Introduction
The incidence of allergic rhinitis (AR) has been
increasing over the past decades in Asian countries (1). In China, the frequency of AR has
increased from 11 to 17% in the past 10 years (2). AR is an immune-related disease that is
characterized as an IgE-modulated type I hypersensitivity disorder
(3). However, type 2 helper T (Th2)
cells have also been proposed to mediate nasal allergic diseases
(4). In addition, IL-13, which is a
cytokine produced by Th2 cells, was verified to a key regulator of
the pathogenesis caused by immune-related inflammation (5). IL-13 can activate eosinophils and
promote the production of mucus and growth factors via the
regulation of epithelial cells (6).
IL-13-treated human nasal epithelial cells (HNECs) are frequently
used as an in vitro model for the study of AR (7). Thus, the modulation of IL-13 is of
great significance for the treatment of AR.
Long non-coding RNAs (lncRNAs) are transcripts of
~200-10,0000 nucleotides in length that are not translated into
proteins (8,9). lncRNAs have been confirmed to serve
important biological roles in numerous biological processes,
including DNA damage, programmed cell death and inflammation
(10–12). Furthermore, lncRNAs have also been
reported to participate in the pathogenesis of AR (13–15).
For example, antisense non-coding RNA in the INK4 locus (ANRIL) was
found to be associated with an increased AR risk and severity, in
addition to an enhanced inflammatory status (16). However, the mechanism by which ANRIL
mediates the progression of AR remains to be determined.
MicroRNAs (miRNAs/miRs) are a class of non-coding
small RNAs that bind to the 3′-untranslated region (3′-UTR) of mRNA
to regulate mRNA expression (17–19).
miRNAs have been proven to be involved in various cellular
processes, including cell growth, differentiation and metabolic
progression (20–24). The dysregulation of miRNAs has also
been confirmed in AR, suggesting the potential function of miRNAs
in AR (25,26). For instance, miR-498 was discovered
to be activated in tissues from patients with AR, and the
suppression of miR-498 promoted inflammatory responses caused by
IL-13 and mucin production in HNECs (15). Moreover, the expression levels of
miR-487b were reported to be upregulated in patients with AR, which
mitigated the pathological alterations of AR by inhibiting the
expression levels of IL-33 and homolog of sulfotransferase
(27). In addition, miR-15a-5p
expression levels were reportedly downregulated in patients with
AR, where served an inhibitory effect in AR (28). However, the association between
ANRIL and miR-15a-5p in AR remains unclear.
JAK2/STAT3 signaling was found to function as a
crucial mediator in the overactivation of macrophages, which caused
an increase in proinflammatory cytokine production (29). In addition, it was reported that
miR-375 alleviated the progression of AR via regulation of the
JAK2/STAT3 axis (30). Based on
these findings, it was hypothesized that JAK2/STAT3 signaling may
serve an important role in the progression of AR.
The present study aimed to confirm the role of ANRIL
in AR. In addition, the study aimed to further determine the
mechanism by which ANRIL mediated the progression of AR. The
results of the current study may provide a novel strategy for the
treatment of AR.
Materials and methods
Cell culture and treatment
Primary HNECs (cat. no. HUM-iCell-m018) and 293T
cell lines (cat. no. ACS-4500) were purchased from iCell
Bioscience, Inc., and the American Type Culture Collection,
respectively. All cells were cultured in RPMI-1640 medium (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco;
Thermo Fischer Scientific, Inc.), 1% penicillin (Invitrogen; Thermo
Fisher Scientific, Inc.) and 1% streptomycin (Invitrogen; Thermo
Fischer Scientific, Inc.), and maintained at 37°C and 5%
CO2. To establish an in vitro model of AR, HNECs
were treated with 50 ng/ml IL-13 (Sigma-Aldrich; Merck KGaA) at
37°C for 24 h, as previously described (31). The control group consisted of
untreated cells.
Cell transfection
Cell transfection was performed according to a
previously reported experimental method (32). The miR-15a-5p inhibitor, miR-15a-5p
mimic and their respective negative controls (NCs) were purchased
from Shanghai GenePharma Co., Ltd. The sequences were as follows:
miR-15a-5p mimics sense, 5′-UAGCAGCACAUAAUGGUUUGUG-3′ and
antisense, 5′-CAAACCAUUAUGUGCUGCUAUU-3′; mimic NC sense,
5′-UUCUCCGAACGUGUCACGUTT-3′ and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′; miR-15a-5p inhibitor,
5′-CACAAACCAUUAUGUGCUGCUA-3′; and inhibitor NC, 5′-
CAGUACUUUUGUGUAGUACAA-3′. For the genetic knockdown of ANRIL, the
corresponding short hairpin RNA (shRNA/sh) targeting ANRIL
(sh-ANRIL) and the non-targeting control shRNA (sh-NC) were cloned
into the pSicoR vector (Addgene, Inc.). pcDNA3.1-JAK2 and the
corresponding control (empty pcDNA3.1 vector) were obtained from
GenScript. HNECs (5×103 per well) were transfected using
Lipofectamine® 2000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.). Briefly, the cells were cultured in
antibiotic-free medium (RPMI-1640) at 37°C for 24 h, and
transfections were performed upon the cell confluence reaching 90%.
The successfully constructed vectors, miR-15a-5p inhibitor/mimics
or controls, and Lipofectamine 2000 were diluted (1:100) in
serum-free Opti-MEM medium (Gibco; Thermo Fisher Scientific, Inc.),
mixed after standing for 5 min, and then incubated for 20 min at
room temperature. Then, 100 µl of the mixed solution was added to
the cells in each well and incubated at 37°C for 48 h. The medium
was replaced with fresh medium after 6 h. Reverse
transcription-quantitative PCR (RT-qPCR) was used to confirm the
transfection efficiency 48 h after transfection.
RT-qPCR
RT-qPCR analysis was used to analyze mRNA expression
levels, as previously described (33). Briefly, total RNA was extracted from
HNECs using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. The
quality and quantity of RNA was detected using 10% agarose gel
electrophoresis. In addition, A260/A280 ratios were calculated
using a spectrophotometer (NanoDrop Technologies; Thermo Fisher
Scientific, Inc.), and a ratio between 1.8 and 2.0 indicated that
the extracted RNA was qualified. Total RNA was reverse transcribed
into cDNA using the PrimeScript RT reagent kit (Takara Bio, Inc.),
according to the manufacturer's protocol. qPCR was subsequently
performed using the SYBR Premix Ex Taq II kit (Takara Bio, Inc.).
The following thermocycling conditions were used for the qPCR:
Initial denaturation for 1 min at 95°C; followed by 35 cycles for
15 sec at 95°C and 30 sec at 60°C. The primer pairs used are
presented in Table I. The
expression levels were quantified using the 2−∆∆Cq
method (34). GAPDH or U6 were used
as the internal loading control for quantification. Each experiment
was performed in triplicate.
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| Antisense
non-coding RNA in the INK4 locus | F:
GCCGGACTAGTGTCCCTTTT |
|
| R:
TCGGGAAAGGATTCCAGCAC |
|
MicroRNA-15a-5p | F:
CGGCTAGCAGCACATAATGG |
|
| R:
GTCGTATCCAGTGCAGGGTCCGAGG |
|
|
TATTCGCACTGGATACGACCACAAA |
| JAK2 | F:
CCAAAGTGGGCAGAATTAGC |
|
| R:
GTGTAGGATCCCGGTCTTCA |
|
Granulocyte-macrophage colony-stimulating
factor | F:
GAGAGCTCCCAGCCTAAGGT |
|
| R:
TGCACACCTCTTGACACTCC |
| Eotaxin-1 | F:
CGGATACCTTGGCGCTAGTA |
|
| R:
TCACTCCGTCTTTTGCACAG |
| Mucin 5AC | F:
AGCCCAAGATGCCCTTCAGT |
|
| R:
CCGTGTTCCTACCCCCAATG |
| U6 | F:
GGTCGGGCAGGAAAGAGGGC |
|
| R:
GCTAATCTTCTCTGTATCGTTCC |
| GAPDH | F:
CTGACTTCAACAGCGACACC |
|
| R:
GTGGTCCAGGGGTCTTACTC |
Western blotting
Protein expression levels were analyzed using
western blotting, according to a previous study (35). Briefly, total protein was extracted
from HNECs using RIPA lysis buffer [50 mM Tris (pH 7.4), 150 mM
NaCl, 1% Triton X-100, 1% sodium deoxycholate, 0.1% SDS, 2 mM
sodium pyrophosphate, 25 mM β-glycerophosphate, 1 mM EDTA, 1 mM
Na3VO4 and 0.5 µg/ml leupeptin] (Beyotime
Institute of Biotechnology). Total protein was quantified using a
bicinchoninic acid protein kit (Pierce; Thermo Fisher Scientific,
Inc.) on a Multiskan™ FC microplate photometer (562 nm; Thermo
Fisher Scientific, Inc.) and 40 µg protein/lane was separated via
10% SDS-PAGE. The separated proteins were subsequently transferred
onto PVDF membranes (Invitrogen; Thermo Fisher Scientific, Inc.)
and blocked with 5% skimmed milk in TBS with 10% Tween 20 for 1 h
at room temperature. The membranes were then incubated with the
following primary antibodies overnight at 4°C (all from Abcam):
Anti-JAK2 (1:1,000; cat. no. ab108596), anti-STAT3 (1:2,000; cat.
no. ab68153), anti-phosphorylated (p)-STAT3 (1:2,000; cat. no.
ab76315) and anti-GAPDH (1:1,000; cat. no. ab9485). Following the
primary antibody incubation, the PVDF membranes were washed with
TBS with Tween-20 and incubated with a HRP-conjugated secondary
antibody (1:5,000; cat. no. ab205718; Abcam) at room temperature
for 1 h. Finally, the protein bands were visualized using an ECL
detection kit (Beyotime Institute of Biotechnology). The relative
protein expression levels were normalized to GAPDH expression
levels. Image-Pro Plus 6.0 (National Institutes of Health) was used
for the densitometry analysis.
Dual luciferase reporter assay
miR-15a-5p binding sites were identified by StarBase
database (http://starbase.sysu.edu.cn). The
dual luciferase reporter assay was performed according to a
previous study (36). ANRIL and
JAK2 3′-UTRs containing the putative binding sites (GCUGCU) of
miR-15a-5p were synthetized and obtained from Sangon Biotech Co.,
Ltd. The aforementioned sequences were cloned into the pmirGLO
vector (Promega Corporation) to construct wild-type (WT) or mutant
(MUT) ANRIL (10 nM) and JAK2 reporter vectors (10 nM). Point
mutations of the miR-15a-5p binding sites were generated using a
Site-Directed Mutagenesis kit (Promega Corporation). The WT or MUT
ANRIL/JAK2 vectors (10 nM) were co-transfected into HNECs
(5×106 per well) with mimics NC or miR-15a-5p mimics (10
nM) using Lipofectamine 2000 reagent for 24 h at 37°C, according to
the manufacturer's instructions. After 24 h, relative luciferase
activities were detected using a Dual-GLO Luciferase assay system
(Promega Corporation). Firefly luciferase activity was normalized
to Renilla luciferase activity.
ELISA
The secretory levels of eotaxin-1 [cat. no.
70-EK1130-96; Hangzhou Multi Sciences (Lianke) Biotech Co., Ltd.),
granulocyte-macrophage colony-stimulating factor [(GM-CSF); cat.
no. 70-EK163HS-96; Hangzhou Multi Sciences (Lianke) Biotech Co.,
Ltd.] and mucin 5AC [(MUC5AC); cat. no. ml-1016017; Shanghai
Enzyme-linked Biotechnology Co., Ltd.] in the supernatant of HNECs
were analyzed using ELISA kits. The supernatants of HNECs were
harvested by centrifugation (100 × g, 20 min, 4°C). Subsequently,
the cells were incubated with a HRP-conjugated secondary antibody,
which was included in the ELISA kits, at room temperature for 1 h.
Finally, after incubation with hydrochloric acid (100 µl) at room
temperature until the solution became discolored, the absorbance
was measured using a microplate reader (450 nm).
Statistical analysis
Statistical analysis was performed using SPSS 22.0
software (IBM Corp.). Data are presented as the mean ± SD and three
technical repeats were performed in three biological replicate
experiments. Statistical differences between 2 groups were analyzed
using an unpaired Student's t-test, while ≥3 groups were analyzed
using a one-way ANOVA followed by a Tukey's post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
ANRIL and JAK2 expression levels are
upregulated in IL-13-induced HNECs, while miR-15a-5p expression
levels are downregulated
To establish an in vitro model of AR, HNECs
were treated with IL-13 for 24 h. As indicated in Fig. 1A, the secretory levels of eotaxin-1,
GM-CSF and MUC5AC in HENC supernatants were significantly
upregulated by IL-13 stimulation. These data suggested that an
in vitro model of AR was successfully established. In
addition, the mRNA expression levels of ANRIL and JAK2 in HNECs
were significantly upregulated in the presence of IL-13 compared
with the control group (Fig. 1B).
In contrast, miR-15a-5p expression levels in HNECs were
significantly downregulated following IL-13 stimulation compared
with the control group (Fig. 1B).
Furthermore, the protein expression levels of JAK2 and
p-STAT3/STAT3 in HNECs were significantly upregulated by IL-13
stimulation compared with the control group (Fig. 1C and D). These data indicated that
ANRIL and JAK2 may promote the occurrence of AR, while miR-15a-5p
exhibited the opposite effects.
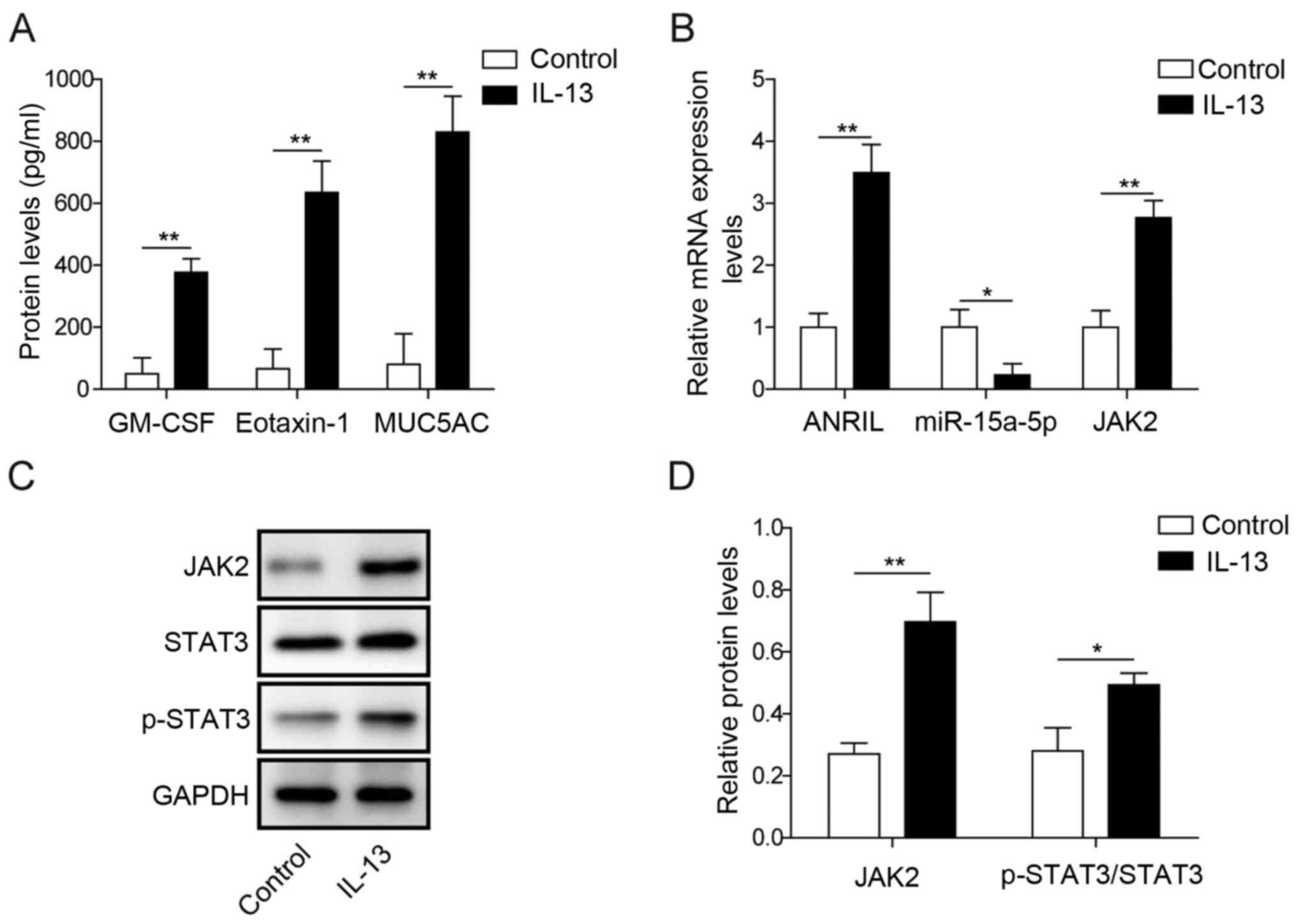 | Figure 1.ANRIL and JAK2 expression levels are
upregulated in IL-13-induced HNECs, while miR-15a-5p expression
levels are downregulated. HNECs were treated with 50 ng/ml IL-13
for 24 h. (A) Secretory levels of eotaxin-1, GM-CSF and MUC5AC in
HNEC supernatants were analyzed using ELISAs. (B) Relative mRNA
expression levels of ANRIL, miR-15a-5p and JAK2 in HNECs were
analyzed using reverse transcription-quantitative PCR. (C) Protein
expression levels of JAK2, STAT3 and p-STAT3 in HNECs were analyzed
using western blotting. (D) Relative protein expression levels from
part (C) were semi-quantified via normalization to GAPDH expression
levels. *P<0.05, **P<0.01. ANRIL, antisense non-coding RNA in
the INK4 locus; HNECS, human nasal epithelial cells; GM-CSF,
granulocyte-macrophage colony-stimulating factor; MUC5AC, mucin
5AC; miR, microRNA; p-, phosphorylated. |
ANRIL sponges miR-15a-5p in HNECs
To confirm the transfection efficiency of sh-ANRIL
in HNECs, RT-qPCR analysis was performed. As shown in Fig. 2A, the expression levels of ANRIL in
HNECs were significantly downregulated following transfection with
sh-ANRIL. These results confirmed that sh-ANRIL was stably
transfected into HNECs. The genetic silencing of ANRIL
significantly upregulated the expression levels of miR-15a-5p in
HNECs (Fig. 2B). These data
suggested that ANRIL may negatively regulate miR-15a-5p expression
levels. The expression levels of miR-15a-5p in HNECs were
significantly upregulated following the transfection with the
miR-15a-5p mimics and downregulated in the presence of the
miR-15a-5p inhibitor compared with their respective NC groups
(Fig. 2C). Furthermore, to identify
the downstream target gene of ANRIL, the StarBase database and dual
luciferase reporter assays were used. The StarBase database
predicted that ANRIL bound to miR-15a-5p (Fig. 2D). Subsequent dual luciferase
reporter assays revealed that the co-transfection with miR-15a-5p
mimics significantly suppressed the relative luciferase activity of
ANRIL-WT compared with cells co-transfected with the ANRIL-WT
vector and mimics NC, while no significant differences were
observed in the relative luciferase activity of ANRIL-MUT between
the cells co-transfected with the mimics NC or miR-15a-5p mimics
(Fig. 2D). These findings suggested
that ANRIL may bind to miR-15a-5p in HNECs.
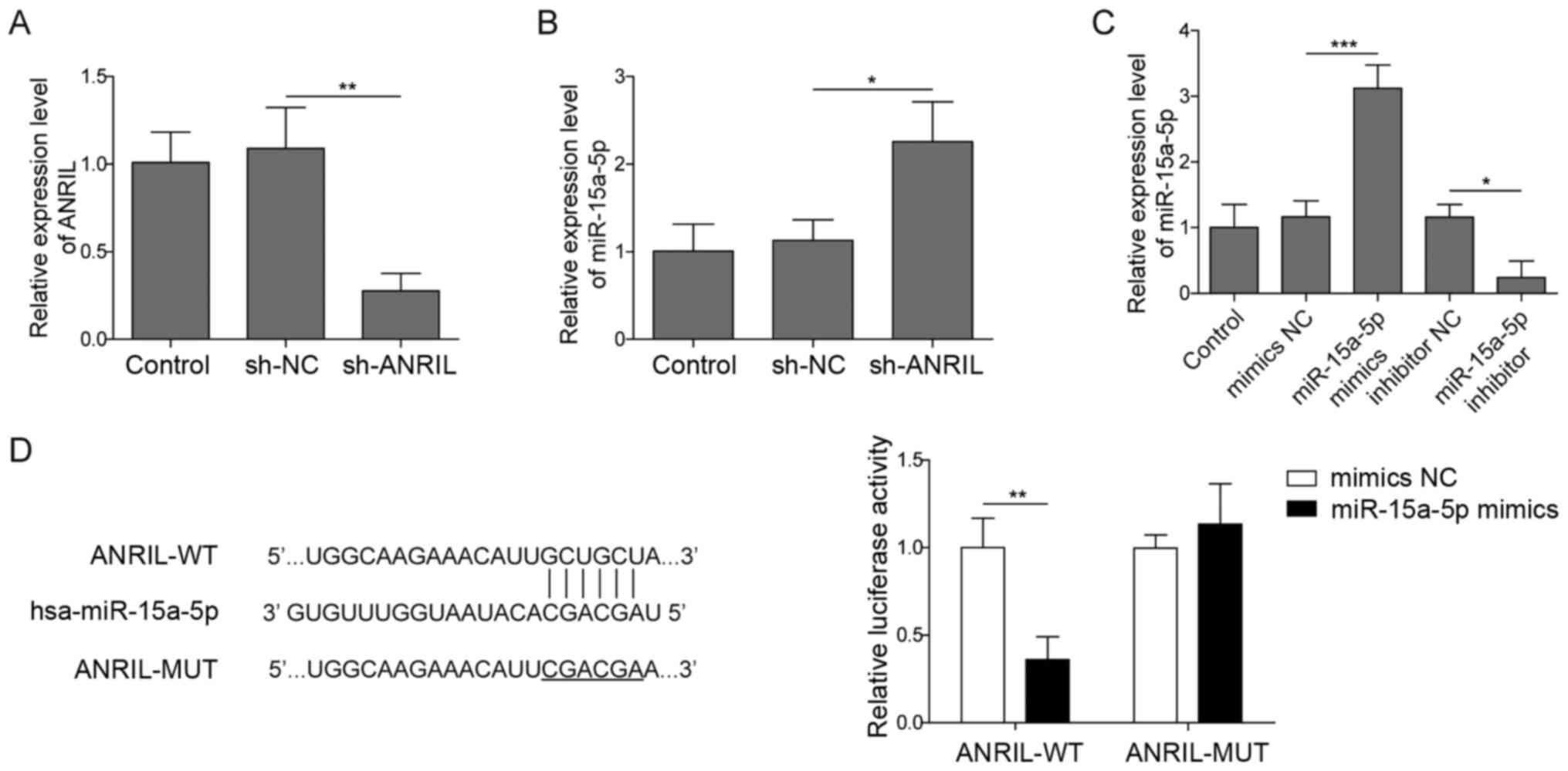 | Figure 2.ANRIL binds to miR-15a-5p in HNECs.
HNECs were transfected with sh-ANRIL or sh-NC for 24 h. (A)
Transfection efficiency was confirmed using RT-qPCR. (B) miR-15a-5p
expression levels in HNECs were analyzed using RT-qPCR. (C) HNECs
were transfected with mimics/inhibitor NC or miR-15a-5p
mimics/inhibitor for 24 h. Then, the transfection efficiency was
determined using RT-qPCR. (D) Complementary binding between ANRIL
and miR-15a-5p was predicted by StarBase. The relative luciferase
activity was analyzed following the co-transfection with mimics NC
or miR-15a-5p mimics and luciferase reporter plasmids carrying the
ANRIL-WT or MUT 3′-untrnaslated region in HNECs using a dual
luciferase reporter assay. *P<0.05, **P<0.01, ***P<0.001.
ANRIL, antisense non-coding RNA in the INK4 locus; HNECS, human
nasal epithelial cells; sh, short hairpin RNA; NC, negative
control; RT-qPCR, reverse transcription-quantitative PCR; miR,
microRNA; WT, wild-type; MUT, mutant. |
ANRIL regulates the production of
inflammatory cytokines and mucin via inhibition of miR-15a-5p
Since eotaxin-1, GM-CSF and MUC5AC have been
discovered to serve promoting roles in AR (15,37),
the effect of ANRIL on these cytokines and mucin was investigated.
As shown in Fig. 3A-C, the mRNA
expression levels of GM-CSF, eotaxin-1 and MUC5AC in HNECs were
significantly upregulated by IL-13 stimulation; however, the
expression levels of the aforementioned genes were significantly
reversed in the presence of sh-ANRIL. Conversely, co-transfection
with the miR-15a-5p inhibitor partially reversed the effect of
ANRIL knockdown on the mRNA expression levels of these cytokines
and mucin. Similar results were obtained regarding the secretory
levels of eotaxin-1, GM-CSF and MUC5AC using ELISAs; the
IL-13-induced increase in eotaxin-1, GM-CSF and MUC5AC
concentrations in HNECs were significantly inhibited by silencing
ANRIL, while the inhibitory effect of ANRIL silencing was partially
rescued following the co-transfection with the miR-15a-5p inhibitor
(Fig. 3D-F). Altogether, these
results suggested that the miR-15a-5p inhibitor may significantly
reduce the anti-inflammatory effect of ANRIL knockdown on
IL-13-induced AR in vitro.
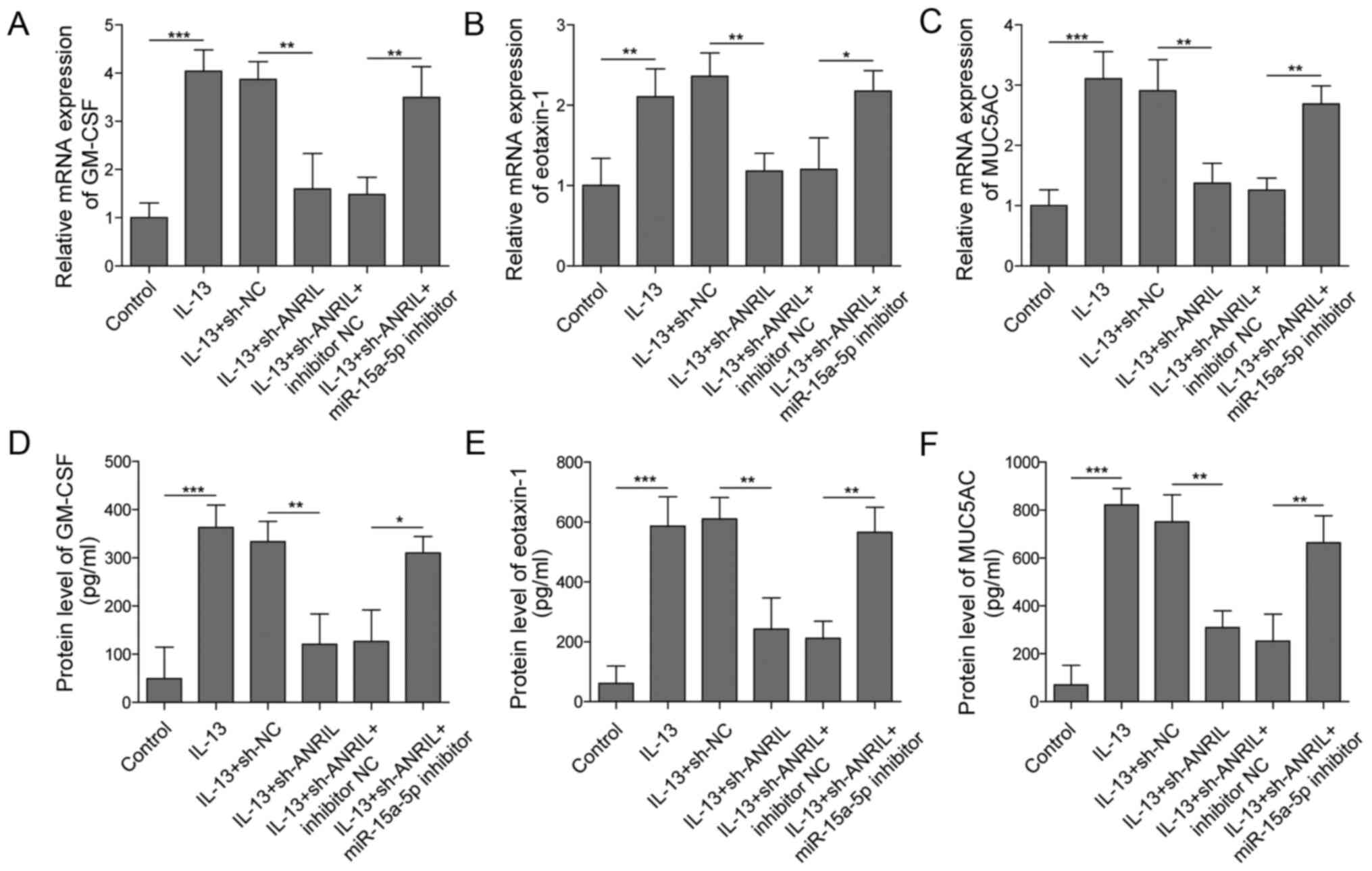 | Figure 3.ANRIL regulates the production of
inflammatory cytokines and mucin by sponging miR-15a-5p. HNECs were
treated with IL-13, IL-13 + sh-NC, IL-13 + sh-ANRIL, IL-13 +
sh-ANRIL+ inhibitor NC or IL-13 + sh-ANRIL + miR-15a-5p inhibitor.
mRNA expression levels of (A) GM-CSF, (B) eotaxin-1 and (C) MUC5AC
in HNECs were analyzed using reverse transcription-quantitative
PCR. Secretory levels of (D) GM-CSF, (E) eotaxin-1 and (F) MUC5AC
in the supernatants of HNECs were detected using ELISAs.
*P<0.05, **P<0.01, ***P<0.001. ANRIL, antisense non-coding
RNA in the INK4 locus; HNECS, human nasal epithelial cells; miR,
microRNA; sh, short hairpin RNA; NC, negative control; GM-CSF,
granulocyte-macrophage colony-stimulating factor; MUC5AC, mucin
5AC. |
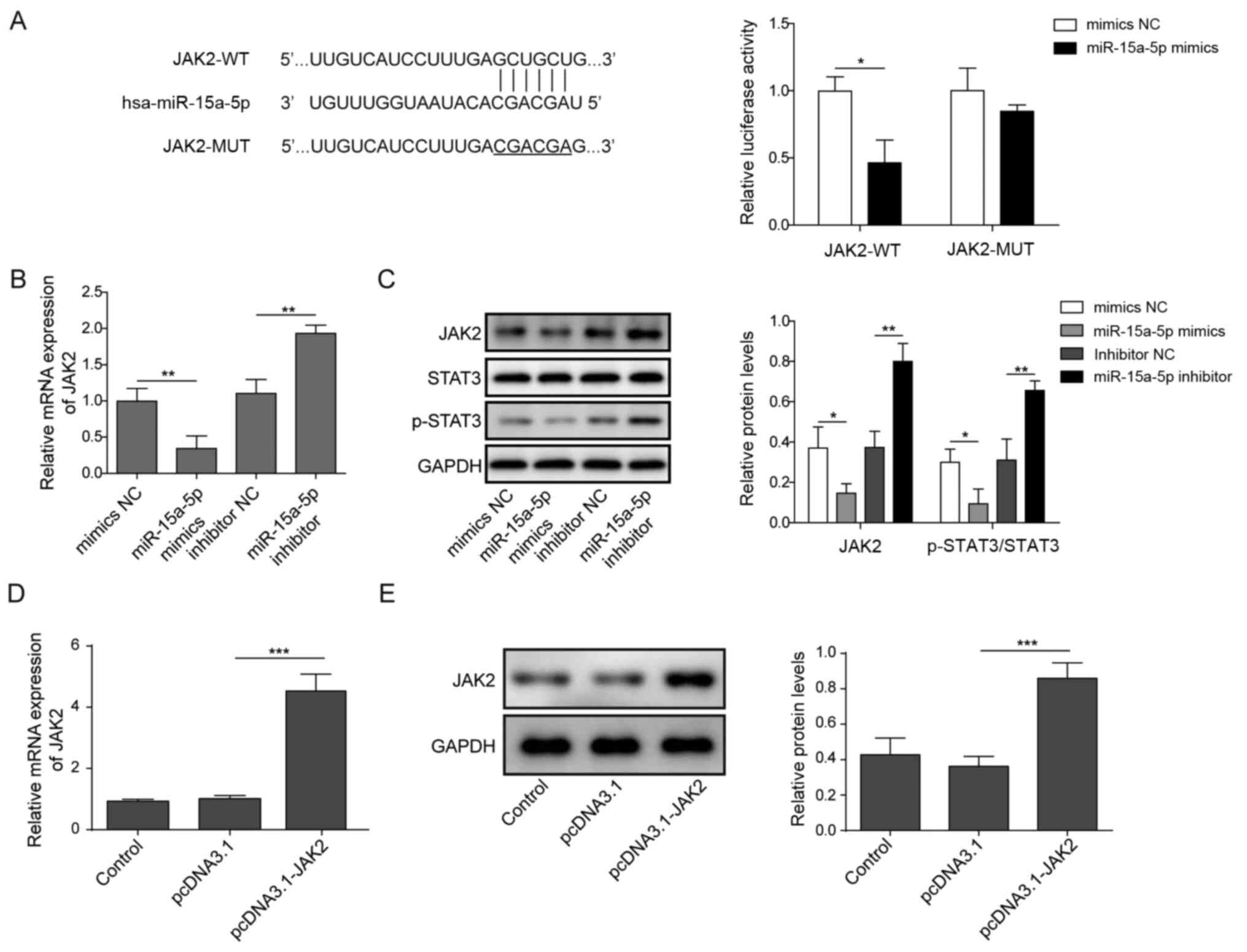
miR-15a-5p inactivates JAK2/STAT3
signaling by directly targeting JAK2
To determine the direct target of miR-15a-5p, the
StarBase database and dual luciferase reporter assays were used.
The analysis indicated that JAK2 might be a direct target of
miR-15a-5p (Fig. 4A). Furthermore,
the co-transfection with the miR-15a-5p mimics significantly
decreased the relative luciferase activity of the JAK2-WT vector
compared with the co-transfection with the mimics NC, but no
significant differences were observed in the relative luciferase
activity of the JAK2-MUT between the mimics NC and miR-15a-5p
mimics group (Fig. 4A). Moreover,
the expression levels of JAK2 in HNECs were significantly
downregulated following the transfection with the miR-15a-5p mimics
compared with the mimics NC group (Fig.
4B). In contrast, the miR-15a-5p inhibitor exhibited the
opposite effect (Fig. 4B). The
protein expression levels of JAK2 and p-STAT3/STAT3 in HNECs were
also significantly downregulated in the miR-15a-5p mimics group
compared with the mimics NC group, while the expression levels were
significantly upregulated by the miR-15a-5p inhibitor compared with
the inhibitor NC (Fig. 4C).
Meanwhile, to investigate the association between miR-15a-5p and
JAK2, HNECs were treated with pcDNA3.1-JAK2 overexpression vector.
The data confirmed that the mRNA and protein expression levels of
JAK2 were significantly upregulated in HNECs transfected with
pcDNA3.1-JAK2 (Fig. 4D and E).
These findings suggested that miR-15a-5p may directly target JAK2
to inhibit the STAT3 signaling pathway.
miR-15a-5p mimics significantly
reverse IL-13-induced inflammatory responses in HNECs by targeting
JAK2
To further confirm the mechanism by which miR-15a-5p
mediated the development of AR in vitro, RT-qPCR was used.
As expected, the overexpression of miR-15a-5p significantly
downregulated the mRNA expression levels of eotaxin-1, GM-CSF and
MUC5AC in IL-13-treated HNECs (Fig.
5A-C). In contrast, the inhibitory effect of miR-15a-5p mimics
was significantly reversed following JAK2 overexpression (Fig. 5A-C). Similarly, the
anti-inflammatory effect of miR-15a-5p on the secretory levels of
these proteins was significantly reversed by JAK2 overexpression
(Fig. 5D-F). All these data
indicated that the upregulation of JAK2 may rescue the
anti-inflammatory effect of miR-15a-5p mimics on AR.
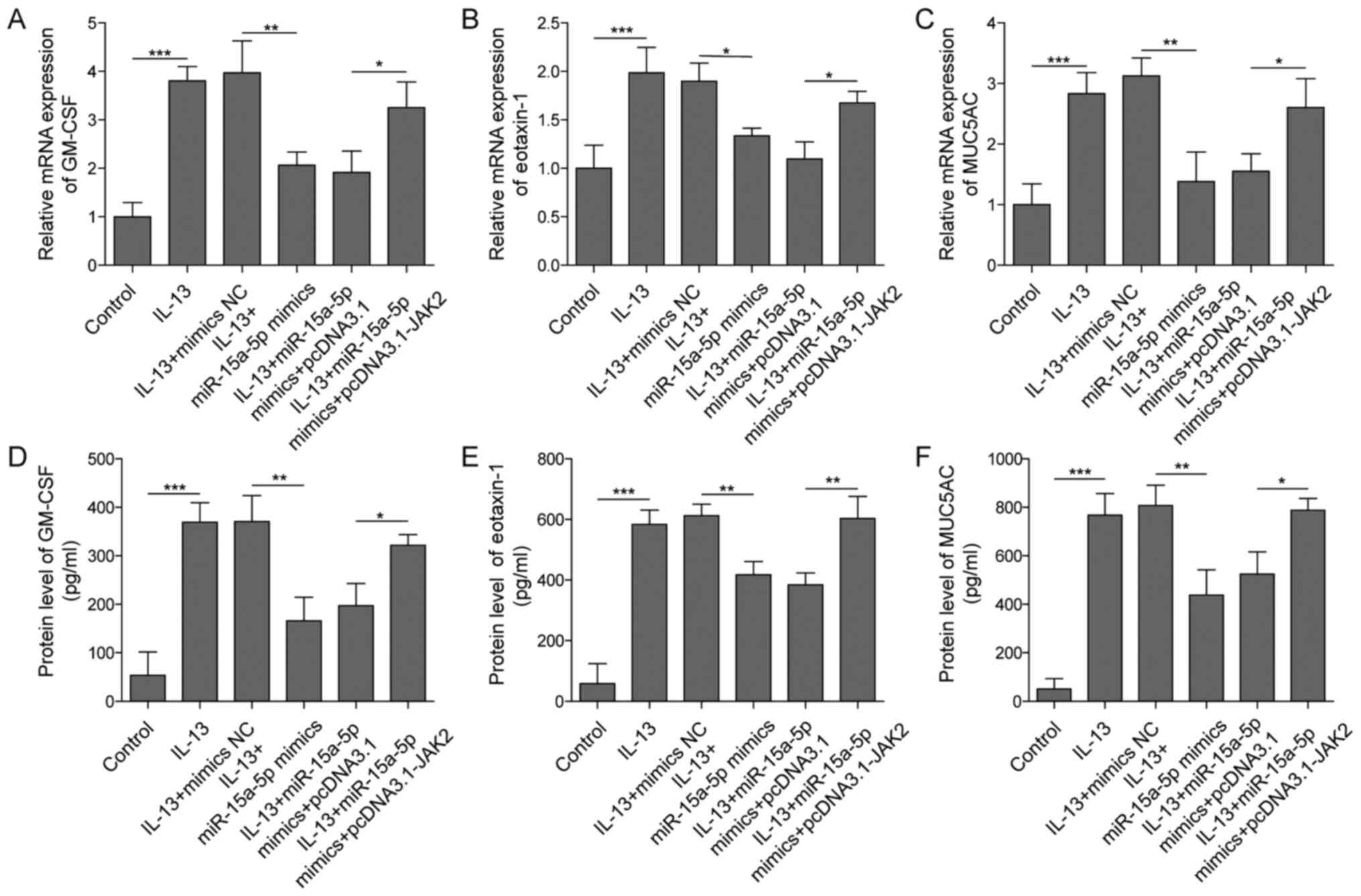 | Figure 5.miR-15a-5p mimics significantly
reverse the IL-13-induced inflammation in HNECs by targeting JAK2.
HNECs were treated with IL-13, IL-13+mimics-NC, IL-13+ miR-15a-5p
mimics, IL-13+miR-15a-5p mimics+pcDNA3.1 or IL-13+miR-15a-5p
mimics+pcDNA3.1-JAK2. The mRNA expression levels of (A) GM-CSF, (B)
eotaxin-1 and (C) MUC5AC in HNECs were analyzed using reverse
transcription-quantitative PCR. The secretory levels of (D) GM-CSF,
(E) eotaxin-1 and (F) MUC5AC in the supernatants of HNECs were
analyzed using ELISA kits. *P<0.05, **P<0.01, ***P<0.001.
HNECS, human nasal epithelial cells; miR, microRNA; NC, negative
control; GM-CSF, granulocyte-macrophage colony-stimulating factor;
MUC5AC, mucin 5AC. |
Discussion
Although increasing efforts have been made to
improve the treatment of AR, the disease remains difficult to
manage and significantly affects the patients' quality of life
(38). Hence, it is necessary to
discover novel methods for AR treatment. In the present study,
HNECs were treated with IL-13 to study AR in vitro. The results
revealed that the production of GM-CSF, eotaxin-1 and MUC5AC was
significantly increased following IL-13 treatment, confirming that
the in vitro model of AR had been successfully established.
GM-CSF, eotaxin-1 and MUC5AC are known as key mediators in AR due
to the fact that upregulation of these three cytokines is
associated with the progression of AR (15,32).
Thus, the present findings suggested that ANRIL could mediate the
progression of AR via regulation of GM-CSF, eotaxin-1 and
MUC5AC.
Previous studies have revealed that lncRNAs
participate in AR. For example, lncRNA growth arrest specific 5 was
found to inhibit the progression of AR via regulation of immune
responses (14). Another previous
study indicated that linc00632 suppressed the inflammatory cytokine
effects caused by IL-13 and mucin production in neuroepithelial
cells (15). In the current study,
ANRIL was found to be activated in IL-13-treated HNECs. This
discrepancy may result from the different functions of lncRNAs on
IL-13-induced inflammatory responses. Qian et al (16) reported that ANRIL promoted AR via
the regulation of inflammatory cytokines (TNF-α, IL-4, IL-6, IL-13,
IL-10 and IL-17). The data of the present study were consistent
with this previous finding, verifying that ANRIL knockdown could
inhibit the occurrence of AR in vitro by downregulating the
production of inflammatory cytokines and mucus. Of note, Zhou et
al (39) discovered that
silencing ANRIL could sponge miR-125a-5p to inhibit the progression
of Alzheimer's disease. However, the findings of the present study
revealed that ANRIL could bind to miR-15a-5p in AR. Therefore,
ANRIL may play a role in different types of diseases by regulating
different miRNAs.
miRNAs are common negative regulators of gene
expression. According to the competitive endogenous RNA (ceRNA)
mechanism, endogenous lncRNAs with miRNA target sites have the
potential to act as natural miRNA sponges, which suppress the
expression of miRNA targets by competitively binding and inhibiting
miRNAs (40). Notably, some lncRNAs
(PVT1 and FENDRR) have been identified to regulate miR-15a-5p
expression via the ceRNA network (41,42).
The current data identified potential crosstalk between ANRIL and
miR-15a-5p, suggesting that ANRIL knockdown inhibited the
progression of AR by preventing the sponging of miR-15a-5p.
Additionally, it has been previously reported that miR-15a-5p could
promote the progression of sepsis by regulating the inflammatory
response in macrophages and targeting TNFAIP3-interacting protein 2
(43). In the present research, the
overexpression of miR-15a-5p could inhibit the development of AR
in vitro by suppressing the secretion of GM-CSF, eotaxin-1
and MUC5AC. Thus, these findings indicated that miR-15a-5p may
serve as a suppressor during inflammation. Wang et al
(28) reported that miR-15a-5p
inhibited IL-13-induced expression of GM-CSF, eotaxin-1 and MUC5AC
in HNECs to alleviate AR via negatively regulating adrenoceptor β2
(ADRB2). The findings of the current study were consistent with
this previous study, indicating that miR-15a-5p may serve as a
suppressor in AR.
In addition, JAK2 was discovered to be directly
targeted by miR-15a-5p. JAK2/STAT3 is an important signaling
pathway involved in multiple inflammatory responses (44,45).
For example, JAK2/STAT3 was reported to promote the inflammatory
responses induced by lipopolysaccharide in human umbilical vein
endothelial cells (46). Moreover,
JAK2/STAT3 has been proven to be a key mediator of Th2-mediated
immune responses (47). Paroxetine
was identified to exert its immunosuppressive effects on immune
responses by activating the JAK2/STAT3 signaling pathway (48), suggesting that the JAK2/STAT3
signaling pathway has immunosuppressive effect, which is the
opposite function found in other previous studies. This phenomenon
may due to the different type of diseases. In the present study,
the overexpression of JAK2 significantly suppressed the
anti-inflammatory effects of miR-15a-5p in AR. Wang et al
(30) reported that miR-375 could
ameliorate AR via the downregulation of the JAK2/STAT3 signaling
pathway. Accordingly, the present findings suggested that
miR-15a-5p may exert its anti-inflammatory effect in IL-13-treated
HNECs by targeting JAK2 and inactivating JAK2/STAT3 signaling.
However, it should be noted that this study only focused on
JAK2/STAT3 signaling. Since IL-4 secreted by Th2 cells has also
been reported to be involved in AR (15), the effect of ANRIL on IL-4
production should be further investigated in the future.
In conclusion, the findings of the present study
suggested that ANRIL may function as a mediator in AR. ANRIL
knockdown was demonstrated to exert its anti-inflammatory effect in
IL-13-treated HNECs via regulation of the miR-15a-5p/JAK2/STAT3
axis. These results suggested that ANRIL may serve as a novel
target for the treatment of AR.
Acknowledgements
Not applicable.
Funding
This work is supported by the Regional Fund of
National Natural Science Foundation of China (grant no.
81860147).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
H-WL designed the study, performed the experiments,
prepared the manuscript and acted as the guarantor of the integrity
of the entire study. Y-QZ designed the study, analyzed the data and
reviewed the manuscript. Z-LH analyzed the data and reviewed the
manuscript. HL, Q-FT and JT performed the experiments. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
These authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AR
|
allergic rhinitis
|
|
HNECs
|
human nasal epithelial cells
|
|
Th2
|
type 2 helper T cells
|
|
ANRIL
|
antisense noncoding RNA in the INK4
locus
|
|
GM-CSF
|
granulocyte-macrophage
colony-stimulating factor
|
|
MUC5AC
|
mucin 5AC
|
References
|
1
|
Greiner AN, Hellings PW, Rotiroti G and
Scadding GK: Allergic rhinitis. Lancet. 378:2112–2122. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li L, Zhang L, Mo JH, Li YY, Xia JY, Bai
XB, Xie PF, Liang JY, Yang ZF and Chen QY: Efficacy of indoor air
purification in the treatment of Artemisia pollen-allergic
rhinitis: A randomised, double-blind, clinical controlled trial.
Clin Otolaryngol. 45:394–401. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pazdro-Zastawny K, Kolator M, Krajewska J,
Basiak-Rasała A, Górna S, Paluszkiewicz P, Zatoński M and Zatoński
T: Lifestyle-related factors differentiating the prevalence of
otorhinolaryngological diseases among 6–17-year-olds from Wrocław,
Poland. Int J Pediatr Otorhinolaryngol. 132:1099342020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Licari A, Castagnoli R, De Filippo M,
Foiadelli T, Tosca MA, Marseglia GL and Ciprandi G: Current and
emerging biologic therapies for allergic rhinitis and chronic
rhinosinusitis. Expert Opin Biol Ther. 20:609–619. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu J, Zhang X, Zhao Y and Wang Y: The
association between allergic rhinitis and sleep: A systematic
review and meta-analysis of observational studies. PLoS One.
15:e02285332020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liang ZY, Deng YQ and Tao ZZ: A quantum
dot-based lateral flow immunoassay for the rapid, quantitative, and
sensitive detection of specific IgE for mite allergens in sera from
patients with allergic rhinitis. Anal Bioanal Chem. 412:1785–1794.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kenyon CC, Maltenfort MG, Hubbard RA,
Schinasi LH, De Roos AJ, Henrickson SE, Bryant-Stephens TC and
Forrest CB: Variability in diagnosed asthma in young children in a
large pediatric primary care network. Acad Pediatr. 20:958–966.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Choi HS, Kim SL, Kim JH and Lee DS: The
FDA-approved anti-asthma medicine ciclesonide inhibits lung cancer
stem cells through Hedgehog signaling-mediated SOX2 regulation. Int
J Mol Sci. 21:10142020. View Article : Google Scholar
|
|
9
|
Yi L, Cui Y, Xu Q and Jiang Y:
Stabilization of LSD1 by deubiquitinating enzyme USP7 promotes
glioblastoma cell tumorigenesis and metastasis through suppression
of the p53 signaling pathway. Oncol Rep. 36:2935–2945. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qin X, Zhu S, Chen Y, Chen D, Tu W and Zou
H: Long non-coding RNA (lncRNA) CASC15 is upregulated in
diabetes-induced chronic renal failure and regulates podocyte
apoptosis. Med Sci Monit. 26:e9194152020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao J, Pu J, Hao B, Huang L, Chen J, Hong
W, Zhou Y, Li B and Ran P: LncRNA RP11-86H7.1 promotes airway
inflammation induced by TRAPM2.5 by acting as a ceRNA of miRNA-9-5p
to regulate NFKB1 in HBECS. Sci Rep. 10:115872020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu X, Zhu N, Zhang B and Xu SB: Long
Noncoding RNA TCONS_00016406 attenuates lipopolysaccharide-induced
acute kidney injury by regulating the miR-687/PTEN pathway. Front
Physiol. 11:6222020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang Y, Zhang Y, Yang Y, Guo J, Yang L, Li
C and Song X: Differential expression of long noncoding RNAs and
their function-related mRNAs in the peripheral blood of allergic
rhinitis patients. Am J Rhinol Allergy. 34:508–518. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu X, Wang X, Wang Y and Zhao Y: Exosomal
long non-coding RNA GAS5 suppresses Th1 differentiation and
promotes Th2 differentiation via downregulating EZH2 and T-bet in
allergic rhinitis. Mol Immunol. 118:30–39. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yue L, Yin X, Hao F, Dong J, Ren X, Xu O
and Shan C: Long noncoding RNA linc00632 inhibits
interleukin-13-induced inflammatory cytokine and mucus production
in nasal epithelial cells. J Innate Immun. 12:116–128. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qian X, Shi S and Zhang G: Long non-coding
RNA antisense non-coding RNA in the INK4 locus expression
correlates with increased disease risk, severity, and inflammation
of allergic rhinitis. Medicine (Baltimore). 98:e152472019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Li J, Xu X, Wei C, Liu L and Wang T: Long
noncoding RNA NORAD regulates lung cancer cell proliferation,
apoptosis, migration, and invasion by the miR-30a-5p/ADAM19 axis.
Int J Clin Exp Pathol. 13:1–13. 2020.PubMed/NCBI
|
|
18
|
Yan Y, Peng Y, Ou Y and Jiang Y:
MicroRNA-610 is downregulated in glioma cells, and inhibits
proliferation and motility by directly targeting MDM2. Mol Med Rep.
14:2657–2664. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chickooree D, Zhu K, Ram V, Wu HJ, He ZJ
and Zhang S: A preliminary microarray assay of the miRNA expression
signatures in buccal mucosa of oral submucous fibrosis patients. J
Oral Pathol Med. 45:691–697. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang H, Guo Y, Mi N and Zhou L: miR-101-3p
and miR-199b-5p promote cell apoptosis in oral cancer by targeting
BICC1. Mol Cell Probes. 52:1015672020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hawkins LJ and Storey KB: MicroRNA
expression in the heart of Xenopus laevis facilitates metabolic
adaptation to dehydration. Genomics. 112:3525–3536. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang X, Zhu X, Yan Z, Li C, Zhao H, Ma L,
Zhang D, Liu J, Liu Z, Du N, et al: miR-489-3p/SIX1 axis regulates
melanoma proliferation and glycolytic potential. Mol Ther
Oncolytics. 16:30–40. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen J, Wang J, Li H, Wang S, Xiang X and
Zhang D: p53 activates miR-192-5p to mediate vancomycin induced
AKI. Sci Rep. 6:388682016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shen ED, Liu B, Yu XS, Xiang ZF and Huang
HY: The effects of miR-1207-5p expression in peripheral blood on
cisplatin-based chemosensitivity of primary gallbladder carcinoma.
OncoTargets Ther. 9:3633–3642. 2016. View Article : Google Scholar
|
|
25
|
Vakkilainen S, Mäkitie R, Klemetti P,
Valta H, Taskinen M, Husebye ES and Mäkitie O: A wide spectrum of
autoimmune manifestations and other symptoms suggesting immune
dysregulation in patients with cartilage-hair hypoplasia. Front
Immunol. 9:24682018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ma Z, Teng Y, Liu X, Li J, Mo J, Sha M and
Li Y: Identification and functional profiling of differentially
expressed long non-coding RNAs in nasal mucosa with allergic
rhinitis. Tohoku J Exp Med. 242:143–150. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu HC, Liao Y and Liu CQ: miR-487b
mitigates allergic rhinitis through inhibition of the IL-33/ST2
signaling pathway. Eur Rev Med Pharmacol Sci. 22:8076–8083.
2018.PubMed/NCBI
|
|
28
|
Wang L, Lv Q, Song X, Jiang K and Zhang J:
ADRB2 suppresses IL-13-induced allergic rhinitis inflammatory
cytokine regulated by miR-15a-5p. Hum Cell. 32:306–315. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhu M, Yang M, Yang Q, Liu W, Geng H, Pan
L, Wang L, Ge R, Ji L, Cui S, et al: Chronic hypoxia-induced
microvessel proliferation and basal membrane degradation in the
bone marrow of rats regulated through the IL-6/JAK2/STAT3/MMP-9
pathway. BioMed Res Int. 2020:92047082020.PubMed/NCBI
|
|
30
|
Wang T, Chen D, Wang P, Xu Z and Li Y:
miR-375 prevents nasal mucosa cells from apoptosis and ameliorates
allergic rhinitis via inhibiting JAK2/STAT3 pathway. Biomed
Pharmacother. 103:621–627. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tang H, Wang H, Bai J, Ding M, Liu W, Xia
W, Luo Q, Xu G, Li H and Fang J; Nasal Health Group China, :
Sensitivity of MUC5AC to topical corticosteroid is negatively
associated with interleukin-17A in patients with allergic rhinitis.
Am J Rhinol Allergy. 26:359–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhai KF, Zheng JR, Tang YM, Li F, Lv YN,
Zhang YY, Gao Z, Qi J, Yu BY and Kou JP: The saponin D39 blocks
dissociation of non-muscular myosin heavy chain IIA from TNF
receptor 2, suppressing tissue factor expression and venous
thrombosis. Br J Pharmacol. 174:2818–2831. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhai KF, Duan H, Cui CY, Cao YY, Si JL,
Yang HJ, Wang YC, Cao WG, Gao GZ and Wei ZJ: Liquiritin from
glycyrrhiza uralensis attenuating rheumatoid arthritis via reducing
inflammation, suppressing angiogenesis, and inhibiting MAPK
signaling pathway. J Agric Food Chem. 67:2856–2864. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−∆∆C(T)) Method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhai KF, Duan H, Chen Y, Khan GJ, Cao WG,
Gao GZ, Shan LL and Wei ZJ: Apoptosis effects of imperatorin on
synoviocytes in rheumatoid arthritis through
mitochondrial/caspase-mediated pathways. Food Funct. 9:2070–2079.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ma Y, Shi L and Zheng C: Microarray
analysis of lncRNA and mRNA expression profiles in mice with
allergic rhinitis. Int J Pediatr Otorhinolaryngol. 104:58–65. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Teng Y, Zhang R, Liu C, Zhou L, Wang H,
Zhuang W, Huang Y and Hong Z: miR-143 inhibits
interleukin-13-induced inflammatory cytokine and mucus production
in nasal epithelial cells from allergic rhinitis patients by
targeting IL13Rα1. Biochem Biophys Res Commun. 457:58–64. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Incorvaia C, Cavaliere C, Frati F and
Masieri S: Allergic rhinitis. J Biol Regul Homeost Agents. 32
(Suppl 1):61–66. 2018.PubMed/NCBI
|
|
39
|
Zhou B, Li L, Qiu X, Wu J, Xu L and Shao
W: Long non-coding RNA ANRIL knockdown suppresses apoptosis and
pro-inflammatory cytokines while enhancing neurite outgrowth via
binding microRNA-125a in a cellular model of Alzheimer's disease.
Mol Med Rep. 22:1489–1497. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ping Y, Zhou Y, Hu J, Pang L, Xu C and
Xiao Y: Dissecting the functional mechanisms of somatic copy-number
alterations based on dysregulated ceRNA networks across cancers.
Mol Ther Nucleic Acids. 21:464–479. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wu H, Tian X and Zhu C: Knockdown of
lncRNA PVT1 inhibits prostate cancer progression in vitro and in
vivo by the suppression of KIF23 through stimulating miR-15a-5p.
Cancer Cell Int. 20:2832020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhu Y, Zhang X, Wang L, Zhu X, Xia Z, Xu L
and Xu J: FENDRR suppresses cervical cancer proliferation and
invasion by targeting miR-15a/b-5p and regulating TUBA1A
expression. Cancer Cell Int. 20:1522020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lou Y and Huang Z: microRNA-15a-5p
participates in sepsis by regulating the inflammatory response of
macrophages and targeting TNIP2. Exp Ther Med. 19:3060–3068.
2020.PubMed/NCBI
|
|
44
|
Yu L, Liu Z, He W, Chen H, Lai Z, Duan Y,
Cao X, Tao J, Xu C, Zhang Q, et al: Hydroxysafflor yellow A confers
neuroprotection from focal cerebral ischemia by modulating the
crosstalk between JAK2/STAT3 and SOCS3 signaling pathways. Cell Mol
Neurobiol. 40:1271–1281. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fang XY, Zhang H, Zhao L, Tan S, Ren QC,
Wang L and Shen XF: Corrigendum to ‘A new xanthatin analogue
1β-hydroxyl-5α-chloro-8-epi-xanthatin induces apoptosis through
ROS-mediated ERK/p38 MAPK activation and JAK2/STAT3 inhibition in
human hepatocellular carcinoma’ [Biochimie 152C (2018) 43–52].
Biochimie. 171-172:21–22. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wang J, Du A, Wang H and Li Y: MiR-599
regulates LPS-mediated apoptosis and inflammatory responses through
the JAK2/STAT3 signaling pathway via targeting ROCK1 in human
umbilical vein endothelial cells. Clin Exp Pharmacol Physiol.
47:1420–1428. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Peng DH, Liu YY, Chen W, Hu HN and Luo Y:
Epidermal growth factor alleviates cerebral ischemia-induced brain
injury by regulating expression of neutrophil gelatinase-associated
lipocalin. Biochem Biophys Res Commun. 524:963–969. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kabiri M, Hemmatpour A, Zare F,
Hadinedoushan H and Karimollah A: Paroxetine modulates immune
responses by activating a JAK2/STAT3 signaling pathway. J Biochem
Mol Toxicol. 34:e224642020. View Article : Google Scholar : PubMed/NCBI
|