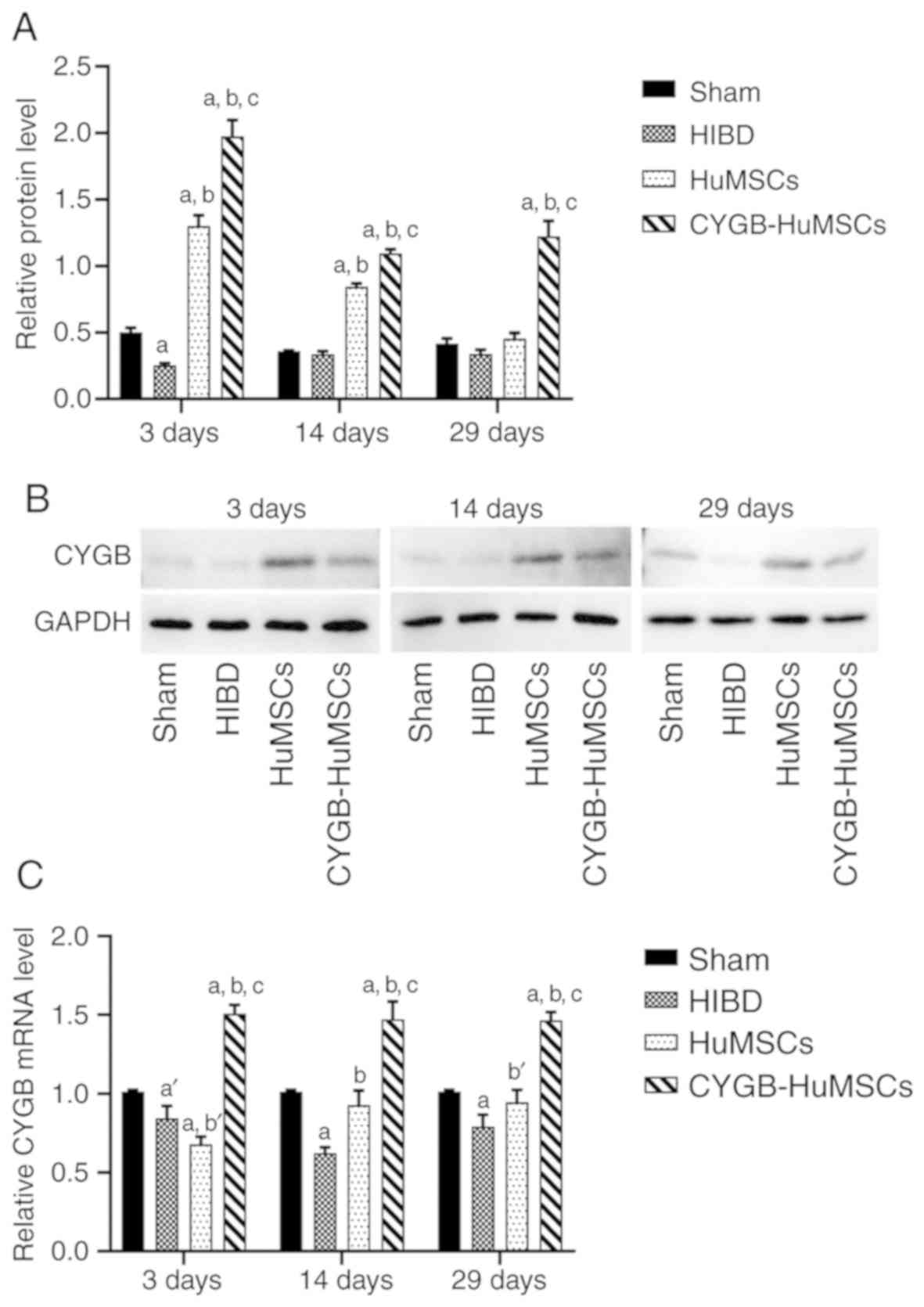
Expression of CYGB after
transplantation of HuMSCs
To investigative the mRNA and protein expression
levels of CYGB following CYGB-HuMSCs transplantation, RT-qPCR and
western blotting were performed on samples collected from all
groups following HI modeling for 3, 14 and 29 days. RT-qPCR results
demonstrated that CYGB mRNA expression was significantly increased
in the CYGB-HuMSCs group compared with the HIBD and HuMSCs groups
(Fig. 5C) at all experimental time
points. CYGB mRNA expression was significantly increased in the
HuMSCs group compared with the HIBD group, but significantly
decreased compared with the CYGB-HuMSC group at 14 and 29 days
(Fig. 5C). Western blotting
results identified that CYGB protein expression was also increased
in the CYGB-HuMSCs group compared with the HIBD and HuMSCs groups.
At all experimental time points, CYGB protein expression was also
increased in the HuMSCs group compared with the HIBD group, while
decreased compared with the CYGB-HuMSCs group. The expression
levels of CYGB protein in both the HuMSCs and CYGB-HuMSCs groups
gradually declined from 3 to 29 days (Fig. 5A and B).
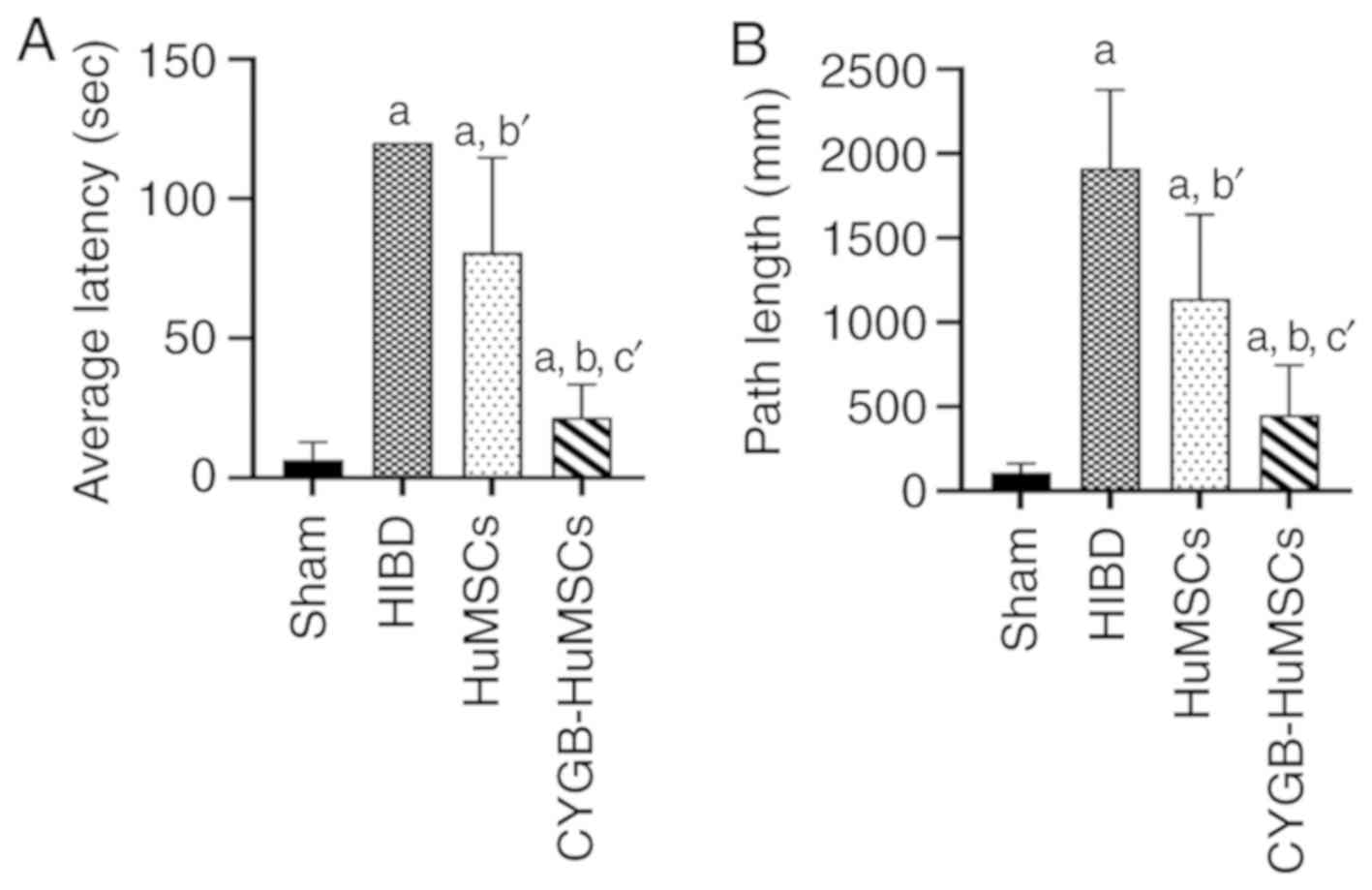
Morris-Water maze test
The Morris-Water maze experiment was performed 29
days after HI to evaluate long term learning and memory, which is
dependent on the function of the hippocampus and cortex (18). The results indicated a longer
escape latency (EL) in the HIBD group compared with the sham group.
EL in the HuMSCs group was significantly shorter compared with the
HIBD group, and in the CYGB-HuMSCs group, EL was shorter compared
with the HuMSCs group (Fig. 6A).
The PL was also recorded. PL was longer in the HIBD group compared
with the sham group (Fig. 6B).
Furthermore, HuMSCs transplantation significantly reduced PL
compared with the HIBD rats, and CYGB-HuMSCs transplantation
resulted in further reduced PL values compared with HuMSCs
transplantation. These results indicated a long-term
neuroprotective effect of either HuMSCs or CYGB-HuMSCs on HIBD.
CYGB-HuMSCs treatment modulates the
p38 pathway
As demonstrated by the western blotting results
(Fig. 7), compared with the Sham
group, the ratio of p-p38/p38 was significantly increased in the
HIBD group after HI modeling for 3, 14 and 29 days (Fig. 7A-C), suggesting that hypoxia and
oxidative stress mediated the stimulation of p38 signaling. HuMSCs
treatment significantly reduced the ratio of p-p38/p38 after HI
modeling for 3 and 29 days (Fig. 7A
and C). The ratio of p-p38/p38 at all experimental time points
in the CYGB-HuMSCs group was significantly lower compared with the
HuMSCs group (Fig. 7). Moreover,
the ratio of p-p38/p38 in the CYGB-HuMSCs group was significantly
lower compared with the sham group on day 3 after transplantation
of CYGB-HuMSCs (Fig. 7A).
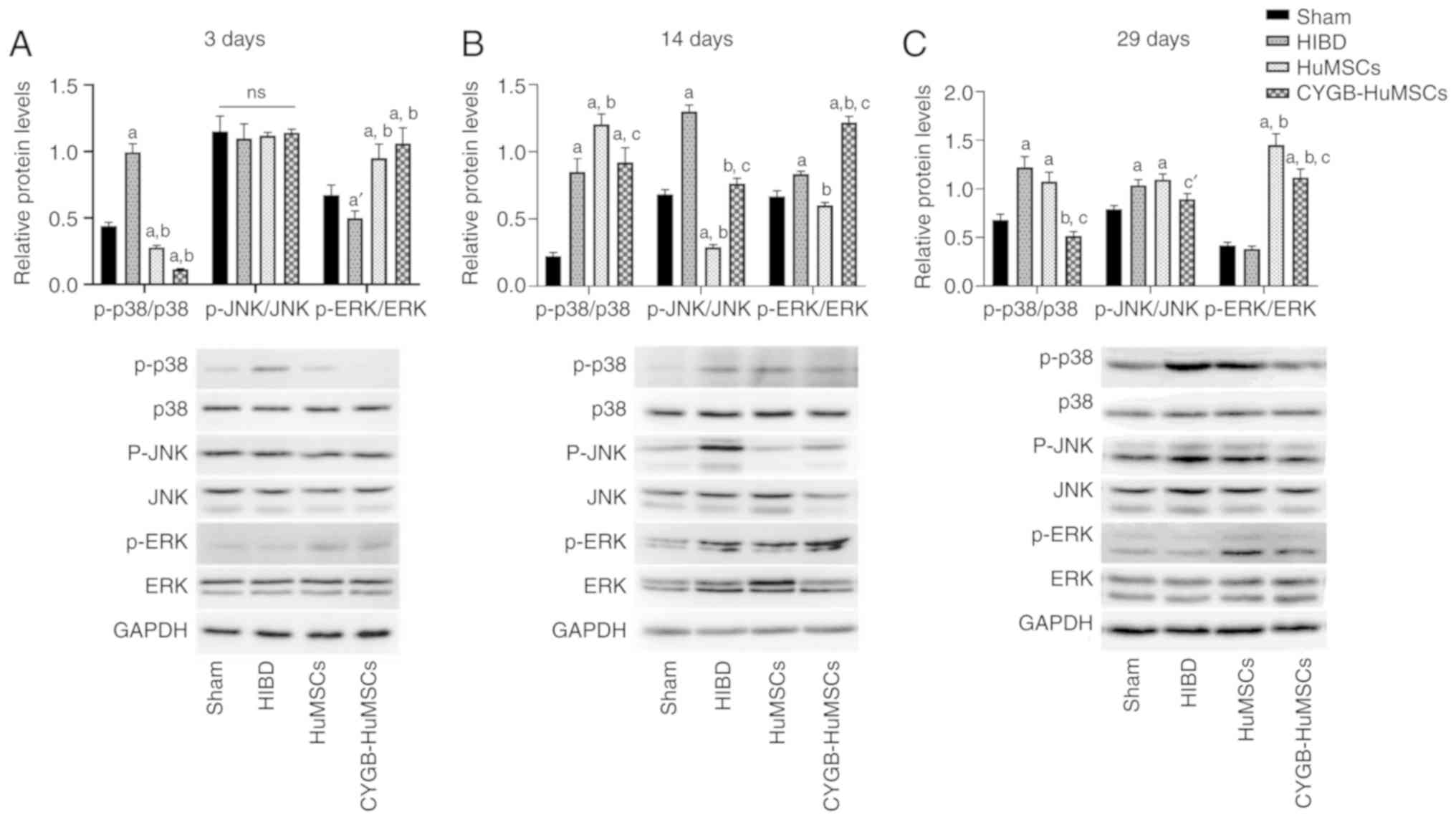 | Figure 7.Expression levels of p-p38, p38, ERK,
p-ERK, JNK and p-JNK after HI modeling for 3, 14 and 29 days (n=5
per group). GAPDH was used as the internal control. (A-C) The ratio
of p-p38/p38 was significantly increased in the HIBD group after HI
modeling for 3, 14 and 29 days. (A and C) The ratio of p-p38/p38
after HI modeling for 3 and 29 days in the HuMSCs group was
decreased. (A-C) The ratio of p-p38/p38 at all experimental time
points in the CYGB-HuMSCs group was significantly lower compared
with the HuMSCs group. (B and C) The ratio of p-JNK/JNK in the HIBD
group was significantly increased compared with the Sham group
after HI modeling for 14 and 29 days. (B) On day 14 after HI
modeling, compared with the HIBD group, the ratio of p-JNK/JNK was
decreased in the HuMSCs and CYGB-HuMSCs groups, but those in the
CYGB-HuMSCs group remained higher than the HuMSCs group. (C) On day
29 after HI modeling, there were no differences in the ratio of
p-JNK/JNK between the HIBD group and the HuMCSs or CYgB-HuMSCs
groups, but the ratio in the CYGB-HuMSCs group was significantly
lower compared with the HuMSCs group. (A, B) The ratio of p-ERK/ERK
in the HIBD group was significantly decreased compared with the
Sham group after HI modeling for 3 days, while increased at day 14
after HI modeling. (A and C) On days 3 and 29 after HI modeling,
compared with the HIBD group, the ratio of p-ERK/ERK was increased
in the HuMSCs and CYGB-HuMSCs groups. (B) On day 14 after HI
modeling, compared with the HIBD group, the ratio of p-ERK/ERK was
decreased in the HuMSCs group, while increased in the CYGB-HuMSCs
group. aP<0.01 vs. Sham group at the same time; bP<0.01 vs.
HIBD group at the same time; cP<0.01 vs. HuMSCs group at the
same time; a'P<0.05 vs. Sham group at the same time; c'P<0.05
vs. HuMSCs group at the same time; ns, no significant difference
vs. other groups. p-, phosphorylated; HI, hypoxia-ischemia injury;
CYGB, Cytoglobin; HuMSC, human umbilical cord-derived mesenchymal
stem cells; HIBD, hypoxic-ischemia brain damage. |
The ratio of p-JNK/JNK in the HIBD group was
significantly increased compared with the Sham group after HI
modeling for 14 and 29 days (Fig. 7B
and C). On day 14 after HI modeling, compared with the HIBD
group, the ratio of p-JNK/JNK was decreased in the HuMSCs group and
CYGB-HuMSCs group; however, the ratio in the CYGB-HuMSCs group
remained higher than the HuMSCs group (Fig. 7B). On day 29 after HI modeling,
there were no significant differences in the ratio of p-JNK/JNK
between the HIBD group and HuMCSs or CYGB-HuMSCs groups, but the
ratio in the CYGB-HuMSCs group was lower compared with the HuMSCs
group (Fig. 7C).
The ratio of p-ERK/ERK in the HIBD group was
significantly decreased compared with the Sham group after HI
modeling for 3 days, while increased at day 14 after HI modeling
(Fig. 7A and B). On days 3 and 29
after HI modeling, compared with the HIBD group, the ratio of
p-ERK/ERK was increased in the HuMSCs group and CYGB-HuMSCs group
(Fig. 7A and C). On day 14 after
HI modeling, compared with the HIBD group, the ratio of p-ERK/ERK
was decreased in the HuMSCs group, while increased in the
CYGB-HuMSCs group (Fig. 7B).
Discussion
HIBD is a significant threat to neonatal health,
which results in disability and mortality in infants and children
(5,6). Despite an increase in the number of
studies that have been performed to investigate the pathogenesis of
HIBD, hypothermia and systemic supportive treatment are the primary
therapeutic options for HIBD (35). To the best of our knowledge, the
present study was the first to demonstrate that intranasal
transplantation of CYGB-HuMSCs exhibited neuroprotective and
antiapoptotic effects in HIBD. In a rat model of HIBD, intranasal
administration of both HuMSCs and CYGB-HuMSCs resulted in reduced
severity of histological and functional deficits. In addition, the
reduction in the CYGB-HuMSCs group was more prominent compared with
the HuMSCs group.
CYGB has been extensively studied in the past 15
years (13), following its
discovery. Different from hemoglobin, CYGB is found outside of red
blood cells and is classified as a non-erythroid globin, similar to
other globins, such as neuroglobin and myoglobin (36). The potential functions of these
non-erythroid globins are associated with tissue and cell
protection under hypoxic conditions, ischemic conditions and during
oxidative stress (37). Due to the
structure of the hemeglobin, CYGB can reversibly bind to oxygen and
other small molecules (38). The
ability of CYGB to store and sense oxygen, as well as its
involvement in nitrite and NO metabolism, are being increasingly
studied and understood (39,40).
It has been reported that, similar to other hemoglobins, CYGB can
scavenge ROS and NO, as well as produce NO from nitrite, thus
reducing oxidative stress and protecting organs and cells (41–44).
Our previous study revealed that pretreatment of
CYGB-overexpression reduced neonatal rat HI injury, as well as
improved long term memory and athletic ability following neonatal
HI (18). However, whether
overexpression of CYGB serves a protective role in neurons
following HIBD has not been previously studied, and there is no
established means of introducing the CYGB gene safely into living
animals.
In the present study, the effect of transplantation
of CYGB-HuMSCs in an HIBD rats model was determined, and was
developed based the application of stem cells for treatment of
various brain diseases and our previous study (18). The current results suggested that
CYGB-HuMSCs reduced neuron cell apoptosis caused by HIBD. The
improvement in long term neurological function 29 days after HIBD
further demonstrated the neuroprotective effect of CYGB-HuMSCs
transplantation. HuMSCs also exhibited a neuroprotective and
antiapoptotic effect in HIBD, but the CYGB-HuMSCs exhibited more
prominent neuroprotective effects. In addition, RT-qPCR and western
blotting results identified that CYGB mRNA and protein expression
levels were increased from day 3 to day 29 after transplantation of
CYGB-HuMSCs. Thus, it was suggested that CYGB functions as an
endogenous neuroprotective protein, and that CYGB-HuMSCs may act as
a gene transfer vector for CYGB, allowing CYGB and HuMSCs to work
together to improve the neuroprotective effects. Therefore, these
results highlight a novel approach for treatment of neonatal
HIBD.
Previous studies have shown that HuMSCs do not
undergo cell replacement via differentiation into neuron-like
cells, but rather exert neuroprotective effects (45,46).
In the present study, HuMSCs or CYGB-HuMSCs administered via nasal
transplantation did not differentiate into cells expressing neural
or glial cell marker proteins; a result that is similar to previous
studies (45,46). Therefore, it was speculated that
the neuroprotective effects of HuMSCs may be achieved via the
action of exosomes to promote endogenous repair of nerve cells
(47–49). Proteins secreted by MSCs can
promote the repair of damaged tissues via a variety of mechanisms,
including prevention of apoptosis, regulation of inflammatory
responses and promotion of endogenous repair mechanisms, such as
angiogenesis and neurogenesis, rather than cell replacement
(48). In a HIBD animal model,
MSCs provide a suitable environment, including increasing the local
levels of brain-derived neurotrophic factor, basic fibroblast
growth factor, neurotrophic factor and nerve growth factor, whilst
downregulating the expression of proinflammatory cytokines, such as
IL-1 and IL-6, which may also contribute to the repair of nerve
cells (47,50). Previous studies have reported that
CYGB serves a neuroprotective role by reducing cerebral infarctions
and apoptosis caused by oxidative stress in vivo, and also
revealed that hypoxia inducible factor-1α/CYGB/vascular endothelial
growth factor signaling may serve an important role in the
CYGB-mediated antioxidant mechanism (18). Therefore, it was hypothesized that
the neuroprotective effects of CYGB-HuMSCs is not via cell
replacement, but via the various cytokines secreted by HuMSCs and
the antioxidant effects of overexpression of CYGB transfected into
animals by HuMSCs.
The p38 mitogen-activated protein kinase (MAPK)
signaling pathway allows cells to interpret a wide range of
external signals and respond appropriately by generating a plethora
of different biological effects (51). It has been revealed that p38 MAPK
is a mediator of hypoxia-induced cerebrovascular inflammation
(52). The p38 MAPK signaling
pathway enhances the production of a range of proinflammatory
cytokines, including IL-β, TNF-α and IL-8 (53,54).
Moreover, p38 MAPK has been suggested to be involved in the context
of ischemia-induced stress in the brain (55). For example, p38 MAPK inhibits the
hypoxia response pathway via EGL-9 in neurons (56). The p38 MAPK can also function as a
mediator of ROS-mediated signaling, and either activate or suppress
cell cycle progression, depending on the activation stimulus
(57). Proteomic and biochemical
analyses further demonstrated that p38 MAPK signaling mediates cell
apoptosis (58), while in
vitro studies have shown that overexpression of CYGB suppressed
p38 expression and protected glomerular mesangial cells from
oxidative stress (59).
In the present study, the ratio of p-p38/p38 was
significantly increased in the HIBD group. However, HuMSCs
supplementation protected against neuron cell apoptosis as
demonstrated by the decreased ratio of p-p38/p38. In addition, the
CYGB-HuMSCs group had an improved antiapoptotic effect accompanied
with a greater degree of decreased p38 expression compared with the
HuMSCs group. The present results also suggested that the
CYGB-HuMSCs group had significantly downregulated phosphorylation
levels of p38, which was accompanied with a significantly increased
expression level of CYGB, compared with the sham group on day 3.
Thus, it was indicated that exogenous CYGB may modulate p38
expression. The current results highlight the involvement of p38
MAPK signaling in HIBD, as well as suggesting that CYGB-HuMSCs
treatment may protect animals from HIBD by avoiding activation of
the p38 MAPK signaling pathway.
In conclusion, to the best of our knowledge, the
present study was the first to demonstrate that intranasal
transplantation of CYGB-HuMSCs can serve as a gene transporter, and
that it exerts neuroprotective and antiapoptotic effects in HIBD.
Additionally, it was identified that CYGB-HuMSCs may exert its
protective properties via the p38 MAPK pathway. The results of the
current study demonstrate a novel therapeutic approach for
treatment of HIBD. However, further investigation is required to
identify the mechanism via which CYGB-HuMSCs contributes to the
molecular pathogenesis of HIBD.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Science and
Technology Project from the Science Technology and Innovation
Committee of Shenzhen Municipality (grant no.
JCYJ20160429141742207), the National Natural Science Foundation of
China (grant nos. 81671525 and 81070478) and the Sanming Project of
Medicine in Shenzhen (grant no. SZSM201512033).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SFT, YBC and HHY designed the study and performed
the experiments. HHY, LM and LCX acquired the data. SFT and HHY
analyzed the data. HHY, YBC and LM prepared the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Committees of
Maternal and Child Health Care Hospital of Shenzhen University
(Guangdong, China), and of the Institutional Animal Care and Use
Committee of Shenzhen University (Guangdong, China). Written
informed consent was obtained from the maternal donors and/or
guardians.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Novak CM, Ozen M and Burd I: Perinatal
brain injury: Mechanisms, prevention, and outcomes. Clin Perinatol.
45:3493–375. 2018. View Article : Google Scholar
|
|
2
|
du Plessis AJ and Volpe JJ: Perinatal
brain injury in the preterm and term newborn. Curr Opin Neurol.
15:151–157. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Triulzi F, Parazzini C and Righini A:
Patterns of damage in the mature neonatal brain. Pediatr Radiol.
36:608–620. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gale C, Statnikov Y, Jawad S, Uthaya SN
and Modi N; Brain Injuries expert working group, : Neonatal brain
injuries in England: Population-based incidence derived from
routinely recorded clinical data held in the National Neonatal
Research Database. Arch Dis Child Fetal Neonatal Ed. 103:301–306.
2018. View Article : Google Scholar
|
|
5
|
Bass JL, Corwin M, Gozal D, Moore C,
Nishida H, Parker S, Schonwald A, Wilker RE, Stehle S and Kinane
TB: The effect of chronic or intermittent hypoxia on cognition in
childhood: A review of the evidence. Pediatrics. 114:805–816. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yildiz EP, Ekici B and Tatli B: Neonatal
hypoxic ischemic encephalopathy: An update on disease pathogenesis
and treatment. Expert Rev Neurother. 17:449–459. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chiang MC, Jong YJ and Lin CH: Therapeutic
hypothermia for neonates with hypoxic ischemic encephalopathy.
Pediatr Neonatol. 58:475–483. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Srinivasakumar P, Zempel J, Wallendorf M,
Lawrence R, Inder T and Mathur A: Therapeutic hypothermia in
neonatal hypoxic ischemic encephalopathy: Electrographic seizures
and magnetic resonance imaging evidence of injury. J Pediatr.
163:465–470. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ma H, Sinha B, Pandya RS, Lin N, Popp AJ,
Li J, Yao J and Wang X: Therapeutic hypothermia as a
neuroprotective strategy in neonatal hypoxic-ischemic brain injury
and traumatic brain injury. Curr Mol Med. 12:1282–1296. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pesce A, Bolognesi M, Bocedi A, Ascenzi P,
Dewilde S, Moens L, Hankeln T and Burmester T: Neuroglobin and
cytoglobin. Fresh blood for the vertebrate globin family. EMBO Rep.
3:1146–1151. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gell DA: Structure and function of
haemoglobins. Blood Cells Mol Dis. 70:13–42. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Burmester T and Hankeln T: Function and
evolution of vertebrate globins. Acta Physiol (Oxf). 211:501–514.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Trent JT III and Hargrove MS: A
ubiquitously expressed human hexacoordinate hemoglobin. J Biol
Chem. 277:19538–19545. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stagner JI, Parthasarathy SN, Wyler K and
Parthasarathy RN: Protection from ischemic cell death by the
induction of cytoglobin. Transplant Proc. 37:3452–3453. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hankeln T, Ebner B, Fuchs C, Gerlach F,
Haberkamp M, Laufs TL, Roesner A, Schmidt M, Weich B, Wystub S, et
al: Neuroglobin and cytoglobin in search of their role in the
vertebrate globin family. J Inorg Biochem. 99:110–119. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Avivi A, Gerlach F, Joel A, Reuss S,
Burmester T, Nevo E and Hankeln T: Neuroglobin, cytoglobin, and
myoglobin contribute to hypoxia adaptation of the subterranean mole
rat Spalax. Proc Natl Acad Sci USA. 107:21570–21575. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Singh S, Manda SM, Sikder D, Birrer MJ,
Rothermel BA, Garry DJ and Mammen PP: Calcineurin activates
cytoglobin transcription in hypoxic myocytes. J Biol Chem.
284:10409–10421. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tian SF, Yang HH, Xiao DP, Huang YJ, He
GY, Ma HR, Xia F and Shi XC: Mechanisms of neuroprotection from
hypoxia-ischemia (HI) brain injury by up-regulation of cytoglobin
(CYGB) in a neonatal rat model. J Biol Chem. 288:15988–16003. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li Y and Chopp M: Marrow stromal cell
transplantation in stroke and traumatic brain injury. Neurosci
Lett. 456:120–123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao L, Johnson T and Liu D: Therapeutic
angiogenesis of adipose-derived stem cells for ischemic diseases.
Stem Cell Res Ther. 8:1252017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park WS, Sung SI, Ahn SY, Yoo HS, Sung DK,
Im GH, Choi SJ and Chang YS: Hypothermia augments neuroprotective
activity of mesenchymal stem cells for neonatal hypoxic-ischemic
encephalopathy. PLoS One. 10:e01208932015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gonzales-Portillo GS, Reyes S, Aguirre D,
Pabon MM and Borlongan CV: Stem cell therapy for neonatal
hypoxic-ischemic encephalopathy. Front Neurol. 5:1472014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
van Velthoven CT, Kavelaars A and Heijnen
CJ: Mesenchymal stem cells as a treatment for neonatal ischemic
brain damage. Pediatr Res. 71:474–481. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang X, Zhang Q, Li W, Nie D, Chen W, Xu
C, Yi X, Shi J, Tian M, Qin J, et al: Therapeutic effect of human
umbilical cord mesenchymal stem cells on neonatal rat
hypoxic-ischemic encephalopathy. J Neurosci Res. 92:35–45. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou X, Gu J, Gu Y, He M, Bi Y, Chen J and
Li T: Human umbilical cord-derived mesenchymal stem cells improve
learning and memory function in hypoxic-ischemic brain-damaged rats
via an IL-8-mediated secretion mechanism rather than
differentiation pattern induction. Cell Physiol Biochem.
35:2383–2401. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim YS, Hwang KA, Go RE, Kim CW and Choi
KC: Gene therapy strategies using engineered stem cells for
treating gynecologic and breast cancer patients (Review). Oncol
Rep. 33:2107–2112. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
High KA and Roncarolo MG: Gene therapy. N
Engl J Med. 381:455–464. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Aiuti A, Slavin S, Aker M, Ficara F, Deola
S, Mortellaro A, Morecki S, Andolfi G, Tabucchi A, Carlucci F, et
al: Correction of ADA-SCID by stem cell gene therapy combined with
nonmyeloablative conditioning. Science. 296:2410–2413. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang H, Qiu X, Ni P, Qiu X, Lin X, Wu W,
Xie L, Lin L, Min J, Lai X, et al: Immunological characteristics of
human umbilical cord mesenchymal stem cells and the therapeutic
effects of their transplantion on hyperglycemia in diabetic rats.
Int J Mol Med. 33:263–270. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kilkenny C, Browne WJ, Cuthill IC, Emerson
M and Altman DG: Improving bioscience research reporting: The
ARRIVE guidelines for reporting animal research. PLoS Biol.
8:e10004122010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rice JE III, Vannucci RC and Brierley JB:
The influence of immaturity on hypoxic-ischemic brain damage in the
rat. Ann Neurol. 9:131–141. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Donega V, Nijboer CH, van Tilborg G,
Dijkhuizen RM, Kavelaars A and Heijnen CJ: Intranasally
administered mesenchymal stem cells promote a regenerative niche
for repair of neonatal ischemic brain injury. Exp Neurol.
261:53–64. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hoogewijs D, Vogler M, Zwenger E, Krull S
and Zieseniss A: Oxygen-dependent regulation of aquaporin-3
expression. Hypoxia (Auckl). 4:91–97. 2016.PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Douglas-Escobar M and Weiss MD:
Hypoxic-ischemic encephalopathy: A review for the clinician. JAMA
Pediatr. 169:397–403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Burmester T, Ebner B, Weich B and Hankeln
T: Cytoglobin: A novel globin type ubiquitously expressed in
vertebrate tissues. Mol Biol Evol. 19:416–421. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ascenzi P, Gustincich S and Marino M:
Mammalian nerve globins in search of functions. IUBMB life.
66:268–276. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Amdahl MB, DeMartino AW, Tejero J and
Gladwin MT: Cytoglobin at the crossroads of vascular remodeling.
Arterioscler Thromb Vasc Biol. 37:1803–1805. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vinogradov SN and Moens L: Diversity of
globin function: Enzymatic, transport, storage, and sensing. J Biol
Chem. 283:8773–8777. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tejero J and Gladwin MT: The globin
superfamily: Functions in nitric oxide formation and decay. Biol
Chem. 395:631–639. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hodges NJ, Innocent N, Dhanda S and Graham
M: Cellular protection from oxidative DNA damage by over-expression
of the novel globin cytoglobin in vitro. Mutagenesis. 23:293–298.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
De Beuf A, Hou XH, D'Haese PC and Verhulst
A: Epoetin delta reduces oxidative stress in primary human renal
tubular cells. J Biomed Biotechnol. 2010:3957852010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lv W, Booz GW, Fan F, Wang Y and Roman RJ:
Oxidative stress and renal fibrosis: Recent insights for the
development of novel therapeutic strategies. Front Physiol.
9:1052018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lilly B, Dammeyer K, Marosis S,
McCallinhart PE, Trask AJ, Lowe M and Sawant D: Endothelial
cell-induced cytoglobin expression in vascular smooth muscle cells
contributes to modulation of nitric oxide. Vascul Pharmacol.
110:7–15. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Hong SQ, Zhang HT, You J, Zhang MY, Cai
YQ, Jiang XD and Xu RX: Comparison of transdifferentiated and
untransdifferentiated human umbilical mesenchymal stem cells in
rats after traumatic brain injury. Neurochem Res. 36:2391–2400.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lin YC, Ko TL, Shih YH, Lin MY, Fu TW,
Hsiao HS, Hsu JY and Fu YS: Human umbilical mesenchymal stem cells
promote recovery after ischemic stroke. Stroke. 42:2045–2053. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
van Velthoven CT, Kavelaars A, van Bel F
and Heijnen CJ: Nasal administration of stem cells: A promising
novel route to treat neonatal ischemic brain damage. Pediatr Res.
68:419–422. 2010.PubMed/NCBI
|
|
48
|
Cunningham CJ, Redondo-Castro E and Allan
SM: The therapeutic potential of the mesenchymal stem cell
secretome in ischaemic stroke. J Cereb Blood Flow Metab.
38:1276–1292. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li Y, Cheng Q, Hu G, Deng T, Wang Q, Zhou
J and Su X: Extracellular vesicles in mesenchymal stromal cells: A
novel therapeutic strategy for stroke. Exp Ther Med. 15:4067–4079.
2018.PubMed/NCBI
|
|
50
|
Gu Y, He M, Zhou X, Liu J, Hou N, Bin T,
Zhang Y, Li T and Chen J: Endogenous IL-6 of mesenchymal stem cell
improves behavioral outcome of hypoxic-ischemic brain damage
neonatal rats by supressing apoptosis in astrocyte. Sci Rep.
6:185872016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cuadrado A and Nebreda AR: Mechanisms and
functions of p38 MAPK signalling. Biochem J. 429:403–417. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sanchez A, Tripathy D, Yin X, Desobry K,
Martinez J, Riley J, Gay D, Luo J and Grammas P: p38 MAPK: A
mediator of hypoxia-induced cerebrovascular inflammation. J
Alzheimers Dis. 32:587–597. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cakmak H, Seval-Celik Y, Arlier S,
Guzeloglu-Kayisli O, Schatz F, Arici A and Kayisli UA: p38
Mitogen-activated protein kinase is involved in the pathogenesis of
endometriosis by modulating inflammation, but not cell survival.
Reprod Sci. 25:587–597. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Amir M, Somakala K and Ali S: p38 MAP
kinase inhibitors as anti inflammatory agents. Mini Rev Med Chem.
13:2082–2096. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Cheung WD and Hart GW: AMP-activated
protein kinase and p38 MAPK activate O-GlcNAcylation of neuronal
proteins during glucose deprivation. J Biol Chem. 283:13009–13020.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Park EC and Rongo C: The p38 MAP kinase
pathway modulates the hypoxia response and glutamate receptor
trafficking in aging neurons. Elife. 5:e120102016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Tormos AM, Taléns-Visconti R, Nebreda AR
and Sastre J: p38 MAPK: A dual role in hepatocyte proliferation
through reactive oxygen species. Free Radic Res. 47:905–916. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang H, Tao L, Jiao X, Gao E, Lopez BL,
Christopher TA, Koch W and Ma XL: Nitrative thioredoxin
inactivation as a cause of enhanced myocardial ischemia/reperfusion
injury in the aging heart. Free Radic Biol Med. 43:39–47. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Xu ZJ, Shu S, Li ZJ, Liu YM, Zhang RY and
Zhang Y: Liuwei Dihuang pill treats diabetic nephropathy in rats by
inhibiting of TGF-β/SMADS, MAPK, and NF-kB and upregulating
expression of cytoglobin in renal tissues. Medicine (Baltimore).
96:e58792017. View Article : Google Scholar : PubMed/NCBI
|