Introduction
Intervertebral disc degeneration (IDD) is a
degenerative disease of the spine originating from the
intervertebral disc, leading to instability of the spine, disc
herniation, spinal stenosis and cervical spondylosis. IDD is one of
the most important causes of musculoskeletal disability and leads
to the wide presence of motor dysfunction in the population
worldwide (1).
Since the pathogenesis of IDD is not fully
understood, current treatments are limited and tend to focus on
pain relief rather than inhibiting disease progression. The main
pathological changes in IDD are nucleus pulpocyte apoptosis and
excessive degradation of extracellular matrix (ECM) (2). The expression of type II collagen and
proteoglycan, which are originally expressed in normal nucleus
pulposus tissue, is upregulated, while the expression of type I
collagen is increased, during intervertebral disc degeneration
(3). Type I collagen was
significantly different due to its biomechanical characteristics
(4). The increase in content of
ECM leads to changes in the biomechanical properties of the disc,
which can cause the development of IDD (5).
A number of studies have been conducted on the role
of microRNAs (miRNAs or miRs) in various diseases (6–8).
miRNAs are a general term for a class of small-molecule non-coding
RNAs that are 20–22 nucleotides in length, as opposed to
mRNA-transcribed proteins. miRNAs do not encode proteins, but
inhibit the expression of multiple target genes by binding to the
3′-untranslated region (3′-UTR) of target mRNAs (9–11).
There is increasing evidence that miRNAs may be involved in the
development and progression of various diseases, such as
cardiovascular disease, cancer and autoimmune diseases (6–8).
Previous study has demonstrated that miR-25-3p can regulate the
proliferation and apoptosis of cancer cells in a variety of cancer
types and is closely associated with the degradation of human
nucleus pulposus cells (12).
Bcl-2 interacting mediator of cell death (Bim) is a
member of the BH3 subfamily of the Bcl-2 family (13). It is an important regulatory
protein of apoptosis and is stable in the homeostasis of
hematopoietic cells, preventing autoimmunity and tumorigenesis
(14). Bim is widely expressed in
normal cells and exists in a variety of isomers (15). Certain apoptotic stimuli can
activate Bim molecules through various signaling pathways (16). Activated Bim molecules activate Bax
through interaction with Bcl-2/Bax, causing apoptosis via the
mitochondrial pathway (17). Bim
is closely associated with the development and treatment of
autoimmune diseases, degenerative diseases and tumors (18). Therefore, basic research into the
role of Bim in apoptosis should provide the theoretical basis and
insights for clinical treatment.
The present study aimed to investigate the role of
miR-25-3p in the pathogenesis of IDD and to explore the underlying
mechanism.
Materials and methods
IDD rat model establishment
According to a previous study, a total of 20 healthy
male Wistar rats (300–350 g; aged 14–16 weeks; Model Animal
Research Center Of Nanjing University, Nanjing, China) were
selected. All rats were housed under standard conditions at room
temperature (22–24°C) and humidity (60–65%) on a 12-h light/dark
cycle with ad libitum supply of food and water. A rat IDD
model was established by puncture method (19). First, the rats were anesthetized
with an intraperitoneal injection of 3% sodium pentobarbital (40
mg/kg). Following anesthesia, the limbs were fixed and placed on
the operating table. X-ray angiography of the rats was performed
using a Faxitron instrument (Faxitron X-ray Corporation) to
identify the segments of the lumbar vertebrae. Then, according to
the angiographic results, the 3–4 lumbar intervertebral discs of
the rats were selected. After determining the position, the needle
of the lumbar vertebra (coccygeal intervertebral levels Co6-7 and
Co8-9) was pierced with a 20-ml needle to cause degeneration of the
intervertebral disc by mechanical damage. After 2 weeks, the extent
of disc degeneration in the experimental rats was determined using
magnetic resonance imaging. The health and behavior of all rats
were monitored every 2 days. No rats died during the experiments.
Experiment was ended when the rats lost >15% of their body
weight. Rats were anesthetized with pentobarbital (40 mg/kg) and
then sacrificed by cervical dislocation, with death defined as the
lack of heartbeat and breathing. All experimental procedures were
performed in accordance with the Recommended Guideline for the Care
and Use of Laboratory Animals issued by the Chinese Council on
Animal Research (20). The current
study was approved by the Animal Ethics Committee of the No. 903
Hospital of People's Liberation Army (approval no. IRB
SOP/01.03/01.1).
Isolation and culture of primary
degenerative nucleus pulposus (NP) cells
After the IDD model was established successfully,
the model and control rats were anesthetized with pentobarbital by
intraperitoneal injection, their limbs were fixed and the
prosthesis was placed on the operating table. The skin and lumbar
vertebrae were cut, the cells were separated from NP tissue and
medium was added to prepare a single cell suspension. Then, the
cells were purified by differential adherent culture to obtain
primary degenerative NP cells. Following successful primary culture
in Dulbecco's modified Eagle's medium (DMEM, Invitrogen; Thermo
Fisher Scientific, Inc.) containing 10% fetal bovine serum (FBS,
Invitrogen; Thermo Fisher Scientific, Inc.) at 37°C for 24 h, the
cells were stained with hematoxylin and eosin at room temperature
for 15 min and morphological identification of NP cells was carried
out by observing the morphology and aggregation of the cells under
an inverted phase contrast microscope (IX51; Olympus
Corporation).
IDD in vitro cell model
establishment
To establish the IDD in vitro cell model,
normal NP cells were treated with 10 ng/ml interleukin (IL)-1β for
24 h. Untreated NP cells were used as control cells.
Cell transfection
Normal NP cells were transfected with a miR-25-3p
inhibitor (5′-UCAGACCGAGACAAGUGCAAUG-3′; Guangzhou RiboBio Co.,
Ltd.), the negative control (NC) of miR-25-3p inhibitor
(5′-CAGUACUUUUGUGUAGUACAA-3′; Guangzhou RiboBio Co., Ltd.),
miR-25-3p inhibitor + control-small interference (siRNA) (cat no.
sc-36869; Santa Cruz Biotechnology, Inc.), miR-25-3p inhibitor +
Bim-siRNA (cat no. sc-29802; Santa Cruz Biotechnology, Inc.),
miR-25-3p mimic (sense, 5′-CAUUGCACUUGUCUCGGUCUGA-3′ and
anti-sense, 5′-UCAGACCGAGACAAGUGCAAUG-3′; Santa Cruz Biotechnology,
Inc.), mimic control (sense, 5′-UUUGUACUACACAAAAGUACUG-3′ and
anti-sense, 5′-CAGUACUUUUGUGUAGUACAAA-3′; Guangzhou RiboBio Co.,
Ltd.), miR-25-3p mimic + control-plasmid (cat no. sc-437275; Santa
Cruz Biotechnology, Inc.) or miR-25-3p mimic + Bim-plasmid (cat no.
sc-419332-ACT; Santa Cruz Biotechnology, Inc.) using Lipofectamine
2000® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) for 48 h following the manufacturer's protocol. At 48 h
post-transfection, the transfection efficiency was detected using
reverse transcription-quantitative PCR (RT-qPCR).
RT-qPCR
Cell total RNA (5×106 cells) was
extracted using the TRIzol® (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
total RNA concentration was detected by NanoDrop 2000
spectrophotometer (NanoDrop Technologies; Thermo Fisher Scientific,
Inc.). Total RNA was stored at −80°C for further use. The synthesis
of cDNA was carried out with the RevertAid™ First Strand cDNA
Synthesis kit (Thermo Fisher Scientific, Inc.). qPCR was performed
using the cDNA by SYBR RT-PCR kit (Takara Bio, Inc.) according to
the manufacturer's protocol. qPCR was performed as follows: 10 min
at 95°C, followed by 35 cycles of 15 sec at 95°C and 40 sec at
55°C. The primer sequences used for RT-qPCR were: miR-25-3p,
forward, 5′-CATTGCACTTGTCTCGGTCTGA-3′ and reverse,
5′-GCTGTCAACGATACGCTACGTAACG-3′; U6, forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′;
Bim, forward, 5′-CACAAACCCCAAGTCCTCCT-3′ and reverse,
5′-ACACCAGGCGGACAATGTAA-3′; caspase-3, forward,
5′-TGTCGATGCAGCAAACCTCA-3′ and reverse,
5′-GACTTCTACAACGATCCCCTC-3′; Bax, forward,
5′-CGTCCACCAAGAAGCTGAGCG-3′ and reverse, 5′-CGTCCACCAAAGCTGAGCG-3′;
Bcl-2, forward, 5′-TTGGATCAGGGAGTTGGAAG-3′ and reverse,
5′-TGTCCCTACCAACCAGAAGG-3′; SOX-9, forward,
5′-GTACCCGCACTTGCACAAC-3′ and reverse, 5′-TCGCTCTCGTTCAGAAGTCTC-3′;
proteoglycan (ACAN), forward, 5′-TTGTGACTCTGCGGGTCATC-3′ and
reverse, 5′-GTCCCTAGGAGGGCCTTCAG-3′; collagen I, forward,
5′-GGCGGCCAGGGCTCCGACCC-3′ and reverse, 5′-AATTCCTCGTCTGGGGCACC-3′;
collagen II, forward, 5′-ATGGCGGCTTCCACTTCAG-3′ and reverse,
5′-CGGTGGCTTCATCCAGGTAG-3′; and GAPDH, forward,
5′-TGCACCACCAACTGCTTAGC-3′ and reverse,
5′-GGCATGGACTGTGGTCATGAG-3′. Relative expression levels were
calculated using the 2−ΔΔCq method (21) following normalization to the
expression of GAPDH or U6. All experiments were performed in
triplicate.
Western blot assay
NP cells were washed 3 times with pre-cooled PBS and
total cellular protein was extracted using a modified RIPA buffer
(Beyotime Institute of Biotechnology) containing 1 mM PMSF for 30
min. A BCA protein quantitative kit (Thermo Fisher Scientific,
Inc.) was applied to detect protein concentration. The proteins (40
µg per lane) were separated by 10% SDS-PAGE and transferred to PVDF
membranes. The membranes were blocked with 5% non-fat milk at room
temperature for 2 h, followed by incubation with primary antibodies
against Bcl-2 (cat no. 4223; 1:1,000; Cell Signaling Technology,
Inc.), Bax (cat no. 5023; 1:1,000; Cell Signaling Technology,
Inc.), Bim (cat no. 2933; 1:1,000; Cell Signaling Technology,
Inc.), cleaved caspase-3 (cat no. 9661; 1:1,000; Cell Signaling
Technology, Inc.), pro-caspase-3 (cat no. ab183179; 1:1,000;
Abcam), caspase-3 (cat no. 9662; 1:1,000; Cell Signaling
Technology, Inc.), SOX-9 (cat no. ab185230; 1:1,000; Abcam), ACAN
(cat no. ab36861; 1:1,000; Abcam), collagen I (cat no. ab34710;
1:1,000; Abcam), collagen II (cat no. ab239007; 1:1,000; Abcam) and
GAPDH (cat no. 5174; 1:1,000; Cell Signaling Technology, Inc.)
overnight for 4°C. The membrane was then incubated for 2 h at room
temperature with a secondary antibody (horseradish
peroxidase-conjugated anti-rabbit immunoglobulin G; cat no. 7074;
1:2,000; Cell Signaling Technology, Inc.). Proteins were detected
using SignalFire™ enhanced chemiluminescence reagent (cat. no.
6883; Cell Signaling Technology, Inc.) and imaged. GAPDH was used
as an internal control. Band densities were quantified using
Gel-Pro Analyzer densitometry software (version 6.3; Media
Cybernetics, Inc.).
MTT assay
MTT assay was performed to detect cell
proliferation. Following treatment, the cells were inoculated in a
96-well plate with 1×104 cells per well and incubated in
a 37°C, 5% CO2 incubator. Then, 20 µl 5 mg/ml MTT agent
(Sigma-Aldrich; Merck KGaA) was added to cells. Following
incubation for 4 h, the formazan crystals were dissolved in 150 µl
DMSO (Sigma-Aldrich; Merck KGaA). The absorbance was measured at a
wavelength of 490 nm using a microplate reader.
Flow cytometry assay
The cells were collected in logarithmic growth phase
by trypsinization, washed 3 times with PBS and trypsinized into
single cell suspensions. Apoptotic cells were detected using the
Annexin V FITC/PI apoptosis detection kit (BD Biosciences; Becton,
Dickinson and Company) according to the manufacturer's protocol.
The cells were stained with Annexin V-FITC and propidium iodide
(PI) for 15 min in the dark at room temperature. Finally, flow
cytometry (BD Biosciences; Becton, Dickinson and Company) was used
to detect cell apoptosis and the data were analyzed using FlowJo
software (version 7.6.1; FlowJo LLC).
Dual luciferase reporter assay
Bioinformatics software (TargetScan 7.2, http://www.targetscan.org/vert_72/) was used to
predict the target gene of miR-25-3p. The results revealed the
binding sites between the 3′-UTR of Bim and miR-25-3p. To confirm
the association between miR-25-3p and Bim, a luciferase reporter
containing the 3′-UTR sequence of Bim was constructed using
pmiR-RB-Report™ Dual-Luciferase reporter gene plasmid vector
(Guangzhou RiboBio Co., Ltd.) according to the manufacturer's
protocol. Cells seeded in 24-well plates were cotransfected with
miR-25-3p mimic or mimic control and the mutant (MUT) or wild type
(WT) 3′-UTR of Bim using Lipofectamine 2000®
(Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h, together
with Renilla luciferase pRL-TK vector as a control.
Following transfection for 48 h, the cells were lysed with RIPA
buffer (Beyotime Institute of Biotechnology). The relative
luciferase activity was detected using the Dual-Luciferase Reporter
Assay System (Promega Corporation) according to the manufacturer's
protocol.
Statistical analysis
Every experiment was performed ≥3 times. All data
are shown as the mean ± standard deviation. The significance of
differences between two groups was measured using the Student's
t-test. Differences between multiple groups were detected using
one-way ANOVA followed by Tukey's test. Data analyses were
performed using GraphPad Prism 6.0 (GraphPad Software, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Expression of miR-25-3p and IDD
markers in rat NP cells
In order to detect the expression of IDD markers and
miR-25-3p in rat NP cells, RT-qPCR and/or western blotting were
performed to compare the expression levels of ACAN, collagen I,
collagen II, SOX-9 and miR-25-3p in degenerative and
non-degenerated NP cells. The RT-qPCR results demonstrated that
miR-25-3p was expressed reduced in rat degenerative NP cells
compared with that in normal NP cells (Fig. 1A). In addition, IDD-associated
markers, including ACAN, collagen I, collagen II and SOX-9 were
highly expressed in rat degenerative NP cells (Fig. 1B-F).
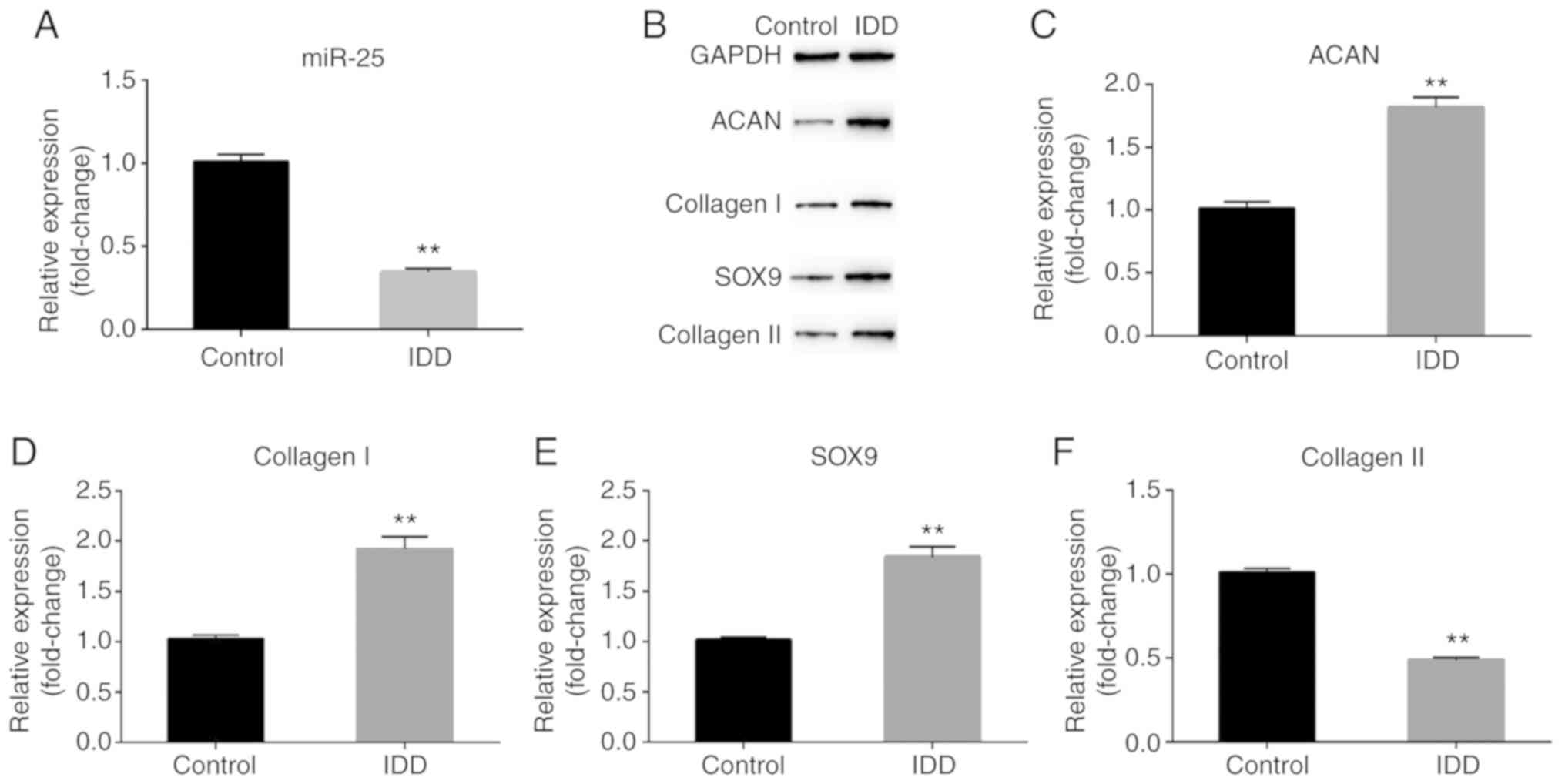 | Figure 1.Expression of miR-25-3p and IDD
markers in rat NP cells. (A) RT-qPCR assay detected the relative
expression of miR-25-3p. (B) Western blot assay detected the
protein expression of ACAN, collagen I, collagen II and SOX-9.
(C-F) RT-qPCR assay detected the relative mRNA expression of ACAN,
collagen I, SOX-9 and collagen II. Control, NP cells from control
rats; IDD, NP cells from rats with IDD. **P<0.01 vs. control.
miR, microRNA; IDD, intervertebral disc degeneration; RT-qPCR,
reverse transcription-quantitative PCR; NP, nucleus pulposus; ACAN,
proteoglycan. |
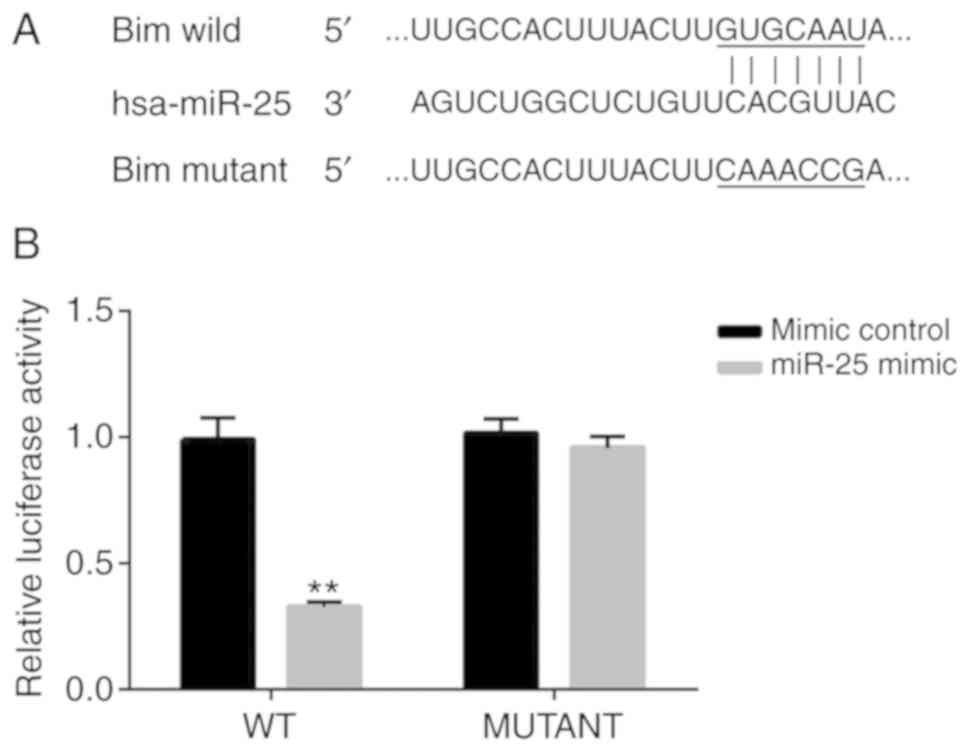
Bim is a direct target of
miR-25-3p
Next, to determine the targets of miR-25-3p,
TargetScan analysis was performed. TargetScan revealed that the
3′-UTR of Bim mRNA contains a putative site that is partially
complementary to miR-25-3p (Fig.
2A). Furthermore, a luciferase reporter assay was performed to
examine whether miR-25-3p interacted directly with the target gene
Bim. The luciferase activity in NP cells transfected with Bim-WT
and miR-25-3p mimic was decreased compared with that in NP cells
transfected with Bim-WT and mimic control. However, no significant
difference was observed in the luciferase activity of NP cells
transfected with Bim-MUT and miR-25-3p mimic and that of NP cells
transfected with Bim-MUT and mimic control (Fig. 2B). Taken together, these results
demonstrated that Bim was the direct target gene of miR-25-3p.
Effect of miR-25-3p downregulation on
the proliferation and apoptosis of normal NP cells
In order to evaluate the role of miR-25-3p
downregulation in normal NP cells, NP cells were transfected with
the miR-25-3p inhibitor, miR-25-3p inhibitor NC, inhibitor +
control-siRNA or inhibitor + Bim-siRNA. The results revealed that
the miR-25-3p inhibitor significantly decreased the level of
miR-25-3p in NP cells (Fig. 3A),
while Bim-siRNA significantly reduced the mRNA level of Bim in NP
cells (Fig. 3B). Then, the effect
of miR-25-3p on the proliferation of NP cells was explored. An MTT
assay indicated that when the cells were transfected with miR-25-3p
inhibitor, the cell viability was decreased compared with the
control (Fig. 3C). To further
determine the apoptotic effect of miR-25-3p, flow cytometry was
performed to detect cell apoptosis. Flow cytometry analysis
demonstrated that transfection with the miR-25-3p inhibitor
significantly induced NP cell apoptosis (Fig. 3D and E). All the effects of
miR-25-3p inhibitor on NP cells were reversed by Bim-siRNA.
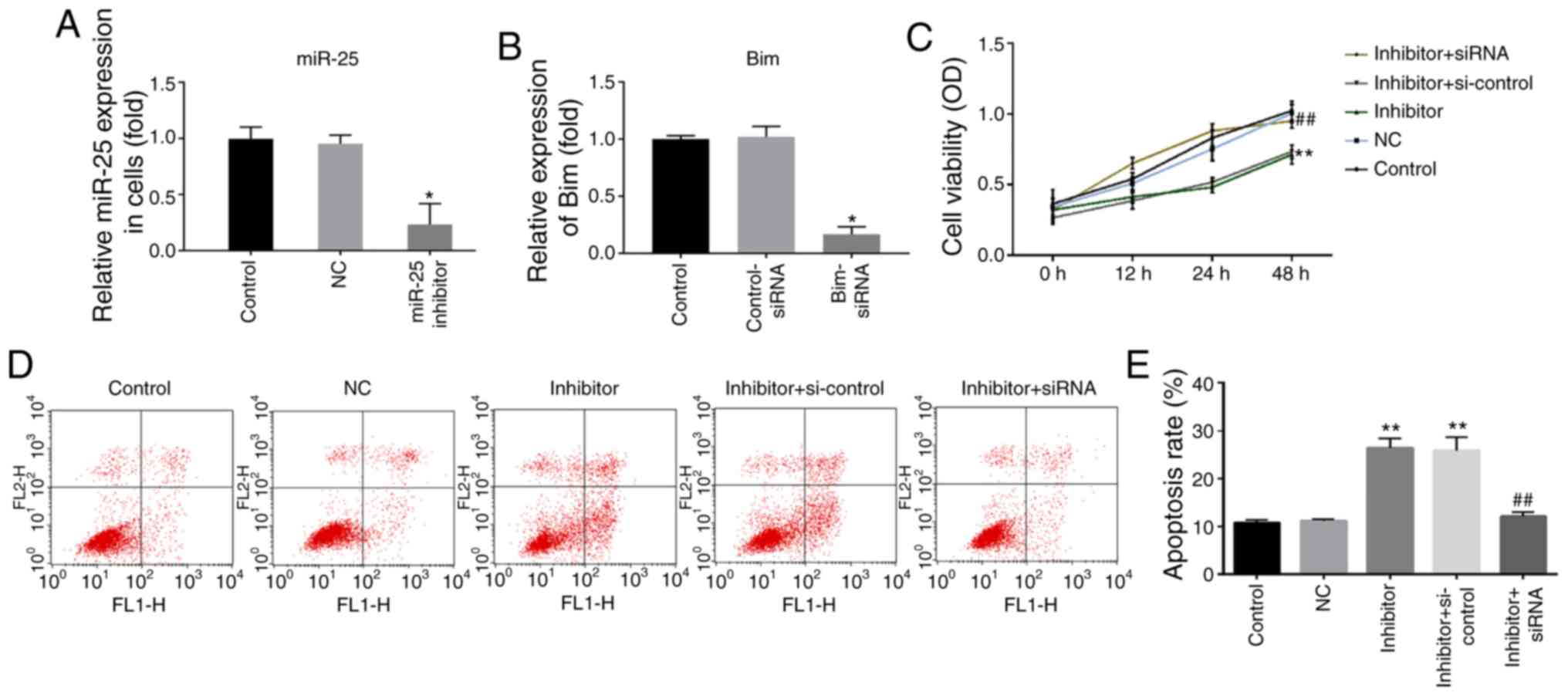 | Figure 3.Effect of miR-25-3p downregulation on
the proliferation and apoptosis of NP cells. (A) The level of
miR-25-3p in NP cells was detected using RT-qPCR. (B) The mRNA
level of Bim in NP cells was detected using RT-qPCR; (C) MTT assay
detected the cell viability of NP cells. (D) Flow cytometry assay
and (E) analysis measured the apoptosis of NP cells. Control, NP
cells without any treatment; NC, NP cells transfected with the NC
of miR-25-3p inhibitor; miR-25-3p inhibitor/inhibitor, NP cells
transfected with miR-25-3p inhibitor; control-siRNA, NP cells
transfected with control-siRNA; Bim-siRNA, NP cells transfected
with Bim-siRNA; inhibitor + si-control, NP cells transfected with
miR-25-3p inhibitor and control-siRNA; inhibitor + siRNA, NP cells
transfected with miR-25-3p inhibitor and Bim-siRNA. *P<0.05 and
**P<0.01 vs. control; ##P<0.01 vs. inhibitor +
si-control. miR, microRNA; NP, nucleus pulposus; RT-qPCR, reverse
transcription-quantitative PCR; Bim, Bcl-2 interacting mediator of
cell death; NC, negative control; si, small interfering; OD,
optical density. |
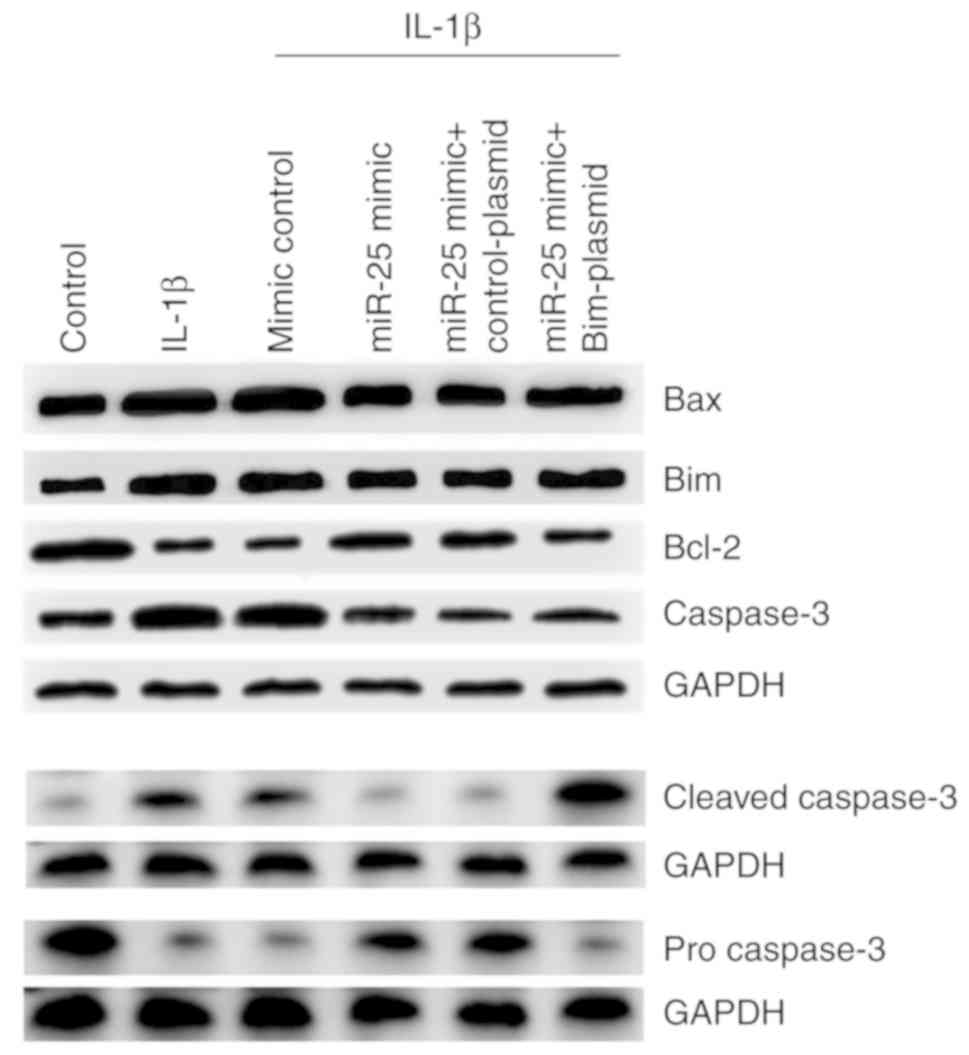
Effect of miR-25-3p downregulation on
the expression level of Bim and apoptosis-associated molecules in
normal NP cells
To further verify the role of miR-25-3p on NP cell
apoptosis, western blot assay and RT-qPCR were performed to detect
the expression of Bim and apoptosis-associated proteins (Bax,
Bcl-2, caspase-3, cleaved caspase-3 and pro-caspase-3). The results
demonstrated that miR-25-3p inhibitor significantly enhanced the
protein expression of Bim, Bax, cleaved caspase-3 and caspase-3,
while Bcl-2 and pro-caspase-3 protein expression was downregulated
(Fig. 4A). Similar results were
obtained with RT-qPCR (Fig. 4B-E).
All the effects of miR-25-3p inhibitor on NP cells were reversed by
Bim-siRNA.
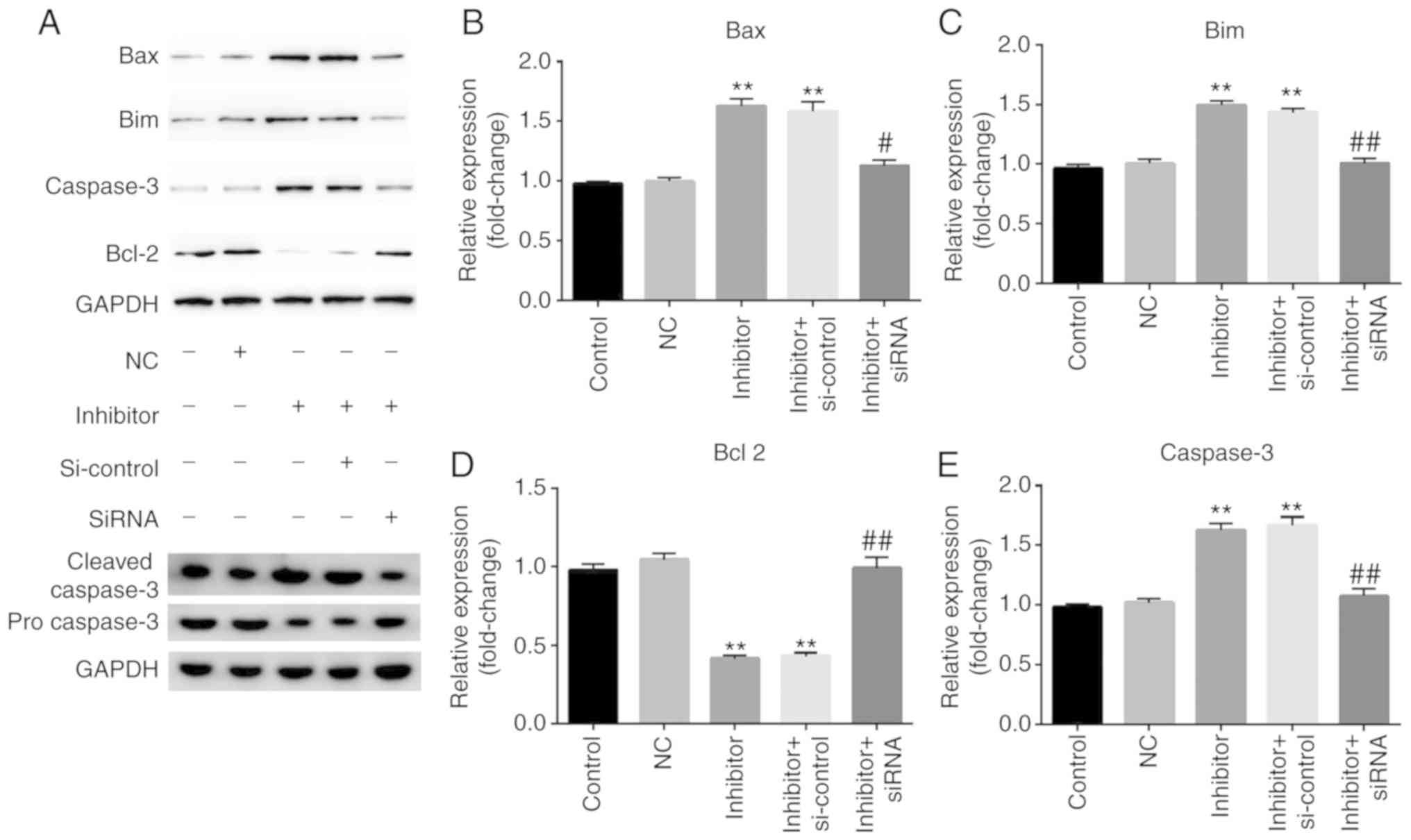 | Figure 4.Effect of miR-25-3p downregulation on
the expression level of Bim and apoptosis-associated molecules in
NP cells. NP cells were transfected with inhibitor, NC, inhibitor +
si-control or miR-25-3p inhibitor + inhibitor + siRNA for 48 h. (A)
A western blot assay was used to detect the protein expression of
Bim and apoptosis-associated molecules such as Bcl-2, Bax, cleaved
caspase-3 and caspase-3. Reverse transcription-quantitative PCR was
used to detect the relative mRNA expression of (B) Bax, (C) Bim,
(D) Bcl-2 and (E) caspase-3. Control, NP cells without any
treatment; NC, NP cells transfected with the NC of miR-25-3p
inhibitor; inhibitor, NP cells transfected with miR-25-3p
inhibitor; control-siRNA, NP cells transfected with control-siRNA;
inhibitor + si-control, NP cells transfected with miR-25-3p
inhibitor and control-siRNA; inhibitor + siRNA, NP cells
transfected with miR-25-3p inhibitor and Bim-siRNA. **P<0.01 vs.
control; #P<0.5, ##P<0.01 vs. inhibitor
+ si-control. miR, microRNA; NP, nucleus pulposus; NC Bim, Bcl-2
interacting mediator of cell death, negative control; si, small
interfering. |
miR-25-3p upregulation inhibits the
effects of IL-1β stimulation on normal NP cells
The effect of miR-25-3p on IL-1β-stimulated NP cells
was explored. The results indicated that miR-25-3p mimic
significantly increased the level of miR-25-3p in NP cells
(Fig. 5A) and Bim-plasmid
significantly enhanced the mRNA level of Bim in NP cells (Fig. 5B). miR-25-3p upregulation
significantly enhanced the viability of NP cells, which
IL-1β-reduced (Fig. 5C) and
reduced IL-1β-induced NP cell apoptosis (Fig. 5D and E), These changes were
reversed by the Bim-plasmid. miR-25-3p upregulation markedly
decreased IL-1β-enhanced expression of Bim, Bax, cleaved caspase-3
and caspase-3, and increased IL-1β-reduced expression of Bcl-2 and
pro-caspase-3 (Fig. 6). These
changes were reversed by Bim-plasmid.
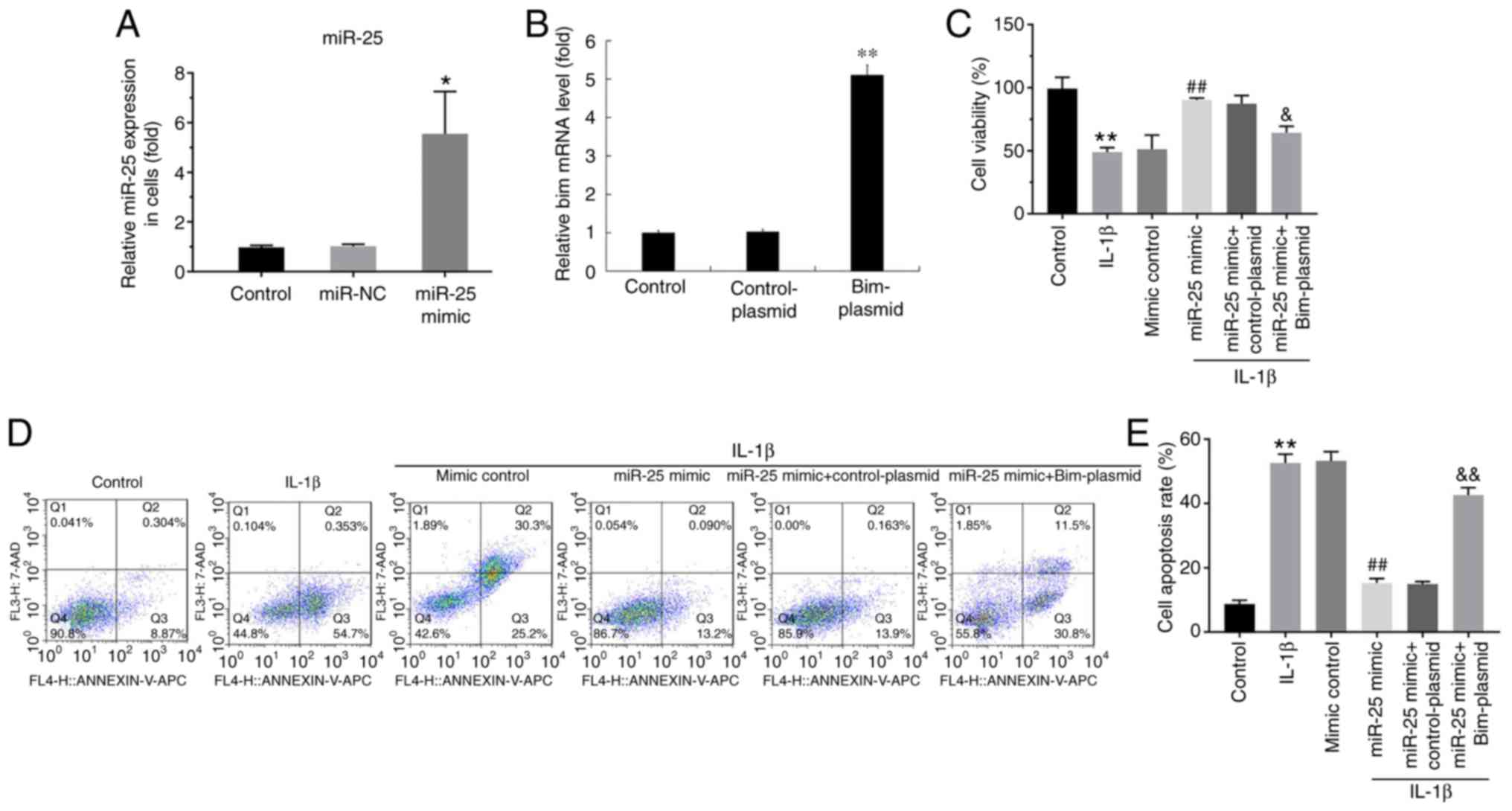 | Figure 5.Effect of miR-25-3p on the
proliferation and apoptosis of IL-1β-induced NP cells. NP cells
were transfected with miR-25-3p mimic, mimic control, miR-25-3p
mimic + control-plasmid or miR-25-3p mimic + Bim-plasmid for 48 h.
Then, NP cells were treated with 10 ng/ml IL-1β for 24 h. (A) The
level of miR-25-3p in NP cells was detected using RT-qPCR. (B) The
mRNA level of Bim in NP cells was detected using RT-qPCR. (C) Cell
Counting Kit-8 assay detected the viability of NP cells transfected
with miR-25-3p mimic, mimic control, miR-25-3p mimic +
control-plasmid or miR-25-3p mimic + Bim-plasmid for 48 h and
treated with 10 ng/ml IL-1β for 24 h. (D) Flow cytometry assay and
(E) analysis measured the apoptosis of NP cells transfected with
miR-25-3p mimic, mimic control, miR-25-3p mimic + control-plasmid
or miR-25-3p mimic + Bim-plasmid for 48 h and treated with 10 ng/ml
IL-1β for 24 h. *P<0.05 and **P<0.01 vs. control;
##P<0.01 vs. IL-1β; &P<0.05 and
&&P<0.01 vs. IL-1β + miR-25 mimic. miR,
microRNA; NP, nucleus pulposus; IL, interleukin; RT-qPCR, reverse
transcription-quantitative PCR; Bim, Bcl-2 interacting mediator of
cell death. |
Discussion
IDD is the main cause of lower back pain and is a
medical condition that constitutes a heavy burden on the global
medical system, with serious socioeconomic consequences (22–24).
At present, due to work and family reasons (25), the incidence of IDD is increasing,
particularly in China. There are numerous studies focusing on the
etiology of IDD, such as genetics (26), mechanical load (27) and environmental factors (28); however, the pathology of IDD is not
fully understood.
A previous study demonstrated that abnormal
expression of miRNA-140 is associated with degenerative diseases
such as osteoarthritis, which is characterized by similar
pathological changes to those in IDD (29). In addition, miRNAs have gained
considerable attention as regulators of gene expression and play
important roles in the prevention and treatment of IDD (30,31).
It has been reported that TNF-α is a key pro-inflammatory cytokine,
which not only has important roles in the inflammatory
microenvironment of cancer and Mycobacterium tuberculosis
infection (32), but also plays a
critical role in IDD via its effects on NP cell apoptosis (33).
Previous research studies have demonstrated that
several miRNAs are dysregulated in IDD, including miR-21, miR-10b,
miR-640 and miR-27 (34–37). miR-200c is upregulated in
degenerative NP tissues (38). In
the present study, miR-25-3p was downregulated in degenerated NP
cells. As expected, IDD-associated markers such as ACAN, collagen
I, collagen II and SOX-9 were highly expressed in degenerated NP
cells. A number of studies have shown that the great majority of
miRNAs play an important role in their biological function by
binding to their target gene (9–11).
Bim is located in the outer mitochondrial membrane
and belongs to the pro-apoptotic Bcl-2 family, which contributes to
TNF-α-induced apoptotic signaling events (39,40).
Bim is closely associated with the development and treatment of
autoimmune diseases, degenerative diseases and tumors. Previous
studies have demonstrated that Bim is a target gene for multiple
miRNAs (41,42). Previous studies have shown that Bim
is a putative miR-24 target (43,44).
One study suggested that miR-24 regulates hepatocyte apoptosis by
suppressing Bim (43). In the
present study, the dual luciferase reporter assay indicated that
Bim was the target for miR-25-3p. Next, to investigate the effect
of miR-25-3p on cell proliferation and apoptosis, normal NP cells
were transfected with miR-25-3p inhibitor, NC, miR-25-3p inhibitor
+ si-control or miR-25-3p inhibitor + siRNA. The results
demonstrated that transfection of miR-25-3p inhibitor could
decrease cell proliferation, induce cell apoptosis and increase the
expression of Bim, Bax, cleaved caspase-3 and caspase-3 and reduce
Bcl-2 and pro-caspase-3 expression in NP cells.
Finally, an IDD in vitro cell model was
established by treating normal NP cells with 10 ng/ml IL-1β for 24
h. Then, the effects of miR-25-3p on IL-1β-treated NP cells were
investigated. The results indicated that miR-25-3p upregulation
enhanced IL-1β-reduced NP cell viability and reduced IL-1β-induced
NP cell apoptosis. miR-25-3p upregulation decreased IL-1β-enhanced
expression of Bim, Bax, cleaved caspase-3 and caspase-3, and
increased IL-1β-reduced expression of Bcl-2 and pro-caspase-3.
Notably, all the effects of miR-25-3p upregulation on NP cells were
reversed by the Bim-plasmid.
Taken together, these results indicated that
miR-25-3p was downregulated in IDD and it was involved in the
development of IDD through regulating NP cell proliferation,
apoptosis and ECM deposition by targeting Bim. The current study
may have provided new insights for the development of novel
therapeutic strategies for IDD. However, this is a preliminary
study on miR-25 in IDD. In order to verify the role of miR-25 in
IDD, further experiments are needed. For example, the expression of
miR-25 in human IDD samples needs to be detected and the role of
miR-25/Bim in human NP cells should be investigated. The
association between the expression of miR-25/Bim and the
clinicopathological features of patients with IDD needs to be
explored. In addition, the effect of miR-25 on IDD should be
studied in vivo. These topics will be investigated in the
future.
Acknowledgements
Not applicable.
Funding
No funding received.
Availability of data and materials
The analyzed data sets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZZ contributed to study design, data collection,
statistical analysis, data interpretation and manuscript
preparation. JZ, YY, KZ and RuW contributed to data collection and
statistical analysis. RaW contributed to data collection,
statistical analysis and manuscript preparation. All authors read
and approved the final version of the manuscript.
Ethics approval and consent to
participate
The current study was approved by the Animal Ethics
Committee of the No. 903 Hospital of People's Liberation Army
(approval no. IRB SOP/01.03/01.1).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tang P, Gu JM, Xie ZA, Gu Y, Jie ZW, Huang
KM, Wang JY, Fan SW, Jiang XS and Hu ZJ: Honokiol alleviates the
degeneration of intervertebral disc via suppressing the activation
of TXNIP-NLRP3 inflammasome signal pathway. Free Radic Biol Med.
120:368–379. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu D, Zheng C, Wu J, Huang R, Chen X,
Zhang T and Zhang L: Molecular biological effects of weightlessness
and hypergravity on intervertebral disc degeneration. Aerosp Med
Hum Perform. 88:1123–1128. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang Y, Jiang T, Chen J, Yin GY and Fan
J: Effects of kartogenin on the attenuated nucleus pulposus cell
degeneration of intervertebral discs induced by interleukin-1β and
tumor necrosis factor-α. Int J Mol Med. 41:749–756. 2018.PubMed/NCBI
|
|
4
|
Viguet-Carrin S, Roux JP, Arlot ME,
Merabet Z, Leeming DJ, Byrjalsen I, Delmas PD and Bouxsein ML:
Contribution of the advanced glycation end product pentosidine and
of maturation of type I collagen to compressive biomechanical
properties of human lumbar vertebrae. Bone. 39:1073–1079. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Vergroesen PP, Kingma I, Emanuel KS,
Hoogendoorn RJ, Welting TJ, van Royen BJ, van Dieën JH and Smit TH:
Mechanics and biology in intervertebral disc degeneration: A
vicious circle. Osteoarthritis Cartilage. 23:1057–1070. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mellis D and Caporali A: MicroRNA-based
therapeutics in cardiovascular disease: screening and delivery to
the target. Biochem Soc Trans. 46:11–21. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ganju A, Khan S, Hafeez BB, Behrman SW,
Yallapu MM, Chauhan SC and Jaggi M: miRNA nanotherapeutics for
cancer. Drug Discov Today. 22:424–432. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Long H, Wang X, Chen Y, Wang L, Zhao M and
Lu Q: Dysregulation of microRNAs in autoimmune diseases:
Pathogenesis, biomarkers and potential therapeutic targets. Cancer
Lett. 428:90–103. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ro S, Park C, Young D, Sanders KM and Yan
W: Tissue-dependent paired expression of miRNAs. Nucleic Acids Res.
35:5944–5953. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mallory AC and Vaucheret H: MicroRNAs:
Something important between the genes. Curr Opin Plant Biol.
7:120–125. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Garzon R, Calin GA and Croce CM: MicroRNAs
in cancer. Annu Rev Med. 60:167–179. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu JY, Yang LL, Ma C, Huang YL, Zhu GX and
Chen QL: miR-25-3p attenuates the proliferation of tongue squamous
cell carcinoma cell line Tca8113. Asian Pac J Trop Med. 6:743–747.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fei Q and Ethell DW: Maneb potentiates
paraquat neurotoxicity by inducing key Bcl-2 family members. J
Neurochem. 105:2091–2097. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bouillet P, Purton JF, Godfrey DI, Zhang
LC, Coultas L, Puthalakath H, Pellegrini M, Cory S, Adams JM and
Strasser A: BH3-only Bcl-2 family member Bim is required for
apoptosis of autoreactive thymocytes. Nature. 415:922–926. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li Y, Deutzmann A and Felsher DW:
BIM-mediated apoptosis and oncogene addiction. Aging (Albany NY).
8:1834–1835. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Guo Y, Schoell MC and Freeman RS: The von
Hippel-Lindau protein sensitizes renal carcinoma cells to apoptotic
stimuli through stabilization of BIM(EL). Oncogene. 28:1864–1874.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Baysan A, Yel L, Gollapudi S, Su H and
Gupta S: Arsenic trioxide induces apoptosis via the mitochondrial
pathway by upregulating the expression of Bax and Bim in human B
cells. Int J Oncol. 30:313–318. 2007.PubMed/NCBI
|
|
18
|
Bouillet P, Cory S, Adams J and Strasser
A: Role of BH3-only protein Bim in autoimmune and degenerative
diseases. Arthritis Res Ther. 5 (Suppl 3):S192003. View Article : Google Scholar
|
|
19
|
Issy AC, Castania V, Castania M, Salmon
CE, Nogueira-Barbosa MH, Bel ED and Defino HL: Experimental model
of intervertebral disc degeneration by needle puncture in Wistar
rats. Braz J Med Biol Res. 46:235–244. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bayne K: Revised guide for the care and
use of laboratory animals available. American physiological
society. Physiologist. 39:199, 208–211. 1996.
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Juniper M, Le TK and Mladsi D: The
epidemiology, economic burden, and pharmacological treatment of
chronic low back pain in France, Germany, Italy, Spain and the UK:
A literature-based review. Expert Opin Pharmacother. 10:2581–2592.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Phillips C, Main C, Buck R, Aylward M,
Wynne-Jones G and Farr A: Prioritising pain in policy making: The
need for a whole systems perspective. Health Policy. 88:166–175.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Waddell G: Low back pain: A twentieth
century health care enigma. Spine (Phila Pa 1976). 21:2820–2825.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang YG, Sun Z, Zhang Z, Liu J and Guo X:
Risk factors for lumbar intervertebral disc herniation in Chinese
population: A case-control study. Spine (Phila Pa 1976).
34:E918–E922. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kalichman L and Hunter DJ: The genetics of
intervertebral disc degeneration. Associated genes. Joint Bone
Spine. 75:388–396. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Walter BA, Korecki CL, Purmessur D,
Roughley PJ, Michalek AJ and Iatridis JC: Complex loading affects
intervertebral disc mechanics and biology. Osteoarthritis
Cartilage. 19:1011–1018. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Battié MC and Videman T: Lumbar disc
degeneration: Epidemiology and genetics. J Bone Joint Surg Am. 88
(Suppl 2):S3–S9. 2006. View Article : Google Scholar
|
|
29
|
Miyaki S, Nakasa T, Otsuki S, Grogan SP,
Higashiyama R, Inoue A, Kato Y, Sato T, Lotz MK and Asahara H:
MicroRNA-140 is expressed in differentiated human articular
chondrocytes and modulates interleukin-1 responses. Arthritis
Rheum. 60:2723–2730. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang HQ, Yu XD, Liu ZH, Cheng X, Samartzis
D, Jia LT, Wu SX, Huang J, Chen J and Luo ZJ: Deregulated mir-155
promotes Fas-mediated apoptosis in human intervertebral disc
degeneration by targeting FAdd and caspase-3. J Pathol.
225:232–242. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ji ML, Lu J, Shi PL, Zhang XJ, Wang SZ,
Chang Q, Chen H and Wang C: Dysregulated miR-98 contributes to
extracellular matrix degradation by targeting IL-6/StAt3 signaling
pathway in human intervertebral disc degeneration. J Bone Miner
Res. 31:900–909. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wibawa T, Pangemanan L, Rachmawaty FJ,
Rintiswati N, Mustofa and Soesatyo MH: Isoniazid (INH) treatment of
INH-resistant Mycobacterium tuberculosis inhibits infected
macrophage to produce TNF-alpha. Southeast Asian J Trop Med Public
Health. 45:1107–1113. 2014.PubMed/NCBI
|
|
33
|
Wang XH, Hong X, Zhu L, Wang YT, Bao JP,
Liu L, Wang F and Wu XT: Tumor necrosis factor alpha promotes the
proliferation of human nucleus pulposus cells via nuclear
factor-κB, c-Jun N-terminal kinase, and p38 mitogen-activated
protein kinase. Exp Biol Med (Maywood). 240:411–417. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yu X, Li Z, Shen J, Wu WK, Liang J, Weng X
and Qiu G: MicroRNA-10b promotes nucleus pulposus cell
proliferation through RhoC-Akt pathway by targeting HOXD10 in
intervetebral disc degeneration. PLoS One. 8:e830802013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu H, Huang X, Liu X, Xiao S, Zhang Y,
Xiang T, Shen X, Wang G and Sheng B: miR-21 promotes human nucleus
pulposus cell proliferation through PTEN/AKT signaling. Int J Mol
Sci. 15:4007–4018. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dong W, Liu J, Lv Y, Wang F, Liu T, Sun S,
Liao B, Shu Z and Qian J: miR-640 aggravates intervertebral disc
degeneration via NF-κB and WNT signalling pathway. Cell Prolif.
52:e126642019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu G, Cao P, Chen H, Yuan W, Wang J and
Tang X: MiR-27a regulates apoptosis in nucleus pulposus cells by
targeting PI3K. PLoS One. 8:e752512013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cheng X, Zhang L, Zhang K, Zhang G, Hu Y,
Sun X, Zhao C, Li H, Li YM and Zhao J: Circular RNA VMA21 protects
against intervertebral disc degeneration through targeting miR-200c
and X linked inhibitor-of-apoptosis protein. Ann Rheum Dis.
77:770–779. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Concannon CG, Tuffy LP, Weisová P, Bonner
HP, Dávila D, Bonner C, Devocelle MC, Strasser A, Ward MW and Prehn
JH: AMP kinase-mediated activation of the BH3-only protein Bim
couples energy depletion to stress-induced apoptosis. J Cell Biol.
189:83–94. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kaufmann T, Jost PJ, Pellegrini M,
Puthalakath H, Gugasyan R, Gerondakis S, Cretney E, Smyth MJ, Silke
J, Hakem R, et al: Fatal hepatitis mediated by tumor necrosis
factor TNFalpha requires caspase-8 and involves the BH3-only
proteins Bid and Bim. Immunity. 30:56–66. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sur S, Steele R, Shi X and Ray RB:
miRNA-29b inhibits prostate tumor growth and induces apoptosis by
increasing Bim expression. Cells. 8:14552019. View Article : Google Scholar
|
|
42
|
Labi V, Peng S, Klironomos F, Munschauer
M, Kastelic N, Chakraborty T, Schoeler K, Derudder E, Martella M,
Mastrobuoni G, et al: Context-specific regulation of cell survival
by a miRNA-controlled BIM rheostat. Genes Dev. 33:1673–1687. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Feng Z, Li Z, Zhu D, Ling W, Zheng L, Pu L
and Kong L: Mir-24 regulates hepatocyte apoptosis via BIM during
acute liver failure. Am J Transl Res. 9:4925–4935. 2017.PubMed/NCBI
|
|
44
|
Pan LJ, Wang X, Ling Y and Gong H: MiR-24
alleviates cardiomyocyte apoptosis after myocardial infarction via
targeting BIM. Eur Rev Med Pharmacol Sci. 21:3088–3097.
2017.PubMed/NCBI
|