Introduction
Thyroid cancer is one of the most commonly diagnosed
endocrine carcinomas and has an increasing incidence; the incidence
and mortality rates are ~9.61/100,000 people and 0.35/100,000
people, respectively, in China annually (1,2).
Papillary thyroid cancer (PTC) is the most common type of thyroid
cancer and accounts for ~90% of all thyroid cancer cases (3). Although the majority of patients with
PTC benefit from surgery, levothyroxine and radioactive iodine
therapy, certain patients still experience a high rate of
metastasis and recurrence (4–7).
Therefore, understanding the mechanisms underlying the development
and progression of PTC may aid the development of novel therapies
in the future.
The gene encoding the vitamin D receptor (VDR),
which is expressed by epithelial cells in both normal and malignant
thyroid glands, is located on chromosome 12q13.1 (8). Previous studies have revealed that
vitamin D, which can be hydroxylated to the active form
1,25(OH)2D3 (referred to as vitamin D or D3 in the
present study), activates VDR (9,10).
Furthermore, VDR expression is associated with cancer risk and
mortality in various types of cancer, including breast, lung and
colorectal cancer (11–14). Other studies suggested that the
vitamin D and VDR complex binds to p21 and p27 proteins, which
suppress DNA synthesis, inactivate mitogenic signals and inhibit
tumor progression (15–18). The vitamin D level is decreased in
PTC tissues compared with paired non-cancerous tissues, and is
found to inhibit the proliferation of PTC cells (19). Furthermore, a previous study has
revealed that VDR is upregulated in human PTC tissues compared with
normal thyroid tissues (20). In
addition, in our preliminary experiments, VDR was upregulated in
human PTC tissues compared with human thyroid tissues and was also
upregulated in PTC cell lines compared with thyroid follicular
cells (Pang et al unpublished data). Furthermore, the
Wnt/β-catenin signaling pathway was reported to serve a critical
role in mediating the development and progression of PTC (21,22).
However, whether VDR influences tumor progression and regulates the
Wnt/β-catenin signaling pathway through its interaction with
vitamin D in PTC cells remains unknown. Therefore, the present
study explored the role of vitamin D and the influence of VDR
knockdown (KD) on cell proliferation, apoptosis, invasion and
Wnt/β-catenin signaling in vitamin D-treated PTC cells.
Materials and methods
Cell culture
The human PTC cell lines MDA-T120 and MDA-T85 were
purchased from the American Type Culture Collection (ATCC), SNU-790
was purchased from the Korean Cell Line Bank (Korean Cell Line
Research Foundation) and IHH4 was purchased from the Japanese
Collection of Research Bioresources Cell Bank. The human thyroid
follicular cell line Nthy-ori 3-1 was purchased from Sigma-Aldrich
(Merck KGaA). 293T cells were purchased from the ATCC. Nthy-ori
3-1, MDA-T120, SNU-790 and MDA-T85 cells were cultured in RPMI-1640
medium (Sigma-Aldrich; Merck KGaA) supplemented with 10% fetal
bovine serum (FBS; Sigma-Aldrich; Merck KGaA). IHH4 and 293T cells
were cultured in DMEM (Sigma-Aldrich; Merck KGaA) supplemented with
10% FBS. All cells were maintained in a humidified incubator at
37°C and 5% CO2.
Gene expression omnibus (GEO)
datasets
The GEO datasets GSE33630 (23), GSE27155 (24), GSE3467 (25) and GSE3678 (26) were downloaded from the GEO database
(http://www.ncbi.nlm.nih.gov/geo)
(18) to assess VDR expression
levels in PTC and healthy tissues. The datasets comprised 49 PTC
tumor samples and 45 normal thyroids samples from GSE33630, 51 PTC
tumor samples and 4 normal samples from GSE27155, 9 PTC tumor
samples and 9 paired normal thyroid tissues from GSE3467, and 7 PTC
samples and 7 paired normal thyroid tissue samples from
GSE3678.
Lentivirus construction and
infection
KD-VDR and non-targeting negative control (KD-NC)
plasmids were cloned into the LV-H1/GFP&Puro plasmid and
constructed by Shanghai GenePharma Co., Ltd. The sequence for
VDR-KD was
5′-CGGCCTGAGATCAATCACATTTAACTCGAGTTAAATGTGATTGATCTCAGGTTTTT-3′ and
for KD-NC was
5′-CACCGTTCTCCGAACGTGTCACGTCGAAACGTGACACGTTCGGAGAA-3′. Then, 0.8 µg
KD-VDR and KD-NC plasmids, as well as the packaging plasmids
pRSV/pREV (Addgene, Inc.), pCMV–VSV-G (Addgene, Inc.) and
pMDLG/pRRE (Addgene, Inc.), were co-transfected into
2×105 293T cells using Lipofectamine® 2000
reagent (Thermo Fisher Scientific, Inc.) and the lentiviral
particles were harvested after 48 h by centrifugation (30,000 × g;
4°C; 4 h). A total of 1×105 SNU-790 and 1×105
IHH4 cells were infected with the lentiviral particles (MOI=20 and
40, respectively) in the presence of 8 µg/ml polybrene
(Sigma-Aldrich; Merck KGaA) at 37°C for 24 h, and then stably
infected cells were selected using puromycin (Thermo Fisher
Scientific, Inc.) for 7 days.
1,25(OH)2D3 treatment
KD-VDR and KD-NC cells that were maintained in
RPMI-1640 medium or DMEM (depending on cell type), supplemented
with 10% FBS containing 10 nM D3 (Sigma-Aldrich; Merck KGaA) were
referred to as the KD-VDR&vitD and KD-NC&vitD groups,
respectively. Whereas, KD-NC cells cultured without D3 were
referred to as the KD-NC group.
Cell Counting Kit-8 (CCK-8) assay
At 0, 24, 48 and 72 h post-treatment, cell
proliferation was determined using the CCK-8 assay (Dojindo
Molecular Technologies, Inc.), according to the manufacturers
instructions. Briefly, 5×103 cells/well were seeded into
96-well plates and incubated at 37°C. Subsequently, 10 µl CCK-8 and
90 µl serum-free medium (RPMI-1640 medium or DMEM depending on the
cell type) were added to each well and incubated at 37°C for 2 h.
The optical density (OD) value was measured using a microplate
reader at a wavelength of 450 nm (BioTek Instruments, Inc.).
Flow cytometric analysis of
apoptosis
At 48 h post-treatment, cells were stained using an
Annexin V-FITC Apoptosis Detection kit (BD Biosciences) according
to the manufacturers instructions. Briefly, cells in each group
were collected by centrifugation (850 × g; 5 min; room
temperature), and then washed with PBS. Subsequently, the cells
were suspended in 100 µl binding buffer, and incubated at 4°C with
5 µl Annexin V and 5 µl propidium iodide in the dark for 15 min.
Following the incubation, 400 µl binding buffer was added and the
apoptotic cells (early + late apoptotic cells were analyzed using a
CytoFLEX™ flow cytometer (Beckman Coulter, Inc.) and FlowJo 7.0
software (FlowJo LLC). PI−/Annexin V+ cells
represented early apoptotic cells and PI+/ Annexin
V+ cells represented late apoptotic cells.
Transwell Matrigel assay
The invasive ability of SNU-790 and IHH4 cells was
detected at 24 h post-transfection. Briefly, 4×104
transfected cells in serum-free media (RPMI-1640 medium or DMEM
depending on the cell type) were seeded into the upper chambers of
Transwell plates (pore size, 8-µm), precoated at 37°C for 1 h with
Matrigel (BD Biosciences). RPMI-1640 medium or DMEM (depending on
the cell type) supplemented with 10% FBS was plated into the lower
chamber. Following incubation for 24 h at 37°C, the non-invasive
cells remaining on the upper membrane were removed with cotton wool
and the invasive cells were fixed with 4% formaldehyde for 15 min
at room temperature and stained with 0.1% crystal violet for 15 min
at room temperature. Stained cells were visualized using an IX73
inverted microscope (magnification, ×200; Olympus Corporation) and
analyzed using Image-Pro Plus 6.0 software (Media Cybernetics,
Inc.).
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from the cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and was reverse transcribed into cDNA using the PrimeScript™
RT Master mix (Takara Biotechnology Co., Ltd.) at 37°C for 20 min,
and then 85°C for 5 sec. qPCR was subsequently performed using the
TB Green Fast qPCR mix (Takara Biotechnology Co., Ltd.) and the
following thermocycling parameters: Initial denaturation at 95°C
for 30 sec; followed by 40 cycles of denaturation at 95°C for 5
sec, annellation and extension at 61°C for 15 sec. VDR, Wnt3 and
CTNNB1 mRNA expression levels were quantified using the
2−ΔΔCq method (27) and
normalized to the internal reference gene GAPDH. The primers used
for the qPCR are presented in Table
I.
 | Table I.Primers used for reverse
transcription-quantitative PCR. |
Table I.
Primers used for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| Vitamin D | F:
GCCGCATCACCAAGGACAA |
| receptor | R:
TCTGAGCAGGAGGAGGAGGA |
| Wnt family | F:
AGCCTGACTTCCGTGCCATC |
| member 3 | R:
CTTCTCCGTCCTCGTGTTGTG |
| Catenin β1 | F:
GCCATTACAACTCTCCACAACCT |
|
| R:
GACAGATAGCACCTTCAGCACTC |
| GAPDH | F:
GACCACAGTCCATGCCATCAC |
|
| R:
ACGCCTGCTTCACCACCTT |
Western blotting
Total protein was extracted using RIPA buffer
(Sigma-Aldrich; Merck KGaA). The protein concentration in each
sample was measured using the Bicinchoninic Acid Protein Assay kit
(Pierce; Thermo Fisher Scientific, Inc.) and 20 µg protein/lane was
separated on 4–12% NuPAGE Bis-Tris Gels (Thermo Fisher Scientific,
Inc.). The separated proteins were transferred to polyvinylidene
fluoride membranes (EMD Millipore) and blocked with 5% BSA (Thermo
Fisher Scientific, Inc.) at 37°C for 2 h. The membranes were
subsequently incubated with the primary antibodies overnight at
4°C. Subsequently, the membranes were incubated with the secondary
antibody for 90 min at 37°C. Protein bands were visualized using
the Novex ECL Chemiluminescent Substrate Reagent kit (Thermo Fisher
Scientific, Inc.) and X-ray film (Kodak). The antibodies and
respective dilutions used in the present study are presented in
Table II.
| Table II.Antibodies used for western blotting
analysis. |
Table II.
Antibodies used for western blotting
analysis.
| A, Primary
antibodya | Cat. no. | Dilution |
|---|
| Anti-VDR; mouse
polyclonal | ab89626 | 1:1,000 |
| Anti-Wnt3; mouse
polyclonal | ab169175 | 1:1,000 |
| Anti-CTNNB1; mouse
monoclonal | ab22656 | 1:1,500 |
| Anti-GAPDH; mouse
monoclonal | ab8245 | 1:5,000 |
|
| B, Secondary
antibodya |
|
| Goat anti-mouse
IgG-HRP |
ab205719 |
1:10,000 |
Statistical analysis
Data are presented as the mean ± standard deviation.
Statistical analyses were performed using GraphPad Prism software
(version 7.02; GraphPad Software, Inc.). Comparisons between a
control group and other experimental groups were determined by
one-way ANOVA followed by Dunnetts test. Comparison between two
groups was assessed using the unpaired Students t-test or paired
t-test if appropriate. Multiple comparisons among groups were
assessed by one-way ANOVA followed by Tukeys test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of VDR in human PTC and
thyroid follicular cells
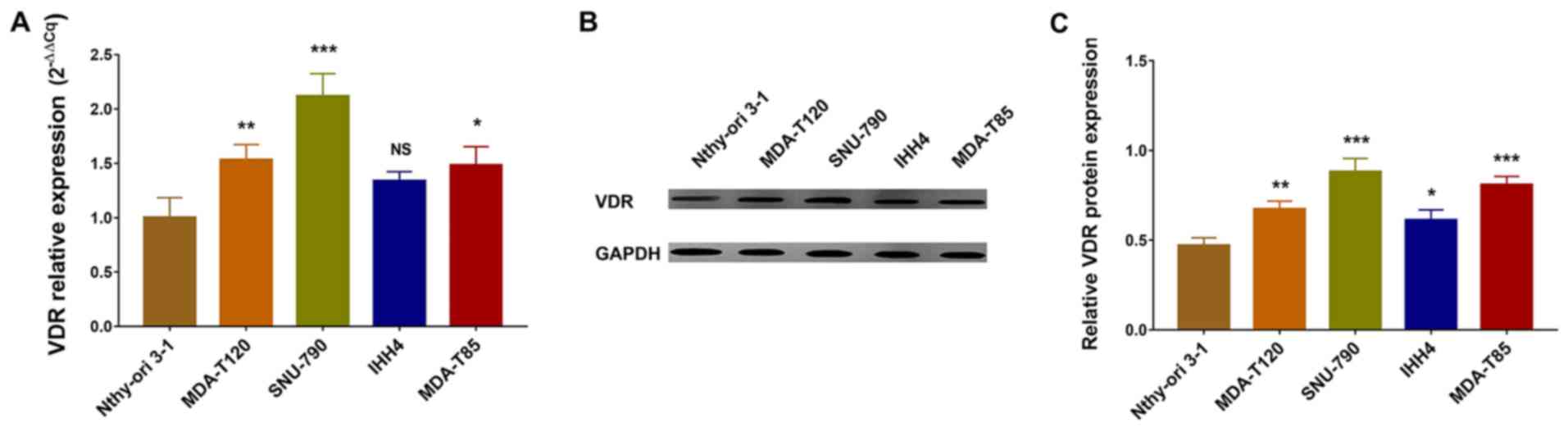
Comparisons between the control thyroid follicular
cell line Nthy-ori 3–1 and four PTC cell lines, revealed that VDR
mRNA expression levels were increased in MDA-T120, SNU-790 and
MDA-T85 cells compared with Nthy-ori 3-1 cells, but similar between
IHH4 and Nthy-ori 3-1 cells (Fig.
1A). Similarly, western blotting revealed that VDR protein
levels were increased in MDA-T120, SNU-790 and MDA-T85 cells
comparing with Nthy-ori 3-1 cells, but similar IHH4 and Nthy-ori
3-1 cells (Fig. 1B and C). These
data indicated that VDR was upregulated in human PTC cell lines
MDA-T120, SNU-790 and MDA-T85 compared with the control thyroid
follicular cell line Nthy-ori 3-1. Considering the observation that
VDR expression was upregulated in SNU-790 cells, and the finding
that VDR expression was similar between IHH4 cell line and thyroid
follicular cells, SNU-790 and IHH4 cells were chosen in the
following functional experiments to determine the influence of
KD-VDR in vitamin D-treated PTC cells.
Expression of VDR in PTC and healthy
tissues
GEO datasets were used to compare VDR expression in
PTC and healthy tissues (Fig.
S1). VDR expression was increased in PTC tissue compared with
healthy tissue in the GSE33630, GSE27155, GSE3467 and GSE3678
datasets. These data demonstrated that VDR was upregulated in PTC
tissues compared with healthy tissues.
KD-VDR in PTC cells
VDR expression was assessed in SNU-790 and IHH4
cells stably infected with KD-VDR and KD-NC plasmids. Comparison
between the two groups showed that VDR mRNA and protein expression
levels were decreased in KD-VDR SNU-790 cells compared with KD-NC
SNU-790 cells (Fig. 2A and B,
respectively). Similarly, VDR mRNA and protein expression levels
were decreased in KD-VDR IHH4 cells compared with KD-NC IHH4 cells
(Fig. 2C and D, respectively).
These results suggested that PTC cells stably infected with KD-VDR
or KD-NC lentiviruses were successfully established.
Effect of vitamin D and KD-VDR on cell
proliferation
To investigate the effect of KD-VDR on cell
proliferation in vitamin D-treated PTC cell lines, CCK-8 assays
were conducted; it was revealed that in SNU-790 (Fig. 3A) and IHH4 cells (Fig. 3B), cell proliferation was decreased
in the KD-NC&vitD group at 48 and 72 h compared with the KD-NC
group, and increased in the KD-VDR&vitD group at 48 and 72 h
compared with the KD-NC&vitD group. These data suggested that
KD-VDR infection attenuated the antiproliferative effect of vitamin
D treatment in PTC cells.
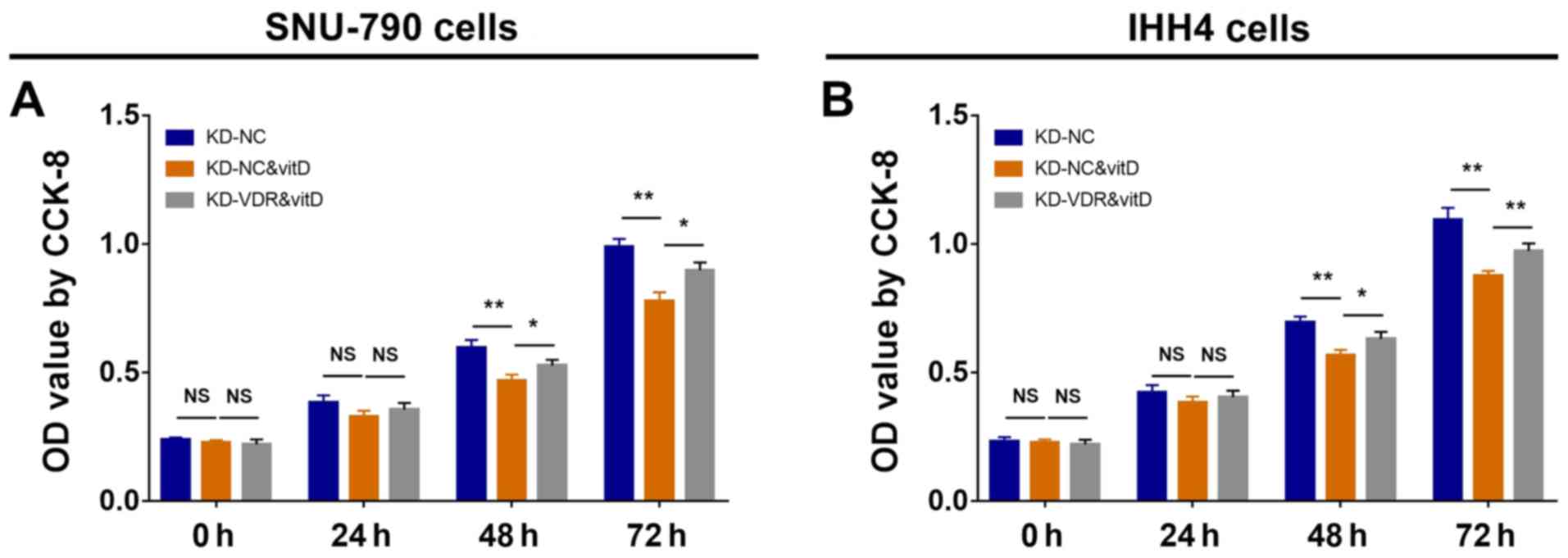 | Figure 3.Cell proliferation in SNU-790 and
IHH4 cells after KD-VDR and vitD treatment. CCK-8 assays for
proliferation were performed, and OD values were compared between
stably infected (A) SNU-790 and (B) IHH4 cells treated with or
without vitD at 0, 24, 48 and 72 h. Multiple comparisons among
groups were assessed by one-way ANOVA followed by Tukeys test.
*P<0.05; **P<0.01. CCK-8, Cell Counting Kit-8; KD, knockdown;
NC, negative control; NS, not significant; OD, optical density;
VDR, vitamin D receptor; vitD, vitamin D. |
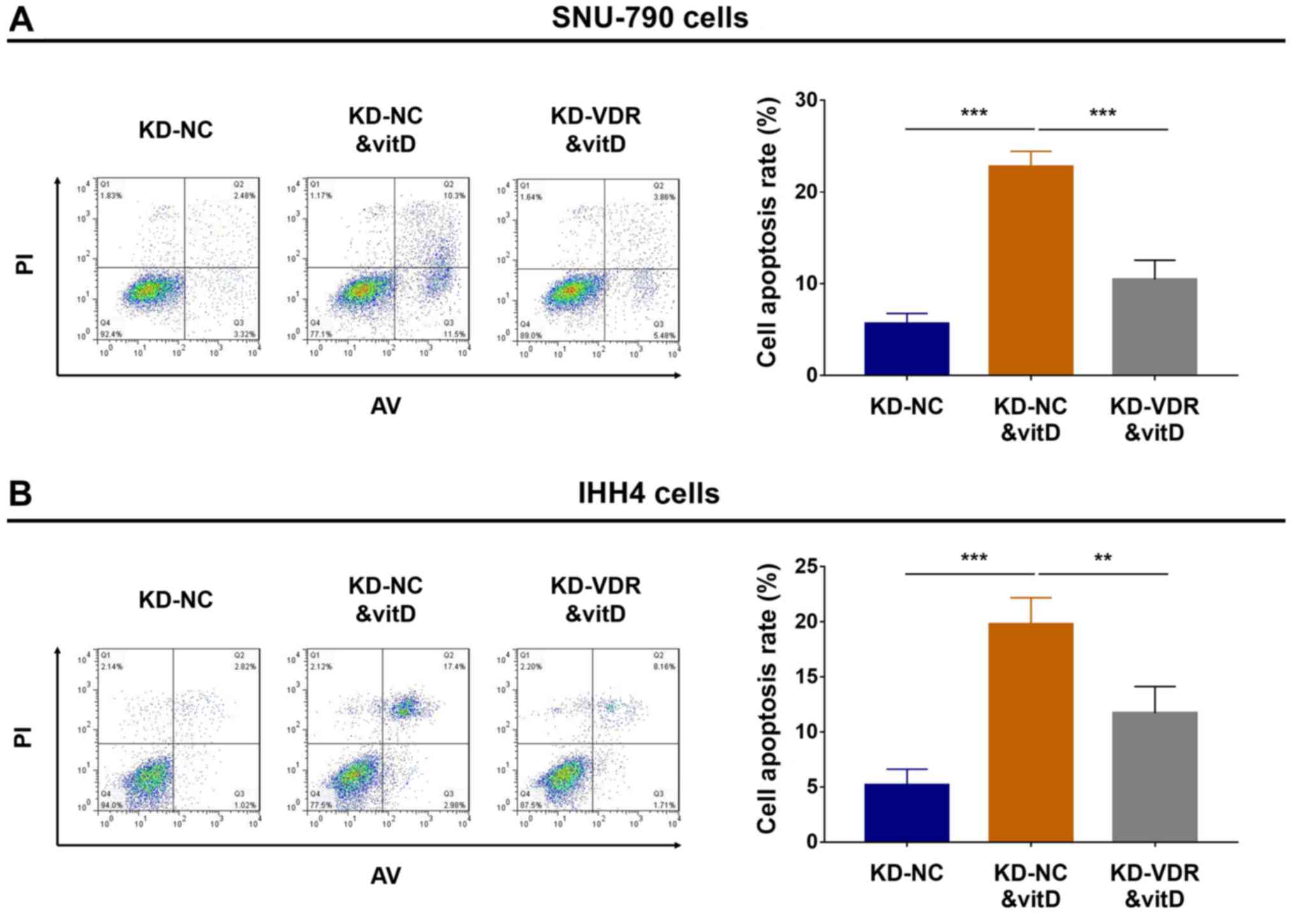
Effect of vitamin D and KD-VDR on
apoptosis
To determine the effect of KD-VDR on the rate of
cell apoptosis in vitamin D-treated PTC cell lines, flow cytometric
analysis of apoptosis was performed; in SNU-790 (Fig. 4A) and IHH4 cells (Fig. 4B), cell apoptosis was increased in
the KD-NC&vitD group compared with the KD-NC group, whereas
cell apoptosis was decreased in the KD-VDR&vitD group compared
with the KD-NC&vitD group. These data indicated that KD-VDR
attenuated the pro-apoptotic effect of vitamin D treatment in PTC
cells.
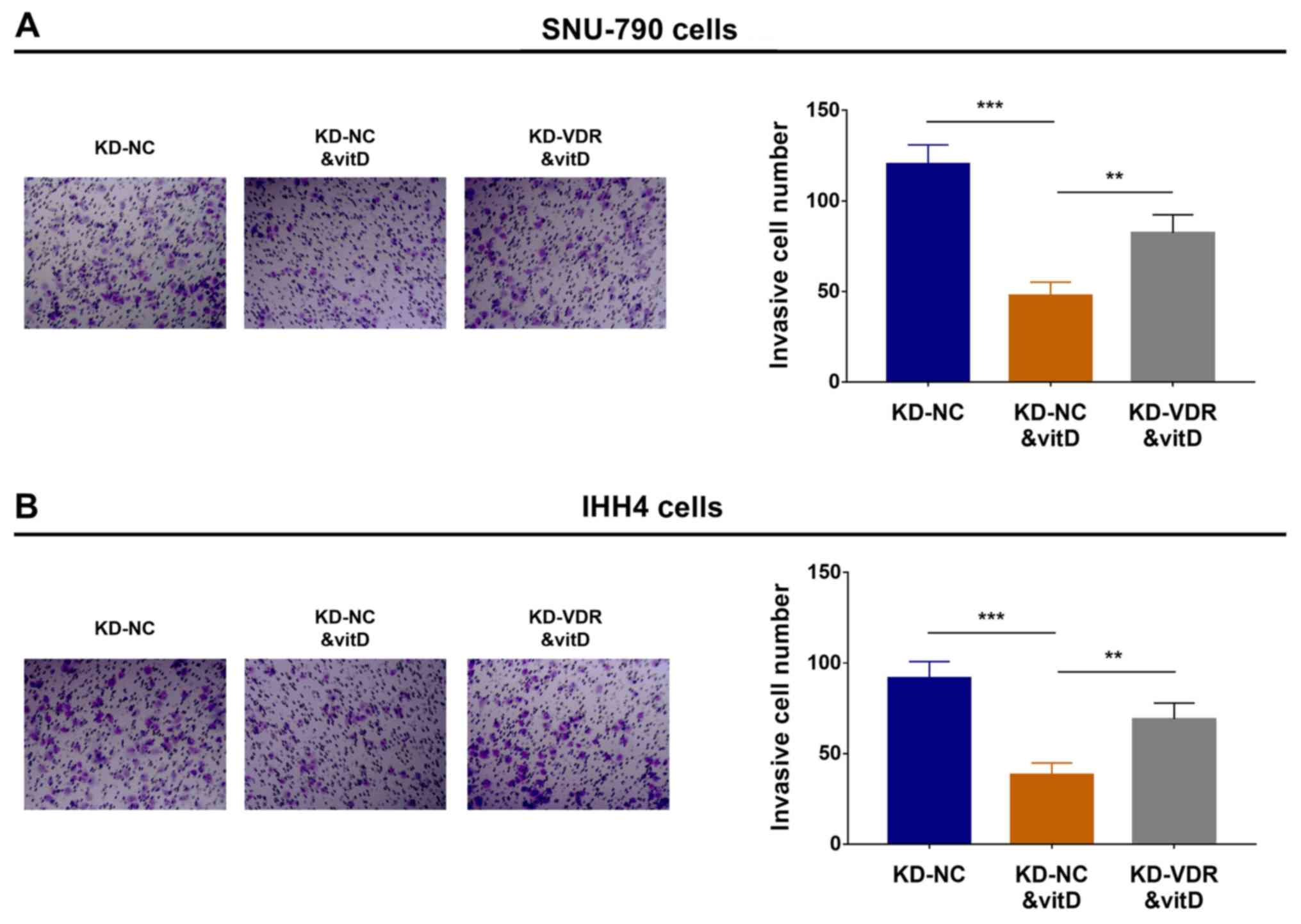
Effect of vitamin D and KD-VDR on cell
invasion
To elucidate the effect of KD-VDR on cell invasion
in vitamin D-treated PTC cell lines, Transwell Matrigel assays were
performed, which revealed that in SNU-790 (Fig. 5A) and IHH4 (Fig. 5B) cells, invasion was decreased in
the KD-NC&vitD group compared with the KD-NC group, but
increased in the KD-VDR&vitD group compared with the
KD-NC&vitD group. These data suggested that KD-VDR attenuated
the anti-invasive effect of vitamin D treatment in PTC cells.
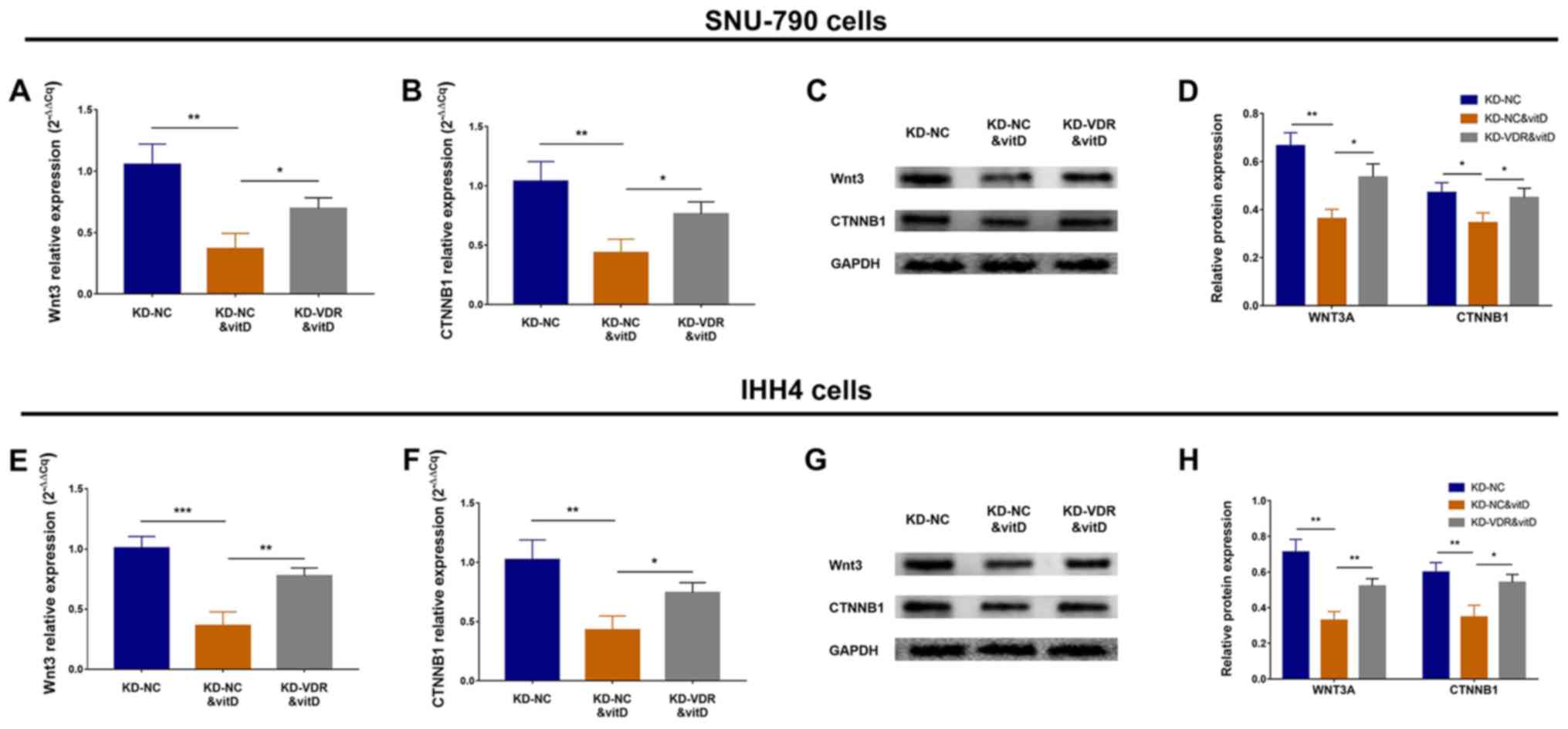
Effect of vitamin D and KD-VDR on the
Wnt/β-catenin signaling pathway
To determine the effect of KD-VDR on the
Wnt/β-catenin signaling pathway in vitamin D-treated PTC cell
lines, RT-qPCR and western blotting assays were performed. The
results revealed that in SNU-790 cells, Wnt3 and CTNNB1 mRNA
expression levels were decreased in the KD-NC&vitD group
compared with the KD-NC group, but increased in the KD-VDR&vitD
group compared with the KD-NC&vitD group (Fig. 6A and B). Wnt3 and CTNNB1 protein
expression levels exhibited a similar trend (Fig. 6C and D). Similarly, in IHH4 cells,
Wnt3 and CTNNB1 mRNA expression levels were decreased in the
KD-NC&vitD group compared with the KD-NC group, but increased
in the KD-VDR&vitD group compared with the KD-NC&vitD group
(Fig. 6E and F), and Wnt3 and
CTNNB1 protein expression levels exhibited a similar trend
(Fig. 6G and H). These data
indicated that KD-VDR attenuated the inhibitory effect of vitamin D
on the Wnt/β-catenin signaling pathway in PTC cells.
Discussion
Vitamin D is synthesized through a series of steps,
starting from a cholesterol precursor molecule that is transformed
into the vitamin D hormone precursor (vitamin D3) and
subsequently hydroxylated to its most active hormone form,
1,25(OH)2D3, in the liver and kidney (14). Vitamin D binds to VDR when it
enters the cells, forming a complex that is reported to influence
cell cycle regulation by decreasing DNA synthesis, inactivating
mitogenic signals, reducing expression of c-Myc and inhibiting cell
differentiation, which inhibit tumor formation in several types of
cancer, including breast, colorectal and lung cancer (13,28,29).
For example, in breast cancer, vitamin D modulates the
intracellular kinase pathways (including p38 mitogen activated
protein kinase and ERK), represses the expression of
proto-oncogenes (such as c-Myc) and decreases the proliferation of
breast cancer cells (15,30). In PTC, vitamin D is associated with
decreased PTC risk, and vitamin D inhibits PTC development and
progression (19). Furthermore,
VDR is reported to be upregulated in PTC tissues compared with
normal thyroid tissue (20,31).
However, the role of VDR in the development and progression of PTC
has not been fully elucidated. The present study analyzed VDR
expression in human PTC and normal thyroid follicular cells, and
revealed that VDR mRNA and protein expression levels were increased
in PTC cells compared with thyroid follicular cells, which was
consistent with previous studies (20,31).
Previous studies had revealed that VDR has the
potential to regulate cancer cellular activities and influence
tumor progression and metabolism by binding to vitamin D (15,20,32,33).
For example, in vitamin D-treated breast cancer cells, silencing of
VDR promotes cancer cell motility and invasiveness by increasing
the expression of proteins involved in cell adhesion,
proliferation, cytoskeletal organization (33). The present study explored the
modulatory effects of vitamin D on cellular activities in PTC
cells, and further investigated whether KD-VDR affected the actions
of vitamin D in PTC cells. In the current study, SNU-790 and IHH4
cells were selected for the functional experiments, based on the
rational that among the MDA-T120, MDA-T85, SNU-790 and IHH4 cell
lines, VDR was more highly expressed in SNU-790 cells compared with
the human thyroid follicular cells Nthy-ori 3-1. Therefore, this
cell line was chosen as it could show an increased phenotype after
VDR knockdown over the remaining cell lines. By contrast, VDR
expression was similar between IHH4 cell line and the human thyroid
follicular cells. Therefore, IHH4 cell line was selected to explore
the regulatory role of VDR knockdown on this cell line expressing
lower VDR levels compared to the remaining PTC cell lines. In the
present study, the two PTC cell lines were selected to exclude the
cell line-dependent phenomenon for the investigation of the effects
of VDR knockdown on cell proliferation, apoptosis, invasion and
Wnt/β-catenin signaling in vitamin D-treated PTC cells. In the
current study, KD-VDR attenuated the antiproliferative,
pro-apoptotic and anti-invasive effects of vitamin D in PTC cells.
In a previous study, vitamin D was shown to reduce cyclin-dependent
kinase activity, regulate the expression of the plasminogen
activator system and increase E-cadherin expression in prostate
cancer (14). Vitamin D may
therefore inhibit PTC cell proliferation and invasion in a similar
manner. A different previous study revealed that VDR was
upregulated in PTC cells compared with thyroid follicular cells and
that the vitamin D and VDR complex exhibited antitumor properties
(34). Therefore, KD-VDR might
decrease the anti-tumor effect by reducing the number of vitamin
D/VDR complexes in PTC cells. The present study revealed that
KD-VDR may attenuate the effect of vitamin D on the oncogenic
Wnt/β-catenin signaling pathway in PTC and inhibit cell
proliferation and invasion but promote cell proliferation.
A previous study revealed that cell proliferation,
apoptosis and epithelial-mesenchymal transition are mediated by
VDR-activated vitamin D and the Wnt/β-catenin signaling pathway in
renal cancer (32). Furthermore,
in uterine fibroid cells, vitamin D reduced the expression levels
of Wnt4 and β-catenin, whereas VDR silencing promoted the
expression levels of Wnt4 and β-catenin, further promoting cell
proliferation and extracellular matrix production (17). Furthermore, in breast and prostate
cancer, VDR silencing was associated with the attenuation of the
Wnt/β-catenin signaling pathway, along with the downregulation of
downstream genes such as cyclin D1 and interleukin-6 (35). In addition, studies have
demonstrated the involvement of the Wnt/β-catenin signaling pathway
in cell growth, invasion and metastasis in several types of cancer,
including PTC (36). A previous
study revealed that activation of the Wnt/β-catenin signaling
pathway contributes to the progression of PTC by promoting cell
proliferation and invasion (22).
Therefore, the present study explored the regulatory effect of
vitamin D on Wnt/β-catenin signaling, and to further investigate
the regulatory effect of KD-VDR on the Wnt/β-catenin signaling
pathway in vitamin D-treated PTC cells. The results revealed that
vitamin D suppressed the expression of Wnt3 and CTNBB1 in PTC
cells, and that KD-VDR attenuated the inhibitory effect of vitamin
D on the expression of Wnt3 and CTNBB1 in PTC cells. This suggested
that KD-VDR might attenuate the antiproliferative, pro-apoptotic
and anti-invasive effects of vitamin D by activating the
Wnt/β-catenin signaling pathway in PTC cells. However, the
mechanism by which VDR alone regulates PTC cells was not
investigated in the present study. However, similar to mechanisms
reported in breast and prostate cancer, inhibiting the VDR may
activate specific downstream promotors of Wnt/β-catenin signaling
pathway in PTC cells, which further stimulates Wnt/β-catenin
signaling (34,37).
Nonetheless, there are several limitations to the
present study; i) The effect of KD-VDR was only investigated in
vitamin D-treated PTC cells, therefore further studies are required
to determine the effect of KD-VDR in vitamin D-treated normal
thyroid cell lines; and ii) the current study did not investigate
the effect of VDR alone in PTC cells, hence further cellular
experiments are required.
In conclusion, KD-VDR attenuated the
antiproliferative, pro-apoptotic and anti-invasive effect of
vitamin D in PTC cells possibly by activating the Wnt/β-catenin
signaling pathway. The results obtained in the present study
suggested that VDR may be a novel and promising therapeutic target
in the treatment of PTC, which might be applied in future clinical
practice and improve the clinical outcomes of patients with
PTC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the Subject of
Heilongjiang Provincal Health and Family Planning Commission (grant
no. 2018043) and the Youth Project of Haiyan Fund (grant no.
JJQN2017-11).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors contributions
RP made substantial contributions to the design of
the present study. Data acquisition and interpretation was
conducted by RP, YX, XH, BL and JY. PR and JY critically revised
the manuscript for intellectual content. All authors agree to be
accountable for all aspects of the work in ensuring that questions
related to the accuracy or integrity of the work are appropriately
investigated and resolved. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wang J, Yu F, Shang Y, Ping Z and Liu L:
Thyroid cancer: Incidence and mortality trends in China, 2005–2015.
Endocrine. 68:163–173. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Haddad RI, Nasr C, Bischoff L, Busaidy NL,
Byrd D, Callender G, Dickson P, Duh QY, Ehya H, Goldner W, et al:
NCCN Guidelines insights: Thyroid carcinoma, version 2.2018. J Natl
Compr Canc Netw. 16:1429–1440. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim HI, Jang HW, Ahn HS, Ahn S, Park SY,
Oh YL, Hahn SY, Shin JH, Kim JH, Kim JS, et al: High serum TSH
level is associated with progression of papillary thyroid
microcarcinoma during active surveillance. J Clin Endocrinol Metab.
103:446–451. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Amouri W, Charfeddine S, Charfi S, Jardak
I, Boudawara T and Guermazi F: Clinicopathological features and
outcomes after radioactive iodine treatment of oncocytic
well-differentiated thyroid carcinomas. Nucl Med Commun.
40:888–893. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Price AK, Randle RW, Schneider DF, Sippel
RS and Pitt SC: Papillary thyroid microcarcinoma: Decision-making,
extent of surgery, and outcomes. J Surg Res. 218:237–245. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lee YC, Na SY, Park GC, Han JH, Kim SW and
Eun YG: Occult lymph node metastasis and risk of regional
recurrence in papillary thyroid cancer after bilateral prophylactic
central neck dissection: A multi-institutional study. Surgery.
161:465–471. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Uitterlinden AG, Fang Y, Van Meurs JB,
Pols HA and Van Leeuwen JP: Genetics and biology of vitamin D
receptor polymorphisms. Gene. 338:143–156. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bizzaro G, Antico A, Fortunato A and
Bizzaro N: Vitamin D and autoimmune diseases: Is Vitamin D receptor
(VDR) polymorphism the culprit? Isr Med Assoc J. 19:438–443.
2017.PubMed/NCBI
|
|
10
|
Hewison M: Vitamin D and the
intracrinology of innate immunity. Mol Cell Endocrinol.
321:103–111. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huss L, Butt ST, Borgquist S, Elebro K,
Sandsveden M, Rosendahl A and Manjer J: Vitamin D receptor
expression in invasive breast tumors and breast cancer survival.
Breast Cancer Res. 21:842019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li M, Liu X, Liu N, Yang T, Shi P, He R
and Chen M: Association between polymorphisms of Vitamin D receptor
and lung cancer susceptibility: Evidence from an updated
Meta-analysis. J Cancer. 10:3639–3649. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Joanna B, Jolanta B, Agnieszka G, Diana HZ
and Krystyna S: Vitamin D, linoleic acid, arachidonic acid and
COX-2 in colorectal cancer patients in relation to disease stage,
tumour localisation and disease progression. Arab J Gastroenterol.
20:121–126. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Trump DL and Aragon-Ching JB: Vitamin D in
prostate cancer. Asian J Androl. 20:244–252. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Feldman D, Krishnan AV, Swami S,
Giovannucci E and Feldman BJ: The role of vitamin D in reducing
cancer risk and progression. Nat Rev Cancer. 14:342–357. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Skrajnowska D and Bobrowska-Korczak B:
Potential Molecular Mechanisms of the Anti-cancer Activity of
Vitamin D. Anticancer Res. 39:3353–3363. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Al-Hendy A, Diamond MP, Boyer TG and
Halder SK: Vitamin D3 inhibits Wnt/β-catenin and mTOR signaling
pathways in human uterine fibroid cells. J Clin Endocrinol Metab.
101:1542–1551. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Samuel S and Sitrin MD: Vitamin Ds role in
cell proliferation and differentiation. Nutr Rev. 66 (Suppl
2):S116–S124. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang T, Zhang H, He L, Wang Z, Dong W,
Sun W and Zhang P: Potential use of 1–25-dihydroxyvitamin D in the
diagnosis and treatment of papillary thyroid cancer. Med Sci Monit.
24:1614–1623. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Izkhakov E, Somjen D, Sharon O, Knoll E,
Aizic A, Fliss DM, Limor R and Stern N: Vitamin D receptor
expression is linked to potential markers of human thyroid
papillary carcinoma. J Steroid Biochem Mol Biol. 159:26–30. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang J, Gill AJ, Issacs JD, Atmore B,
Johns A, Delbridge LW, Lai R and McMullen TP: The Wnt/β-catenin
pathway drives increased cyclin D1 levels in lymph node metastasis
in papillary thyroid cancer. Hum Pathol. 43:1044–1050. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Y, Huang H, Hu F, Li J, Zhang L and
Pang H: CITED1 contributes to the progression of papillary thyroid
carcinoma via the Wnt/β-catenin signaling pathway. Onco Targets
Ther. 12:6769–6777. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tomas G, Tarabichi M, Gacquer D, Hebrant
A, Dom G, Dumont JE, Keutgen X, Fahey TJ III, Maenhaut C and
Detours V: A general method to derive robust organ-specific gene
expression-based differentiation indices: Application to thyroid
cancer diagnostic. Oncogene. 31:4490–4498. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Giordano TJ, Kuick R, Thomas DG, Misek DE,
Vinco M, Sanders D, Zhu Z, Ciampi R, Roh M, Shedden K, et al:
Molecular classification of papillary thyroid carcinoma: Distinct
BRAF, RAS, and RET/PTC mutation-specific gene expression profiles
discovered by DNA microarray analysis. Oncogene. 24:6646–6656.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He H, Jazdzewski K, Li W, Liyanarachchi S,
Nagy R, Volinia S, Calin GA, Liu CG, Franssila K, Suster S, et al:
The role of microRNA genes in papillary thyroid carcinoma. Proc
Natl Acad Sci USA. 102:19075–19080. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhu X, Yao J and Tian W: Microarray
technology to investigate genes associated with papillary thyroid
carcinoma. Mol Med Rep. 11:3729–3733. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tagliabue E, Raimondi S and Gandini S:
Vitamin D, cancer risk, and mortality. Adv Food Nutr Res. 75:1–52.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Robsahm TE, Tretli S, Torjesen PA,
Babigumira R and Schwartz GG: Serum 25-hydroxyvitamin D levels
predict cancer survival: A prospective cohort with measurements
prior to and at the time of cancer diagnosis. Clin Epidemiol.
11:695–705. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rohan JN and Weigel NL:
1Alpha,25-dihydroxyvitamin D3 reduces c-Myc expression, inhibiting
proliferation and causing G1 accumulation in C4-2 prostate cancer
cells. Endocrinology. 150:2046–2054. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Choi JY, Yi JW, Lee JH, Song RY, Yu H,
Kwon H, Chai YJ, Kim SJ and Lee KE: VDR mRNA overexpression is
associated with worse prognostic factors in papillary thyroid
carcinoma. Endocr Connect. 6:172–178. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu KH, Fu J, Zhou N, Yin W, Yang YY,
Ouyang SX and Liang YM: 1,25-Dihydroxyvitamin D3 prevents
epithelial-mesenchymal transition of HMrSV5 human peritoneal
mesothelial cells by inhibiting histone deacetylase 3 (HDAC3) and
increasing Vitamin D receptor (VDR) expression through the
Wnt/β-catenin signaling pathway. Med Sci Monit. 25:5892–5902. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Horas K, Zheng Y, Fong-Yee C, Macfarlane
E, Manibo J, Chen Y, Qiao J, Gao M, Haydar N, McDonald MM, et al:
Loss of the vitamin d receptor in human breast cancer cells
promotes epithelial to mesenchymal cell transition and skeletal
colonization. J Bone Miner Res. 34:1721–1732. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim D: The role of Vitamin D in thyroid
diseases. Int J Mol Sci. 18:19492017. View Article : Google Scholar
|
|
35
|
Zheng Y, Trivedi T, Lin RC, Fong-Yee C,
Nolte R, Manibo J, Chen Y, Hossain M, Horas K, Dunstan C, et al:
Loss of the vitamin D receptor in human breast and prostate cancers
strongly induces cell apoptosis through downregulation of
Wnt/β-catenin signaling. Bone Res. 5:170232017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang X, Li M, Zuo K, Li D, Ye M, Ding L,
Cai H, Fu D, Fan Y and Lv Z: Upregulated miR-155 in papillary
thyroid carcinoma promotes tumor growth by targeting APC and
activating Wnt/β-catenin signaling. J Clin Endocrinol Metab.
98:E1305–E1313. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Welsh J: Vitamin D and breast cancer: Past
and present. J Steroid Biochem Mol Biol. 177:15–20. 2018.
View Article : Google Scholar : PubMed/NCBI
|