Introduction
The incidence of colorectal cancer (CRC) increased
from 12.8 in 2003 to 16.8 per 100,000 in 2011 in China over the
past few decades. Although the incidence of CRC is lower in China
than that noted in other countries worldwide, the case-fatality and
mortality/incidence ratios are higher (1). Overall, ~50% of patients with CRC may
develop lymph node, liver, lung, peritoneum, brain and bone
metastases; metastasis is associated with worse survival compared
with that in patients without metastases (2). The liver is the most frequent target
organ of CRC distant metastases, and ~1/3 of all patients with CRC
present with or subsequently develop colorectal liver metastases
(3). Only a limited number of
patients with CRC present with lung metastasis. Although
metastasectomy can uniformly improve cancer-specific survival in
patients with liver metastases, it does not have the same effects
for patients with lung metastasis and combined liver and lung
metastases (4). To date, although
several studies have focused on the liver metastasis of CRC, the
mechanism underlying CRC lung metastasis is not fully
understood.
The microenvironment has been recognized to serve an
important role in tumor metastasis (5). Stromal cell reprogramming is a key
factor in the remodeling of the internal microenvironment of tumors
and in intercellular interactions (6). The stroma consists of fibroblasts,
immune cells, blood lymphatic vessels and the extracellular matrix
(ECM) (7). The complex composition
of the microenvironment, notably its dynamic feature transformed by
cancer cells, can enhance malignant progression. Mounting evidence
has revealed that fibroblasts in normal tissues function as resting
mesenchymal cells embedded within the interstitial fibrillar ECM,
which can be activated to cancer-associated fibroblasts (CAFs) in a
context-dependent manner during wound healing, tissue inflammation
and organ fibrosis (8–10). Subsequently, CAFs can function
synergistically with cancer cells to form an environment conducive
to proliferation and metastasis (11).
It has been reported that tumor-derived exosomes are
necessary for pre-metastatic microenvironment formation (12). Exosomes are a subset of
extracellular vesicles (30–150 nm) that are released from cells
upon fusion of an intermediate endocytic compartment, known as the
multivesicular body, with the plasma membrane (13). They contain proteins, mRNA,
microRNAs (miRNAs/miRs), small RNAs and/or DNA fragments that
facilitate pre-metastatic niche formation by mediating
communication between tumor cells and surrounding components, or by
horizontally transferring their contents into recipient cells
(14). miRNAs are small non-coding
RNAs, which suppress gene expression at the post-transcriptional
level via sequence-specific interactions with the 3′untranslated
region of cognate mRNA targets (15). Previous studies have reported that
exosomes contain a high level of miRNAs (16–18).
These molecules have been shown to contribute to the development of
metastasis in several types of cancer. For example,
exosome-mediated transfer of miR-193a-3p, miR-210-3p and miR-5100
has been shown to promote metastasis of lung cancer cells by
activating STAT3 signaling-induced epithelial-to-mesenchymal
transition (EMT) (19).
Tumor-derived exosomal-miR-1247-3p has also been shown to induce
CAF activation and thus facilitate lung metastasis of liver cancer
(20). However, the mechanisms
underlying the activation of fibroblasts by CRC primary cells
remain unclear, and are particularly obscure with regards to lung
metastasis of CRC cancer.
Our previous study demonstrated that miR-10a
expression was higher in primary CRC tissues compared with that
noted in lymph node metastatic tissues. In addition, miR-10a was
shown to suppress CRC metastasis to the liver by modulating EMT and
the process of anoikis of CRC cells in vitro and in a nude
mouse model (21). In the present
study, the function of exosomal-miR-10a derived from CRC primary
cells was investigated on a normal human lung fibroblast (NHLF)
cell line established from the normal lung tissues. Initially, the
expression levels of miR-10a were detected in serum and cancer
tissues from patients with CRC, and the analysis aimed to assess
whether miR-10a may be considered a blood-based biomarker for the
diagnosis of patients with CRC. Subsequently, a NHLFs cell line was
established from a patient with lung fibroma, and the ability of
NHLFs to absorb exosomes from SW480 cells and the effects of
exosomal-miR-10a on cell proliferation, migration and
pro-inflammatory cytokine expression were assessed.
Materials and methods
Clinical specimens and cell lines
Human serum was collected from 20 healthy subjects
and 40 patients with CRC who had not received chemotherapy or
radiotherapy prior to radical resection at Tangshan People's
Hospital (Tangshan, China). The patients with CRC were aged between
32 and 78 years old and included male and female patients, they
were admitted to the Tangshan People's Hospital between April and
October 2017. The CEA, CA-199 and CA-724 of the patients with CRC
were detected by an electrochemical luminescence analyzer (E601;
Roche Diagnostics) when they were admitted to the hospital. The TNM
stage was determined according to The Eighth Edition AJCC Cancer
Staging Manual (22). The healthy
subjects were between 41 and 73 years old, which included male and
female subjects, from the same hospital. Furthermore, 15 pairs of
tissue samples from the 40 patients with CRC were collected
(including adjacent normal tissues and cancer tissues). The use of
clinical samples was approved by the Ethical Committee of Tangshan
People's Hospital, and written informed consent was obtained from
the individuals.
The NHLF cell line was established via the tissue
block adherent method using tissues obtained from a 51-year-old
male patient pathologically diagnosed with benign lung tumors in
August 2017. The collection of a fresh clinical sample from this
patient was approved by the Ethical Committee of Tangshan People's
Hospital and written informed consent was obtained. Firstly, fresh
normal lung tissue was collected >2.0-cm away from the lesion
and rapidly added to Dulbecco's modified Eagle's F12 medium
(DMEM/F12; Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% fetal bovine serum (FBS; Gibco; Thermo Fisher Scientific,
Inc.), 100 U/ml penicillin and 0.1 mg/ml streptomycin. Within 20
min, the sample was washed twice with PBS, cut into
1.5–2.0-mm3 fragments and centrifuged at 200 × g for 5
min at 26°C. The small lung tissue fragments (1.5–2.0
mm3) were then collected into a cell culture flask
filled with DMEM/F12 supplemented with 20% FBS, 100 U/ml penicillin
and 0.1 mg/ml streptomycin, and cultured at 37°C in a humidified
incubator with 5% CO2 for 1 h. Subsequently, the culture
flask was inverted for 24 h, after which the culture flask was
placed in the humidified incubator and the culture medium was
replaced with DMEM/F12 supplemented with 10% FBS, 100 U/ml
penicillin and 0.1 mg/ml streptomycin. During the process of cell
culture, DMEM/F12 was replaced every 3–4 days until cell confluence
reached 80%. For the present study, passage 3–7 primary NHLFs were
used.
The human colon cancer SW480 cell line was donated
by Professor Hua Tang (Tianjin Medical University, Tianjin, China).
The cells were conventionally cultured in minimum essential medium
α (Gibco; Thermo Fisher Scientific, Inc.) containing 10% FBS, 100
U/ml penicillin and 100 µg/ml streptomycin, and incubated at 37°C
in a humidified incubator supplemented with 5% CO2.
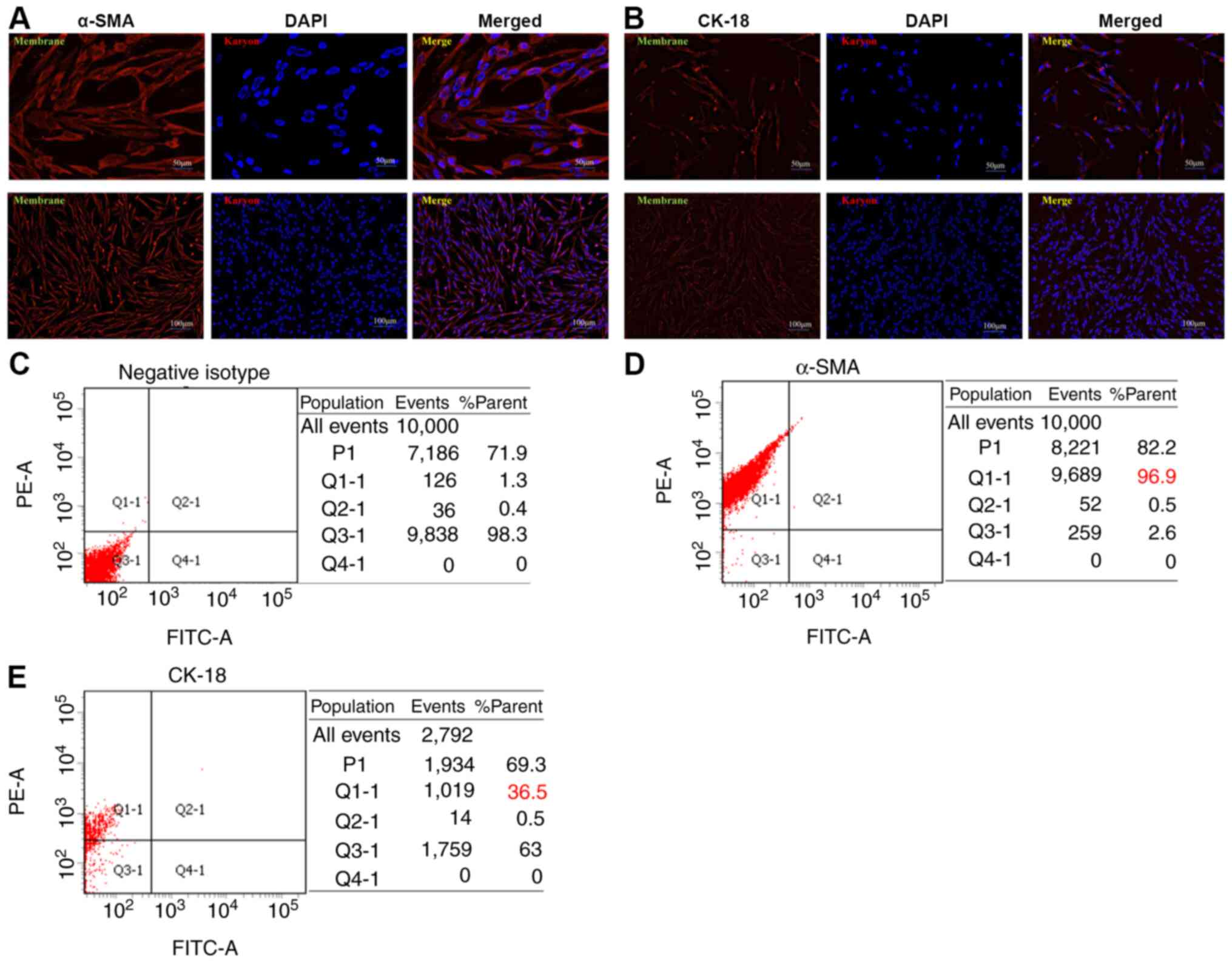
Cell immunofluorescence
Third generation NHLFs were identified by cell
immunofluorescence. Cells were seeded into 6-well plates at a
density of 50% and incubated in a 37°C incubator containing 5%
CO2 for 3–5 days until they had reached 80% confluence.
Subsequently, cells were fixed with 2% paraformaldehyde in PBS for
10 min, permeabilized for 10 min in PBS-0.01 Triton X-100
(Sigma-Aldrich; Merck KGaA), and then blocked with 5% bovine serum
albumin (cat. no. 0332-100G; Amresco LLC) to suppress non-specific
reactions at room temperature for 60 min. Cells were then incubated
with anti-cytokeratin-18 (CK-18) (1:500; cat. no. ab32118; Abcam)
and anti-α-smooth muscle actin (α-SMA; 1:500; cat. no. SRP05217;
Tianjiin Saierbio) overnight at 4°C, followed by incubation with
Alexa Fluor® 555-conjugated secondary anti-rabbit
antibody (1:500; cat. no. A-21-430; Invitrogen; Thermo Fisher
Scientific, Inc.) for 60 min at room temperature in the dark. The
negative isotype was incubated with PBS in the same conditions as
the CK-18 and α-SMA antibodies. Finally, DAPI was used for nuclear
staining at room temperature for 2 min. Cells were observed under
an IX71 fluorescence microscope (Olympus Corporation).
Flow cytometry
For identifying primary cells, third generation
NHLFs were cultured in DMEM/F12 (Gibco; Thermo Fisher Scientific,
Inc.) with 10% FBS, 100 U/ml penicillin and 0.1 mg/ml streptomycin
in 6-well plates for 3–5 days, and 1×106 cells were
harvested to detect α-SMA and CK-18 cell makers. Cells were blocked
with blocking buffer (0.5% BSA in 1X PBS) after one wash, and were
fixed with 2% paraformaldehyde at room temperature for 10 min.
Subsequently, cells were washed once and permeabilized with 0.5 ml
permeabilization solution (cat. no. 340973; BD Biosciences) at room
temperature for 10 min. Cells were then washed once, incubated in
blocking buffer for 30 min at room temperature, and incubated with
α-SMA (1:20; cat. no. SRP05217; Tianjiin Saierbio) and CK-18 (1:20;
cat. no. ab32118; Abcam) primary antibodies for 30 min at room
temperature. Finally, cells were washed twice and incubated with
Alexa Fluor® 555-conjugated secondary antibody (1:500)
for 30 min at room temperature. Cells were resuspended in 1X PBS
and analyzed using a BD FACSAria™ II (BD Biosciences) with BD
FACSDiva™ software (BD Biosciences).
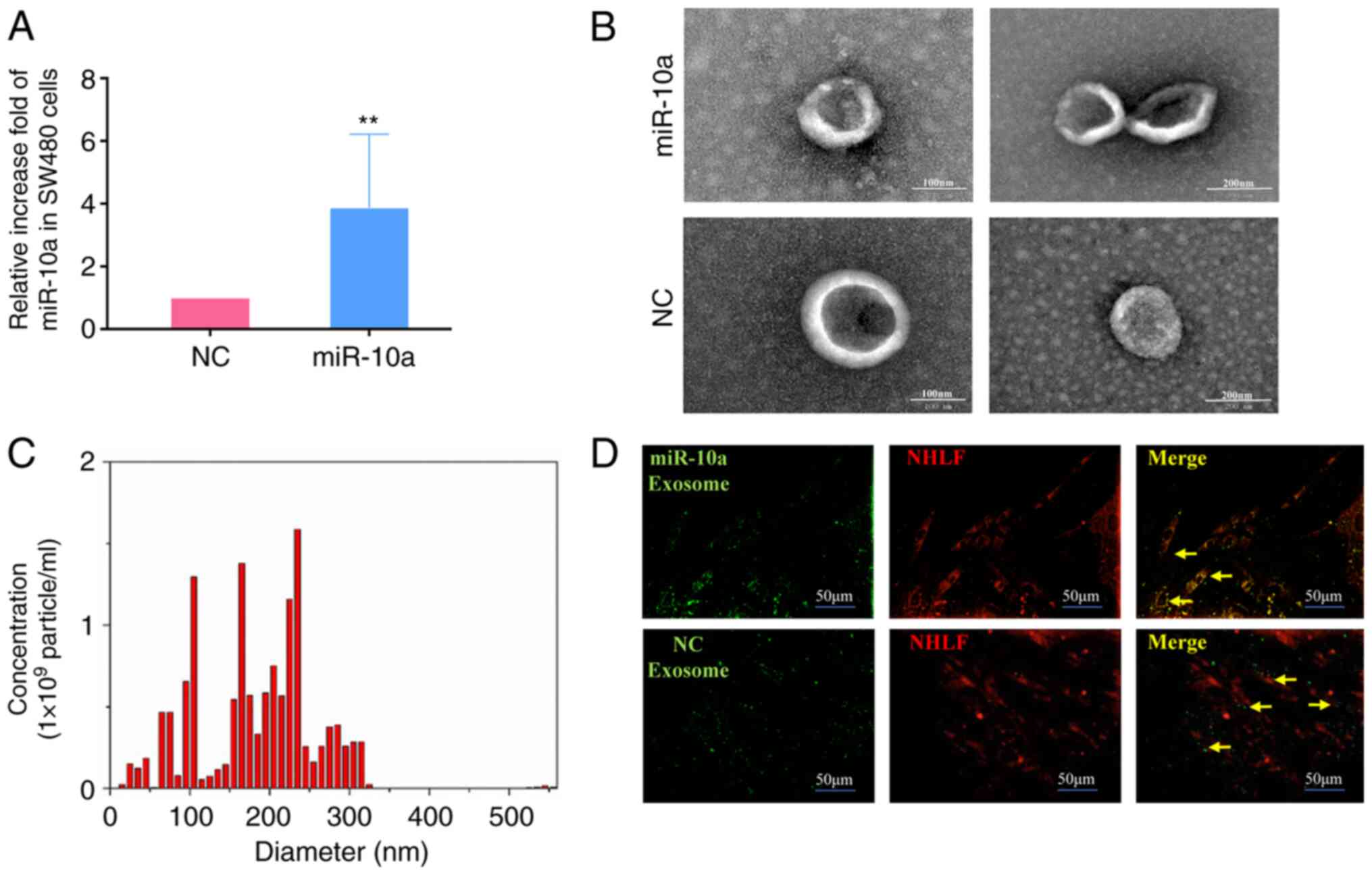
Extraction and identification of
exosomes
SW480 cells at a density of 80% were cultured in
cell culture flasks containing fresh DMEM/F-12 supplemented with
exosome-depleted FBS. The cells were transfected with miR-10a
mimics (5′-UACCCUGUAGAUCCGAAUUUGUGCAAAUUCGGAUCUACAGGGUAUU-3′) and
negative control (NC) mimics
(5′-UUCUCCGAACGUGUCACGUTT3ACGUGACACGUUCGGAGAATT-3′), purchased from
Shanghai GenePharma Co., Ltd., at 37°C for 48 h, according to the
manufacturer's protocol, using Lipofectamine® 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). After 48 h,
supernatants from the different groups were collected, centrifuged
at 300 × g at 25°C for 10 min and filtered through 0.45-µM filters.
Supernatants were again collected, and centrifuged at 2,000 × g for
10 min, 10,000 × g for 30 min and twice at 100,000 × g for 70 min
using the Optima XPN-100 (Beckman Coulter, Inc.) all at room
temperature. The exosome pellet was resuspended in PBS,
ultra-centrifuged at 100,000 × g for 70 min, resuspended in 100 µl
1X PBS and stored at −80°C. Exosomes were observed under a
JEM-1200EX transmission electron microscope (JEOL, Ltd.) and
quantified using NanoSight LM10 (Malvern Instruments, Ltd.), which
was conducted by Guangzhou EpiBiotek Co., Ltd..
RNA extraction and reverse
transcription quantitative PCR (RT-qPCR)
The blood samples were collected from the patients
with CRC, and then the serums were extracted from the blood at room
temperature at 200 × g for 10 min within 30 min. Total RNA was
extracted from tissues and serum using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. RNA was tested for quality via 1% agarose
gel electrophoresis. cDNA was synthesized using the PrimeScript™
1st Strand cDNA Synthesis kit including DNase (Takara Bio., Inc.)
from 3 µg total RNA with a thermocycler (Arktik™ 96; Thermo Fisher
Scientific, Inc.) Stem-loop RT-qPCR (Suzhou GenePharma Co., Ltd.)
analysis for the detection of miR-10a in tissues, cells and
exosomes was conducted as previously described (21). The qPCR assay was performed as
follows: 95°C for 3 min, and 40 cycles of 95°C for 12 sec and 62°C
for 40 sec. The fold-change was analyzed using the
2−∆∆Cq method (23).
Cel-miR-39 (Shanghai GenePharma Co., Ltd.) is a miRNA that is not
expressed in human cells, so it was used as an external parameter
for miR-10a in serum samples, and U6 was used as a housekeeping
gene for miR-10a in tissues and cells.
TRIzol was used for RNA extraction from the NHLF
cells. RT-qPCR analysis of IL-6, IL-8 and IL-1β was performed using
PrimeScript™ 1st Strand cDNA Synthesis kit (Takara Bio, Inc.) and
random primers for RT. The process of RT was performed at 65°C for
10 min, 25°C for 5 min, 0°C for 2 min, 42°C for 30 min, 70°C for 10
min. qPCR was conducted using SYBR Premix Ex Taq (Takara Bio, Inc.)
and a PikoReal 96 RT-PCR system (Thermo Fisher Scientific, Inc.).
qPCR was performed at 95°C for 4 min, then 33 cycles at 95°C for 1
min, 55°C for 1 min and 72°C for 1 min, and finally 72°C for 10
min. β-actin was used to normalize the expression levels of IL-6,
IL-8 and IL-1β. The primer sequences are provided in Table I. All experiments were carried out
at least in triplicate (24).
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′-3′) |
|---|
| miR-10a | F:
TGCGGTACCCTGTAGATCCG |
|
| R:
CCAGTGCAGGGTCCGAGGT |
| U6 | F:
ATTGGAACGATACAGAGATT |
|
| R:
GGAACGCTTCACGAATTTG |
| Cel-miR-39 |
UCACCGGGUGUAAAUCAGCUUG |
| IL-6 | F:
ACTCACCTCTTCAGAACGAATTG |
|
| R:
CCATCTTTGGAAGGTTCAGGTTG |
| IL-8 | F:
TTTTGCCAAGGAGTGCTAAAGA |
|
| R:
AACCCTCTGCACCCAGTTTTC |
| IL-1β | F:
ATGATGGCTTATTACAGTGGCAA |
|
| R:
GTCGGAGATTCGTAGCTGGA |
| β-actin | F:
ACTGTGCCCATCTACGAGG |
|
| R:
GAAAGGGTGTAACGCAACTA |
Trace experiment of exosomes
SW480 cells in the miR-10a mimics and NC groups were
pretreated with 6.5 mmol/l DiO (Beyotime Institute of
Biotechnology) at 37°C for 20 min, according to the manufacturer's
instructions. Subsequently, the exosomes were collected from the
SW480 supernatants as aforementioned. The obtained DiO-labeled
exosomes were incubated with NHLFs pretreated with 6.5 mmol/l DiL
(Beyotime Institute of Biotechnology) at 37°C for 10 min. The
location of exosomes was observed using an IX71 fluorescence
microscope (Olympus Corporation).
Proliferation assay
NHLFs (4×103/well) were seeded into
96-well plates, and equal quantities of exosomes from SW480 cells
in the miR-10a mimics and NC groups were added into the wells with
an exosome concentration of 10 µg/ml. To detect the proliferative
ability of NHLFs, 10% CCK-8 (100 µl; Dojindo Molecular
Technologies, Inc.) was added into the well and incubated with
NHLFs at 37°C for 3 h. Absorbance was measured using a microplate
reader at a wavelength of 450 nm (Multiskan FC; Thermo Fisher
Scientific, Inc.). Each experiment was carried out in three
replicate wells and was repeated three times.
Transwell migration assay
NHLFs were plated in 24-well Transwell plates at a
density of 1×104/well in DMEM/F12 with 10% FBS, 100 U/ml
penicillin and 0.1 mg/ml streptomycin. The medium in the upper
chamber was free of FBS, whereas the medium in the lower chamber
was supplemented with 10% FBS. For detecting exosome function, 6 µg
SW480-derived exosomes were added to the inserts. After incubation
at 37°C for 48 h, the cell inserts were fixed with 33% (v/v) acetic
acid (glacial acetic acid: Methyl alcohol was 1:3) and stained with
0.1% crystal violet (Beijing Solarbio Science & Technology Co.,
Ltd.). Both the fixing and staining assays were conducted at room
temperature for 10 min. Images of representative fields were
captured with a light microscope, and the number of migrated cells
per field was counted. The procedure was conducted as previously
described (21), and the data were
analyzed from three different wells per group.
Wound-healing assay
NHLFs at a density of 1×105 per well were
plated into six-well plates in DMEM/F12 supplemented with 3% FBS,
100 U/ml penicillin and 0.1 mg/ml streptomycin. The SW480-derived
exosomes at a concentration of 10 µg/ml were added to each well at
37°C for 24 and 48 h. Subsequently, a wound was generated on the
cell monolayers using a 200-µl pipette tip, and the NHLFs that
migrated into the wounded area were observed under a light
microscope at the specific time points. Wound closure was
calculated with ImageJ software (version 1.46; National Institutes
of Health).
Expression of inflammatory
factors
To detect the effect of exosomes on the expression
levels of cytokines, an equal number (1×106/flask) of
NHLFs was plated in a 25 ml cell culture flask, and miR-10a
exosomes and NC exosomes of the same quantities were added to the
flasks. After incubation at 37°C for 48 h, the NHLFs in each group
were collected and RT-qPCR was performed to detect the expression
levels of the cytokines IL-6, IL-8 and IL-1β.
Statistical analysis
Data analysis was performed using SPSS software
version 17.0 (SPSS, Inc.). Each experiment was carried out at least
in triplicate and all results are presented as the mean ± SD. A
paired Student's t-test was used to compare the paired tissue
samples. Student's t-test was used to assess statistical
significance. Significant association between miR-10a expression
and clinicopathological parameters were assessed using the
independent samples t-test. An ANOVA was performed to compare three
groups followed by Scheffe post hoc test. P<0.05 was considered
to indicate a statistically significant difference.
Results
miR-10a levels are expressed at lower
levels in the serum and cancer tissues of patients with CRC
Our previous study discovered that miR-10a was
highly expressed in primary CRC tissues compared with the
corresponding expression in metastatic lymph node tissues (21). Therefore, the present study examined
whether miR-10a was a biomarker for early diagnosis of patients
with CRC. The expression levels of miR-10a were detected in mixed
serum samples from 40 patients with CRC and 20 healthy subjects.
The expression levels of miR-10a were lower in 32 out of the 40
serum samples from patients with CRC compared with those in the
serum samples from the healthy subjects (not all 40 patient samples
are shown; Fig. 1A). The expression
levels of miR-10a in the serum samples from patients with CRC were
negatively associated with the invasion depth of CRC; however, no
significant association was noted between miR-10a expression and
other clinicopathological indices (Table II).
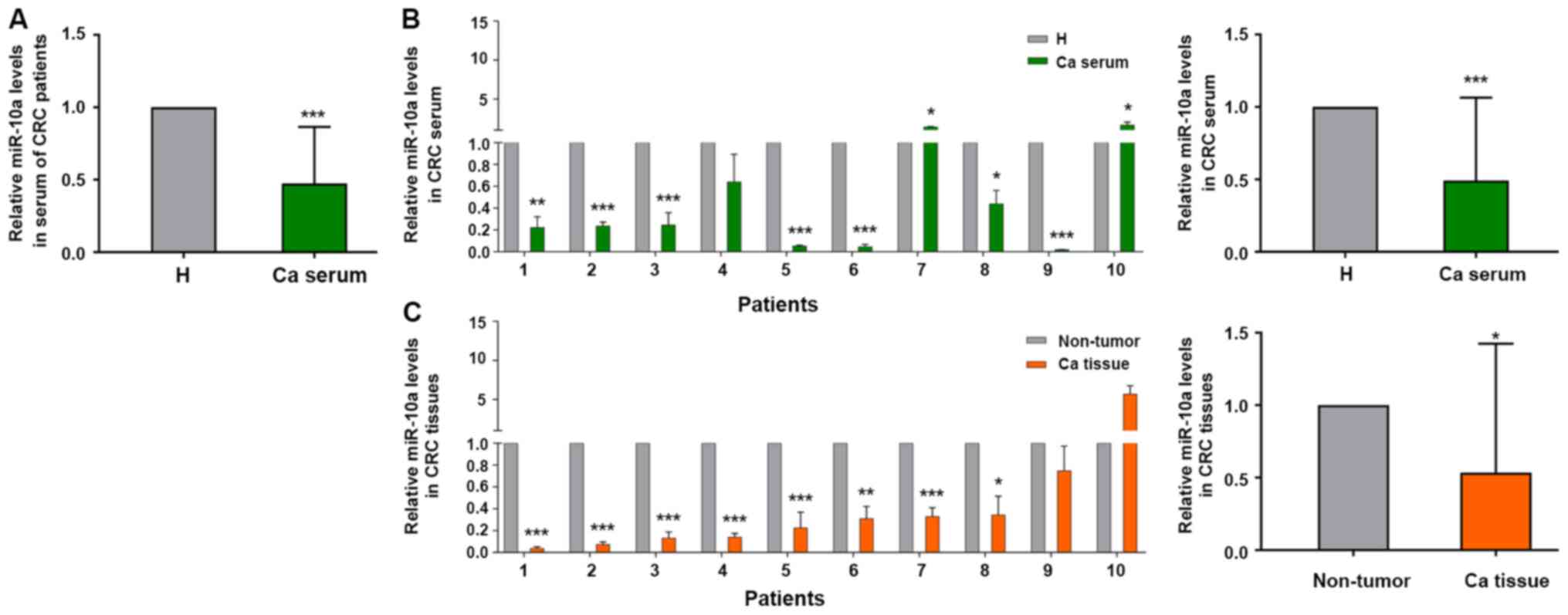 | Figure 1.Relative expression levels of miR-10a
in serum and cancer tissues from patients with CRC. (A) Reverse
transcription-quantitative PCR analysis of miR-10a in serum samples
from patients with CRC compared with serum from healthy subjects,
nCRC=40, nH=20. (B) Expression levels of
miR-10a in serum samples from randomly selected patients with CRC
and healthy subjects, n=10. *P<0.05, **P<0.01, ***P<0.001
vs. H. (C) Expression levels of miR-10a in cancer tissues compared
with adjacent non-tumor tissues from the same patients with CRC
patient as (B), n=10. The expression of miR-10a in serum and
tissues from patients with CRC was normalized to cel-miR-39 and U6
snRNA, respectively. The results are presented as the mean ± SD.
All experiments were performed at least in triplicate, and
Student's t-test was used to analyze the data. *P<0.05,
**P<0.01, ***P<0.001 vs. non-tumor tissues. Ca, patients with
CRC; H, healthy subjects; CRC, colorectal cancer; miR-10a,
microRNA-10a. |
| Table II.Association between miR-10a serum
expression and clinicopathological features of patients with
colorectal cancer. |
Table II.
Association between miR-10a serum
expression and clinicopathological features of patients with
colorectal cancer.
| Variables | N (40) | miR-10a
expression |
P-valuesa |
|---|
| Age, years |
|
| 0.861 |
|
≤65b | 20 | 0.49±0.44 |
|
|
>65 | 20 | 0.46±0.35 |
|
| Sex |
|
| 0.172 |
|
Male | 23 | 0.42±0.40 |
|
|
Female | 17 | 0.57±0.34 |
|
| Tumor
sitec |
|
| 0.978 |
|
Colon | 20 | 0.48±0.43 |
|
|
Rectum | 20 | 0.47±0.36 |
|
| Tumor type |
|
| 0.372 |
|
Adenocarcinoma | 37 | 0.49±0.39 |
|
|
Mucinouse | 3 | 0.37±0.28 |
|
| Invasion depth |
|
| 0.039d |
|
T1+T2 | 11 | 0.30±0.26 |
|
|
T3+T4 | 29 | 0.58±0.37 |
|
| No. of positive
nodes |
|
| 0.558 |
| 0 | 29 | 0.41±0.33 |
|
|
1-3 | 6 | 0.53±0.45 |
|
| 3+ | 5 | 0.65±0.55 |
|
| Distant
metastasis |
|
| 0.095 |
| M0 | 38 | 0.45±0.39 |
|
| M1 | 2 | 0.92±0.16 |
|
| TNM stage |
|
| 0.301 |
|
I+II | 27 | 0.43±0.38 |
|
|
III+IV | 13 | 0.57±0.42 |
|
| CEA, ng/ml |
|
| 0.593 |
|
0-5 | 22 | 0.44±0.33 |
|
|
>5 | 18 | 0.51±0.46 |
|
| CA-199, U/ml |
|
| 0.611 |
|
0-39 | 36 | 0.46±0.39 |
|
|
>39 | 4 | 0.57±0.45 |
|
| CA-724, U/ml |
|
| 0.260 |
|
0-6.9 | 37 | 0.49±0.39 |
|
|
>6.9 | 3 | 0.27±0.23 |
|
A total of 15 pairs of tissue samples were selected
from the 40 patient samples, in order to detect the expression
levels of miR-10a. After carefully checking the integrity of the
tissue RNA, only 10 cases from these samples could be used for
further study (Fig. 1B). The
expression levels of miR-10a were detected in serum and tissue
samples. The expression levels of miR-10a were lower in the serum
samples from patients with CRC compared with those detected in the
healthy subjects (Fig. 1B). A total
of eight patients out of 10 exhibited lower miR-10a expression in
cancer tissues compared with those noted in the adjacent normal
tissues, which was consistent with the results obtained from serum
samples (Fig. 1C). These results
indicated that miR-10a expression was lower in serum and cancer
tissue samples from patients with CRC, thus suggesting that this
miRNA may be a potential biomarker for the diagnosis of CRC.
Establishment and identification of
NHLFs
Fibroblasts have been recognized to be the dominant
component of the tumor stroma. Previous studies have suggested a
prominent functional role for these cells in cancer progression and
metastasis (25–27). In the present study, NHLFs were
established using the tissue block adherent method and the third
generation of NHLFs was identified. The NHLFs exhibited classic
spindle-shape morphology with a potential for planar polarity
(Fig. 2A and B). α-SMA and CK-18
are the classic makers used for human fibroblast identification.
The expression levels of α-SMA and CK-18 were examined in NHLFs by
cellular immunofluorescence assays. α-SMA was expressed mainly in
the cytoplasm and cellular membrane, whereas it was weakly
expressed in the cell nucleus of NHLFs (Fig. 2A). CK-18 was weakly expressed in the
cytoplasm, cellular membrane and cell nucleus of NHLFs (Fig. 2B). Consistent with the
aforementioned results, flow cytometry indicated that the ratios of
negative isotype (Fig. 2C), α-SMA
(Fig. 2D) and CK-18 (Fig. 2E) expressed in NHLFs were 96.9 and
36.5%, respectively. Since low expression of CK-18 in NHLFs is a
known marker for human epithelial cells (28,29),
the expression analysis of these markers indicated that primary
NHLFs were successfully established.
Identification and tracer technique of
exosomes from SW480 cells
To further investigate the effects of miR-10a
derived from SW480 cell exosomes, miR-10a was overexpressed in
these cells by transfection with miR-10a mimics. Subsequently,
exosomes were isolated. RT-qPCR indicated that the expression
levels of miR-10a were significantly increased in exosomes from
transfected cells compared with those noted in control subjects
(Fig. 3A). Subsequently, electron
microscopy and NanoSight equipment were employed to identify the
shape and size distribution of the exosomes secreted from SW480
cells. The exosomes exhibited a hemispherical shape (Fig. 3B and C). The diameters of all
exosomes were mainly concentrated at 0–300 nm and the majority of
them were centered at 100–200 nm. Furthermore, it was shown that
exosomes released by SW480 cells could be absorbed by NHLFs
(Fig. 3D). Taken together, these
data suggested that exosomes carrying miR-10a could form an
important tool for communication between primary CRC cells and
NHLFs.
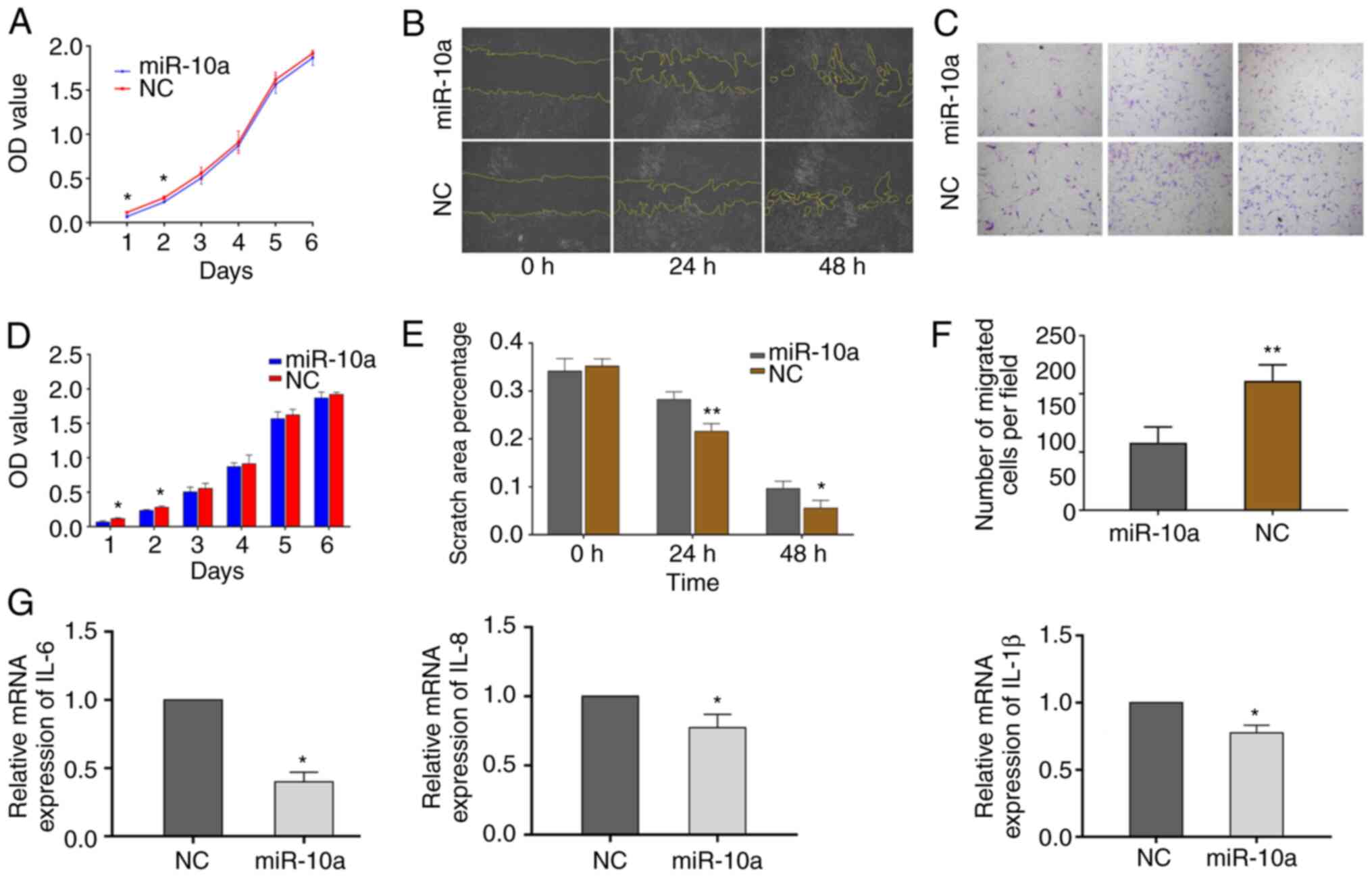
Exosomal-miR-10a from SW480 cells
reduces NHLF proliferation, migration, and IL-6, IL-8 and IL-1β
expression
In order to investigate the effect of
exosomal-miR-10a from the primary CRC cells on fibroblasts in lung
metastatic loci, the effects of exosomes derived from
miR-10a-overexpressing SW480 cells were examined on NHLF features,
including cell proliferation, migration and the expression of
pro-inflammatory cytokines. NHLFs were respectively treated with
the same quantity of exosomes from the miR-10a mimics and NC
groups. The proliferative activity of the miR-10a group NHLFs was
significantly decreased compared with that of the NC group within
the first 2 days (Fig. 4A and D).
In addition, wound-healing assays indicated that NHLFs exhibited
reduced migratory activity when treated with exosomes
overexpressing miR-10a at 24 and 48 h (Fig. 4B and E). The Transwell migration
assay indicated similar results as those noted from the
wound-healing assay at 48 h (Fig. 4C
and F). In order to determine the effects of exosomes derived
from miR-10a-overexpressing SW480 cells on the expression of
inflammatory cytokines in NHLFs, the mRNA expression levels of
IL-6, IL-8 and IL-1β were detected. The results demonstrated that
all three inflammatory cytokines exhibited decreased expression
levels in the miR-10a-overexpressing group (Fig. 4G). These data suggested that
exosomal-miR-10a from SW480 cells may not only reduce NHLF
proliferation and migration, but could also inhibit the mRNA
expression levels of IL-6, IL-8 and IL-1β.
Discussion
It is well known that cancer metastasis is a very
complex process. Various hypotheses have been proposed to explain
this biological process. It has been proposed that metastatic
dissemination largely depends on mechanical factors that result
from the anatomical structure of the vascular system (30). In addition, it has been reported
that cancer metastasis is initiated by numerous subpopulations of
cells that have different biological characteristics, including
metastatic potential (31). The
most accepted hypothesis is Paget's ‘seed and soil’ hypothesis,
which suggests that the outcome of metastasis is not due to chance,
but due to certain tumor cells having specific affinity for the
microenvironment of certain organs (32). Previously, the ‘seed and soil’
hypothesis was revised, and it has been proposed that primary
neoplasms and metastases consist of both tumor cells and
microenvironment cells (33). In
the present study, fibroblasts were used, which are dominant,
long-lived and highly plastic cells present within the tumor
microenvironment. The experiments aimed to determine the mechanism
by which primary tumor cell exosomal-miR-10a regulates lung
metastasis in patients with CRC.
Previous studies have suggested that miR-10a
expression is upregulated in various tumor types, including breast
(34), pancreatic cancer (35), hepatocellular carcinoma (36) and non-small cell lung cancer (NSCLC)
(37,38). In addition, higher expression levels
of miR-10a have been reported to be positively correlated with
advanced tumor stage and positive lymph node metastasis in NSCLC
(38). In contrast to these
observations, a different role of miR-10a was reported in our
previous study, which revealed a negative correlation between
miR-10a expression and distant metastasis and invasion depth in
patients with CRC (21). In the
present study, results indicated that miR-10a expression was lower
in the serum samples of patients with CRC compared with the levels
noted in healthy subjects. In addition, lower expression levels of
miR-10a were detected in cancer tissues compared with those noted
in the paired adjacent normal tissues of patients with CRC. The
expression of miR-10a was also inversely associated with the
invasion depth of CRC. The different functions of miR-10a in
various types of cancer may be due to the biological heterogeneity
of tumor cells. Due to these differences, our future studies aim to
provide a deeper understanding of the effects of miR-10a on lung
metastasis of CRC. Additional experiments will be conducted in the
future to clarify the mechanism underlying lower miR-10a expression
in the serum of patients with CRC.
To address the role of miR-10a in the process
underlying lung cancer metastasis of CRC, NHLFs were established
and identified. Fibroblasts are associated with all stages of
disease progression, including cancer metastasis, and they are
considered a component of the general host response to tissue
damage caused by cancer cells (39). Notably, a more accurate definition
of a fibroblast is a resting mesenchymal cell with the potential to
be activated by appropriate stimuli and thus become a mesenchymal
stem cell (39). In the present
study, exosomes were used, which are considered a significant
factor within the microenvironment, to investigate the specific
function of miR-10a on regulating the crosstalk between the primary
CRC tumor site and lung metastatic loci. The results revealed that
miR-10a in exosomes secreted by CRC cells could suppress the
proliferative activity of NHLFs. In contrast to these findings,
miR-10a-overexpressing exosomes promoted cell proliferative
activity within 2 days, and there was no significant difference in
proliferation between the miR-10a and NC groups after 2 days, which
was possibly caused by non-continuous supplementation of exosomes.
In addition, the data demonstrated that miR-10a reduced the
migratory ability of NHLFs within 48 h, as determined by
wound-healing and Transwell assays. These reduced proliferative and
migratory capabilities of NHLFs may be adverse factors for CRC
primary cell metastasis.
IL-6 is a pro-inflammatory cytokine that has been
positively associated with tumor progression and metastasis in
various types of cancer (40,41).
It has been reported that high levels of IL-6 and IL-8 in the
circulation are associated with decreased overall survival of
patients with melanoma (42). The
present study demonstrated that exosomal-miR-10a derived from SW480
cells reduced IL-6 and IL-8 expression levels in NHLFs. IL-1β
facilitates invasion and extravasation in the early stages of
metastasis (43), and promotes EMT
(44). The IL-1β inflammatory
response has been reported to be driven by primary breast cancer
and is considered to be dissemination-supportive and
colonization-suppressive factor, thus leading to the suppressed
metastasis of breast cancer (45).
Moreover, microenvironment-secreted IL-1β may promote breast cancer
metastatic colonization in the bone via activation of Wnt signaling
(46). In the present study, IL-1β
expression was downregulated in NHLFs treated with exosomes from
SW480 cells overexpressing miR-10a, thus suggesting that the IL-1β
expression of NHLFs may mediate an inflammatory response, which may
partly contribute to CRC cell metastasis. The present study was
limited by the lack of evaluation of downstream pathway mechanisms
of miR-10a, which could reduce proliferation and migration of NHLFs
and decrease the expression of inflammatory cytokines. The exact
mechanism will be assessed in follow-up studies.
In conclusion, the present study provided evidence
that miR-10a was expressed at lower levels in the serum and cancer
tissues of patients with CRC, and its expression was inversely
associated with invasion depth of CRC. Furthermore,
exosomal-miR-10a derived from CRC cells reduced the proliferative
and migratory activities, and the expression of IL-6, IL-8 and
IL-1β, in NHLFs. These data may provide a novel insight into the
mechanism underlying lung metastasis and may provide molecular
biomarkers for the assessment of tumor metastasis to the lung
tissues of patients with CRC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Talent
Training Subsidy Project of Hebei Province (grant no. A201902029),
the Natural Science Foundation of Hebei Province (grant no.
H2018105049) and the Science and Technology Project of Tangshan
City (grant no. 19150246E).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YufengL and YankunL conceived the project and
supervised the experiments. JW, YingL and XZ performed the cell
experiments and extraction exosomes assay. YuantingL and JG
performed the surgery to collect the samples and performed the
RT-qPCR assay. YanL, ZW and JZ collected the clinical serum and
tissue samples and the information of patients with CRC for the
experiments, and performed the FACS assay. YW and WH performed
statistical analysis. YankunL and YufengL wrote the manuscript with
help from all of the authors. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
The use of clinical samples was approved by the
Ethical Committee of Tangshan People's Hospital, and written
informed consent was obtained from the individuals.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Zhu J, Tan Z, Hollis-Hansen K, Zhang Y, Yu
C and Li Y: Epidemiological trends in colorectal cancer in China:
An ecological study. Dig Dis Sci. 62:235–243. 2017. View Article : Google Scholar
|
|
2
|
Chakedis J and Schmidt CR: Surgical
treatment of metastatic colorectal cancer. Surg Oncol Clin N Am.
27:377–399. 2018. View Article : Google Scholar
|
|
3
|
Engstrand J, Nilsson H, Strömberg C, Jonas
E and Freedman J: Colorectal cancer liver metastases-a
population-based study on incidence, management and survival. BMC
Cancer. 18:782018. View Article : Google Scholar
|
|
4
|
Siebenhüner AR, Güller U and Warschkow R:
Population-based SEER analysis of survival in colorectal cancer
patients with or without resection of lung and liver metastases.
BMC Cancer. 20:2462020. View Article : Google Scholar
|
|
5
|
Quail DF and Joyce JA: Microenvironmental
regulation of tumor progression and metastasis. Nat Med.
19:1423–1437. 2013. View
Article : Google Scholar
|
|
6
|
Najafi M, Mortezaee K and Majidpoor J:
Stromal reprogramming: A target for tumor therapy. Life Sci.
239:1170492019. View Article : Google Scholar
|
|
7
|
Rønnov-Jessen L, Petersen OW and Bissell
MJ: Cellular changes involved in conversion of normal to malignant
breast: Importance of the stromal reaction. Physiol Rev. 76:69–125.
1996. View Article : Google Scholar
|
|
8
|
Chen X and Song E: Turning foes to
friends: Targeting cancer-associated fibroblasts. Nat Rev Drug
Discov. 18:99–115. 2019. View Article : Google Scholar
|
|
9
|
Wu H, Ma S, Xiang M and Tong S: HTRA1
promotes transdifferentiation of normal fibroblasts to
cancer-associated fibroblasts through activation of the NF-κB/bFGF
signaling pathway in gastric cancer. Biochem Biophys Res Commun.
514:933–939. 2019. View Article : Google Scholar
|
|
10
|
Affo S, Yu LX and Schwabe RF: The role of
cancer-associated fibroblasts and fibrosis in liver cancer. Annu
Rev Pathol. 12:153–186. 2017. View Article : Google Scholar
|
|
11
|
Öhlund D, Elyada E and Tuveson D:
Fibroblast heterogeneity in the cancer wound. J Exp Med.
211:1503–1523. 2014. View Article : Google Scholar
|
|
12
|
Javeed N and Mukhopadhyay D: Exosomes and
their role in the micro-/macro-environment: A comprehensive review.
J Biomed Res. 31:386–394. 2017.
|
|
13
|
Roy S, Lin HY, Chou CY, Huang CH, Small J,
Sadik N, Ayinon CM, Lansbury E, Cruz L, Yekula A, et al: Navigating
the landscape of tumor extracellular vesicle heterogeneity. Int J
Mol Sci. 20:13492019. View Article : Google Scholar
|
|
14
|
Mathivanan S, Ji H and Simpson RJ:
Exosomes: Extracellular organelles important in intercellular
communication. J Proteomics. 73:1907–1920. 2010. View Article : Google Scholar
|
|
15
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar
|
|
16
|
Lin J, Li J, Huang B, Liu J, Chen X, Chen
XM, Xu YM, Huang LF and Wang XZ: Exosomes: Novel biomarkers for
clinical diagnosis. ScientificWorldJournal. 2015:6570862015.
View Article : Google Scholar
|
|
17
|
Zhang J, Li S, Li L, Li M, Guo C, Yao J
and Mi S: Exosome and exosomal microRNA: Trafficking, sorting, and
function. Genomics Proteomics Bioinformatics. 13:17–24. 2015.
View Article : Google Scholar
|
|
18
|
Yu X, Odenthal M and Fries JW: Exosomes as
miRNA carriers: Formation-function-future. Int J Mol Sci.
17:20282016. View Article : Google Scholar
|
|
19
|
Zhang X, Sai B, Wang F, Wang L, Wang Y,
Zheng L, Li G, Tang J and Xiang J: Hypoxic BMSC-derived exosomal
miRNAs promote metastasis of lung cancer cells via STAT3-induced
EMT. Mol Cancer. 18:402019. View Article : Google Scholar
|
|
20
|
Fang T, Lv H, Lv G, Li T, Wang C, Han Q,
Yu L, Su B, Guo L, Huang S, et al: Tumor-derived exosomal
miR-1247-3p induces cancer-associated fibroblast activation to
foster lung metastasis of liver cancer. Nat Commun. 9:1912018.
View Article : Google Scholar
|
|
21
|
Liu Y, Zhang Y, Wu H, Li Y, Zhang Y, Liu
M, Li X and Tang H: miR-10a suppresses colorectal cancer metastasis
by modulating the epithelial-to-mesenchymal transition and anoikis.
Cell Death Dis. 8:e27392017. View Article : Google Scholar
|
|
22
|
Amin MB, Greene FL, Edge SB, Compton CC,
Gershenwald JE, Brookland RK, Meyer L, Gress DM, Byrd DR and
Winchester DP: The eighth edition AJCC cancer staging manual:
Continuing to build a bridge from a population-based to a more
‘personalized’ approach to cancer staging. CA Cancer J Clin.
67:93–99. 2017. View Article : Google Scholar
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
24
|
Liu Y, Han S, Li Y, Liu Y, Zhang D, Li Y
and Zhang J: MicroRNA-20a contributes to cisplatin-resistance and
migration of OVCAR3 ovarian cancer cell line. Oncol Lett.
14:1780–1786. 2017. View Article : Google Scholar
|
|
25
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006. View Article : Google Scholar
|
|
26
|
desJardins-Park HE, Foster DS and Longaker
MT: Fibroblasts and wound healing: An update. Regen Med.
13:491–495. 2018. View Article : Google Scholar
|
|
27
|
Spaw M, Anant S and Thomas SM: Stromal
contributions to the carcinogenic process. Mol Carcinog.
56:1199–1213. 2017. View
Article : Google Scholar
|
|
28
|
Strutz F, Okada H, Lo CW, Danoff T, Carone
RL, Tomaszewski JE and Neilson EG: Identification and
characterization of a fibroblast marker: FSP1. J Cell Biol.
130:393–405. 1995. View Article : Google Scholar
|
|
29
|
Liu SC, Lu HH, Fan HC, Wang HW, Chen HK,
Lee FP, Yu CJ and Chu YH: The identification of the TRPM8 channel
on primary culture of human nasal epithelial cells and its response
to cooling. Medicine (Baltimore). 96:e76402017. View Article : Google Scholar
|
|
30
|
Ewing J: Neoplastic diseases; a treatise
on tumors PA. WB Saunders; Philadelphia, London: 1928
|
|
31
|
Fidler IJ: Critical factors in the biology
of human cancer metastasis: Twenty-eighth G.H.A. Clowes memorial
award lecture. Cancer Res. 50:6130–6138. 1990.
|
|
32
|
Paget S: The distribution of secondary
growths in cancer of the breast. 1889. Cancer Metastasis Rev.
8:98–101. 1989.
|
|
33
|
Seretis F, Seretis C, Youssef H and
Chapman M: Colorectal cancer: Seed and soil hypothesis revisited.
Anticancer Res. 34:2087–2094. 2014.
|
|
34
|
Chang CH, Fan TC, Yu JC, Liao GS, Lin YC,
Shih AC, Li WH and Yu AL: The prognostic significance of RUNX2 and
miR-10a/10b and their inter-relationship in breast cancer. J Transl
Med. 12:2572014. View Article : Google Scholar
|
|
35
|
Ohuchida K, Mizumoto K, Lin C, Yamaguchi
H, Ohtsuka T, Sato N, Toma H, Nakamura M, Nagai E, Hashizume M and
Tanaka M: MicroRNA-10a is overexpressed in human pancreatic cancer
and involved in its invasiveness partially via suppression of the
HOXA1 gene. Ann Surg Oncol. 19:2394–2402. 2012. View Article : Google Scholar
|
|
36
|
Varnholt H, Drebber U, Schulze F,
Wedemeyer I, Schirmacher P, Dienes HP and Odenthal M: MicroRNA gene
expression profile of hepatitis C virus-associated hepatocellular
carcinoma. Hepatology. 47:1223–1232. 2008. View Article : Google Scholar
|
|
37
|
Yu T, Liu L, Li J, Yan M, Lin H, Liu Y,
Chu D, Tu H, Gu A and Yao M: MiRNA-10a is upregulated in NSCLC and
may promote cancer by targeting PTEN. Oncotarget. 6:30239–30250.
2015. View Article : Google Scholar
|
|
38
|
Bao M, Pan S, Yang W, Chen S, Shan Y and
Shi H: Serum miR-10a-5p and miR-196a-5p as non-invasive biomarkers
in non-small cell lung cancer. Int J Clin Exp Pathol. 11:773–780.
2018.
|
|
39
|
Kalluri R: The biology and function of
fibroblasts in cancer. Nat Rev Cancer. 16:582–598. 2016. View Article : Google Scholar
|
|
40
|
Ara T and Declerck YA: Interleukin-6 in
bone metastasis and cancer progression. Eur J Cancer. 46:1223–1231.
2010. View Article : Google Scholar
|
|
41
|
Lou W, Ni Z, Dyer K, Tweardy DJ and Gao
AC: Interleukin-6 induces prostate cancer cell growth accompanied
by activation of stat3 signaling pathway. Prostate. 42:239–242.
2000. View Article : Google Scholar
|
|
42
|
Tobin RP, Jordan KR, Kapoor P, Spongberg
E, Davis D, Vorwald VM, Couts KL, Gao D, Smith DE, Borgers JSW, et
al: IL-6 and IL-8 Are linked with myeloid-derived suppressor cell
accumulation and correlate with poor clinical outcomes in melanoma
patients. Front Oncol. 9:12232019. View Article : Google Scholar
|
|
43
|
Spiegel A, Brooks MW, Houshyar S,
Reinhardt F, Ardolino M, Fessler E, Chen MB, Krall JA, DeCock J,
Zervantonakis IK, et al: Neutrophils suppress intraluminal NK
cell-mediated tumor cell clearance and enhance extravasation of
disseminated carcinoma cells. Cancer Discov. 6:630–649. 2016.
View Article : Google Scholar
|
|
44
|
Chaffer CL, San Juan BP, Lim E and
Weinberg RA: EMT, cell plasticity and metastasis. Cancer Metastasis
Rev. 35:645–654. 2016. View Article : Google Scholar
|
|
45
|
Castaño Z, San Juan BP, Spiegel A, Pant A,
DeCristo MJ, Laszewski T, Ubellacker JM, Janssen SR, Dongre A,
Reinhardt F, et al: IL-1β inflammatory response driven by primary
breast cancer prevents metastasis-initiating cell colonization. Nat
Cell Biol. 20:1084–1097. 2018. View Article : Google Scholar
|
|
46
|
Eyre R, Alférez DG, Santiago-Gómez A,
Spence K, McConnell JC, Hart C, Simões BM, Lefley D, Tulotta C,
Storer J, et al: Microenvironmental IL1β promotes breast cancer
metastatic colonisation in the bone via activation of Wnt
signalling. Nat Commun. 10:50162019. View Article : Google Scholar
|