Introduction
Prostate cancer (PCa), which is a clinically
heterogeneous-multifocal disease, is the second most frequently
diagnosed cancer in males worldwide (1). The most recent US statistic reported
that in 2017 the number of new cases and cases of mortality
associated with PCa in the US were 161,360 and 26,730, respectively
(1). microRNAs (miRNAs or miRs)
are small noncoding RNAs (length ~22 nt) expressed in animal and
plant cells (2,3). miRNAs are the smallest known carriers
of gene-encoded, post-transcriptional regulatory information in
plants and animals (4), and
suppress a variety of gene expression at the post-transcriptional
level by pairing with complementary nucleotide sequences in the
3′-untranslated region (UTR) of specific target genes (5). Accumulated evidence suggests that
miRNAs serve as tumor suppressor or oncogenes depending on the
target gene (6,7) and dysregulation of miRNAs may serve
important roles in the initiation and progression of types of
cancer of various tissue origins (8).
Similar to other malignancies, PCa has a distinctive
miR expression profile, which has been the basis for the functional
study of miRNA in PCa (7–9). Porkka et al (7) demonstrated that 37 miRNAs were
downregulated and 14 upregulated in PCa tissues compared with
benign tissues. A previous study obtained differential expression
data of miRNAs in PCa determined by miRNA microarray analyses and
reported that 11 miRNAs were upregulated and 17 miRNAs were
downregulated in PCa (10).
However, these previous gain-of-function studies did not include
loss-of-function analyses using miRNA knockout and applying the
clustered regularly interspaced short palindromic repeats (CRISPR)
and CRISPR-associated (Cas) 9 system, which can effectively,
specifically and stably suppress gene expression in vitro
and in vivo (11,12). CRISPR/Cas9 is a recently discovered
genome editing system, which has markedly changed the way that
researchers study genes and their functions in mammalian systems
(12,13). It is derived from the CRISPR/Cas
bacterial-acquired immune system and Cas9 is directed by guide
(g)RNAs, which match the DNA targeted in cleavage to modify the
respective gene (11,13).
To classify rapidly the differential functions of
miRNAs, which were identified through miRNA expression profiling in
PCa, the CRISPR/Cas9 system was used to knockout the expression of
PCa-associated miRNAs, including miR-205, miR-221, miR-455-3p,
miR-222, miR-224, miR-505, miR-23b, miR-30c, miR-1225-5p and
miR-663a in LNCaP cells. The proliferation, invasion and metabolic
changes were determined.
Materials and methods
Prostate cancer-associated miRNA
Data
The prostate cancer-associated miRNA is the
differentially expressed miRNA in the PCa tissue compared with the
adjacent benign prostate gland tissues, which is from our miRNA
expression profiling microarray data (GEO, http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE34932)
and the Taylor database (GEO, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE21032;
Table I) (10).
 | Table I.Differentially expressed miRNAs in
PCa (Fold change ≥1.5, P<0.05). |
Table I.
Differentially expressed miRNAs in
PCa (Fold change ≥1.5, P<0.05).
|
| miRNA microarray
data | Taylor
database |
|
|---|
|
|
|
|
|
|---|
| Name | Regulation | Fold change | P-value | Fold change | P-value | Chromosomal
location |
|---|
| hsa-miR-205 | Down | 58.96 | 0.009 | 3.19 | <0.001 |
1:209605478-209605587[+] |
| hsa-miR-221 | Down | 5.15 | 0.036 | 3.24 | <0.001 |
X:45605585-45605694[-] |
| hsa-miR-455-3p | Down | 4.39 | 0.014 | 1.63 | 0.003 |
9:116971714-116971809[+] |
| hsa-miR-222 | Down | 3.94 | 0.036 | 3.55 | <0.001 |
X:45606421-45606530[-] |
| hsa-miR-221 | Down | 3.50 | 0.050 | 1.59 | 0.008 |
X:45605585-45605694[-] |
| hsa-miR-224 | Down | 3.32 | 0.011 | 2.56 | <0.001 |
X:151127050-151127130[-] |
| hsa-miR-505 | Down | 2.72 | 0.038 | 1.52 | <0.001 |
X:139006307-139006390[-] |
| hsa-miR-23b | Down | 2.70 | 0.033 | 1.86 | <0.001 |
9:97847490-97847586[+] |
| hsa-miR-30c | Down | 2.09 | 0.018 | 1.60 | <0.001 |
1:41222956-41223044[+] |
|
hsa-miR-1225-5p | Up | 3.37 | 0.041 | 1.56 | 0.005 |
16:2140196-2140285[-] |
| hsa-miR-663a | Up | 2.71 | 0.044 | 2.64 | <0.001 |
20:26188822-26188914[-] |
Cell culture
The human PCa cell line, LNCaP was purchased from
the American Type Culture Collection and cultured in RPMI-1640
medium (HyClone; Cytiva) supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.), 2 mM L-glutamine and
1% penicillin/streptomycin antibiotics (Thermo Fisher Scientific,
Inc.). Cells were maintained at 37°C in a humidified chamber
supplemented with 5% CO2.
Single guide (sg)RNA design and
plasmid construction
Target sequences of miR-205, miR-221, miR-455-3p,
miR-222, miR-224, miR-505, miR-23b, miR-30c, miR-1225-5p and
miR-663a for CRISPR interference were designed. Taking miR-205 as
an example (Gene ID406988; http://www.ncbi.nlm.nih.gov/gene/406988): The gene
sequence
5′-AAAGATCCTCAGACAATCCATGTGCTTCTCTTGTCCTTCATTCCACCGGAGTCTGTCTCATACCCAACCAGATTTCAGTGGAGTGAAGTTCAGGAGGCATGGAGCTGACA-3′
was retrieved and downloaded from GeneBank and the sgRNA was
designed by Zhang lab (https://zlab.bio/guide-design-resources) using Target
Finder (version 2014, http://targetfinder.flycrispr.neuro.brown.edu/) and
DNA 2.0 gRNA (https://www.dna20.com). A total of
four optimal target sequences for each miRNA were selected and four
scramble sequences were used as controls (Table SI). Subsequently, two
complementary oligonucleotides with Bbsl restriction sites
for gRNAs were synthesized and cloned into CRISPR/Cas9
lentiCRISPR-v2 vector (cat. no. 52961; Addgene, Inc.) by HYYMed
Company using T4 DNA ligase (cat. no. D2011B; Takara Biotechnology
Co., Ltd.) (14).
Cell line construction and
transfection
LNCaP cells were seeded in 6-well plates
(3×105), grown to ~70% confluence and transfected with
the CRISPR/Cas9 lentiCRISPR-v2 vector plasmid construct (2 µg)
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols. On the
day following transfection, cells were treated with 10 mg/ml of
puromycin (Beyotime Institute of Biotechnology) and maintained at
37°C in a humidified chamber supplemented with 5% CO2
for >2 days. Subsequently, the cells were harvested or cultured
for further experiments.
T7 endonuclease I (T7E1) assays for
knockout efficiency
T7E1 digestion assay was performed to analyze the
gene knockout efficiency as previously described (15,16).
Genomic DNA of transfected cells was extracted using HiPure Tissue
DNA Mini kit (cat. no. D3121-03; Angen Biotech Co., Ltd.) according
to the manufacturer's protocols. The target site of sgRNA was
amplified by PCR and DNA fragments were subjected to digestion with
the mismatch-sensitive T7E1 (cat. no. E3321; New England BioLabs,
Inc.). For T7E1 digestion, amplified PCR products were denatured at
95°C for 5 min and slowly cooled to room temperature to allow
formation of heteroduplex DNA; the annealed products were incubated
with 0.5 µl T7E1 for 30 min at 37°C and the digested DNA was
separated on 2% agarose gels. Based on relative band intensities,
the small insertion and deletion (indel) percentage was calculated
using the formula, 1-√ 1-(b+c)/(a+b+c) ×100, where a is the
integrated intensity of the undigested PCR product, and b and c are
the integrated intensities of each cleavage product (14).
Cell proliferation assay
For cell viability assays, cells were seeded in
96-well plates at 5,000 cells/200 µl per well and cultured for 24,
48 and 72 h. Cells were then incubated with 20 µl Cell Counting
Kit-8 (CCK-8) solution (cat. no. C0038; Beyotime Institute of
Biotechnology) for 2 h at 37°C according to the manufacturer's
protocol. The absorbance was measured at 450 nm using a
spectrophotometer.
Cell invasion assay
Transwell inserts (pore size, 8 µm) were filled with
50 µl of a mixture of serum-free RPMI-1640 medium and Matrigel
(ratio, 1:10; BD Biosciences) for 30 min at 37°C. The inserts were
placed in 24-well tissue culture plates (Transwell; Corning Inc.)
containing 10% FBS-medium. Following solidification by incubation
in 37°C for 4 h, 8×104 cells in 200 µl medium were
placed in upper chambers. Following a 48-h incubation at 37°C with
5% CO2, culture medium with mitomycin to halt the
mitosis was added and the membranes were fixed with 10% formalin
for 5 min at room temperature and stained with 0.05% crystal violet
for 10 min at room temperature. Migrated cells were assessed and
the data were expressed as the mean ± standard deviation.
Detection of lactic acid
The culture medium was collected and the
concentration of lactic acid was determined using the GEM Premier
3000 Blood Gas analyzer (Instrumentation Laboratory Inc.). Lactate
was measured by amperometry based on the following principle:
Lactate oxidase, immobilized on a lactate biochip sensor,
selectively converted lactate into pyruvate and hydrogen peroxide
(H2O2), the released
H2O2 oxidizes on a platinum electrode and
produces an electric current that is proportional to the lactate
concentration in the blood sample.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Data analysis was performed by using one-way ANOVA followed by
Tukey's post hoc test or independent samples t-test between
control and experimental groups using SPSS 17.0 (SPSS, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Prostate cancer-associated miRNAs
selection
The PCa differentially expressing miRNAs with fold
change >1.5 were selected for miRNA expression profiling
(Table I). The following
differentially expressing miRNAs were selected: hsa-miR-205,
hsa-miR-221, hsa-miR-455-3p, hsa-miR-222, hsa-miR-224, hsa-miR-505,
hsa-miR-23b, hsa-miR-30c, hsa-miR-1225-5p and hsa-miR-663a
(downregulated, 8; upregulated, 2; Table I) (10). To further study the above miRNAs
function in prostate cancer cell, the use of CRISPR/Cas9-mediated
gene knockout technology for the rapid classification of the
function of differential miRNAs was assessed in the following.
miRNAs knockout by CRISPR/Cas9
system
In the present study, a total of four optimal target
sgRNA sequences for each miRNA were designed (Table SI), and every sgRNA position in
genomic DNA was illustrated in Fig.
SIA. The sgRNA sequence was synthesized (Table SII) and cloned into the
CRISPR/Cas9 lentiCRISPR-v2 vector as demonstrated in Fig. S1B (14). To verify the knockout efficiency of
miRNAs in human LNCaP cells, a mismatch-sensitive T7E1 assay was
conducted to analyze the sgRNA target genomics profiling as
previously described (15). The
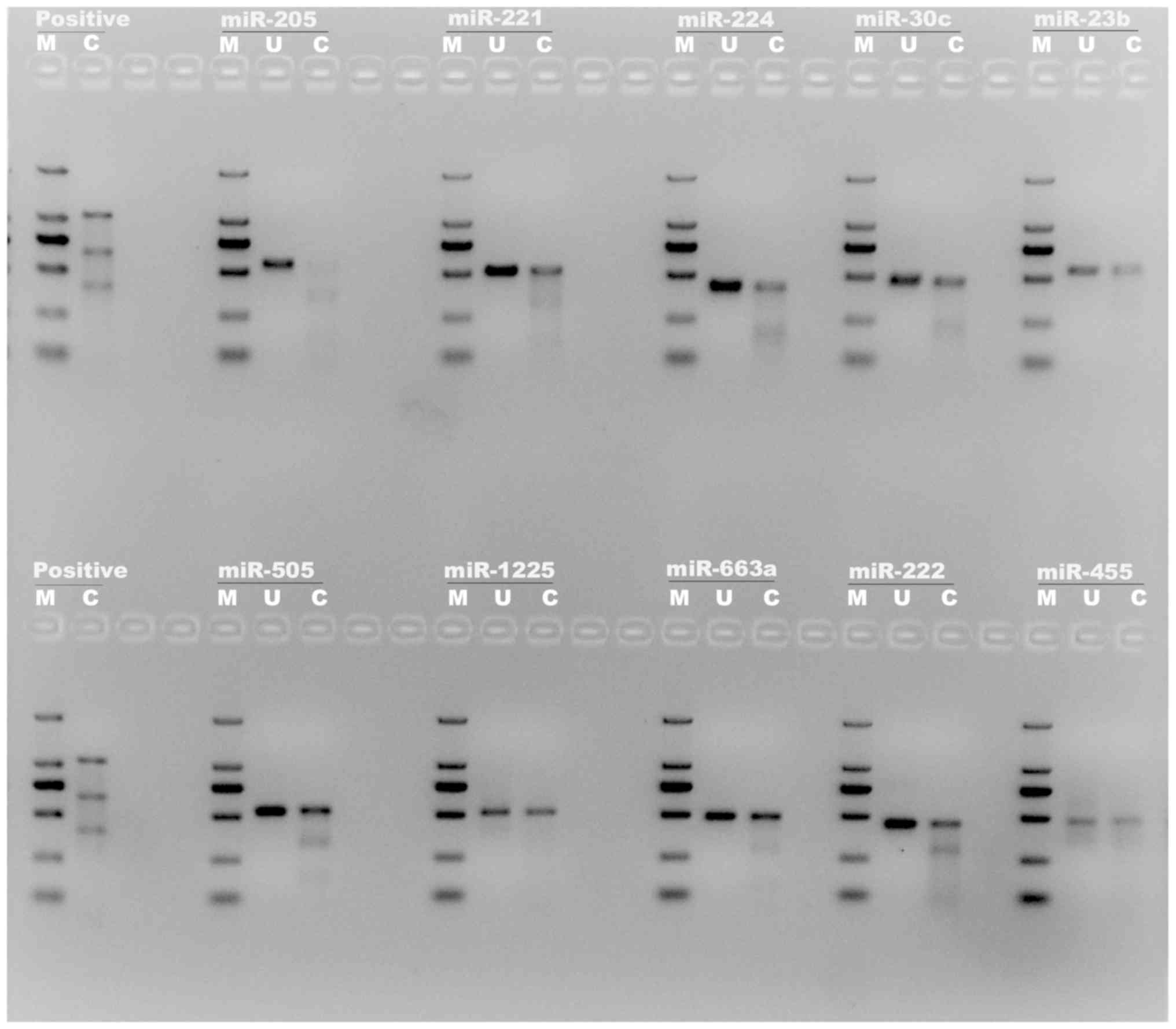
PCR primers for T7E1 assay of miRNAs were designed (Table SIII and Fig. S2). The T7E1 assay result is
presented in Fig. 1. By comparing
the bright lanes of ‘uncut’ (U) and ‘cut’ (C), it demonstrated that
the mixture of the miR-205 sgRNA vectors demonstrated effective
knockout of the miR-205 gene in LNCaP cells with >90% indel
percentage and the other miRNAs also demonstrated a positive
knockout effect.
miRNAs affect prostate cancer cell
proliferation
To study the influence of the differentially
expressed miRNAs on PCa cells, a lentiviral vector knockout system
for the miRNAs using the CRISPR/Cas9 system was applied and stable
LNCaP cells were established following lentivector transfection.
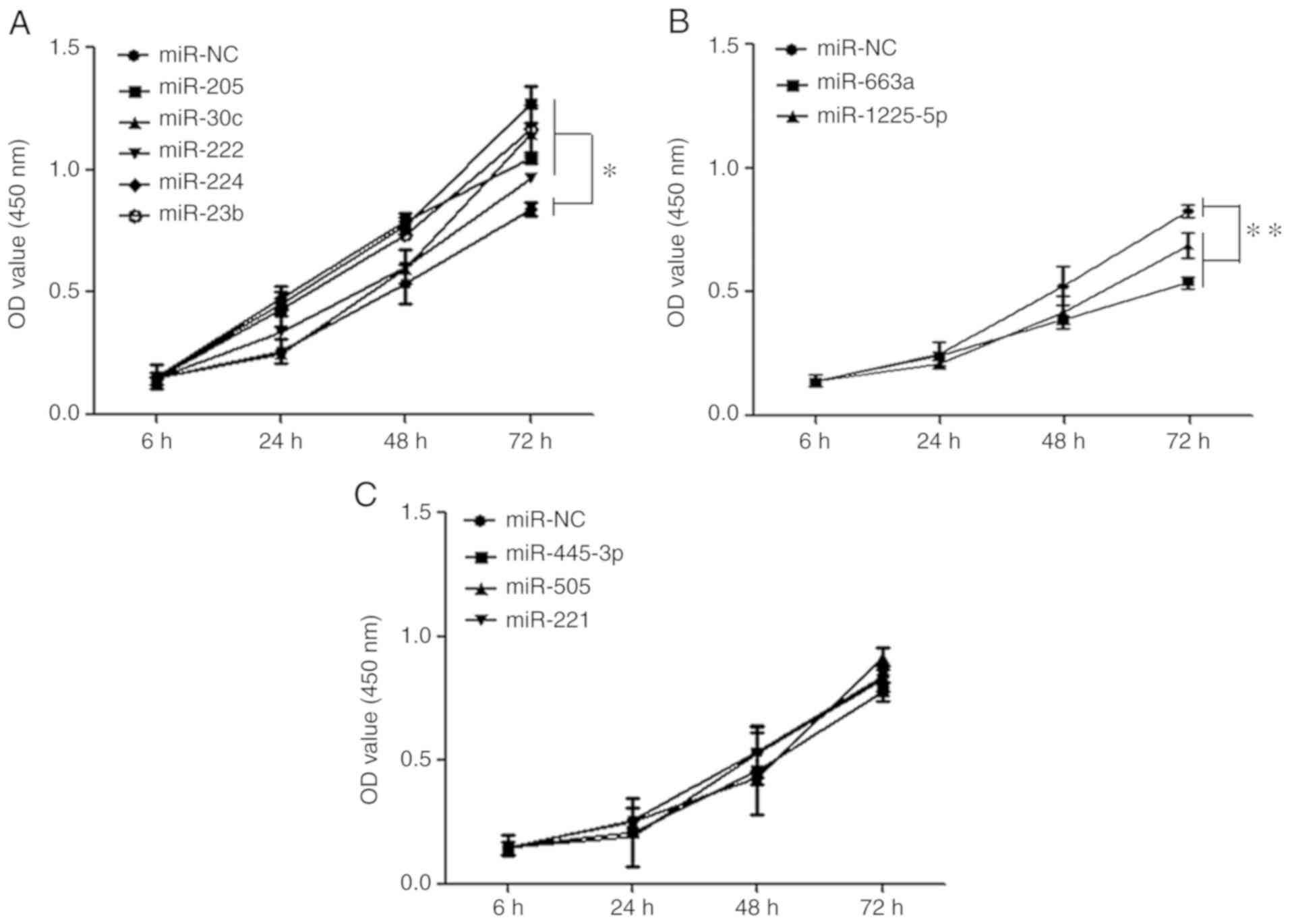
Proliferation was investigated by CCK-8 assay. The results
indicated that the proliferative abilities of miR-222, miR-224,
miR-23b, miR-205 and miR-30c knockout cells were significantly
enhanced (P<0.05) compared with the blank vector groups in LNCaP
cells lines at 72 h following transfection (Fig. 2A). In contrast, the proliferative
abilities of miR-1225-5p and miR-663a knockout were significantly
reduced (P<0.01; Fig. 2B). The
other group of miR-221, miR-455-3p, miR-505 showed no function on
the cell proliferation of prostate cancer (P>0.05; Fig. 2C).
miRNAs affect prostate cancer cell
invasion
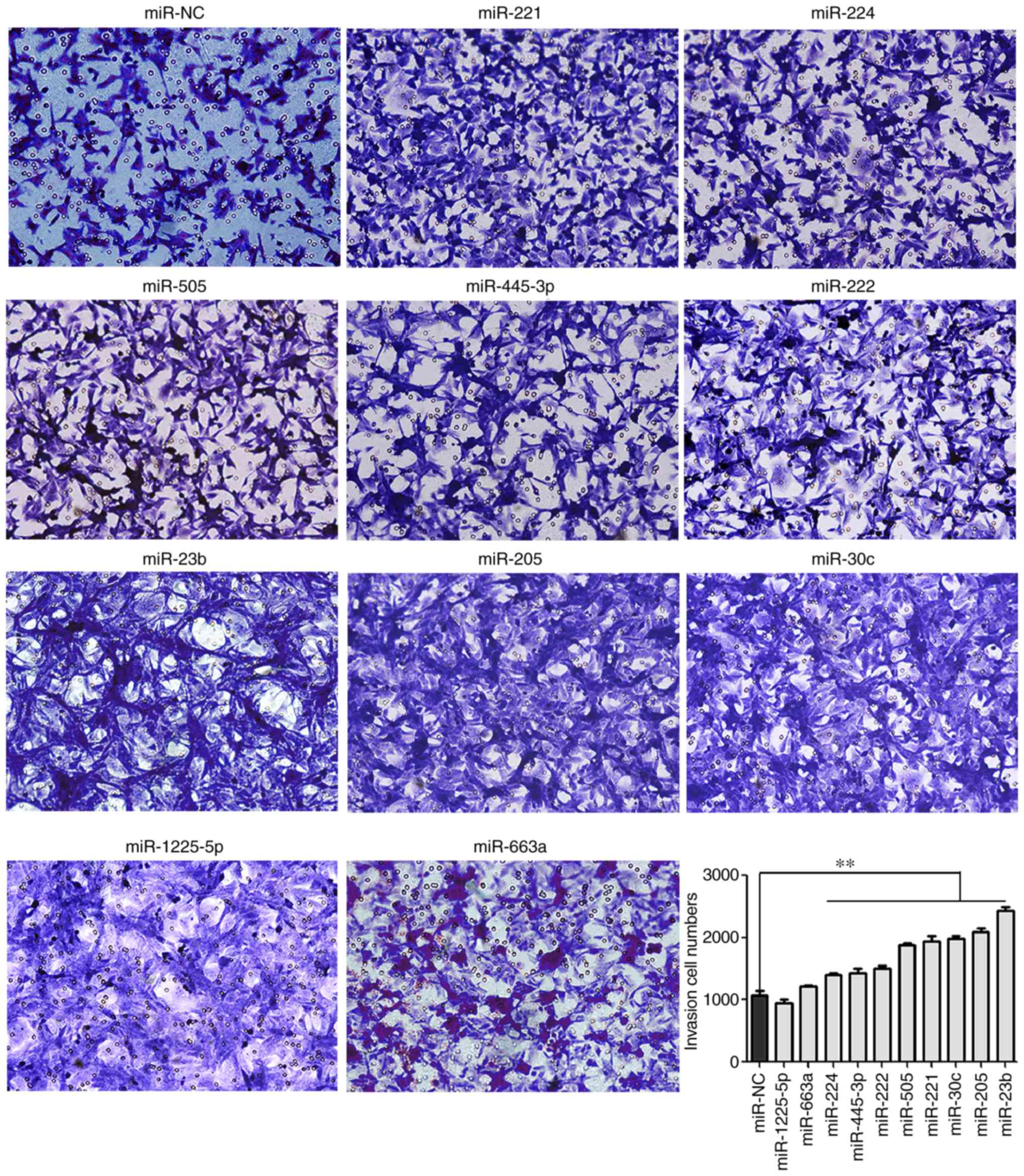
In the present study, Transwell assays were
performed to study the effect of various miRNAs on PCa cell
invasion. The results revealed that knockout of miR-205, miR-221,
miR-455-3p, miR-222, miR-224, miR-505, miR-23b and miR-30c
significantly increased invasive activities of LNCaP cells compared
with the control cells (P<0.01; Fig. 3), which suggested that these miRNAs
may act as tumor suppressors in PCa.
miRNAs affect prostate cancer cell
metabolism
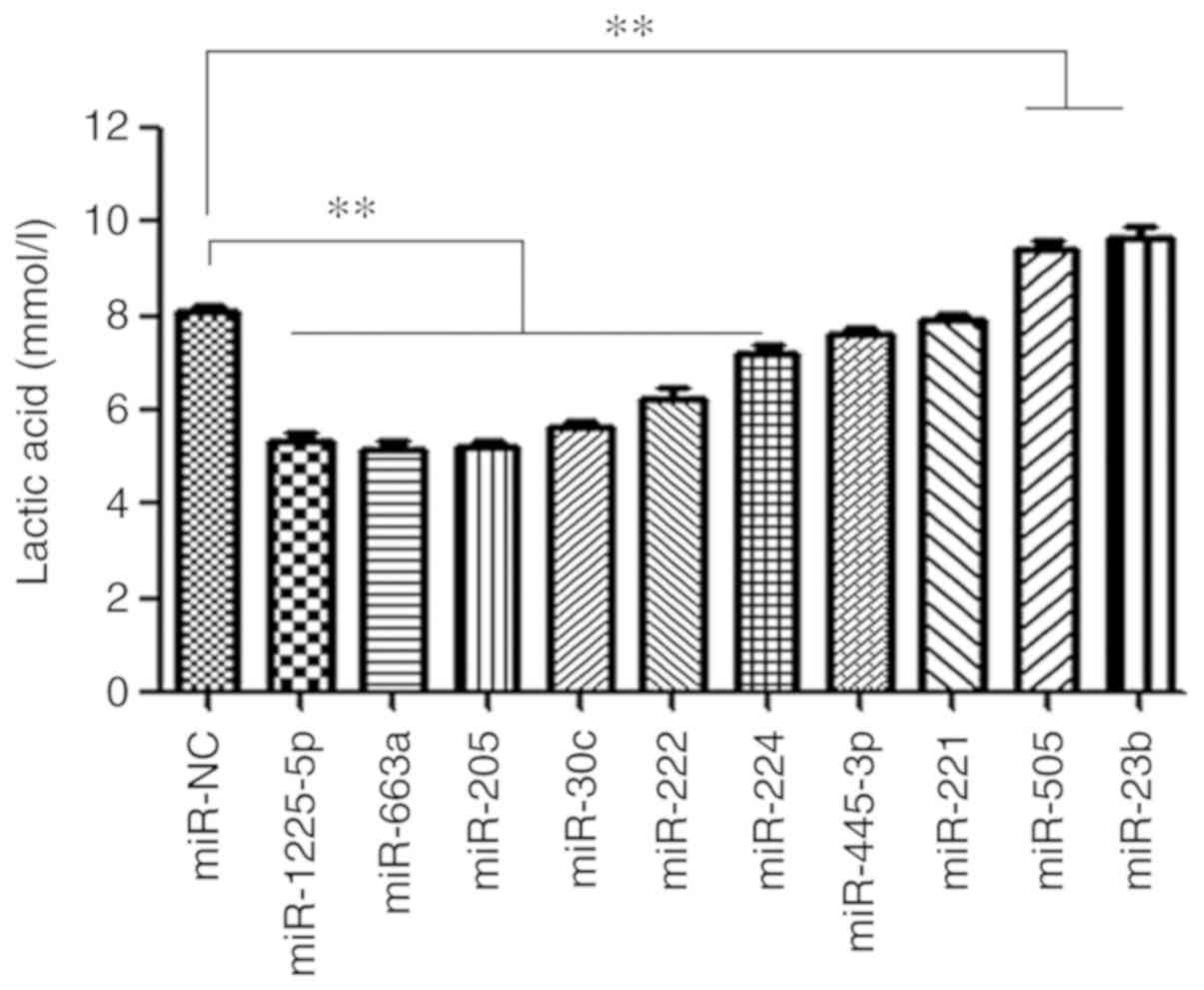
In the present study, the lactic acid concentration
in culture medium of miRNA-knockout LNCaP cells was detected in
vitro to represent the cell metabolism. As presented in
Fig. 4, knockout of miR-505 and
miR-23b demonstrated increased lactate production in LNCaP cells
in vitro (P<0.01), while knockout of miR-1225-5p,
miR-663a, miR-205, miR-30c, miR-222 and miR-224 significantly
decreased the lactic acid concentration (P<0.01).
Discussion
PCa is one of the most common malignancies in Europe
and the US and poses a serious threat to the health of older males
(1,17). miRNAs are small non-coding RNA
molecules that regulate gene expression by base pairing with their
target mRNAs and they contribute to cancer initiation, progression
and metastasis by directly modulating oncogenic or tumor suppressor
pathways (18). Previous studies
demonstrated that miRNA expression patterns serve as phenotypic
signatures of various types of cancer and may be used as
diagnostic, prognostic and therapeutic tools (9,10,19).
Previous studies have analyzed global miRNA expression profiles or
the functional role of miRNAs in PCa; however, results are
inconsistent (20,21).
In the present study, eight down- and two
upregulated miRNAs in PCa tissues were selected based on previous
miRNA expression profiling microarray data and the Taylor database.
Downregulation of miR-205, miR-221, miR-455-3p, miR-222, miR-224,
miR-505, miR-23b and miR-30c in PCa tissues led to the hypothesis
that these miRNAs may function as tumor suppressors.
CRISPR/Cas9-mediated gene knockout technology was applied to
classify the differentially expressed miRNA functions.
Proliferation and invasive abilities of the miRNAs were
subsequently validated by CCK-8 and Transwell assays. According the
Warburg effect, aggressive tumors frequently exhibit metabolic
alteration and reveal an increasing dependence on the glycolytic
pathway to generate ATP, even in the presence of oxygen (22). The increased glycolysis may be a
response to the hypoxic conditions characterizing the
microenvironment of malignant cells (22). The generation and accumulation of
lactic acid leads to microenvironmental acidosis and facilitates
tumor proliferation, invasion and metastasis (23). In the present study, the lactic
acid concentration of the miRNA knockout LNCaP cells was detected
in vitro.
miR-205 is reported to be downregulated in patients
with PCa and acts as a tumor suppressor (24–26).
Verdoodt et al (27)
reported that miRNA-205 is a novel regulator of the anti-apoptotic
protein B-cell lymphoma 2 and promotes apoptosis in PCa cells in
response to DNA damage by cisplatin and doxorubicin treatment, and
it further inhibits proliferation in PC3 and LNCaP cells. Recently,
studies (28,29) suggested that miRNA-205 serves as a
prognostic factor and suppresses proliferation and invasion in
various types of cancers, which is consistent with the results of
the present study. Downregulation of miR-221 and miR-222 are
frequently observed in PCa samples and certain differentially
expressed gene targets are associated with PCa (9,10,21,30).
The results of the present study indicated that the downregulation
of miR-30c promoted cell proliferation and invasion, which was
consistent with previous studies (31,32).
Other studies (33,34) suggest that miR-224 directly targets
the Ras-association domain family and acts as a tumor promoter in
cervical and gastric cancer progression. Liu et al (35) indicated that miR-224 inhibits
proliferation and migration of breast cancer cells by
downregulating Fizzled-5 expression. The present study demonstrated
that miR-224 was downregulated in PCa and knockout of miR-224
promoted proliferation and invasion in LNCaP cells. Recently,
miRNA-455-3p was described to be markedly downregulated in PCa
cells and clinical tumor specimens and it functions as a tumor
suppressor by targeting eukaryotic translation initiation factor 4E
and by inhibiting proliferation of PCa cells (36). Notably, the present study revealed
that knockout of miRNA-455-3p promoted invasion of LNCaP cells. It
was reported that miR-505 functions as a tumor suppressor in
endometrial cancer by targeting tumor growth factor α and miR-505
modulated cancer proliferation and migration in human non-small
cell lung cancer through inverse regulation of FZD4
(37). However, the functions
remain to be elucidated in human PCa.
The present study described that knockout of miR-505
efficiently enhanced PCa cell invasion. A downregulated expression
of miR-23b was described for malignant PCa tissues. The
miR-23b/-27b cluster functions as a metastasis-suppressor by
decreasing Huntingtin-interacting protein 1-related protein levels
in preclinical models of PCa (38), which is consistent with the results
of the present study that miR-23b inhibited PCa proliferation and
invasion. Knockout of miR-505 and miR-23b demonstrated increased
lactate production in LNCaP cells in vitro, which indicated
that miR-505 and miR-23b may be involved in the metabolic
regulation. The difference in lactic acid levels between the
control and miRNA-knockout groups was not as significant as
anticipated, this may be a limitation in the detection method of
the GEM Premier 3,000 Blood Gas analyzer; the lactic acid levels
were detected in conditional medium but not in cells. A better
method and devices, such as Seahorse XF Analyzers, can be applied
to live-cell metabolic assays.
miRNA-663a has been revealed to be downregulated in
non-small cell lung cancer and to suppress cell proliferation and
invasion by targeting JunD (39)
and miR-1225-5p was revealed to serve to constrain gastric
carcinoma growth and the metastatic potential via inhibition of
insulin receptor substrate-1 and β-catenin signaling (40). The role of miR-663a and miR-1225-5p
in PCa has not been previously addressed. The data from the present
study indicated that miR-663a and miR-1225-5p were upregulated in
PCa tissues and cell proliferation in miR-663a and miR-1225-5p
knockout PCa cells was significantly lower compared with miR-NC
cells. In addition, knockout of miR-1225-5p and miR-663a
significantly decreased the lactate production in the LNCaP cells
in vitro. Collectively, the data demonstrated that miR-663a
and miR-1225-5p functioned as tumor promoters in PCa progression.
The results provided a starting point for future research into the
function of miR-663a and miR-1225-5p and suggested that miR-663a
and miR-1225-5p upregulation may be involved in the progression of
PCa and this may promote the clinical application of miR-663a and
miR-1225-5p as PCa biomarkers.
In conclusion, the present study offered a simple
and efficient method for rapidly classifying miRNA function using
the CRISPR/Cas9 system in LNCaP cells. miR-205, miR-221, miR-222,
miR-30c, miR-224, miRNA-455-3p, miR-23b and miR-505 downregulation
in patients with PCa were experimentally validated to function as
tumor suppressors in PCa cells. The data from the present study,
for the first time to the best of the authors' knowledge, suggested
that the aberrant expression of miR-663a and miR-1225-5p may be
involved with PCa progression, implying their potential as
candidate markers for this type of cancer. However, the precise
roles of miR-663a and miR-1225-5p in accelerating the development
of PCa and promoting tumor progression require further
clarification. Limitations of the present study were that only one
cell line, LNCap, was investigated and that the CRISPR/Cas9 system
has a potential off-target problem, which may cause the cell
function to change by the knockout of another unexpected gene, thus
the detailed function of miRNA requires further study by
overexpression and knockout of miRNA in vitro and in
vivo.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Professor Yanqiong
Zhang (Institute of Chinese Materia Medica, China Academy of
Chinese Medical Sciences) who provided guidance and revised the
language.
Funding
The present study was supported by grants from
National Natural Science Foundation of China (grant nos. 81660426
and 81873608), the Natural Science Foundation of Guangdong Province
(grant nos. 2014A030304068, 2014A030310088 and 2014A020212471), the
High-level Innovative Talent Project of Guizhou Province in 2018
[grant no. (2018)5639] and the Science and Technology Project of
HuaDu District (grant no. HD15CXY005).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
FNJ, JGZ and WDZ designed the study and edited the
manuscript. YXL, WW, CYZ, GXC, YPW, ZZL, YY and ZDH performed the
experiments and analyzed the data. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bartel DP: MiRNAs: Target recognition and
regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ayub SG, Kaul D and Ayub T:
Microdissecting the role of miRNAs in the pathogenesis of prostate
cancer. Cancer Genet. 208:289–302. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao Y, Cong L and Lukiw WJ: Plant and
Animal microRNAs (miRNAs) and their potential for inter-kingdom
communication. Cell Mol Neurobiol. 38:133–140. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Volinia S, Calin GA, Liu CG, Ambs S,
Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, et
al: A microRNA expression signature of human solid tumors defines
cancer gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Porkka KP, Pfeiffer MJ, Waltering KK,
Vessella RL, Tammela TL and Visakorpi T: MicroRNA expression
profiling in prostate cancer. Cancer Res. 167:6130–6135. 2007.
View Article : Google Scholar
|
|
8
|
Plaisier CL, Pan M and Baliga NS: A
miRNA-regulatory network explains how dysregulated miRNAs perturb
oncogenic processes across diverse cancers. Genome Res.
22:2302–2314. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Song CJ, Chen H, Chen LZ, Ru GM, Guo JJ
and Ding QN: The potential of microRNAs as human prostate cancer
biomarkers: A meta-analysis of related studies. J Cell Biochem.
119:2763–2786. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He HC, Han ZD, Dai QS, Ling XH, Fu X, Lin
ZY, Deng YH, Qin GQ, Cai C, Chen JH, et al: Global analysis of the
differentially expressed miRNAs of prostate cancer in Chinese
patients. BMC Genomics. 14:7572013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cho SW, Kim S, Kim JM and Kim JS: Targeted
genome engineering in human cells with the Cas9 RNA-guided
endonuclease. Nat Biotechnol. 31:230–232. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cong L, Ran FA, Cox D, Lin S, Barretto R,
Habib N, Hsu PD, Wu X, Jiang W, Marraffini LA and Zhang F:
Multiplex genome engineering using CRISPR/Cas systems. Science.
339:819–823. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mali P, Yang L, Esvelt KM, Aach J, Guell
M, DiCarlo JE, Norville JE and Church GM: RNA-guided human genome
engineering via Cas9. Science. 339:823–826. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sanjana NE, Shalem O and Zhang F: Improved
vectors and genome-wide libraries for CRISPR screening. Nat
Methods. 11:783–784. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mussolino C, Morbitzer R, Lütge F,
Dannemann N, Lahaye T and Cathomen T: A novel TALE nuclease
scaffold enables high genome editing activity in combination with
low toxicity. Nucleic Acids Res. 39:9283–9293. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim D, Bae S, Park J, Kim E, Kim S, Yu HR,
Hwang J, Kim JI and Kim JS: Digenome-seq: Genome-wide profiling of
CRISPR-Cas9 off-target effects in human cells. Nat Methods.
12:237–243. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Baranwal S and Alahari SK: miRNA control
of tumor cell invasion and metastasis. Int J Cancer. 126:1283–1290.
2010.PubMed/NCBI
|
|
19
|
Saini S, Majid S and Dahiya R: Diet,
microRNAs and prostate cancer. Pharm Res. 27:1014–1026. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Szczyrba J, Löprich E, Wach S, Jung V,
Unteregger G, Barth S, Grobholz R, Wieland W, Stöhr R, Hartmann A,
et al: The microRNA profile of prostate carcinoma obtained by deep
sequencing. Mol Cancer Res. 8:529–538. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schaefer A, Jung M, Mollenkopf HJ, Wagner
I, Stephan C, Jentzmik F, Miller K, Lein M, Kristiansen G and Jung
K: Diagnostic and prognostic implications of microRNA profiling in
prostate carcinoma. Int J Cancer. 126:1166–1176. 2010.PubMed/NCBI
|
|
22
|
Gatenby RA and Gillies RJ: Why do cancers
have high aerobic glycolysis? Nat Rev Cancer. 4:891–899. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Airley RE and Mobasheri A: Hypoxic
regulation of glucose transport, anaerobic metabolism and
angiogenesis in cancer: Novel pathways and targets for anticancer
therapeutics. Chemotherapy. 53:233–256. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gandellini P, Folini M, Longoni N, Pennati
M, Binda M, Colecchia M, Salvioni R, Supino R, Moretti R, Limonta
P, et al: miR-205 exerts tumor-suppressive functions in human
prostate through down-regulation of protein kinase Cepsilon. Cancer
Res. 69:2287–2295. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Majid S, Dar AA, Saini S, Yamamura S,
Hirata H, Tanaka Y, Deng G and Dahiya R: MicroRNA-205-directed
transcriptional activation of tumor suppressor genes in prostate
cancer. Cancer. 116:5637–5649. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang N, Li Q, Feng NH, Cheng G, Guan ZL,
Wang Y, Qin C, Yin CJ and Hua LX: miR-205 is frequently
downregulated in prostate cancer and acts as a tumor suppressor by
inhibiting tumor growth. Asian J Androl. 15:735–741. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Verdoodt B, Neid M, Vogt M, Kuhn V,
Liffers ST, Palisaar RJ, Noldus J, Tannapfel A and
Mirmohammadsadegh A: MicroRNA-205, a novel regulator of the
anti-apoptotic protein Bcl2, is downregulated in prostate cancer.
Int J Oncol. 43:307–314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pang H and Yue X: MiR-205 serves as a
prognostic factor and suppresses proliferation and invasion by
targeting insulin-like growth factor receptor 1 in human cervical
cancer. Tumour Biol. 39:10104283177013082017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen S, Jin L, Nie S, Han L, Lu N and Zhou
Y: MiR-205 inhibits growth and invasion of neuroblastoma by
targeting cAMP responsive element binding protein 1. Oncol Res.
26:445–455. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tong AW, Fulgham P, Jay C, Chen P, Khalil
I, Liu S, Senzer N, Eklund AC, Han J and Nemunaitis J: MicroRNA
profile analysis of human prostate cancers. Cancer Gene Ther.
16:206–216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ling XH, Han ZD, Xia D, He HC, Jiang FN,
Lin ZY, Fu X, Deng YH, Dai QS, Cai C, et al: MicroRNA-30c serves as
an independent biochemical recurrence predictor and potential tumor
suppressor for prostate cancer. Mol Biol Rep. 41:2779–2788. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang J, Wang X, Wang Y, Peng R, Lin Z,
Wang Y, Hu B, Wang J and Shi G: Low expression of microRNA-30c
promotes prostate cancer cells invasion involved in downregulation
of KRAS protein. Oncol Lett. 14:363–368. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Huang Y, Li Y, Wang FF, Lv W, Xie X and
Cheng X: Over-expressed miR-224 promotes the progression of
cervical cancer via targeting RASSF8. PLoS One. 11:e01623782016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
He C, Wang L, Zhang J and Xu H:
Hypoxia-inducible microRNA-224 promotes the cell growth, migration
and invasion by directly targeting RASSF8 in gastric cancer. Mol
Cancer. 16:352017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu F, Liu Y, Shen J, Zhang G and Han J:
MicroRNA-224 inhibits proliferation and migration of breast cancer
cells by down-regulating Fizzled 5 expression. Oncotarget.
7:49130–49142. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao Y, Yan M, Yun Y, Zhang J, Zhang R, Li
Y, Wu X, Liu Q, Miao W and Jiang H: MicroRNA-455-3p functions as a
tumor suppressor by targeting eIF4E in prostate cancer. Oncol Rep.
37:2449–2458. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Negrete-Garcia MC, Ramírez-Rodriguez SL,
Rangel-Escareño C, Muñoz-Montero S, Kelly-García J,
Vázquez-Manríquez ME, Santillán P, Ramírez MM, Ramírez-Martínez G,
Ramírez- Venegas A and Ortiz-Quintero B: Deregulated MicroRNAs in
cancer-associated fibroblasts from front tumor tissues of lung
adenocarcinoma as potential predictors of tumor promotion. Tohoku J
Exp Med Oct. 246:107–120. 2018. View Article : Google Scholar
|
|
38
|
Rice MA, Ishteiwy RA, Magani F, Udayakumar
T, Reiner T, Yates TJ, Miller P, Perez-Stable C, Rai P, Verdun R,
et al: The microRNA-23b/-27b cluster suppresses prostate cancer
metastasis via Huntingtin-interacting protein 1-related. Oncogene.
35:4752–4761. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang Y, Xu X, Zhang M, Wang X, Bai X, Li
H, Kan L, Zhou Y, Niu H and He P: MicroRNA-663a is downregulated in
non-small cell lung cancer and inhibits proliferation and invasion
by targeting JunD. BMC Cancer. 16:3152016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zheng H, Zhang F and Lin X, Huang C, Zhang
Y, Li Y, Lin J, Chen W and Lin X: MicroRNA-1225-5p inhibits
proliferation and metastasis of gastric carcinoma through
repressing insulin receptor substrate-1 and activation of β-catenin
signaling. Oncotarget. 7:4647–4663. 2016. View Article : Google Scholar : PubMed/NCBI
|