Introduction
Colorectal cancer (CRC) is one of the most common
digestive carcinomas. According to the statistics, there were
>1.8 million new colorectal cancer cases and 881,000 deaths
occurred in 2018 worldwide (1).
Although therapeutic strategies have been considerably improved,
the 5-year survival rate of patients with CRC with distant
metastasis was estimated to be 11.7% due to the unclear molecular
mechanisms underlying tumor metastasis and progression are unclear
(2,3). Therefore, further investigation of the
underlying molecular mechanisms associated with the progression of
CRC is required for improving disease prognosis.
Long non-coding RNAs (lncRNAs) are a type of
non-coding RNA with an approximate length of >200 nucleotides
(4). Previously, it was confirmed
that lncRNAs are dysregulated in various human cancers, suggesting
that they may participate in several biological processes in
cancers, including cancer progression (5). For instance, Hua et al
(6) demonstrated that
overexpression of NNT-AS1 could enhance cell proliferation and
invasion via the Wnt/b-catenin signaling pathway. Ge et al
(7) revealed that CAR10 expression
is increased in lung adenocarcinoma and that it is correlated with
the proliferation, migration and invasiveness of lung
adenocarcinoma cells. Small nucleolar RNA host gene 20 (SNHG20),
located on chromosome 17q25.2, was initially identified as an
oncogenic gene involved in hepatocellular carcinoma (8). Subsequent studies demonstrated that
SNHG20 is associated with the progression of various types of
cancers. For example, SNHG20 expression is significantly
upregulated in non-small cell lung cancer (NSCLC), suggesting that
it acts as an oncogene (9). SNHG20
promotes oral squamous cell carcinoma cell proliferation and tumor
growth by competitively sponging microRNA (miR/miRNA)-197 (10). Wu et al (11) demonstrated that SNHG20 upregulation
contributes to the growth and metastasis of prostate cancer cells
via the miR-6516-5p/SCGB2A1 pathway. In addition, a previous study
indicated that SNHG20 expression is notably upregulated in
colorectal cancer. Elevated levels of SNHG20 are correlated with
advanced tumor node metastasis (TNM) stage and poor survival
(12). However, the pivotal roles
and potential molecular mechanisms of SNHG20 regulation in CRC have
not been fully elucidated. Therefore, the present study aimed to
investigate the underlying molecular mechanisms of SNHG20 in CRC
progression.
In the present study, the data revealed that the
expression levels of SNHG20 were significantly elevated in CRC
tissues and cell lines. In addition, functional experiments
indicated that SNHG20 knockdown inhibited CRC cell proliferation,
migration and invasion. It is important to note that silencing of
SNHG20 expression significantly inhibited tumor growth and
metastasis in vivo. Knockdown of SNHG20 expression
suppressed CRC progression. This was achieved by suppressing STAT3
expression via sponging miR-495.
Materials and methods
Tissue samples and cell culture
A total of 68 matched CRC tissues and corresponding
normal tissues were collected at The First Affiliated Hospital of
the University of South China (Hengyang, China). Signed informed
consent had been provided by all patients. None of the patients
received chemotherapy or radiotherapy prior to surgical resection.
The present study was approved by the Medical Ethics Committee of
The First Affiliated Hospital of University of South China
(approval no. LL20181102013).
Normal colorectal epithelial cells (FHC) and human
CRC cells (HCT116, SW480, SW620, HT-29) were provided by The Cell
Bank of Type Culture Collection of the Chinese Academy of Sciences.
The cell lines were authenticated by STR profiling. The cells were
grown in RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (Invitrogen; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin antibiotics
(Invitrogen; Thermo Fisher Scientific, Inc.) in the presence of 5%
CO2 at 37°C.
Cell transfection
Lentiviral knockdown vector specifically targeting
SNHG20 [small interfering (si) RNA-SNHG20] and negative control
(si-NC) were provided by Shanghai GenePharma Co., Ltd. miR-495
mimics, miR-495 inhibitor and their NCs were purchased from
Shanghai GenePharma Co., Ltd. STAT3 overexpression plasmid
[pcDNA3.1(+) STAT3] and its NC [pcDNA3.1(+)] were constructed by
Shanghai GenePharma Co., Ltd. Before transfection, cells were
subcultured in 6-well plates (Corning, Inc.) ~50% confluence
(3×105). RNAs (100 nM) or miR-495 mimics (50 nM) or
miR-495 inhibitor (150 nM) or plasmids (1.5 µg per well) were
transfected into cells. The lentivirus was prepared according to
the User Manual of the Lenti-Pac™ HIV Expression Packaging kit
(GeneCopoeia, Inc.). The viral packaging was performed in 293T
cells after co-transfection of the Lv3-si-SNHG20, or empty
lentiviral vector, and Lenti-Pac™ HIV packaging mix (GeneCopoeia,
Inc.) using EndoFectin™ Lenti transfection reagent (GeneCopoeia,
Inc.). The medium containing the retroviral supernatant was
harvested 48–72 h after transfection. The retroviruses were added
into HCT116 and SW480 cells for the infection. The cells were
selected by culture in the presence of puromycin (5 µg/ml;
Invitrogen; Thermo Fisher Scientific, Inc.) for up to 2 weeks. The
sequences were as follows: si-SNHG20, 5′-GCCUAGGAUCAUCCAGGUUTT-3′;
si-NC, 5′-UUCUCCGAACGUGUCACGUU-3′; miR-495 mimic,
5′-UGUGACGAAACAAACAUGGUGCACU-3′; miR-NC,
5′-CAGUACUUUUGUGUAGUACAA-3′; miR-495 inhibitor,
5′-GCCGAATTCTGGCTGCTATGATCTGAACT-3′; and inhibitor NC,
5′-CAGUACUUUUGUGUAGUACAA-3′. In order to establish the
STAT3-overexpression cell model, the full-length cDNAs of human
STAT3 were cloned into the pc-DNA3.1 vector. Transfection was
performed when the cell density was 50%. All cells transfections
were conducted using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.). Transfection was performed at room
temperature for 30 min. After incubation for 48 h, the cells were
collected for the subsequent experiments.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was isolated from tissue and cells with
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The mRNA expression levels of SNHG20 and STAT3 were detected
following RNA extraction and RT-qPCR. Total RNA was reverse
transcribed into cDNA using a Reverse Transcription kit (Takara
Bio, Inc.). The PCR protocol consisted of cycling at 94°C for 3
min, followed by 30 cycles of 94°C for 30 sec, 55°C for 30 sec and
72°C for 60 sec and a final extension at 72°C for 5 min. RT-qPCR
was carried out with SYBR Green (Takara Bio, Inc.). The expression
levels of miR-495 were measured using the miRNA isolation kit
(Guangzhou RiboBio Co., Ltd.). GAPDH and U6 were used as internal
controls for RNAs and miRNAs, respectively. The relative expression
levels of the genes were determined using the 2−ΔΔCq
method (13). The primers used are
shown in Table I.
 | Table I.Primers used for reverse
transcription-quantitative PCR analysis. |
Table I.
Primers used for reverse
transcription-quantitative PCR analysis.
| Gene | Primer sequences
(5′→3′) |
|---|
| SNHG20 | F:
ATGGCTATAAATAGATACACG |
|
| R:
GGTACAAACAGGGAGGGA |
| miR-495 | F:
GCGGAAACAAACATGGTGCA |
| U6 | F: CTCGCTTCGGCA
GCACA |
| STAT3 | F:
CAGCAGCTTGACACACGGTA |
|
| R:
AAACACCAAAGTGGCATGTGA |
| GAPDH | F:
GCGAGATCGCACTCATCATCT |
|
| R:
TCAGTGGTGGACCTGACC |
Cell viability assays
The cell proliferative rates were assessed by MTS
and colony formation assays. The transfected cells were seeded in
96-well plates (2×103 cells/well) with five replicate
wells. At the time of harvest, 10 µl MTS was added to each well.
Following incubation for 2 h, the absorbance was read at 490 nm on
a microplate reader (Bio-Rad Laboratories, Inc.). The colony
formation assay was performed with transfected cells seeded in
6-well plates (800/well) and incubated for 2 weeks. Subsequently,
the cells were fixed with ethanol at room temperature for 10 min
and stained with 0.1% crystal violet at room temperature for 10
min. when a single colony cell number >50 was achieved. Finally,
colony counting was performed under an inverted light microscope
(magnification, ×200; CKX41, Olympus).
Transwell migration and invasion
assays
The Transwell assay was carried out to assess the
migratory (without Matrigel) and invasive (with Matrigel; BD
Biosciences) ability of the cells. The BD Matrigel was placed in a
4°C refrigerator for 24 h. It was diluted to 1:9 ratios with
RPMI-1640. The upper Transwell chamber was filled with 50 µl of the
diluent. The Transwell insert was incubated for 3 h at 37°C until
the Matrigel clotted. Briefly, the transfected cells
(5×104 cells/well) were seeded into the upper Transwell
chamber with 250 µl serum-free RPMI-1640 medium, and the lower
chamber was filled with 750 µl RPMI-1640 medium supplemented with
10% FBS. Following incubation for 48 h at 37°C, the invaded cells
were fixed with ethanol at room temperature for 10 min and stained
with 0.1% crystal violet at room temperature for 10 min. The
stained cells were counted using an inverted light microscope
(magnification, ×200; CKX41, Olympus Corporation).
Dual-luciferase reporter assay
The StarBase software V2.0 (http://starbase.sysu.edu.cn) was applied to analyze
the potential miRNA target of SNHG20. The TargetScan software was
performed to predict the potential mRNA target of miR-495. The
wild-type (WT) or mutant (MUT) SNHG20 sequences containing the
miR-495 binding sites were cloned into the pmir-GLO Dual-luciferase
vector (Promega Corporation) and co-transfected with the miR-495
mimics or corresponding control sequences into the cells with
Lipofectamine 2000. Following transfection, the cells were
incubated for 48 h and the luciferase activity was measured with
the Dual Luciferase Reporter Assay System (Promega Corporation).
Similarly, pmirGLO-STAT3-WT or pmirGLO-STAT3-MUT were constructed
and co-transfected with miR-495 mimics or the corresponding control
into the cells. Following transfection, the cells were incubated
for 48 h and the luciferase activity was measured. The
Renilla luciferase was used as an internal control to
homogenize the detection of the reporter gene.
Western blot analysis
Total protein from tissues or cells was extracted
using pre-cooled RIPA lysis. Protein concentration was evaluated
using the BCA protein assay regents (cat. no. 23225, Pierce; Thermo
Fisher Scientific, Inc.). Equal amounts (50 µg/lane) of protein
aliquots were loaded on to 10% SDS-PAGE gels, then, proteins
transferred onto PVDF membranes. After blocking with 5% skimmed
milk for 2 h at room temperature, the membranes were then incubated
with primary antibodies at 4°C overnight. Next, the membranes were
incubated with secondary antibody for 1 h at room temperature. The
primary antibodies used in the present study were as follows: STAT3
(1:1,000; cat. no. 9139; Cell Signaling Technology, Inc.) and GAPDH
(1:3,000; cat. no. 8245; Abcam). The secondary antibody used was
HRP-conjugated goat anti-rabbit IgG (1:2,000; cat. no. 6728;
Abcam). The immunoblots were detected using LI-COR
Odyssey® CLX Two-colour infrared laser imaging system
(LI-COR Biosciences). ImageJ software V1.8.0 (National Institutes
of Health) was implemented to perform densitometric analysis.
RNA immunoprecipitation (RIP)
assay
RIP assays were performed using the Magna RIP™ kit
(EMD Millipore). Briefly, the cells were lysed using complete RIP
lysis buffer (4°C; 1,000 × g; 10 min) and incubated with containing
magnetic beads (2 µg) conjugated with human anti-Ago2 antibody
(Abcam; cat. no. ab186733) or anti-IgG antibody (EMD Millipore;
cat. no. MA5-27548) for overnight at 4°C. Samples were washed with
ethanol and sodium acetate and incubated with proteinase K at 55°C
for 30 min to isolate the RNA-protein complexes from beads (4°C;
1,000 × g; 15 min). Lastly, RT-qPCR assays were performed to
determine SNHG20 and miR-495 expression.
Tumor xenograft mouse models
Ten male nude mouse (age, 5–6 weeks; weight, 18–20
g) were housed in an isolated, clean, air-conditioned room at a
temperature of 25–26°C and a relative humidity of ~50%, light 12
h/day. The mice were randomly divided into two groups (n=5/group).
All rats were given free access to water and food. SW480 cells were
stably transfected with si-SNHG20 lentivirus or si-NC
(2×106 cells/mouse; n=5 for each group). A total of
5×106 cells suspended in 100 µl PBS were subcutaneously
inoculated into the right flanks of each nude mouse (the mice were
provided by the Experimental Animal Center of the Chinese Academy
of Sciences). Tumor size was measured once every 4 days following
the appearance of the tumors and calculated using the following
formula: V (mm3)=width2 (mm2) ×
length (mm)/2. Following 4 weeks of tumor growth, the mice mouse
was euthanized by cervical dislocation under anesthesia with 1%
pentobarbital (50 mg/kg) by intraperitoneal injection. The
following signs were used to verify death of the mice: Eyes had
turned pale, no heartbeat, and they did not respond to external
stimuli. The tumors were collected for weight assessment and for
RT-qPCR and western blot analyses. All the animal experiments were
carried out in accordance with the Ethics Committee of The First
Affiliated Hospital of University of South China.
In vivo metastasis study
Ten male nude mice were randomly divided into two
groups (n=5/group). SW480 cells that exhibited stable knockdown of
SNHG20 expression were transfected with firefly luciferase vector
(LV16; 100 nM; Guangzhou RiboBio Co., Ltd.). Nude mouse
(BALB/c-nu/nu; age, 5–6 weeks old; weight, 18–20 g) were
anesthetized by intraperitoneal injection of 1% pentobarbital (50
mg/kg) and cells (2×106 LV16-si-SNHG20 SW480 cells) were
inoculated into the tail vein. Lung metastases were monitored by
bioluminescent imaging for 6 consecutive weeks. All mice were
sacrificed at 6 weeks following injection. Before collection of the
lungs, the mice were euthanized by cervical dislocation under
anesthesia with 1% pentobarbital (50 mg/kg) by intraperitoneal
injection. The aforementioned indicators were used to verify death.
The lung tissue were fixed using 4% paraformaldehyde at 4°C for 24
h and sections were cut into 4 µm pieces. The lung metastatic
sites, the number of cells and the corresponding signal intensity
were recorded and analyzed by using a small animal live imaging
system (PerkinElmer, Inc.; Lumina X5).
Statistical analysis
All statistical analyses were carried out with
GraphPad Prism 8.0 software (GraphPad Software, Inc.) or SPSS 20.0
(IBM Corp.). The data are presented as the mean ± SD. A paired
Student's t-test was applied for the comparison of matched samples
(CRC and normal adjacent tissues). An unpaired t-test was applied
for the other comparisons between two groups. For the comparison of
multiple groups, one-way ANOVA analysis was performed followed by
Tukey's post hoc test. The statistical analysis between the
clinical parameters of the patients with CRC and the SNHG20
expression groups was analyzed using a chi-squared test or the
Fisher's exact test. Pearson correlation analysis was used to
estimate the relationship between the expression level of different
gene. All experiments were carried out at least three times.
P<0.05 was considered to indicate a statistically significant
difference.
Results
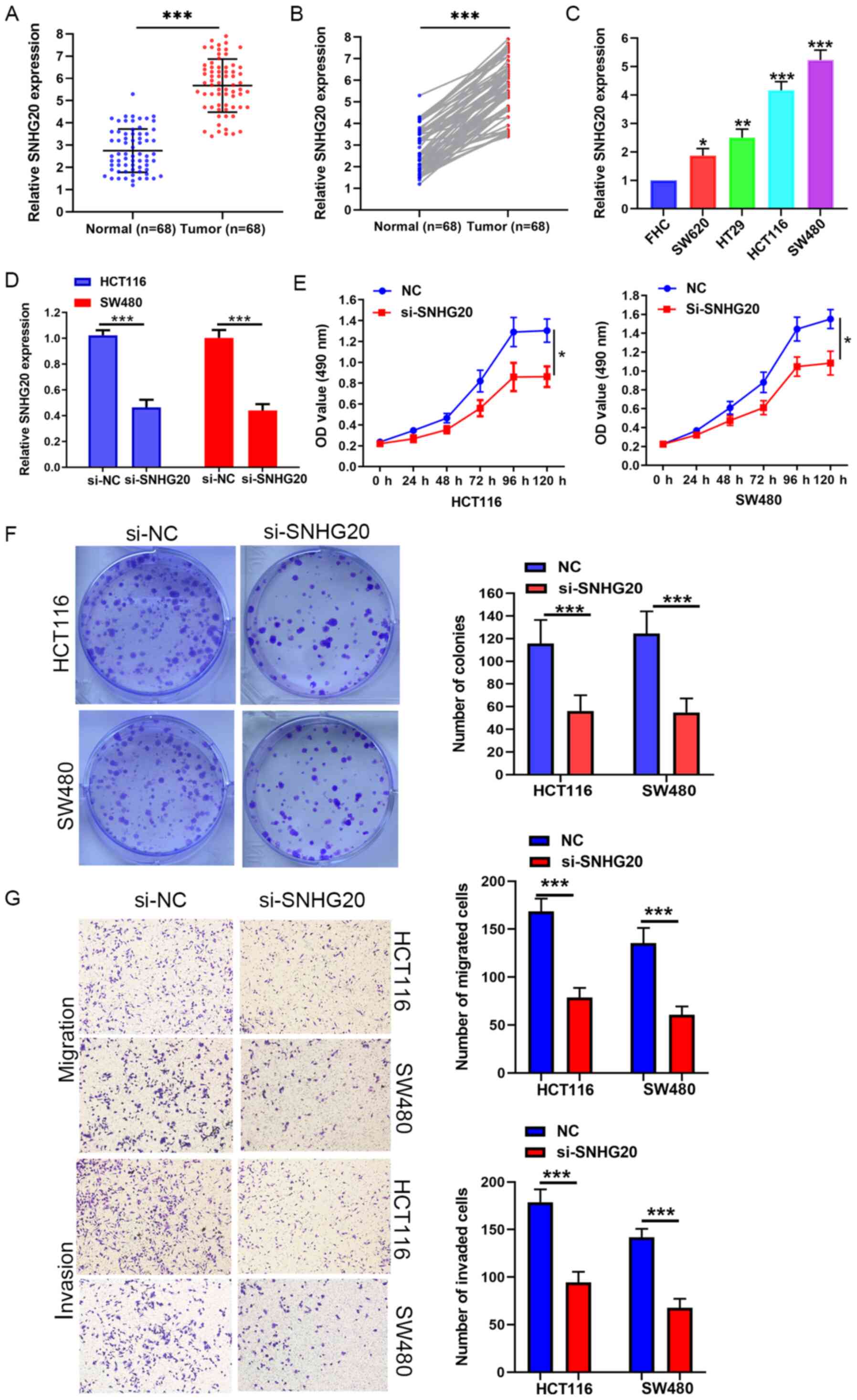
SNHG20 is highly expressed in CRC and
is associated with poor prognosis in patients with CRC
The expression levels of SNHG20 were measured in 68
pairs of CRC and adjacent normal tissues by RT-qPCR. The results
indicated that SNHG20 was highly expressed in CRC tissues (Fig. 1A and B). Similarly, SNHG20
expression was significantly increased in CRC cell lines (HCT116,
SW480, SW620 and HT-29) compared with that of the FHC cell line
(Fig. 1C). To further explore the
clinical significance of SNHG20 expression in CRC, 68 patients with
CRC were divided into a high (n=34) and a low expression group
(n=34), according to the median expression levels of SNHG20 in all
CRC samples. Notably, the clinicopathological data demonstrated
that high expression of SNHG20 was significantly associated with
tumor size (P=0.014), tumor invasion depth (P=0.019), positive
lymph node status (P=0.022), distant metastasis (P=0.017) and TNM
stage (P=0.038; Table II).
| Table II.Association between SNHG20 expression
and clinicopathological factors of CRC. |
Table II.
Association between SNHG20 expression
and clinicopathological factors of CRC.
|
|
| SNHG20
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Number of
cases | Low | High | P-value |
|---|
| Age, years |
|
|
| 0.627 |
|
<60 | 32 | 15 | 17 |
|
|
≥60 | 36 | 19 | 17 |
|
| Sex |
|
|
| 0.874 |
|
Male | 30 | 14 | 16 |
|
|
Female | 38 | 17 | 21 |
|
| Tumor size, cm |
|
|
| 0.014a |
|
<5 | 28 | 19 | 9 |
|
| ≥5 | 40 | 12 | 28 |
|
| Tumor invasion
depth |
|
|
| 0.019a |
|
T1+T2 | 22 | 15 | 7 |
|
|
T3+T4 | 46 | 19 | 27 |
|
| Lymph node
status |
|
|
| 0.022a |
|
Negative | 19 | 12 | 7 |
|
|
Positive | 49 | 16 | 33 |
|
| Distant
metastasis |
|
|
| 0.017a |
| No | 29 | 18 | 11 |
|
|
Yes | 39 | 14 | 25 |
|
| TNM stage |
|
|
| 0.038a |
|
I+II | 26 | 16 | 10 |
|
|
III+IV | 42 | 15 | 27 |
|
Knockdown of SNHG20 causes significant
suppression of cell viability, migration and invasion in vitro
Loss of function assays were performed to further
assess the biological function of SNHG20. SNHG20 expression was
significantly reduced in HCT116 and SW480 cells that were stably
transfected with si-SNHG20 (Fig.
1D). MTS and colony formation assays indicated that SNHG20
knockdown significantly suppressed the viability of HCT116 and
SW480 cells (Fig. 1E and F).
Moreover, Transwell assays revealed that SNHG20 knockdown resulted
in a significant reduction of the migratory and invasive activities
of HCT116 and SW480 cells (Fig.
1G).
SNHG20 interacts with miR-495 to
regulate CRC cell viability, invasion and migration
It has been reported that specific lncRNAs serve as
competing endogenous RNAs (ceRNAs) by sponging miRNAs (14). StarBase software was used to predict
the target miRNAs of SNHG20. miR-495 was characterized as a
potential target of SNHG20 due to its potential complementary sites
(Fig. 2A) for SNHG20 and its role
as a tumor suppressor in CRC (15).
The present study indicated that miR-495 expression levels were
significantly downregulated in CRC tissues and cell lines (HCT116,
SW480, SW620 and HT-29) (Fig. 2B and
C). The miR-495 mimic and inhibitor were transfected into
HCT116 and SW480 cell lines. The expression of miR-495 was
significantly increased in miR-495 mimic-transfected cells, whereas
miR-495 expression was significantly decreased in miR-495
inhibitor-transfected cells (Fig.
2D). The dual luciferase reporter assays revealed that miR-495
mimics significantly reduced the relative luciferase activity in
HCT116 and SW480 cells transfected with pmirGLO-SNHG20-WT (Fig. 2E). In addition, the expression of
miR-495 were significantly increased following SNHG20 knockdown
(Fig. 2F). Furthermore, Pearson's
correlation analysis revealed that miR-495 expression was inversely
associated with SNHG20 in CRC tissues (r=−0.53; P<0.001;
Fig. 2G). RIP assays indicated that
SNHG20 and miR-495 expression levels were significantly enriched in
Ago2-containing microribonucleoprotein complexes (Fig. 2H). Taken together, the data clearly
indicated that SNHG20 directly interacted with miR-495.
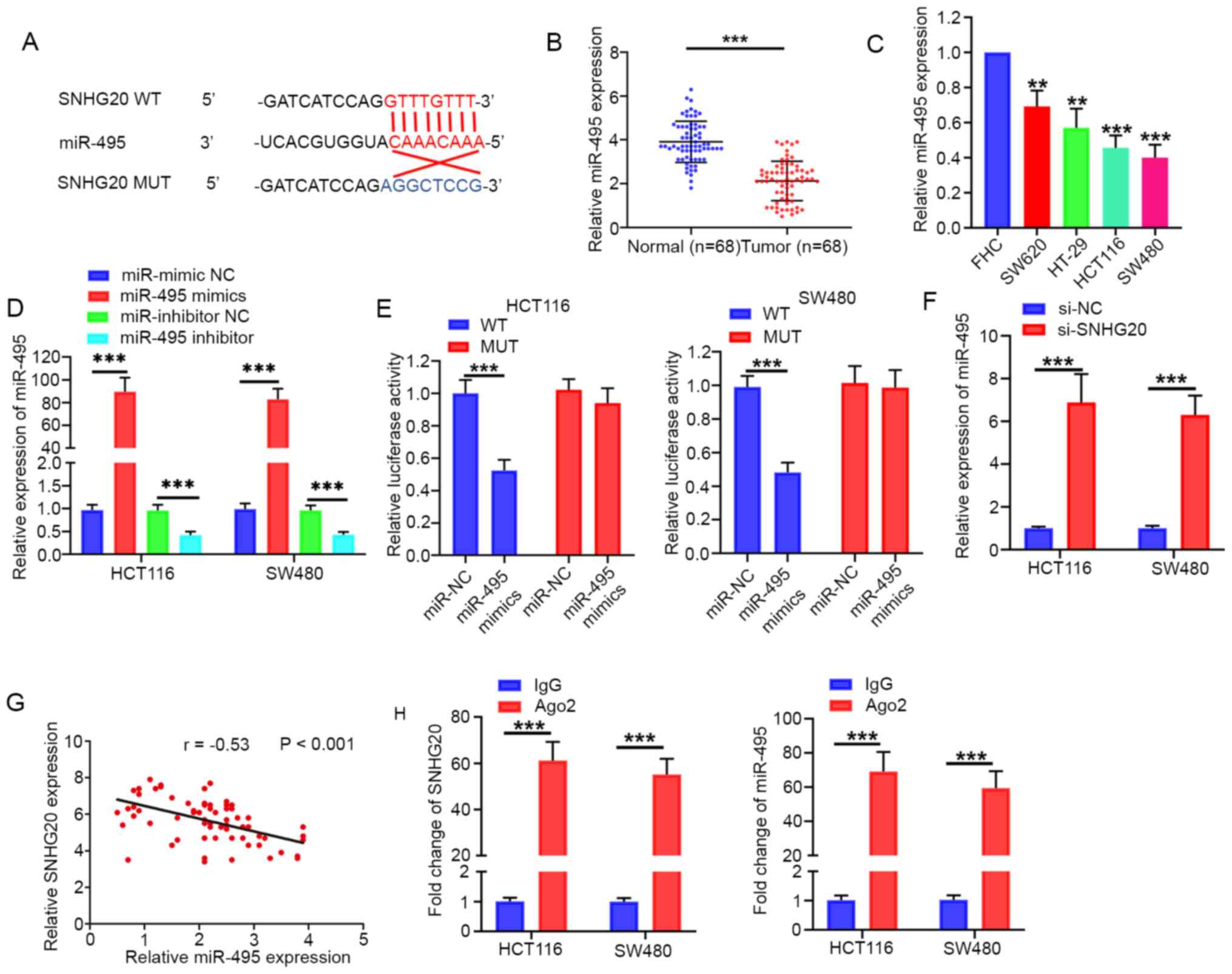 | Figure 2.SNHG20 acts as a sponge for miR-495.
(A) Predicted binding site of SNHG20 with miR-495, as determined by
bioinformatics analysis. (B and C) miR-495 expression levels were
significantly decreased in CRC tissues and CRC cell lines (SW620,
HT-29, HCT116, SW480). (D) Reverse transcription-quantitative PCR
analysis indicated that miR-495 expression was significantly
increased in miR-495 mimic-transfected cells, whereas miR-495
expression was significantly decreased in miR-495
inhibitor-transfected cells. (E) Luciferase reporter assay
indicated that SNHG20-WT activity was inhibited following
transfection of the cells with miR-495 mimics in HCT116 and SW480
cells. (F) SNHG20 knockdown increased the relative expression of
miR-495 in HCT116 and SW480 cells. (G) Pearson's correlation
analysis indicated that SNHG20 expression was negatively associated
with miR-495 in CRC tissues. (H) RNA immunoprecipitation assay
demonstrated enrichment of SNHG20 and miR-495 in Ago2-containing
beads. **P<0.01 and ***P<0.001 vs. FHC cells or as indicated.
SNHG20, small nucleolar RNA host gene 20; CRC, colorectal cancer;
miR, microRNA; WT, wild-type; MUT, mutant; si-, small interfering
RNA; NC, negative control. |
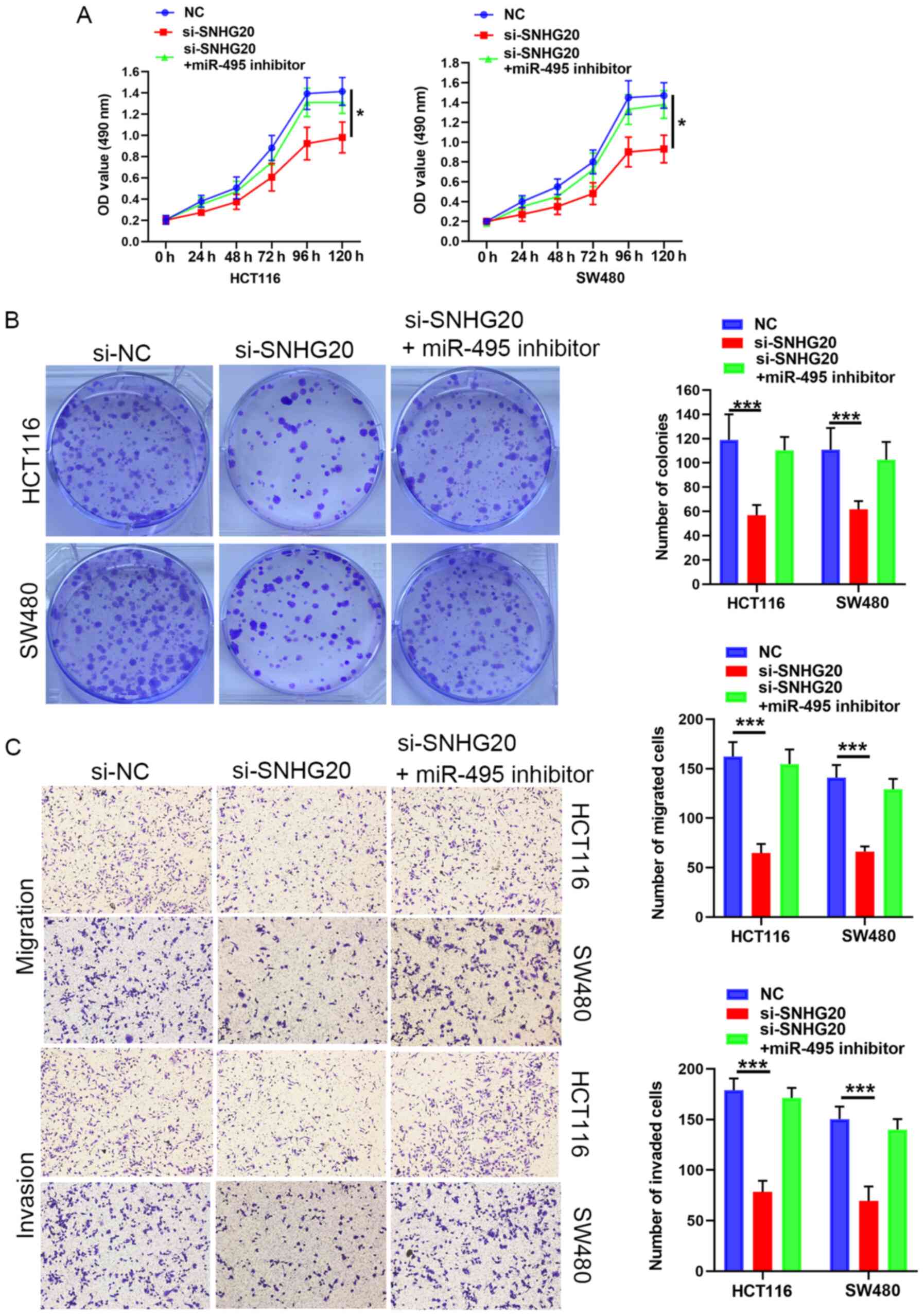
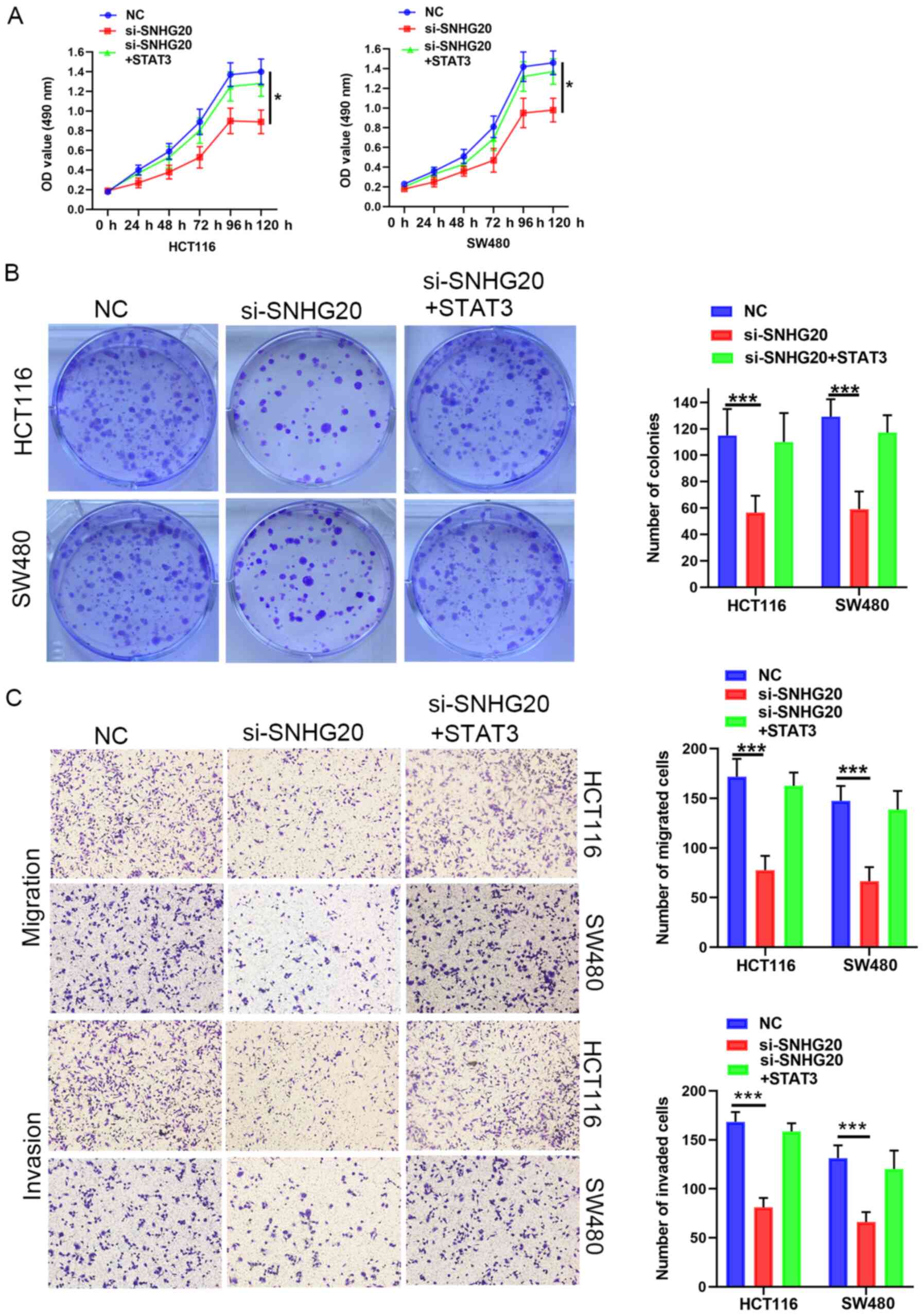
Subsequently, rescue experiments were performed by
transfecting miR-495 inhibitor into SNHG20-knockdown cells. The
results suggested that the miR-495 inhibitor reversed the
suppressive effects of SNHG20 knockdown on CRC cell viability
(Fig. 3A and B), migration and
invasion (Fig. 3C). In summary,
SNHG20 expression regulated the progression of CRC cells via
sponging miR-495.
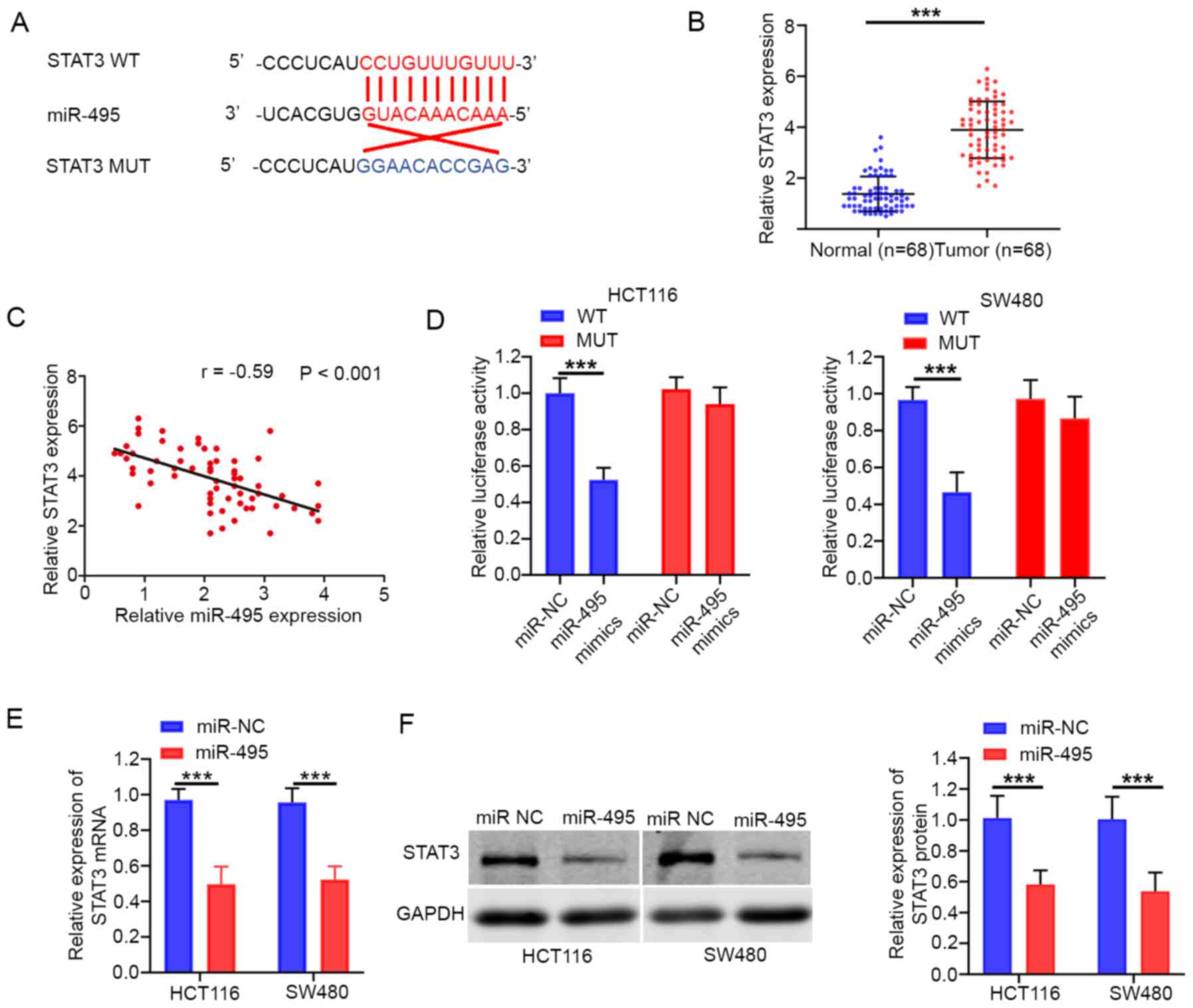
miR-495 targets STAT3 in CRC
The present study demonstrated that STAT3 was a
potential target of miR-495 (Fig.
4A). A Previous study reported that STAT3 serves as an oncogene
in various cancer types, including CRC (16). The data demonstrated that STAT3
expression levels were significantly upregulated in CRC tissues
(Fig. 4B). In addition, Pearson's
correlation analysis confirmed that miR-495 expression was
associated with STAT3 in CRC tissues (r=−0.59; P<0.001; Fig. 4C). The luciferase reporter assay
revealed that miR-495 mimics significantly reduced the luciferase
activity of STAT3-WT, whereas this effect was not noted for
STAT3-MUT (Fig. 4D). In addition,
overexpression of miR-495 resulted in a significant decrease in the
mRNA and protein expression of STAT3 in both HCT116 and SW480 cells
(Fig. 4E and F).
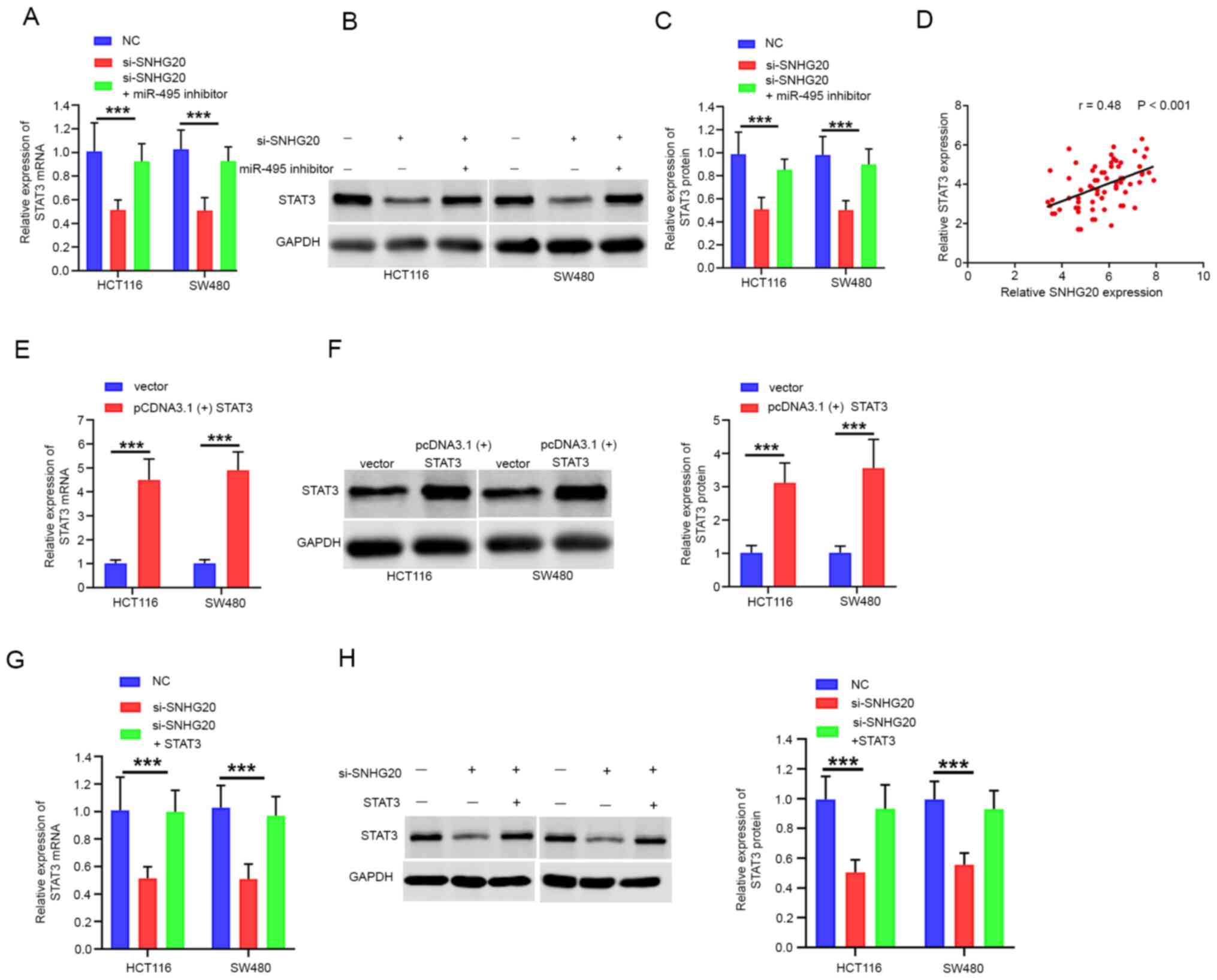
SNHG20 positively regulates STAT3
expression by competitively binding to miR-495
Accumulating evidence has revealed that lncRNAs
regulate tumor progression by competing for target miRNAs and
modulating mRNA expression (17).
Therefore, the present study aimed to investigate whether SNHG20
expression contributed to CRC progression by modulating STAT3
expression. The data indicated that STAT3 mRNA and protein levels
were significantly reduced following SNHG20 knockdown. This effect
could be partially reversed by the addition of miR-495 inhibitor to
the cells (Fig. 5A-C). Moreover,
Pearson's correlation analysis confirmed that STAT3 expression was
positively correlated with SNHG20 expression in CRC tissues
(r=0.48; P<0.001; Fig. 5D). To
further investigate the role of STAT3 and SNHG20 in CRC
progression, pcDNA3.1(+) STAT3 was transfected into cells. The mRNA
and protein levels of STAT3 were increased significantly following
transfection of the cells with pcDNA3.1(+) STAT3 (Fig. 5E and F). In addition, the mRNA and
protein levels of STAT3 were significantly reduced following SNHG20
knockdown, whereas co-transfection of pcDNA3.1(+) STAT3 and
si-SNHG20 reversed this inhibitory effect (Fig. 5G and H). Functional assays indicated
that STAT3 overexpression could rescue the inhibitory effect of
SNHG20 knockdown on cell viability (Fig. 6A and B), migration and invasion
(Fig. 6C). In summary, these data
demonstrated that SNHG20 facilitated CRC progression by modulating
the miR-495/STAT3 axis.
SNHG20 knockdown inhibits tumor growth
and metastasis formation in vivo
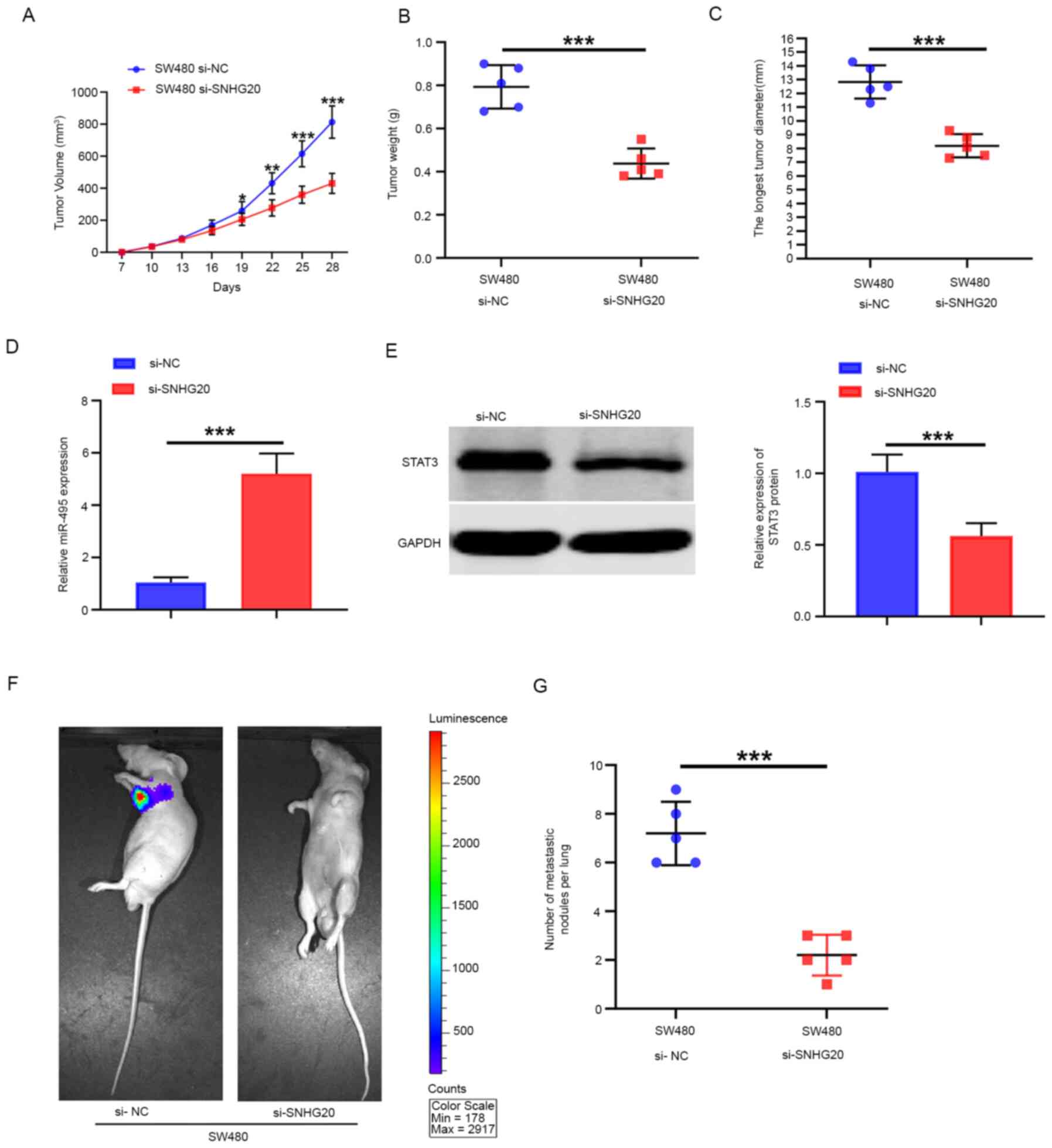
The tumor volume was significantly smaller in the
SNHG20-knockdown group than that noted in the NC group (Fig. 7A) In addition, the average weight of
the tumors in the SNHG20-knockdown group was significantly lower
than that noted in the NC group (Fig.
7B). The tumor diameters of the tumors in the SNHG20-knockdown
group were significantly smaller than that in the NC group
(Fig. 7C). The expression levels of
miR-495 in tumor tissues from the SNHG20-knockdown group were
higher than those of the NC group as determined by RT-qPCR
(Fig. 7D), which was consistent
with the in vitro data. However, the expression levels of
STAT3 in the SNHG20-knockdown group were significantly lower than
those of the NC group (Fig. 7E).
The results derived from the in vivo fluorescence imaging
indicated that the lowest number of metastatic sites was present in
the SNHG20-knockdown group, whereas the highest number was found in
the NC group (Fig. 7F). In
addition, SNHG20 knockdown significantly reduced the number of
metastatic pulmonary nodules compared with that in the NC group
(Fig. 7G). Taken together, the
results indicated that SNHG20 knockdown inhibited CRC growth and
metastasis in vivo.
Discussion
A previous study reported that SNHG20 expression was
significantly upregulated in CRC tissues and that this lncRNA was
involved in the development and progression of CRC (12). However, the aforementioned study was
primarily focused on the association between SNHG20 expression and
the clinical features of the subjects. The specific molecular
mechanisms of SNHG20 in the progression of CRC are still unclear.
In the present study, the expression of SNHG20 was investigated and
the data demonstrated that it was significantly increased in CRC.
Chi-square and Fisher's exact tests indicated that SNHG20
expression was associated with tumor size, tumor invasion depth,
TNM stage, lymph node status and distant metastasis. Functional
experiments demonstrated that SNHG20 silencing suppressed
viability, migration and invasion of CRC cells in vitro, and
it inhibited CRC growth and metastasis in vivo.
Previous studies have revealed that SNHG20 plays an
important role in the development of a multitude of cancers. In
gastric cancer, Cui et al (18) demonstrated that high SNHG20
expression was positively associated with poor clinical outcomes.
This previous study further demonstrated that SNHG20 silencing
inhibited proliferation and invasion of gastric cancer cells. In
osteosarcoma, Wang et al (19) reported that SNHG20 silencing notably
repressed proliferation and invasion, while enhancing the apoptotic
fraction of osteosarcoma cells. In glioma, Li et al
(20) demonstrated that SNHG20
expression was upregulated in glioma tissues and cells.
Furthermore, they also revealed that SNHG20 silencing led to
attenuated vasculogenic mimicry formation in glioma cells (20). The present study reported that
SNHG20 expression was significantly upregulated in CRC, which was
consistent with the results reported previously.
Mounting evidence has demonstrated that lncRNA acts
as a sponge for miRNAs, which further modulate gene expression
levels and subsequently participate in several biological
processes, including cell proliferation and metastasis (7,14).
However, the target miRNAs of SNHG20 have not been previously
identified in CRC. Based on the StarBase software, miR-495 was
predicted as a potential target of SNHG20. miR-495 is involved in
the development and progression of several tumors. For example, Li
et al (21) revealed that
miR-495 acted as a tumor suppressor by directly targeting protein
tyrosine phosphatase type IVA 3 in human gastric cancer. miR-495
was also reported as an antitumor miRNA in prostate cancer
(22), NSCLC (23) and endometrial cancer (24). However, in breast cancer,
overexpression of miR-495 was demonstrated to facilitate cell
invasion (25). Moreover, Bai et
al (26) reported that miR-495
overexpression markedly inhibited invasion and the
epithelial-mesenchymal transition of CRC cells. A study by Yan
et al (15) indicated that
miR-495 expression levels were considerably reduced in CRC tissues
and cell lines compared with those of adjacent normal tissues and
cell lines. Overexpression of miR-495 significantly inhibited the
proliferation, colony formation, migration and invasion of CRC
cells (15). He et al
(27) demonstrated that miR-495
levels were significantly decreased in CRC cells, and that they
were associated with significant suppression of CRC progression. In
the present study, the results of the luciferase reporter and RIP
assays demonstrated that SNHG20 interacted with miR-495 in CRC
cells. In addition, Pearson's correlation analysis further
confirmed that SNHG20 expression was negatively associated with
miR-495 expression in CRC tissues. Moreover, SNHG20 knockdown
significantly increased miR-495 expression in CRC cells. All these
data confirmed that SNHG20 contributed to CRC progression via
sponging miR-495.
STAT3 is a transcription factor that acts as an
oncogene and its expression is upregulated in various human cancers
(28). A previous study conducted
by Morikawa et al (29) used
724 CRC samples and demonstrated that STAT3 activation was
associated with adverse clinical outcomes in CRC. It was reported
that inactivation of STAT3 in CRC cells markedly inhibited tumor
growth both in vitro and in vivo (30). In the present study, STAT3 was
confirmed as a downstream target of miR-495 in CRC via luciferase
reporter assays. In addition, silencing of SNHG20 expression
reduced STAT3 expression, whereas co-transfection of the cells with
the miR-495 inhibitor reversed the inhibitory effects of SNHG20
silencing on STAT3 expression. Moreover, functional experiments
illustrated that co-transfection of the cells with pcDNA3.1-STAT3
rescued the effects of SNHG20 knockdown on viability, migration and
invasion of CRC cells.
In summary, the results of the present study
demonstrated that SNHG20 promoted viability, migration and invasion
of CRC cells. SNHG20 functioned as a ceRNA for miR-495 and
upregulated STAT3 expression, which in turn facilitated CRC
progression. The SNHG20/miR-495/STAT3 axis may aid the
understanding of the molecular mechanism of CRC progression and
provide a novel therapeutic target for CRC treatment.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW and ZL were responsible for conception of the
study and the drafting of the manuscript. JF and LY helped to
design the study and performed the statistical analysis. ZL revised
the manuscript. All authors read and approved the final version of
the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Committee of The First Affiliated Hospital of University of South
China (Hengyang, China). Signed informed consent had been provided
by all patients. All of the animals experiments were performed
under protocols approved by the Institutional Animal Care and Use
Committee of The First Affiliated Hospital of the University of
South China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dienstmann R, Vermeulen L, Guinney J,
Kopetz S, Tejpar S and Tabernero J: Consensus molecular subtypes
and the evolution of precision medicine in colorectal cancer. Nat
Rev Cancer. 17:79–92. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sridharan M, Hubbard JM and Grothey A:
Colorectal cancer: How emerging molecular understanding affects
treatment decisions. Oncology (Williston Park). 28:110–118.
2014.PubMed/NCBI
|
|
4
|
Fatica A and Bozzoni I: Long non-coding
RNAs: New players in cell differentiation and development. Nat Rev
Genet. 15:7–21. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shi X, Sun M, Liu H, Yao Y and Song Y:
Long non-coding RNAs: A new frontier in the study of human
diseases. Cancer Lett. 339:159–166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hua F, Liu S, Zhu L, Ma N, Jiang S and
Yang J: Highly expressed long non-coding RNA NNT-AS1 promotes cell
proliferation and invasion through wnt/β-catenin signaling pathway
in cervical cancer. Biomed Pharmacother. 92:1128–1134. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ge X, Li GY, Jiang L, Jia L, Zhang Z, Li
X, Wang R, Zhou M, Zhou Y, Zeng Z, et al: Long noncoding RNA CAR10
promotes lung adenocarcinoma metastasis via miR-203/30/SNAI axis.
Oncogene. 38:3061–3076. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang D, Cao C, Liu L and Wu D:
Up-Regulation of lncRNA SNHG20 predicts poor prognosis in
hepatocellular carcinoma. J Cancer. 7:608–617. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen Z, Chen X, Chen P, Yu S, Nie F, Lu B,
Zhang T, Zhou Y, Chen Q, Wei C, et al: Long non-coding RNA SNHG20
promotes non-small cell lung cancer cell proliferation and
migration by epigenetically silencing of P21 expression. Cell Death
Dis. 8:e30922017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu J, Zhao W, Wang Z, Xiang X, Zhang S and
Liu L: Long non-coding RNA SNHG20 promotes the tumorigenesis of
oral squamous cell carcinoma via targeting miR-197/LIN28 axis. J
Cell Mol Med. 23:680–688. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu X, Xiao Y, Zhou Y, Zhou Z and Yan W:
lncRNA SNHG20 promotes prostate cancer migration and invasion via
targeting the miR-6516-5p/SCGB2A1 axis. Am J Transl Res.
11:5162–5169. 2019.PubMed/NCBI
|
|
12
|
Li C, Zhou L, He J, Fang XQ, Zhu SW and
Xiong MM: Increased long noncoding RNA SNHG20 predicts poor
prognosis in colorectal cancer. BMC Cancer. 16:6552016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolf PP: A ceRNA hypothesis: The rosetta stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yan LK, Yao JF and Qiu J: miRNA-495
suppresses proliferation and migration of colorectal cancer cells
by targeting FAM83D. Biomed Pharmacother. 96:974–981. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou C, Tong Y, Wawrowsky K and Melmed S:
PTTG acts as a STAT3 target gene for colorectal cancer cell growth
and motility. Oncogene. 33:851–861. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cai Y, Sheng Z, Chen Y and Wang J: LncRNA
HMMR-AS1 promotes proliferation and metastasis of lung
adenocarcinoma by regulating miR-138/sirt6 axis. Aging (Albany NY).
11:3041–3054. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cui N, Liu J, Xia H and Xu D: LncRNA
SNHG20 contributes to cell proliferation and invasion by
upregulating ZFX expression sponging miR-495-3p in gastric cancer.
J Cell Biochem. 120:3114–3123. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang W, Luo P, Guo W, Shi Y, Xu D, Zheng H
and Jia L: LncRNA SNHG20 knockdown suppresses the osteosarcoma
tumorigenesis through the mitochondrial apoptosis pathway by
miR-139/RUNX2 axis. Biochem Biophys Res Commun. 503:1927–1933.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li X, Xue Y, Liu X, Zheng J, Shen S, Yang
C, Chen J, Li Z, Liu L, Ma J, Ma T and Liu Y: ZRANB2/SNHG20/FOXK1
axis regulates vasculogenic mimicry formation in glioma. J Exp Clin
Cancer Res. 38:682019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Z, Cao Y, Jie Z, Liu Y, Li Y, Li Y, Li
J, Zhu G, Liu Z, Tu Y, et al: miR-495 and miR-551a inhibit the
migration and invasion of human gastric cancer cells by directly
interacting with PRL-3. Cancer Lett. 323:41–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li JZ, Wang ZL, Xu WH, Li Q, Gao L and
Wang ZM: MicroRNA-495 regulates migration and invasion in prostate
cancer cells via targeting akt and mTOR signaling. Cancer Invest.
34:181–188. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chu H, Chen X, Wang H, Du Y, Wang Y, Zang
W, Li P, Li J, Chang J, Zhao G and Zhang G: miR-495 regulates
proliferation and migration in NSCLC by targeting MTA3. Tumour
Biol. 35:3487–3494. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu YY, Tian J, Hao Q and Yin LR:
MicroRNA-495 downregulates FOXC1 expression to suppress cell growth
and migration in endometrial cancer. Tumour Biol. 37:239–251. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hwang-Verslues WW, Chang PH, Wei PC, Yang
CY, Huang CK, Kuo WH, Shew JY, Chang KJ, Lee EY and Lee WH: miR-495
is upregulated by E12/E47 in breast cancer stem cells, and promotes
oncogenesis and hypoxia resistance via downregulation of E-cadherin
and REDD1. Oncogene. 30:2463–2474. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bai Z, Wang J, Wang T, Li Y, Zhao X, Wu G,
Yang Y, Deng W and Zhang Z: The miR-495/Annexin A3/P53 axis
inhibits the invasion and EMT of colorectal cancer cells. Cell
Physiol Biochem. 44:1882–1895. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He Z, Dang J, Song A, Cui X, Ma Z and
Zhang Z: NEAT1 promotes colon cancer progression through sponging
miR-495-3p and activating CDK6 in vitro and in vivo. J Cell
Physiol. 234:19582–19591. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yuan J, Zhang F and Niu R: Multiple
regulation pathways and pivotal biological functions of STAT3 in
cancer. Sci Rep. 5:176632015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Morikawa T, Baba Y, Yamauchi M, Kuchiba A,
Nosho K, Shima K, Tanaka N, Huttenhower C, Frank DA, Fuchs CS and
Ogino S: STAT3 expression, molecular features, inflammation
patterns and prognosis in a database of 724 colorectal cancers.
Clin Cancer Res. 17:1452–1462. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang P, Zhao Y, Zhu X, Sedwick D, Zhang X
and Wang Z: Cross-Talk between phospho-STAT3 and PLCγ1 plays a
critical role in colorectal tumorigenesis. Mol Cancer Res.
9:1418–1428. 2011. View Article : Google Scholar : PubMed/NCBI
|