Introduction
Breast cancer is a malignant disease that seriously
damages the health of women worldwide and is considered to be the
second leading cause of cancer mortality in women according to the
2018 American Cancer Report (1).
Patients with breast cancer undergo surgery or radiation therapy
combined with chemotherapy; despite the improved survival rates
with these treatments, drug resistance and recurrence in certain
patients continues to lead to poor prognosis and treatment failure
(2). Therefore, developing
effective adjuvant therapy with fewer side-effects compared with
main treatments to reduce the progression of breast cancer has
become an urgent need.
Natural products have been attracting increasing
interest worldwide due to their extensive biological activities and
minimum toxicity (3,4). These natural products have
contributed to improving the quality of life and several of them
have been used clinically to treat various diseases, including
cancer (5). Matrine is a bioactive
compound found in the traditional Chinese medicine Sophora
flavescens (Fig. 1). It
demonstrates versatile pharmacological properties, including
anti-inflammatory (6), antiviral
(7), antitumor (8) and anti-fibrotic properties (9). Matrine has been demonstrated to
effectively influence the development and progression of cancer
involving the blockade of cell cycle progression (10) and the induction of apoptosis
(11). Previous studies also
confirmed that matrine suppressed proliferation, invasion and
metastasis, regulating the expression of oncogenes and inhibiting
the production of cytokines of breast cancer in vitro and
in vivo (12,13). Due to such promising efficacy,
matrine is currently used as an adjuvant in China to improve 5-year
survival rates and quality of life in breast cancer patients and to
enhance their immunity (6).
Nevertheless, the functional role and exact mechanisms of matrine
against breast cancer remain to be elucidated.
Autophagy is an evolutionarily conserved mechanism
responsible for cellular homeostasis in eukaryotic cells. It is a
form of programmed cell death that regulates tumor development and
progression (14,15). In addition, autophagy can have
opposing roles in different cell types (16), for example activation of autophagy
can contribute to the survival of human lung carcinoma A549 cells
(17), or promote cell death of
U251 (14) and SMMC7721 cells
(15). Autophagy triggered in
response to anticancer therapy can be associated with cell death,
leading to the activation of the pro-death signaling pathway in
cancer cells (18). Previous
studies demonstrated that autophagy is involved in breast cancer
biology (19,20) and in the development of therapy
resistance in breast cancer (21).
Recently, studies demonstrated that autophagy served a crucial role
in breast cancer therapeutics. For example, baicalein was revealed
to suppress cell proliferation and induce apoptosis and autophagy
in breast cancer cells by inhibiting the PI3K/AKT pathway both
in vitro and in vivo (22). In another study, corilagin was
revealed to inhibit breast cancer growth via reactive oxygen
species (ROS)-dependent apoptosis and autophagy (23). Studies have demonstrated that
matrine possesses potential antitumor activities in breast cancer
cells (5,24). However, the exact effects of
autophagy in regulating the cellular sensitivity of breast cancer
to matrine treatment remain to be elucidated.
Therefore, the present study investigated the
influence of matrine on breast cancer and explored the potential
mechanisms of autophagy involved in the anti-breast cancer activity
of matrine. To this end, a widely used epithelial cancer cell line
MCF-7, derived from breast adenocarcinoma, was employed as an in
vitro model to investigate the anticancer effects of matrine on
breast cancer by focusing on its influence on autophagy. The
findings of the present study may afford new insights into the
mechanisms of matrine in inhibiting breast cancer.
Materials and methods
Materials
Matrine was purchased from Shanghai Yuanye
Biotechnology Co., Ltd., (purity 98%). DMEM was obtained from Gibco
(Thermo Fisher Scientific, Inc.). Penicillin-streptomycin was
purchased from Beijing Solarbio Science and Technology Co., Ltd.
Fetal bovine serum (FBS) was purchased from Zhejiang Tianhang
Biotechnology Co., Ltd. BCA protein assay kit and Annexin V-FITC
apoptosis detection kit were provided by Beyotime Institute of
Biotechnology. Cell Counting Kit-8 (CCK-8) was obtained from
Dojindo Molecular Technologies, Inc. Antibodies to light chain 3
(LC3)-II rabbit mAb (cat. no. 3868s), p62 rabbit mAb (cat. no.
5114), phosphorylated (p-)AKT rabbit mAb (cat. no. 13038),
p-mammalian target of rapamycin (mTOR) rabbit mAb (cat. no. 5536),
β-actin rabbit mAb (cat. no. 8457) and anti-rabbit IgG alkaline
phosphatase (AP)-linked antibodies (cat. no. 7054) were obtained
from Cell Signaling Technology, Inc.
Cell culture
MCF-7 cells were provided by Professor Jianqiang Liu
(Guangdong Medical University, China). The cells were maintained in
DMEM supplemented with 10% FBS and 1% penicillin-streptomycin at
37°C in a humidified atmosphere of 5% CO2.
Cell viability assay
MCF-7 cells were seeded into the 96-well plate at
5×104 cells/ml. After the cells were attached, they were
incubated with 2, 4, 8 and 10 mM matrine for 24, 48 and 72 h. Cell
viability of MCF-7 was determined using the CCK-8 assay kit in
accordance with the manufacturer's protocol. The absorbance at 450
nm was measured with a microplate reader.
Morphological changes of MCF-7
cells
MCF-7 cells (1×105 cells/ml) were seeded
into 12-well plates and the morphological changes were observed by
optical microscope at ×200 magnification and images were captured
after the cells had been treated with matrine for 24 h.
Apoptosis analysis
Following matrine exposure, MCF-7 were collected by
scraping with a rubber policeman on the ice followed by
centrifugation at 1,000 × g for 5 min at 4°C and re-suspended in
fresh medium at a density of 2×105 cells/ml. Apoptosis
was assessed according to the manufacturer's instructions. The
cells were labeled with 5 µl Annexin V-FITC and 5 µl of propidium
iodide (PI) for 10 min in the dark. Samples were analyzed using
flow cytometry (FACSVerse; BD Biosciences) and data were analyzed
with FlowJo 7.6.1 software (Tree star, Inc.). Annexin
V+/PI− cells were identified as early
apoptotic cells, whereas Annexin V+/PI+ cells
were designated as late apoptotic cells.
Hoechst 33342 staining
Apoptotic condensed nuclear changes were identified
with Hoechst 33342 staining. The MCF-7 cells were washed twice with
PBS and then incubated in Hoechst 33342 at 37°C for 30 min
following matrine treatment. Then, alterations in nuclear
morphology were imaged under a Olympus IX73 fluorescence microscope
at 377 nm excitation and 447 nm emission wavelengths (Olympus
Corporation) from five random fields (magnification, ×200).
Western blot analysis
Total protein was extracted using RIPA buffer
containing 1 mM phenylmethylsulphonyl fluoride (Beyotime Institute
of Biotechnology) at 4°C for 30 min. The concentrations of the
protein were determined using a BCA protein assay kit. Equal
amounts of proteins (20 µg) were subjected to 10%
SDS-polyacrylamide gel electrophoresis and electro-transferred to
Immuno-Blot PVDF membranes. Membranes were blocked with 5% (w/v)
bovine serum albumin (Beyotime Institute of Biotechnology, China)
for 2 h at 4°C. Subsequently, the membranes were incubated with
primary antibody (LC3-II, p62, p-AKT, p-mTOR, and β-actin; all
1:1,000) overnight at 4°C. After washing with 0.1% (v/v) Tween-20
in TBS, the anti-rabbit IgG AP-linked secondary antibody was
applied for 1 h at room temperature. The bands were developed using
the ECL detection system (Bio-Rad Laboratories, Inc.) and
densitometry analysis was performed by ImageJ software (version
1.46, National Institutes of Health). β-actin served as the
internal reference concurrently. All data were expressed as the
relative intensity compared to the control group for the
statistical analyses.
Statistical analysis
Data were reported as the means ± standard deviation
of at least three independent experiments conducted in triplicate.
Statistical analyses were performed using GraphPad Prism 8.0
software (GraphPad Software, Inc.). Differences between groups were
performed using Student's t-test (2-group comparisons) or one-way
analysis of variance followed by Tukey's post hoc test (>2
groups). P<0.05 was considered to indicate a statistically
significant difference.
Results
Matrine decreases cell viability of
MCF-7 cells
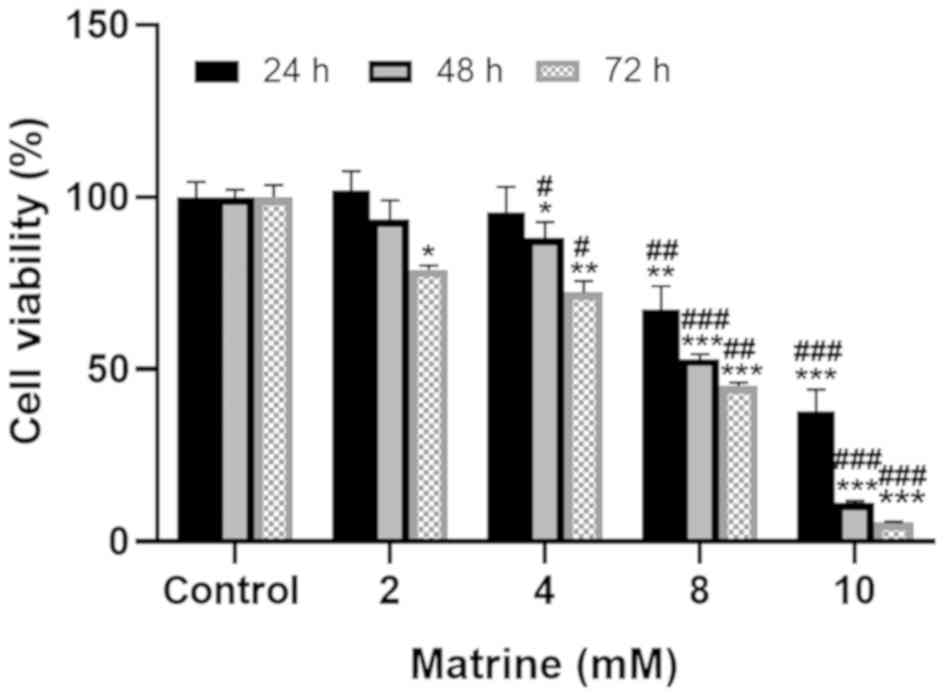
To elucidate the impact of matrine on the cell
viability of human breast cancer MCF-7 cells, the cells were
cultured with 2, 4, 8 and 10 mM matrine for 24, 48 and 72 h,
following which the CCK-8 assay was conducted. As revealed in
Fig. 2, matrine significantly
caused the inhibition of the viability of MCF-7 cells (P<0.05)
and this effect was dose-dependent. Over the course of extended
treatments (72 h), matrine exhibited a greater inhibitory effect
compared with that at 24 and 48 h (P<0.01). These results
indicated that matrine exhibited anti-cell growth effects on breast
cancer MCF-7 cells in a dose- and time-dependent manner.
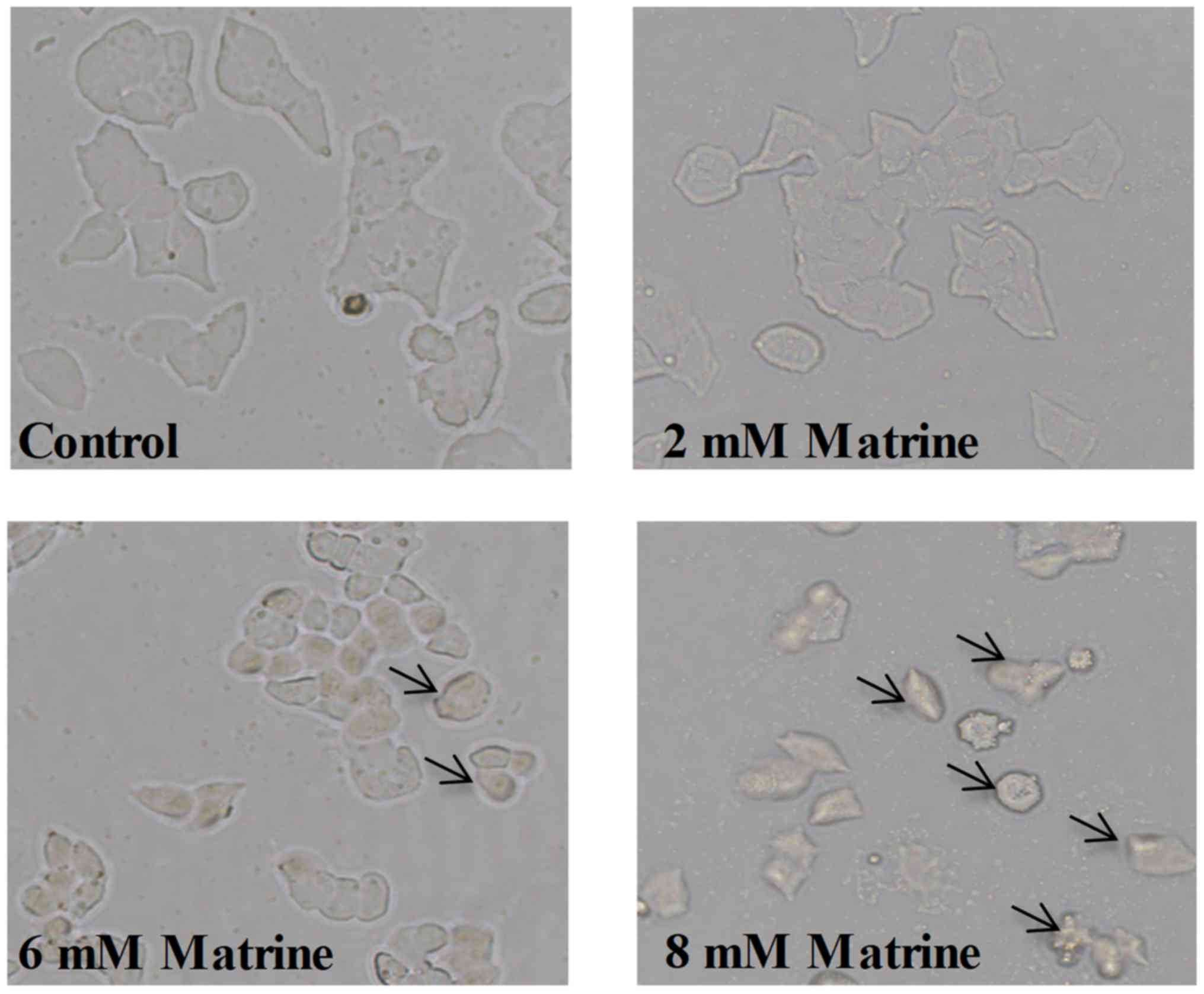
Matrine induces morphologic changes of
MCF-7 cells
Morphological features were observed in MCF-7 cells
following treatment with various concentrations of matrine. MCF-7
cells demonstrated shrinkage, membrane blebbing, ballooning and
partial detachment after treatment with matrine for 24 h (Fig. 3).
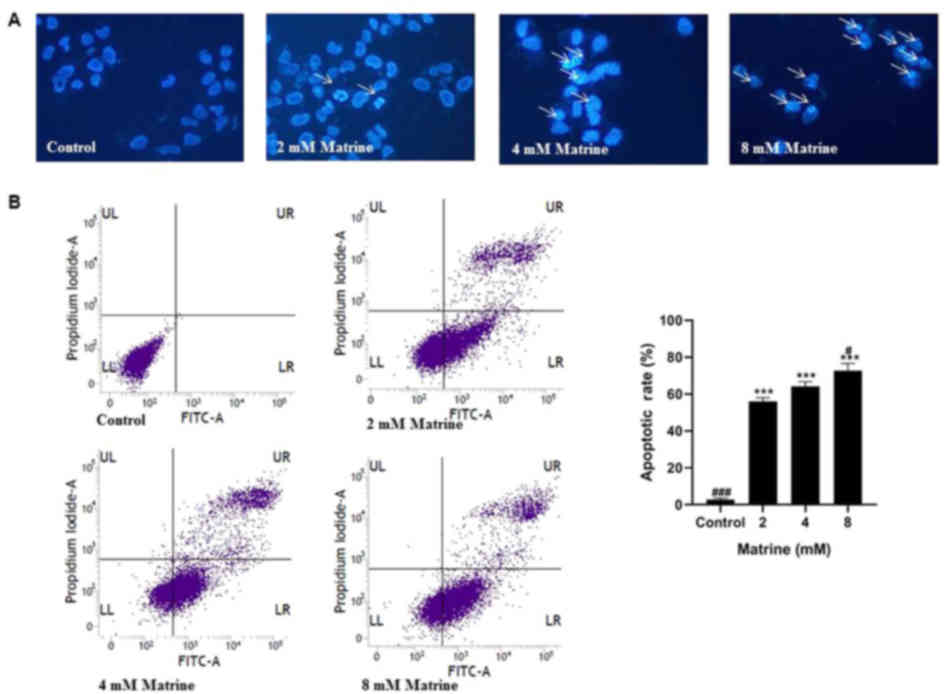
Matrine induces cell apoptosis in
MCF-7 cells
To evaluate the effect of matrine on the apoptosis
of breast cancer MCF-7 cells, Hoechst 33342 staining, which is
sensitive to DNA, was used to estimate the changes of nuclear
morphology. As revealed in Fig.
4A, Hoechst 33342 staining of the control MCF-7 cells exhibited
a uniformly blue dyed pattern in the nuclei whereas the cells
incubated with matrine demonstrated typical characteristics of
apoptosis, namely nuclear condensation/fragmented chromatin and
apoptotic bodies with bright blue fluorescent dots. These data
indicated that matrine could effectively induce cellular apoptosis.
Apoptotic responses were also assessed by flow cytometry following
Annexin V/PI staining. As is revealed in Fig. 4B, exposure to matrine led to a
significant increase in the percentage of apoptotic cells in MCF-7
cells compared with the control. The apoptotic rates were
56.04±2.00% (2 mM), 64.28±2.68% (4 mM) and 72.81±3.83% (8 mM),
respectively. These results further confirmed that matrine could
induce cellular apoptosis. Based on these findings, matrine could
induce an apoptotic response in MCF-7 cells.
Matrine induces autophagy in MCF-7
cells
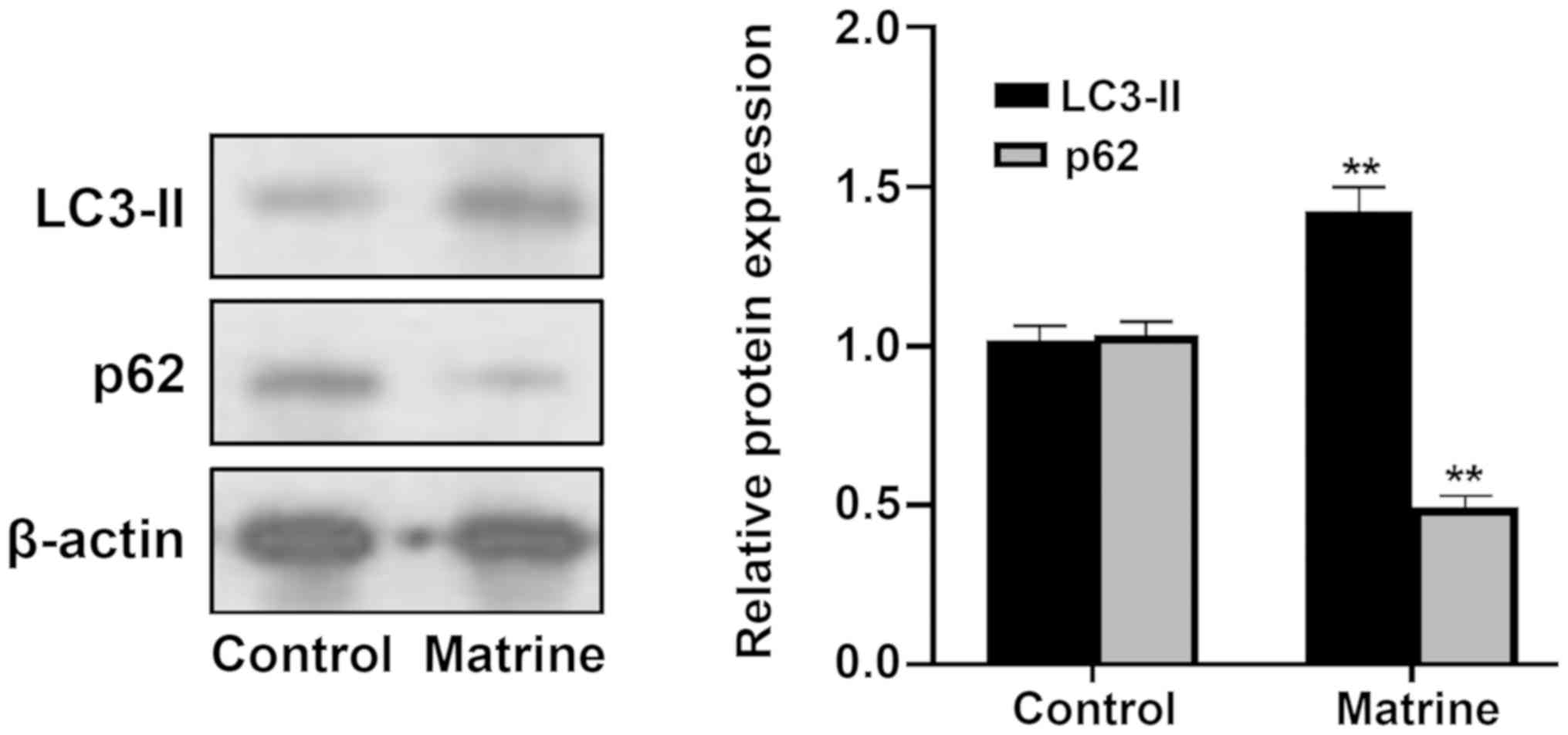
To determine the effect of matrine on the induction
of autophagy, expression of LC3-II, a protein recruited to
autophagosomal membranes as an indication of autophagy (25), was assessed using western blotting.
As demonstrated in Fig. 5, in
MCF-7 cells, matrine enhanced LC3-II expression compared with the
control group, indicating that matrine induced autophagic flux. To
further determine whether matrine modulates autophagic degradation,
the expression levels of p62, also known as sequestosome-1, which
is associated with LC3-II turnover and functions as a marker of
autophagic degradation (26) were
assessed. MCF-7 cells incubated with matrine resulted in
significantly decreased expression levels of p62, indicating an
inductive effect of matrine on autophagic flux. Collectively, these
results indicated that matrine was an inducer of autophagy.
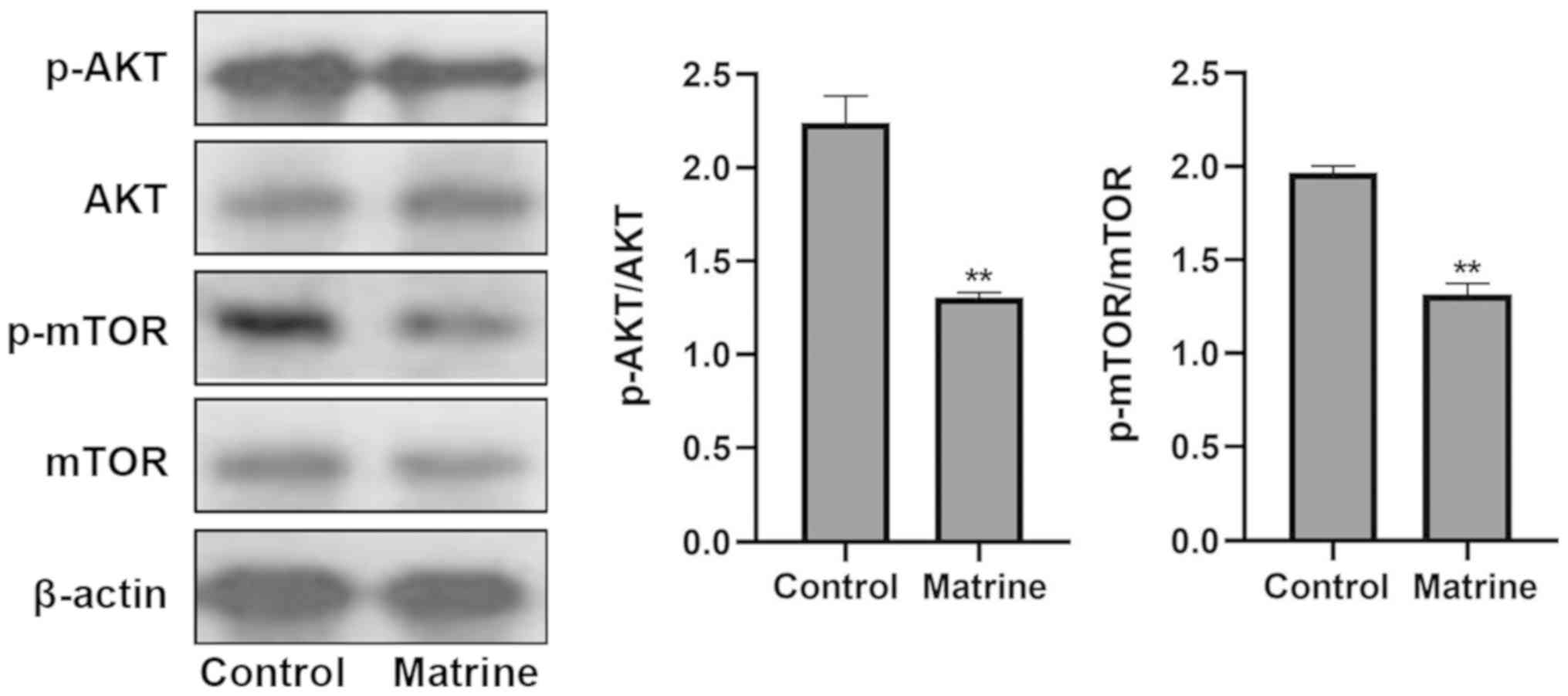
Matrine induces autophagy via the
AKT/mTOR pathway
Next, the pathways involving the activation of
autophagy induced by matrine analysis were explored using western
blotting. It was identified that, compared with the control group,
levels of phosphorylation of AKT and mTOR were significantly
downregulated in MCF-7 cells following incubation with matrine
(Fig. 6). Matrine treatment
significantly decreased the ratio of p-AKT/AKT in MCF-7 cells.
Variation in the p-mTOR/mTOR ratio was consistent with the change
in the p-AKT/AKT ratio. Collectively, these results indicated that
matrine-induced autophagy was due, at least in part, to inhibition
of the AKT/mTOR pathway.
Discussion
Breast cancer is the commonest type of malignancy in
females worldwide (27). Recently,
phytochemicals have become a substantial source of cancer
treatments (28). Matrine, an
alkaloid component extracted from the Chinese herb Sophora
flavescens, is considered to improve the quality of life and
prognosis of cancer patients when combined with standard therapies
(29). Matrine exhibits
pro-apoptotic (30),
anti-proliferation (31) and
metastasis-suppression effects (32,33)
in various cancer cells such as SGC-7901 human gastric cancer cell
and osteosarcoma cells (12,13).
However, the mechanisms underlying the anticancer properties of
matrine in MCF-7 breast cancer cells remain to be elucidated.
Therefore, the potential mechanism of matrine against MCF-7 breast
cancer cells was explored in the present study.
To determine the role of matrine in MCF-7 breast
cancer cells, CCK-8 assay was performed. Consistent with previous
studies (6,34), matrine significantly suppressed
cell viability in a dose- and time-dependent manner in MCF-7 breast
cancer cells. Furthermore, the morphological changes of the MCF-7
breast cancer cells cultured in the presence of matrine also
revealed its cytotoxic effect on MCF-7 cells. Apoptosis induction
serves a key role in cancer therapy (35). In the present study, Hoechst 33342
staining and flow cytometry revealed that matrine could cause
cellular apoptosis in MCF-7 cells. These findings supported the
hypothesis that inhibition of cellular growth in MCF-7 by matrine
was at least partly attributed to its induction of apoptosis.
Autophagy is closely associated with cancer
therapeutics (36). Previous
studies have revealed that autophagy enhancers can be used as
potential treatment strategies for cancer in precancerous lesions
(37,38). In advanced cancer, intervening in
autophagy has been suggested as one of the effective measures for
cancer treatment (39,40). Resveratrol has been revealed to
inhibit the proliferation of A549 lung cancer cells and to promote
cell death by an autophagic mechanism (41). Juglone conferred protection in
human hepatocellular carcinoma cells, mediated by its ability to
induce apoptosis and autophagy by adjustment of mitogen-activated
protein kinase pathways (42).
Recently, several lines of evidence have also demonstrated that
matrine has potential anticancer properties by enhancing autophagy.
Matrine can induce apoptosis and autophagy of glioma cell line U251
via modulation of circular RNA-104075/BCL-9 (43) and induced protective autophagy via
ERK activation in osteosarcoma cells (30). However, there is insufficient
evidence to determine the role of matrine interaction between
autophagy and breast cancer. To the best of the authors' knowledge,
the present study is the first time to identify matrine as an
inducer of autophagy in breast cancer MCF-7 cells. LC3, a cytosolic
ubiquitin-like protein, is expressed in the majority of cell types
(44). During autophagosome
formation, cytosolic LC3-I is transformed into its enzymatic LC3-II
counterpart that aggregates on membranes of autophagosomes
(45); p62, a ubiquitin-binding
scaffold protein, has an LC3-interacting domain and is degraded
during autophagy progression (46). LC3-II and p62 are therefore
reliable autophagy markers. Thus, to determine the role of matrine
in autophagy induction, expression levels of LC3-II and p62 were
assessed using western blotting. The results demonstrated that
matrine upregulated the expression of LC3-II and concomitantly
decreased p62 expression. These results indicated that the
cytotoxic effects of matrine in breast cancer MCF-7 cells may at
least in part associated with the induction of autophagy.
Autophagy is regulated by a number of upstream
signaling pathways (47). The
AKT/mTOR pathway is a crucial signaling cascade that regulates
numerous cellular processes in both normal and cancer cells,
including cell proliferation, the cell cycle, synthesis of protein
and angiogenesis (48). mTOR is
phosphorylated by activated AKT and p-mTOR negatively regulates
autophagy (49). A previous study
suggested that the AKT/mTOR signaling pathway serves a vital role
in autophagy (50). Inhibition of
the AKT/mTOR pathway was revealed to promote autophagy and
apoptosis to produce anticancer effects (51). Triptolide suppressed tumor growth
by inducing apoptosis and autophagy by activating the ROS/JNK and
blocking the AKT/mTOR signaling pathways in glioma cells (52). Curcumin demonstrated its antitumor
activity by regulating autophagy in human lung cancer A549 cells by
inhibiting the PI3K/AKT/mTOR pathway (53). The present study determined whether
the AKT/mTOR signaling pathway could be responsible for the
autophagy upregulation efficacy of matrine. Matrine treatment
significantly inhibited the phosphorylation of AKT and mTOR in
breast cancer MCF-7 cells. Collectively, the data from the present
study indicated that the AKT/mTOR signaling pathway participated in
matrine-induced apoptosis and protective autophagy in breast cancer
MCF-7 cells. The present study demonstrated that, in breast cancer
MCF-7 cells, matrine could suppress cellular growth by triggering
apoptosis and autophagy, both of which are regulated by the
AKT/mTOR signaling pathway. Consequently, these findings indicated
that matrine has a potential application in the treatment of human
breast cancer. However, the several factors involved in these
events are complex processes that require further characterization.
Detailed in vivo investigations into the therapeutic
benefits of matrine should be pursued.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Guangdong
Natural Science Foundation (grant nos. 2018A030313084 and
2018A030310112), the Administration of Traditional Chinese Medicine
of Guangdong Province of China (grant nos. 20181150 and 20191187),
the ‘Group-type’ Special Support Project for Education Talents in
Universities (grant no. 4SG19045G), Public Service Platform Open
Project Fund of South China Sea for R&D Marine Biomedicine
Resources (grant no. 2HC18010) and the Shenzhen Bao'an Science and
Technique Project (grant no. 2019JD214).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JD, LinL and LiL conceived the study. BL and LiL
designed the study. JL, DS, QL and LiL performed the experiments.
JL and LiL analyzed the data. JD and LiL wrote the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 60:277–300. 2018.
|
|
2
|
Yang LH, Ho YJ, Lin JF, Yeh CW, Kao SH and
Hsu LS: Butein inhibits the proliferation of breast cancer cells
through generation of reactive oxygen species and modulation of ERK
and p38 activities. Mol Med Rep. 6:1126–1132. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Du J, Li L and Zhou S: Microbial
production of cyanophycin: From enzymes to biopolymers. Biotechnol
Adv. 37:1074002019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Du J, Yang D, Luo ZW and Lee SY: Metabolic
engineering of Escherichia coli for the production of indirubin
from glucose. J Biotechnol. 267:19–28. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Brower V: Back to nature: Extinction of
medicianl plants threathens drug discovery. J Natl Cancer Inst.
100:838–839. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou BG, Wei CS, Zhang S, Zhang Z and Gao
HM: Matrine reversed multidrug resistance of breast cancer
MCF-7/ADR cells through PI3K/AKT signaling pathway. J Cell Biochem.
119:3885–3891. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang YB, Zhan LQ, Li GQ, Wang F, Wang Y,
Li YL, Ye WC and Wang GC: Dimeric matrine-type alkaloids from the
roots of Sophora flavescens and their anti-hepatitis B virus
activities. J Org Chem. 81:6273–6280. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Y, Zhang H, Yu P, Liu Q, Liu K, Duan
H, Luan G, Yagasaki K and Zhang G: Effects of matrine against the
growth of human lung cancer and hepatoma cells as well as lung
cancer cell migration. Cytotechnology. 59:191–200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y, Cui L, Guan G, Wang J, Qiu C,
Yang T, Guo Y and Liu Z: Matrine suppresses cardiac fibrosis by
inhibiting the TGF-β/Smad pathway in experimental diabetic
cardiomyopathy. Mol Med Rep. 17:1775–1781. 2018.PubMed/NCBI
|
|
10
|
Zhang P, Wang Z, Chong T and Ji Z: Matrine
inhibits proliferation and induces apoptosis of the
androgen-independent prostate cancer cell line PC-3. Mol Med Rep.
5:783–787. 2012.PubMed/NCBI
|
|
11
|
Zhou N, Li J, Li T, Chen G, Zhang Z and Si
Z: Matrine-induced apoptosis in Hep3B cells via the inhibition of
MDM2. Mol Med Rep. 15:442–450. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang J and Xu H: Matrine: Bioactivities
and structural modifications. Curr Top Med Chem. 16:3365–3378.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yong J, Wu X and Lu C: Anticancer advances
of matrine and its derivatives. Curr Pharm Des. 21:3673–3680. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xue S, Zhou Y, Zhang J, Xiang Z, Liu Y,
Miao T, Liu G, Liu B, Liu X, Shen L, et al: Anemoside B4 exerts
anti-cancer effect by inducing apoptosis and autophagy through
inhibiton of PI3K/Akt/mTOR pathway in hepatocellular carcinoma. Am
J Transl Res. 11:2580–2589. 2019.PubMed/NCBI
|
|
15
|
Guo Z, Guozhang H, Wang H, Li Z and Liu N:
Ampelopsin inhibits human glioma through inducing apoptosis and
autophagy dependent on ROS generation and JNK pathway. Biomed
Pharmacother. 116:1085242019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu J, Fan L, Wang H and Sun G: Autophagy,
a double-edged sword in anti-angiogenesis therapy. Med Oncol.
33:102016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kaminskyy VO, Piskunova T, Zborovskaya IB,
Tchevkina EM and Zhivotovsky B: Suppression ofbasal autophagy
reduces lung cancer cell proliferation and
enhancescaspase-dependent and -independent apoptosis by stimulating
ROS formation. Autophagy. 8:1032–1044. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Leidal AM, Levine B and Debnath J:
Autophage and the cell biology of age-related disease. Nat Cell
Biol. 20:1338–1348. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Maycotte P, Gearheart CM, Barnard R, Aryal
S, Mulcahy Levy JM, Fosmire SP, Hansen RJ, Morgan MJ, Porter CC,
Gustafson DL and Thorburn A: STAT3-mediated autophagy dependence
identifies subtypes of breast cancer where autophagy inhibition can
be efficacious. Cancer Res. 74:2579–2590. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lefort S, Joffre C, Kieffer Y, Givel AM,
Bourachot B, Zago G, Bieche I, Dubois T, Meseure D, Vincent-Salomon
A, et al: Inhibition of autophagy as a new means of improving
chemotherapy efficiency in high-LC3B triple-negative breast
cancers. Autophagy. 10:2122–2142. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cook KL, Wärri A, Soto-Pantoja DR, Clarke
PA, Cruz MI, Zwart A and Clarke R: Hydroxychloroquine inhibits
autophagy to potentiate antiestrogen responsiveness in ER+ breast
cancer. Clin Cancer Res. 20:3222–3232. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yan W, Ma X, Zhao X and Zhang S: Baicalein
induces apoptosis and autophagy of breast cancer cells via
inhibiting PI3K/AKT pathway in vivo and vitro. Drug Des Devel Ther.
12:3961–3972. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tong Y, Zhang G, Li Y, Xu J, Yuan J, Zhang
B, Hu T and Song G: Corilagin inhibits breast cancer growth via
reactive oxygen species-dependent apoptosis and autophagy. J Cell
Mol Med. 22:3795–3807. 2018. View Article : Google Scholar
|
|
24
|
Xiao X, Ao M, Xu F, Li X, Hu J, Wang Y, Li
D, Zhu X, Xin C and Shi W: Effect of matrine against breast cancer
by downregulating the vascular endothelial growth factor via the
Wnt/β-catenin pathway. Oncol Lett. 15:1691–1697. 2018.PubMed/NCBI
|
|
25
|
Xie Z, Xie Y, Xu Y, Zhou H, Xu W and Dong
Q: Bafilomycin A1 inhibits autophagy and induces apoptosis in MG63
osteosarcoma cells. Mol Med Rep. 10:1103–1107. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yue Z: Regulation of neuronal autophagy in
axon: Implication of autophagy in axonal function and
dysfunction/degeneration. Autophagy. 3:139–141. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancerstatistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Efferth T, Konkimalla VB, Wang YF,
Sauerbrey A, Meinhardt S, Zintl F, Mattern J and Volm M: Prediction
of broad spectrum resistance of tumors towards anticancer drugs.
Clin Cancer Res. 14:2405–2412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Niu H, Zhang Y, Wu B, Zhang Y, Jiang H and
He P: Matrine induces the apoptosis of lung cancer cells through
downregulation of inhibitor of apoptosis proteins and the Akt
signaling pathway. Oncol Rep. 32:1087–1093. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma K, Huang MY, Guo YX and Hu GQ:
Matrine-induced autophagy counteracts cell apoptosis via the ERK
signaling pathway in osteosarcoma cells. Oncol Lett. 12:1854–1860.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang W, Hosford SR, Traphagen NA, Shee K,
Demidenko E, Liu S and Miller TW: Autophagy promotes escape from
phosphatidylinositol 3-kinase inhibition in estrogen
receptor-positive breast cancer. FASEB J. 32:1222–1235. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu P, Liu Q, Liu K, Yagasaki K, Wu E and
Zhang G: Matrine suppresses breast cancer cell proliferation and
invasion via VEGF-Akt-NF-kappaB signaling. Cytotechnology.
59:219–229. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li H, Tan G, Jiang X, Qiao H, Pan S, Jiang
H, Kanwar JR and Sun X: Therapeutic effects of matrine on primary
and metastatic breast cancer. Am J Chin Med. 38:1115–1130. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li H, Li X, Bai M, Suo Y, Zhang G and Cao
X: Matrine inhibited proliferation and increased apoptosis in human
breast cancer MCF-7 cells via upregulation of Bax and
downregulation of Bcl-2. Int J Clin Exp Pathol. 8:14793–14799.
2015.PubMed/NCBI
|
|
35
|
Tsuji T, Ozasa H, Aoki W, Aburaya S,
Yamamoto Funazo T, Furugaki K, Yoshimura Y, Yamazoe M, Ajimizu H,
Yasuda Y, et al: YAP1 mediates survival of ALK-rearranged lung
cancer cells treated with alectinib via pro-apoptotic protein
regulation. Nat Commun. 11:742020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang S, Wang K, Wang H, Han J and Sun H:
Autophagy is essential for flavopiridol-induced cytotoxicity
against MCF-7 breast cancer cells. Mol Med Rep. 16:9715–9720. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Poillet-Perez L and White E: Role of tumor
and host autophagy in cancer metabolism. Genes Dev. 33:610–619.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen J, Zhang L, Zhou H, Wang W, Luo Y,
Yang H and Yi H: Inhibition of autophagy promotes cisplatin-induced
apoptotic cell death through Atg5 and Beclin 1 in A549 human lung
cancer cells. Mol Med Rep. 17:6859–6865. 2018.PubMed/NCBI
|
|
39
|
Vidoni C, Ferraresi A, Secomandi E,
Vallino L, Dhanasekaran DN and Isidoro C: Epigenetic targeting of
autophagy for cancer prevention and treatment by natural compounds.
Semin Cancer Biol. 2019.(Epub ahead of print). PubMed/NCBI
|
|
40
|
Udristioiu A and Nica-Badea D: Autophagy
dysfunctions associated with cancer cells and their therapeutic
implications. Biomed Pharmacother. 115:1088922019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang J, Li J, Cao N, Li Z, Han J and Li L:
Resveratrol, an activator of SIRT1, induces protective autophagy in
non-small-cell lung cancer via inhibiting Akt/mTOR and activating
p38-MAPK. Onco Targets Ther. 11:7777–7786. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang P, Gao C, Wang W, Yao LP, Zhang J,
Zhang SD, Li J, Fang SH and Fu YJ: Juglone induces apoptosis and
autophagy via modulation of mitogen-activated protein kinase
pathways in human hepatocellular carcinoma cells. Food Chem
Toxicol. 116:40–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chi G, Xu D, Zhang B and Yang F: Matrine
induces apoptosis and autophagy of glioma cell line U251 by
regulation of circRNA-104075/BCL-9. Chem Biol Interact.
308:198–205. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Saha S, Panigrahi DP, Patil S and Bhutia
SK: Autophagy in health and disease: A comprehensive review. Biomed
Pharmacother. 104:485–495. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tazawa H, Kuroda S, Hasei J, Kagawa S and
Fujiwara T: Impact of autophagy in oncolytic adenoviral therapy for
cancer. Int J Mol Sci. 18:14792017. View Article : Google Scholar
|
|
46
|
Cohen-Kaplan V, Livneh I, Avni N, Fabre B,
Ziv T, Kwon YT and Ciechanover A: p62- and ubiquitin-dependent
stress-induced autophagy of the mammalian 26S proteasome. Proc Natl
Acad Sci USA. 113:E74902016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Chen Y, Zhao X, Li J, Zhang L, Li R, Zhang
H, Liao R, Liu S, Shi W and Liang X: Amino acid starvation promotes
podocyte autophagy through mammalian target of rapamycin inhibition
and transcription factor EB activation. Mol Med Rep. 18:4342–4348.
2018.PubMed/NCBI
|
|
48
|
Heras-Sandoval D, Pérez-Rojas JM,
Hernández-Damián J and Pedraza-Chaverri J: The role of
PI3K/AKT/mTOR pathway in the modulation of autophagy and the
clearance of protein aggregates in neurodegeneration. Cell Signal.
26:2694–2701. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Zhang XR, Wang SY, Sun W and Wei C:
Isoliquiritigenin inhibits proliferation and metastasis of MKN28
gastric cancer cells by suppressing the PI3K/AKT/mTOR signaling
pathway. Mol Med Rep. 18:3429–3436. 2018.PubMed/NCBI
|
|
50
|
Yu H, Qiu Y, Pang X, Li J, Wu S, Yin S,
Han L, Zhang Y, Jin C, Gao X, et al: Lycorine promotes autophagy
and apoptosis via TCRP1/Akt/mTOR axis inactivation in human
hepatocellular carcinoma. Mol Cancer Ther. 16:2711–2723. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Hu JL, Hu XL, Guo AY, Wang CJ, Wen YY and
Cang SD: Endoplasmic reticulum stress promotes autophagy and
apoptosis and reverses chemoresistance in human ovarian cancer
cells. Oncotarget. 8:49380–49394. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liu X, Zhao P, Wang X, Wang L, Zhu Y and
Gao W: Triptolide induces glioma cell autophagy and apoptosis via
upregulating the ROS/JNK and downregulating the Akt/mTOR signaling
pathways. Front Oncol. 9:3872019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Liu F, Gao S, Yang Y, Zhao X, Fan Y, Ma W,
Yang D, Yang A and Yu Y: Antitumor activity of curcumin by
modulation of apoptosis and autophagy in human lung cancer A549
cells through inhibiting PI3K/Akt/mTOR pathway. Oncol Rep.
39:1523–1531. 2018.PubMed/NCBI
|