Introduction
Sevoflurane is an inhaled anesthetic introduced into
clinical practice >20 years ago (1). It is a sweet-smelling, fast-onset and
recovery agent, with a low blood-gas partition coefficient and
limited cardiorespiratory depression properties (2). The use of sevoflurane for general
anesthesia in the pediatric population has become common (3). However, an increasing number of
studies on rodents and nonhuman primates suggested that sevoflurane
can cause neuronal apoptosis in the developing brain and result in
learning and memory deficits later in adulthood (4–7).
Sevoflurane has also been demonstrated to inhibit the proliferation
of neural progenitor cells, reduce the self-renewal capacity of
neural stem cells and induce neuroinflammation through microglial
cells in mice (8–11). These findings have raised concern
about the detrimental effects of sevoflurane on brain development
and neurocognitive function in children.
Dexmedetomidine is an α2 adrenoceptor agonist that
has been used as an anesthetic agent and sedative for several years
(12,13). In clinical practice, dexmedetomidine
is used to prevent sevoflurane-related agitation in children
(14,15). Previous studies have suggested that
dexmedetomidine could suppress sevoflurane-induced neuronal
apoptosis and neurocognitive impairment in neonatal rats.
Importantly, our preliminary study also indicated that
dexmedetomidine could attenuate sevoflurane-induced learning and
memory impairment in mice. However, the mechanism underlying the
neuroprotective effect of dexmedetomidine remains poorly
understood. Dexmedetomidine is an α2 adrenergic receptor agonist,
and α2 adrenoceptors are known to act as trophic factors in the
central nervous system (16).
Moreover, adrenoceptors activate endogenous norepinephrine, which
promotes cell survival, notably through the Ras-Raf-ERK pathway
(17,18).
Several studies have suggested that dexmedetomidine
has neuroprotective effects against ischemic cerebral injury
through activation of α2 adrenergic receptors and binding to
imidazoline-1 and −2 receptors (19,20).
It was suggested that neuroinflammation and oxidative stress may
cause synapse dysfunction, which results in cognitive dysfunction
(21–25). The aim of the present study was to
investigate the effect of dexmedetomidine on sevoflurane-induced
neuroinflammation, oxidative stress and neuroapoptosis. The role of
α2 adrenoceptors in the neuroprotective effect of dexmedetomidine
was also examined.
Materials and methods
Animals
A total of 60 of postnatal day 6 (P6) C57BL/6 male
mice (weight, ~1.7 g) were purchased from Changzhou Cavens
Laboratory Animal Co., Ltd. Mice were housed with their mothers for
4 weeks for 12:12 h of light and dark cycles at a temperature of
24±2°C and 60±10% humidity prior to sevoflurane exposure. All
animals had free access to food and water.
Experimental procedures
All animal procedures were approved by the Animal
Experimental Ethics Committee of The Huai'an Maternity and Child
Clinical College of Xuzhou Medical University and were performed in
strict accordance with the guidelines of University Laboratory
Animal Management.
In a first experiment, animals were divided into
four groups: NS + Air (control group), NS + Sev, Dex20 + Sev and
Dex20 + Sev + Yoh (n=6/group). P6 mice received an intraperitoneal
injection of 20 µg/kg dexmedetomidine (Jiangsu Hengrui Medicine
Co., Ltd.) or normal saline 2 h prior to sevoflurane exposure
(26,27). The mice were then exposed to either
6 h of 3% sevoflurane in 25% oxygen or to air in a
temperature-controlled chamber, and injection of yohimbine (1
mg/kg) 15 min before sevoflurane exposure for Dex20 + Sev + Yoh
group. A Morris water maze (MWM) test was conducted to study
hippocampal-dependent learning and memory ability from P35 till
P41.
In a separate experiment, animals were allocated
into six groups: NS + Air, NS + Sev, Dex10 + Sev, Dex20 + Sev,
Dex20 + Sev + Yoh and Dex + Air. Normal saline or 5, 10 or 20 µg/kg
dexmedetomidine with or without injection of a2-adrenoceptor
antagonist yohimbine (1 mg/kg; Absin) were administered by
intraperitoneal injection 2 h before exposure, and the mice were
then exposed to either 6 h of 3% sevoflurane in 25% oxygen or to
air. At the end of sevoflurane exposure, all mice were sacrificed
by removal of the brain under anesthesia by intraperitoneal
injection of 100 mg/kg sodium pentobarbital. The hippocampus was
then dissected out on ice for subsequent experiments.
MWM test
The MWM test was conducted in a circular tank filled
with 20°C water opacified with titanium dioxide (diameter, 1.8 m;
depth, 60 cm). In the center of the tank, a 11×11 cm platform was
located 1.0 cm from the board of the tank The mice were tested on
the MWM four times a day, from P35 to P41 (7 days in total). Mice
were randomly placed in the pool. If a mouse found the platform, it
was allowed to stay on it for 15 sec. If the mouse was not able to
find the platform within 90 sec, it was guided to the platform and
allowed to stay on it for 15 sec. The swimming process was recorded
by a video tracking system, and the data were captured using
motion-detection software (Biobserve FST Analysis). The platform
was removed from the pool after the reference training, and the
mice were placed in the opposite quadrant. Both the number of
crossings completed within 60 sec and the crossing time were
recorded. At the end of the test, each mouse was wiped dry to
prevent hypothermia.
Immunohistochemistry
The hippocampus tissue was cut into 5-µm sections,
which were fixed in 4% paraformaldehyde overnight at 4°C and
embedded in paraffin. Then, tissue was de-paraffinized and
rehydrated. After 24 h, the sections were dried at 37°C, then
incubated with 0.3% hydrogen peroxidase in methanol for 30 min at
room temperature and washed in PBS blocked with 1% bovine serum
albumin (BSA, MP Biomedicals, LLC) in PBS at room temperature for
60 min. The sections were then incubated with goat-anti-cleaved
caspase-3 primary antibody (1:200; cat. no. sc-166589; Santa Cruz
Biotechnology, Inc.) at 4°C overnight, then with a
Vectastain® Avidin-Biotin Complex staining kit (Vector
Laboratories, Inc. PK-6100) for 40 min at room temperature in the
dark. Tissue sections were then stained with diaminobenzidine
(Vector Laboratories, Inc.), then placed in a gradient of ethanol
solutions (70–100%) and finally covered with a coverslip using
neutral resin. A light microscope (magnification, ×200) was used to
observe sections, NIS-Elements BR image processing and analysis
software (cat. no. E100; Nikon Corporation) was used to quantify
the 3 fields of cleaved caspase-3-positive cells in the very 3
hippocampal CA1.
ELISA
The levels of tumor necrosis factor-α (TNF-α),
interleukin (IL)-6 and IL)-1β in the hippocampus were determined
using ELISA kits purchased from R&D Systems (cat. no. MTA00B,
M6000B and MLB00C for TNF-α, IL-6 and IL-1β, respectively),
according to the manufacturer's instructions. The hippocampal
tissue was homogenized using ice-cold lysis buffer (Promega
Corporation) and an electric homogenizer and centrifuged at 7,155 ×
g for 5 min at 4°C and the protein concentration was quantified
using a Pierce™ BCA Protein Assay kit (Thermo Fisher Scientific,
Inc.).
Superoxide dismutase (SOD) activity
measurement
SOD is an enzyme that catalyzes the dismutation of
superoxide radicals into either oxygen or hydrogen peroxide
(26). SOD activity was analyzed as
described according to the SOD kit procedure (cat. no. bc0170;
Beijing Solarbio Science & Technology Co., Ltd.), Samples were
prepared, and analyzed based on the procedure measured at 560
nm.
Measurement of malondialdehyde (MDA)
levels
MDA is a marker of oxidative stress-mediated lipid
peroxidation (26). MDA levels were
measured using the thiobarbituric acid reaction method from Beijing
Solarbio Science & Technology Co., Ltd. MDA kit (cat. no.
BC0025). A total of ~0.1 g tissue was weighed and 1 ml extract was
added for ice bath homogenate. Following centrifugation at 8,000 ×
g at 4°C for 10 min, the samples were taken re-suspended and placed
on ice to be measured. Microplate reader was used, absorbance was
read at a wavelength of 450, 532 and 600 nm. The MDA levels (in
nmol/mg protein) were calculated.
Flow cytometry
The frequency of apoptotic cells in the brain was
assessed by flow cytometry. Briefly, hippocampus were harvested on
ice immediately after sacrifice. Hippocampus cells were isolated
into a single-cell suspension using 10% trypsin at 37°C for 15 min.
An annexin V-FITC and propidium iodide apoptosis detection kit (BD
Biosciences, cat. no. 556547 was used to stain apoptotic cells. A
total of 3×104 single cells per sample were analyzed by
flow cytometry (BD Accuri, C6) and FlowJo 8.6 software (both Becton
Dickinson & Company).
Western blot analysis
Western blotting was used to examine phosphorylated
(p)-cAMP response element-binding protein (CREB) levels after
dexmedetomidine treatment. Hippocampus were centrifuged at 12,000 ×
g at 4°C for 5 min immediately after mice brains were dissected and
digested by RIPA lysis buffer (Sangon Biotech) with 1% PMSF on ice
for 15 min. BCA assay was used to determine protein concentration.
Then, the lysate was heated at 95°C for 10 min and 30 µg was loaded
on a 10% gel. All protein samples were separated by SDS-PAGE, then
transferred to a nitrocellulose membrane with 5% BSA TBST (0.1%
Tween-20) for 60 min. After incubation with rabbit anti-mice
primary antibodies (1:2,000; p-CREB, cat. no. ab32096; CREB, cat.
no. ab32515; and β-actin, cat. no. ab6276; all Abcam) at 4°C
overnight and horseradish peroxidase-conjugated secondary antibody
[Goat Anti-Rabbit and Anti-Mouse (1:2,000; cat. nos. ab205718 and
ab205719, respectively; both Abcam)] for 1 h at room temperature,
all membranes were exposed in a dark room with ECL reagent and
imaged using a Tanon 1600/1600R Gel Imaging System (UVP, LLC).
Statistical analysis
All data are presented as the mean ± SD. All
experiments were repeated at least twice. Student's t-test was
performed to compare the difference between two groups. Multigroup
comparisons were performed using one-way ANOVA followed by Tukey's
post hoc test. GraphPad Prism 5 (GraphPad Software, Inc.) was used
to conduct the analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
Dexmedetomidine reverses
sevoflurane-induced learning and memory impairment via α2
adrenoceptors
P6 mice were treated with 3% sevoflurane for 6 h,
and neurocognitive function was tested using a MWM from P35-P41.
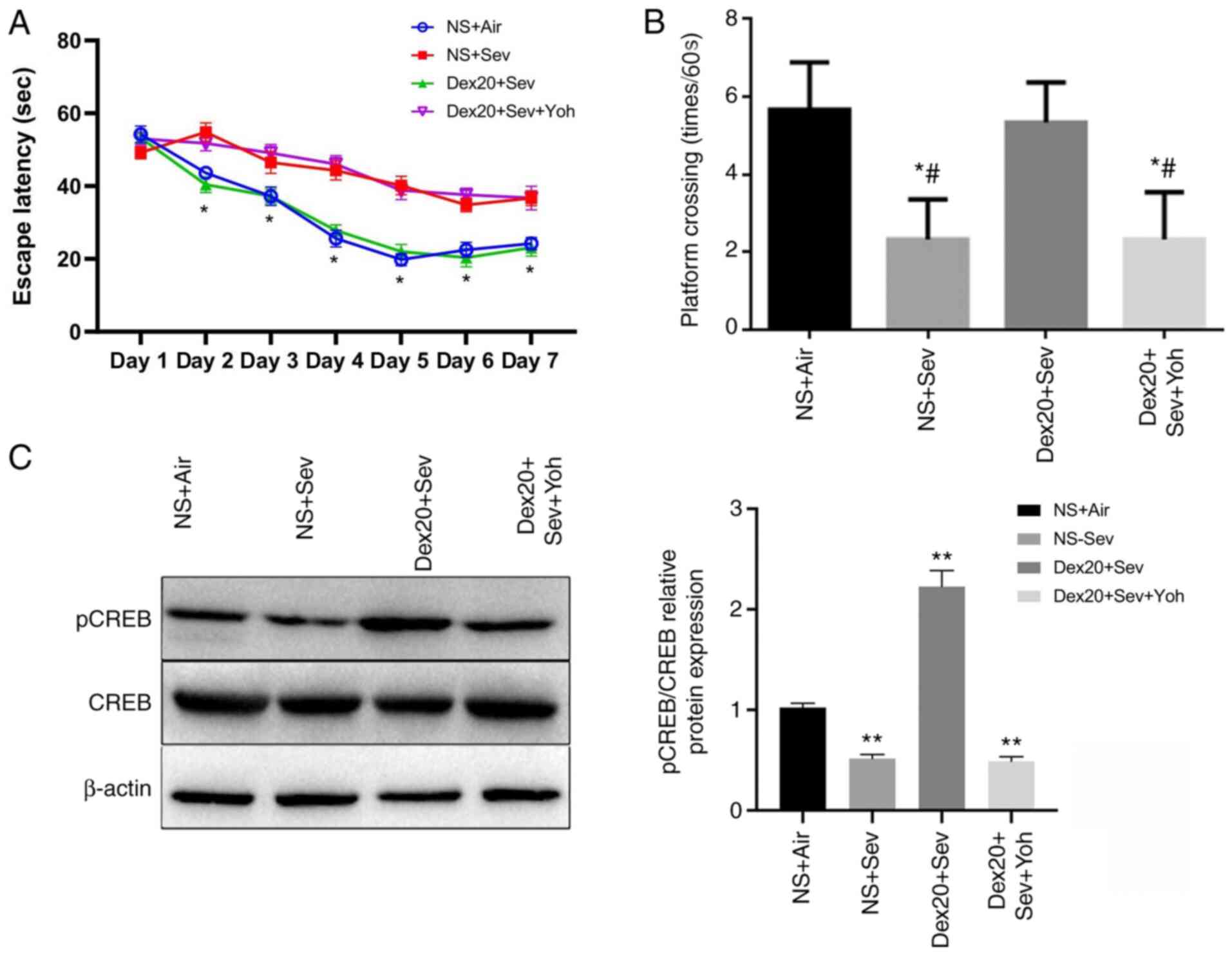
The mice exposed to sevoflurane displayed significantly increased
escape latency times from days 2–7 compared with control mice
exposed to air (Fig. 1A). Moreover,
sevoflurane-treated mice also completed significantly fewer
crossings compared with the control group (Fig. 1B). These observations suggested that
sevoflurane exposure in young mice could induce cognitive
impairment after three weeks. However, mice treated with
dexmedetomidine 2 h prior to sevoflurane exposure displayed
significantly reduced cognitive impairment, as indicated by shorter
escape latency times and increased numbers of crossings, compared
with mice receiving sevoflurane alone. There were no significant
differences between the dexmedetomidine treatment group and the
control group. By contrast, the α2 adrenoceptor antagonist
yohimbine significantly inhibited the neuroprotective effect of
dexmedetomidine. No significant differences in escape latency and
number of platform crossings were observed between the
yohimbine-treated group and mice exposed only to sevoflurane,
indicating that the neuroprotective effect of dexmedetomidine may
be mediated by α2 adrenoceptors.
pCREB is a molecular marker for memory processing in
space learning in the hippocampus (28). Compared with the control group,
Western blot analysis demonstrated that pCREB levels were
significantly decreased in the brain following sevoflurane
exposure, but restored by dexmedetomidine treatment. However, this
effect was inhibited by yohimbine pretreatment (Fig. 1C). These results indicated that
dexmedetomidine could reverse learning and memory impairment caused
by sevoflurane.
Dexmedetomidine attenuates
sevoflurane-induced neuronal apoptosis through α2 adrenoceptors in
6-day-old mice
The CA1 pyramidal cell layer has neurophysiological
signature characteristics, which serve a role in the hippocampal
memory circuit as an outstanding output node (27). A 6-h exposure to 3% sevoflurane
resulted in a significant increase in caspase-3-positive cells in
the CA1 layer of the hippocampus, compared with air-exposed control
mice. Pretreatment with dexmedetomidine significantly decreased the
sevoflurane-induced increase in caspase-3-positive cells (Fig. 2A). However, yohimbine attenuated the
effect of dexmedetomidine treatment on the number of
caspase-3-postive cells, suggesting a role for α2 adrenoceptors in
dexmedetomidine-mediated neuroprotection. Moreover, dexmedetomidine
significantly reduced sevoflurane-induced apoptosis in the brain,
and this effect was partially inhibited by yohimbine (Fig. 2B).
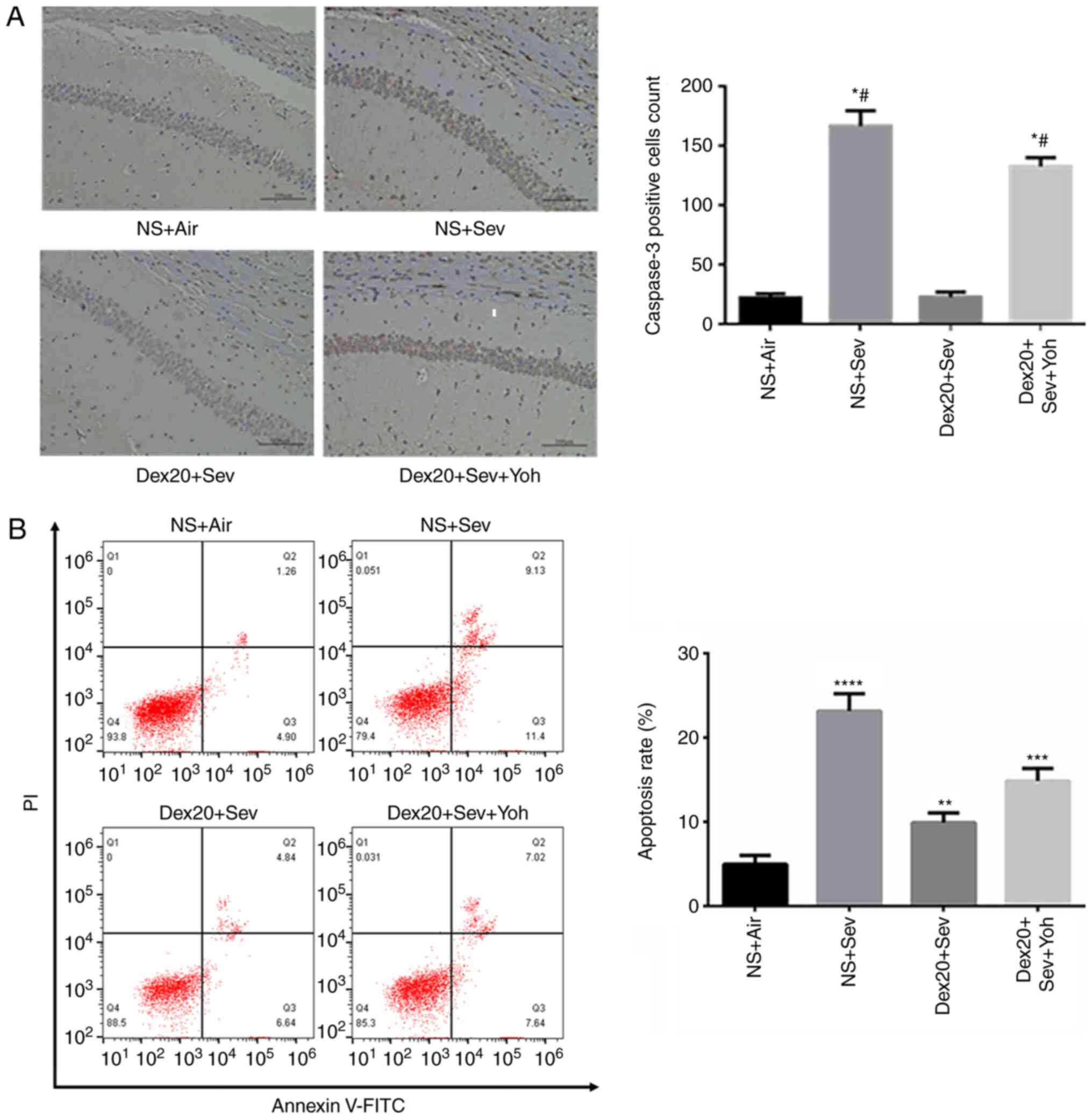 | Figure 2.Dexmedetomidine reverses
sevoflurane-induced neuroapoptosis. (A) Caspase-3 expression
increased in mice exposed to 3% sevoflurane alone, compared with NS
+ Air. Dex20 treatment reduced sevoflurane-induced neuroapoptosis.
Scale bar, 200 µm; (B) Apoptosis rate changes following
dexmedetomidine (20 µg/kg) with air, Sev or Sev + Yoh treatment.
n=6 in each group. PI/FITC. +/− (Q1), percentage of necrotic cells;
PI/FITC +/+ (Q2), percentage of late apoptotic cells; PI/FITC −/−
(Q3), percentage of viable cells; and PI/FITC −/+ (Q4), percentage
of early apoptotic cells. *P<0.05, **P<0.01; ***P<0.001;
****P<0.0001 vs. NS + Air; #P<0.05 vs. Dex20 +
Sev. Dex20, dexmedetomidine 20 µg/kg; Sev, sevoflurane; Yoh,
yohimbine; NS, normal saline; PI, propidium iodide. |
Dexmedetomidine attenuates
sevoflurane-induced proinflammatory cytokine release through α2
adrenoceptors in 6-day-old mice
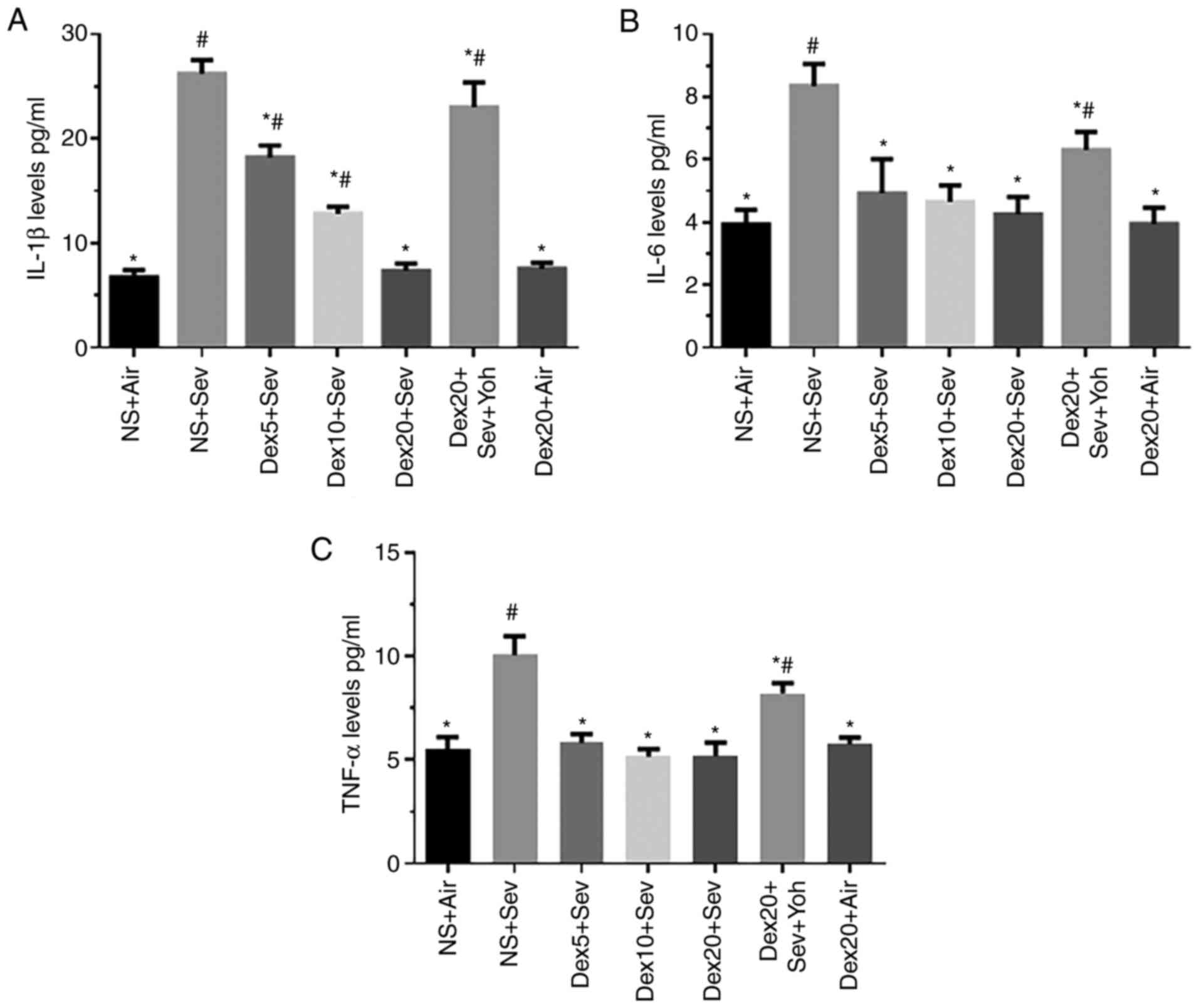
Mice exposed to 3% sevoflurane for 6 h displayed a
significant increase in IL-1β, IL-6 and TNF-α levels compared with
control mice exposed to air (Fig.
3). However, pretreatment with dexmedetomidine significantly
reduced the sevoflurane-induced release of the proinflammatory
cytokines. In particular, dexmedetomidine decreased the levels of
IL-1β in a dose-dependent manner. Yohimbine significantly increased
the levels of IL-1β, IL-6 and TNF-α, restoring the expression of
these pro-inflammatory cytokines to levels comparable to
sevoflurane alone. Thus, α2 adrenoceptors may mediate the
anti-inflammatory effect of dexmedetomidine (Fig. 3).
Dexmedetomidine attenuates
sevoflurane-induced oxidative stress through α2 adrenoceptors in
6-day-old mice
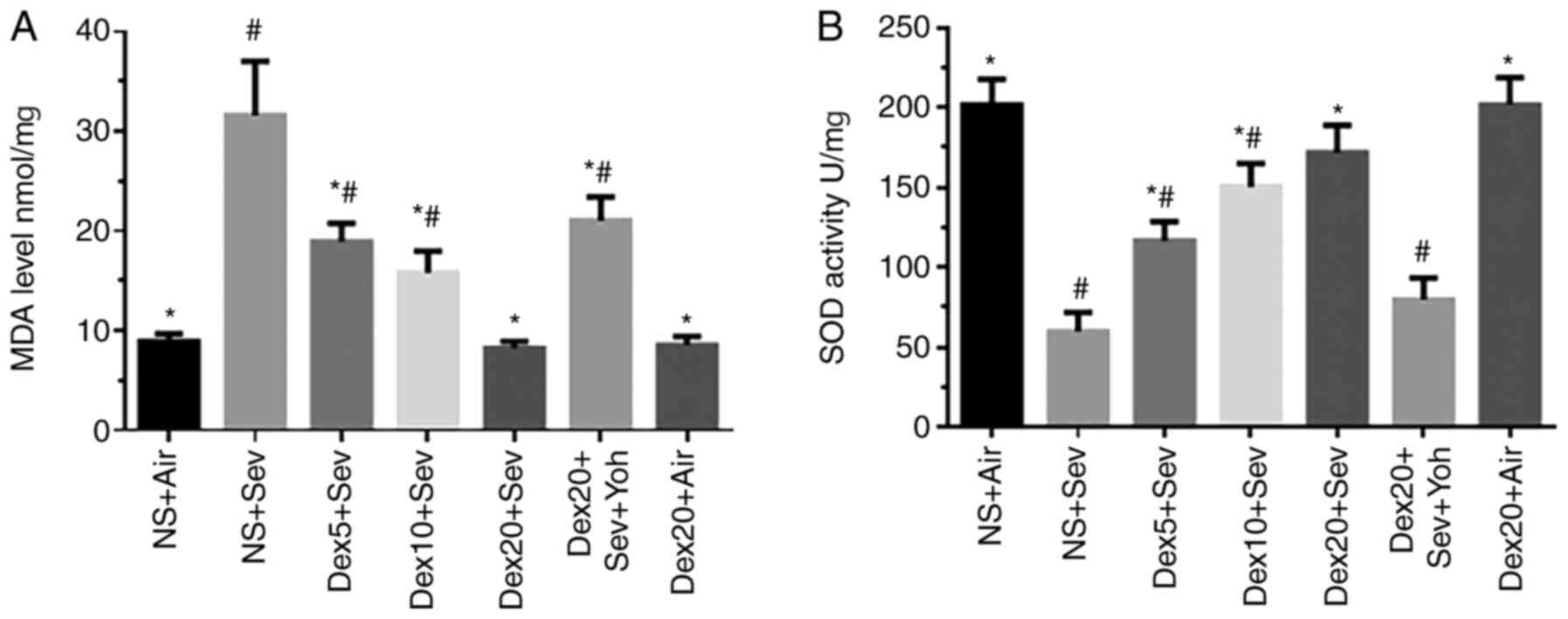
Exposure to 3% sevoflurane for 6 h significantly
increased oxidative stress, as indicated by increased MDA levels
and reduced SOD activity, compared with control mice exposed to air
(Fig. 4). By contrast, pretreatment
with dexmedetomidine significantly decreased sevoflurane-induced
oxidative stress in a dose-dependent manner. However, the
protective effects of dexmedetomidine on oxidative stress were
inhibited by yohimbine, indicating that dexmedetomidine could
modulate oxidative stress through α2 adrenoceptors.
Discussion
The present study demonstrated that pretreatment
with dexmedetomidine could attenuate sevoflurane-induced
neurotoxicity, as indicated by reduced learning and memory
impairment, and decreased neuronal cell apoptosis, inflammation and
oxidative stress. Moreover, the neuroprotective effects of
dexmedetomidine was reversed by yohimbine, an α2-adrenoceptor
antagonist, suggesting that the effects of dexmedetomidine may be
mediated by α2 adrenoceptors.
The MWM is broadly used for the evaluation of
learning and memory function in mice, particularly spatial learning
and memory (28,29). In the present study, the MWM test
was used to determine the effect of dexmedetomidine on
sevoflurane-induced cognitive impairment. Consistent with previous
studies, exposure to sevoflurane in the developing brain induced
learning and memory functional impairment in adulthood (4,30).
Shan et al (31) suggested
that dexmedetomidine ameliorated sevoflurane-induced neurocognitive
impairment. Although the methods used to evaluate neurocognitive
function differed, the present study also indicated that
dexmedetomidine could reverse sevoflurane-induced cognitive
impairment.
Neuroapoptosis is strongly associated with
neurocognitive function (32). In
the present study, sevoflurane exposure significantly increased
neuroapoptosis. Moreover, dexmedetomidine decreased neuroapoptosis
induced by sevoflurane. These findings were consistent with
previous studies by Shan et al (31) and Li et al (11), which demonstrated that
dexmedetomidine ameliorated isoflurane-induced neuroapoptosis.
In addition, cognitive impairment is associated with
neuroapoptosis, inflammation and oxidative stress (22,23,25).
Proinflammatory cytokines, such as TNF-α and IL-6, are associated
with neuroinflammation and lead to cognitive impairment following
surgery under cbupivacine anesthesia (33). In the present study, sevoflurane
increased IL-1β, IL-6 and TNF-α levels in the hippocampus, which
was reversed by dexmedetomidine pretreatment. Microglial cells, the
resident macrophages of the central nervous system, play an
important role in innate immunity and neuroinflammatory processes
in the brain (34). Although
microglial cells can promote healing, activation of microglia can
generate cytotoxic mediators, such as IL-1β, IL-6, and TNF-α, which
may be toxic to neighboring neurons (35). Thus, dexmedetomidine might function
by inhibiting the activation of microglia. However, in the present
study, the potential effect of dexmedetomidine on microglia was not
evaluated, and further study would be required to validate this
hypothesis.
An imbalance between radical-generating and
radical-scavenging systems causes oxidative stress (36). Increased reactive oxygen species
induce lipid peroxidation of polyunsaturated fatty acid in biofilms
and plasma lipoproteins, which may lead to multiple organ
dysfunction (37). Sevoflurane can
impair the function and affect the morphology of immature neuronal
mitochondria (38). In the present
study, sevoflurane increased MDA levels and decreased SOD activity
in the hippocampus. However, dexmedetomidine pretreatment decreased
MDA levels and increased SOD activity, compared with mice exposed
to sevoflurane alone. Thus, dexmedetomidine attenuated oxidative
stress.
In the present study, yohimbine, an α2 adrenoceptor
inhibitor, significantly attenuated the positive effects of
dexmedetomidine on neurocognitive impairment, neuroapoptosis,
neuroinflammation and oxidative stress. These findings suggested
that α2 adrenoceptors might mediate the protective effects
dexmedetomidine, consistent with previous studies (19,20).
In conclusion, the present study suggested that
dexmedetomidine could provide neuroprotection against
sevoflurane-induced neuroapoptosis, neuroinflammation, oxidative
stress and neurocognitive impairment by activating α2
adrenoceptors. These findings may provide insight into the
development of treatment options that could prevent neurotoxicity
caused by sevoflurane.
Acknowledgements
Not applicable.
Funding
The present study was supported by The City Council
of Science Research Plan (grant no. HAB201824).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ, ML, EC and JS conceptualized the study and
designed the experiments. YZ, ML, EC, HZ, XZ and GZ performed the
experiments. JZ and MY analyzed the data. YZ, ML, EC and JS wrote
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal procedures were approved by the Animal
Experimental Ethics Committee of The Huai'an Maternity and Child
Clinical College of Xuzhou Medical University and were performed in
strict accordance with the University Laboratory Animal
Management.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Brioni JD, Varughese S, Ahmed R and Bein
B: A clinical review of inhalation anesthesia with sevoflurane:
From early research to emerging topics. J Anesth. 31:764–778. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lerman J, Sikich N, Kleinman S and Yentis
S: The pharmacology of sevoflurane in infants and children.
Anesthesiology. 80:814–824. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Devroe S, Van de Velde M and Rex S:
General anesthesia for caesarean section. Curr Opin Anaesthesiol.
28:240–246. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shen X, Liu Y, Xu S, Zhao Q, Guo X, Shen R
and Wang F: Early life exposure to sevoflurane impairs adulthood
spatial memory in the rat. Neurotoxicology. 39:45–56. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Satomoto M, Satoh Y, Terui K, Miyao H,
Takishima K, Ito M and Imaki J: Neonatal exposure to sevoflurane
induces abnormal social behaviors and deficits in fear conditioning
in mice. Anesthesiology. 110:628–637. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Qiu J, Shi P, Mao W, Zhao Y, Liu W and
Wang Y: Effect of apoptosis in neural stem cells treated with
sevoflurane. BMC Anesthesiol. 15:252015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu F, Rainosek SW, Frisch-Daiello JL,
Patterson TA, Paule MG, Slikker W Jr, Wang C and Han X: Potential
adverse effects of prolonged sevoflurane exposure on developing
monkey brain: From abnormal lipid metabolism to neuronal damage.
Toxicol Sci. 147:562–572. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nie H, Peng Z, Lao N, Dong H and Xiong L:
Effects of sevoflurane on self-renewal capacity and differentiation
of cultured neural stem cells. Neurochem Res. 38:1758–1767. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang L, Zhang J, Yang L, Dong Y, Zhang Y
and Xie Z: Isoflurane and sevoflurane increase interleukin-6 levels
through the nuclear factor-kappa B pathway in neuroglioma cells. Br
J Anaesth. 110 (Suppl 1):i82S–i91S. 2013. View Article : Google Scholar
|
|
10
|
Zhang Y, Dong Y, Zheng H, Shie V, Wang H,
Busscher JJ, Yue Y, Xu Z and Xie Z: Sevoflurane inhibits
neurogenesis and the Wnt-catenin signaling pathway in mouse neural
progenitor cells. Curr Mol Med. 13:1446–1154. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li Y, Zeng M, Chen W, Liu C, Wang F, Han
X, Zuo Z and Peng S: Dexmedetomidine reduces isoflurane-induced
neuroapoptosis partly by preserving PI3K/Akt pathway in the
hippocampus of neonatal rats. PLoS One. 9:e936392014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Weerink MAS, Struys MMRF, Hannivoort LN,
Barends CRM, Absalom AR and Colin P: Clinical pharmacokinetics and
pharmacodynamics of dexmedetomidine. Clin Pharmacokinet.
56:893–913. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Di Cesare Mannelli L, Micheli L, Crocetti
L, Giovannoni MP, Vergelli C and Ghelardini C: α2 Adrenoceptor: A
target for neuropathic pain treatment. Mini Rev Med Chem.
17:95–107. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun L, Guo R and Sun L: Dexmedetomidine
for preventing sevoflurane-related emergence agitation in children:
A meta-analysis of randomized controlled trials. Acta Anaesthesiol
Scand. 58:642–650. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tan D, Xia H, Sun S and Wang F: Effect of
ancillary drugs on sevoflurane related emergence agitation in
children undergoing ophthalmic surgery: A Bayesian network
meta-analysis. BMC Anesthesiol. 19:1382019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Winzer-Serhan U and Leslie F: Expression
of alpha2A adrenoceptors during rat neocortical development. J
Neurobiol. 38:259–269. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Philipp M, Brede ME, Hadamek K, Gessler M,
Lohse MJ and Hein L: Placental alpha (2)-adrenoceptors control
vascular development at the interface between mother and embryo.
Nat Genet. 31:311–315. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Q, Lu R, Zhao J and Limbird LE:
Arrestin serves as a molecular switch, linking endogenous
α2-adrenergic receptor to SRC-dependent, but not SRC-independent,
ERK activation. J Biol Chem. 281:25948–25955. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dahmani S, Paris A, Jannier V, Hein L,
Rouelle D, Scholz J, Gressens P and Mantz J: Dexmedetomidine
increases hippocampal phosphorylated extracellular signal-regulated
protein kinase 1 and 2 content by an
alpha2-adrenoceptor-independent mechanismevidence for the
involvement of imidazoline I1 receptors. Anesthesiology.
108:457–466. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma D, Hossain M, Rajakumaraswamy N, Arshad
M, Sanders RD, Franks NP and Maze M: Dexmedetomidine produces its
neuroprotective effect via the α2A-adrenoceptor subtype. Eur J
Pharmacol. 502:87–97. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cibelli M, Fidalgo AR, Terrando N, Ma D,
Monaco C, Feldmann M, Takata M, Lever IJ, Nanchahal J, Fanselow MS
and Maze M: Role of interleukin-1beta in postoperative cognitive
dysfunction. Ann Neurol. 68:360–368. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cruickshank AM, Fraser WD, Burns HJ, Van
Damme J and Shenkin A: Response of serum interleukin-6 in patients
undergoing elective surgery of varying severity. Clin Sci (Lond).
79:161–165. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tan H, Bi J, Wang Y, Zhang J and Zuo Z:
Transfusion of old RBCs induces neuroinflammation and cognitive
impairment. Crit Care Med. 43:e276–e286. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Terrando N, Eriksson LI, Ryu JK, Yang T,
Monaco C, Feldmann M, Jonsson Fagerlund M, Charo IF, Akassoglou K
and Maze M: Resolving postoperative neuroinflammation and cognitive
decline. Ann Neurol. 70:986–995. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wan Y, Xu J, Ma D, Zeng Y, Cibelli M and
Maze M: Postoperative impairment of cognitive function in rats: A
possible role for cytokine-mediated inflammation in the
hippocampus. Anesthesiology. 106:436–443. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lv Q, Guo Y, Zhu M, Geng R, Cheng X, Bao
C, Wang Y, Huang X, Zhang C, Hao Y, et al: Predicting individual
responses to lithium with oxidative stress markers in drug-free
bipolar disorder. World J Biol Psychiatry. 20:778–789. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Soltesz I and Losonczy A: CA1 pyramidal
cell diversity enabling parallel information processing in the
hippocampus. Nat Neurosci. 21:484–493. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Crawley JN: Behavioral phenotyping
strategies for mutant mice. Neuron. 57:809–818. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
D'Hooge R and De Deyn PP: Applications of
the Morris water maze in the study of learning and memory. Brain
Res Brain Res Rev. 36:60–90. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao Y, Chen K and Shen X: Environmental
enrichment attenuated sevoflurane-induced neurotoxicity through the
PPAR-γ signaling pathway. Biomed Res Int. 2015:1071492015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shan Y, Yang F, Tang Z, Bi C, Sun S, Zhang
Y and Liu H: Dexmedetomidine ameliorates the neurotoxicity of
sevoflurane on the immature brain through the BMP/SMAD signaling
pathway. Front Neurosci. 12:9642018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu D, Li L and Yuan W: Neonatal anesthetic
neurotoxicity: Insight into the molecular mechanisms of long-term
neurocognitive deficits. Biomed Pharmacother. 87:196–199. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu Z, Dong Y, Wang H, Culley DJ,
Marcantonio ER, Crosby G, Tanzi RE, Zhang Y and Xie Z: Peripheral
surgical wounding and age-dependent neuroinflammation in mice. PLoS
One. 9:e967522014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Block ML: Neuroinflammation: Modulating
mighty microglia. Nat Chem Biol. 10:988–989. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qiu LL, Ji MH, Zhang H and Yang JJ, Sun
XR, Tang H, Wang J, Liu WX and Yang JJ: NADPH oxidase 2-derived
reactive oxygen species in the hippocampus might contribute to
microglial activation in postoperative cognitive dysfunction in
aged mice. Brain Behav Immun. 51:109–118. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sriram N, Kalayarasan S and Sudhandiran G:
Enhancement of antioxidant defense system by
epigallocatechin-3-gallate during bleomycin induced experimental
pulmonary fibrosis. Biol Pharm Bull. 31:1306–1311. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Del RD, Stewart AJ and Pellegrini N: A
review of recent studies on malondialdehyde as toxic molecule and
biological marker of oxidative stress. Nutr Metab Cardiovasc Dis.
15:316–328. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xu F, Armstrong R, Urrego D, Qazzaz M,
Pehar M, Armstrong JN, Shutt T and Syed N: The mitochondrial
division inhibitor Mdivi-1 rescues mammalian neurons from
anesthetic-induced cytotoxicity. Mol Brain. 9:352016. View Article : Google Scholar : PubMed/NCBI
|