Introduction
Pulmonary arterial hypertension (PAH) is a severe
pathophysiological syndrome characterized by pulmonary
vasoconstriction and structural remodeling of blood vessel walls,
which in turn lead to increased vascular resistance and right
ventricular dysfunction (1,2). The causes of PAH are complex, with
hypoxia being one of the most important factors (3,4). As
the main cells in pulmonary arteries, pulmonary artery smooth
muscle cells are sensitive to hypoxia (5). Under hypoxic conditions, abnormal
proliferation and migration of pulmonary artery smooth muscle cells
(PASMCs) cause pulmonary stenosis or occlusion and further induce
the occurrence and development of PAH (6). Therefore, strategies to inhibit the
abnormal proliferation and migration of PASMCs and reverse
pulmonary vascular remodeling are particularly important in the
treatment of PAH.
MicroRNAs (miRNAs/miRs) are small, endogenous RNAs
~22 nucleotides in length (7).
miRNAs serve an important regulatory role in organism development,
cell differentiation, proliferation and apoptosis, as well as in
pathological and physiological activities, including tumorigenesis
(8–10). miRNAs commonly negatively regulate
the expression of target genes by specifically binding to the
3′-untranslated region (3′-UTR) of the target mRNA molecule
(11). Recently, it has been
reported that a number of miRNAs serve a vital role in the
occurrence and development of PAH, highlighting their potential use
as a biomarker and therapeutic tool for PAH (12,13).
In addition, miRNAs regulate the PASMC phenotype and pulmonary
vascular remodeling under hypoxia (14). For example, decreased expression of
miR-34a in PASMCs induces cell proliferation, increases the
expression of platelet growth factor receptor α and promotes the
development of PAH; this phenomenon is reversed by overexpression
of miR-34a (15). miR-143-5p
modulates pulmonary artery smooth muscle cell functions in hypoxic
pulmonary hypertension through targeting hypoxia-inducible factor-1
(HIF-1)α (16). miR-153 is involved
in different pathophysiological processes in human diseases, such
as suppressing lung cancer (17,18),
inhibiting pulmonary fibrosis (19)
and inhibiting angiogenesis under hypoxic conditions (20). However, the effect of miR-153 on PAH
has yet to be reported.
ρ-associated, coiled-coil-containing protein kinase
1 (ROCK1) and nuclear factor of activated T cells cytoplasmic 3
(NFATc3) serve an important role in the formation of hypoxic
pulmonary hypertension. Inhibitors of these molecules significantly
inhibit the proliferation and migration of pulmonary smooth muscle
cells, constraining the development of pulmonary hypertension
(21–23). miRNAs are used to target NFATc3 to
inhibit PAH induced by the proliferation of human (H)PASMCs
(24). The present study explored
the ability of miR-153 to inhibit pulmonary vascular remodeling
induced by hypoxia by targeting ROCK1 and NFATc3, providing new
ideas for treatment of PAH.
Materials and methods
Animal experiment
Healthy male Sprague-Dawley (SD) rats (6–8 week-old;
180–200 g; n=30) were purchased from the Experimental Animal Center
of Jinzhou Medical University (Jinzhou, China). The present study
adhered to the Guide for the Care and Use of Laboratory Animals
published by the US National Institutes of Health (NIH Publication
No. 85-23, revised 1996) (25) and
all experimental protocols were approved by the Animal
Experimentation Ethics Committee of Jinzhou Medical University
(approval no. 2019065). The rats were randomly divided into five
groups including one control group (0 day) and four hypoxia groups
(1, 2, 4 and 6 weeks). Subsequentially, six rats in each group were
sacrificed for investigation at every time-point (0 day, 1, 2, 4
and 6 weeks). All rats in the normal and control groups survived
until sacrifice. The hypoxic rats were housed in a gas chamber with
a gas control delivery system (Oxycycler model A84XOV; BioSpherix,
Ltd.) for 12 h per day. The duration of the experiments was ~6
weeks (including 0 day, 1, 2, 4 and 6 weeks). During exposure to
hypoxia, oxygen concentration was set at 10±0.5% and age-matched
male rats were housed under normoxic conditions (21% oxygen) as
normal controls. All animals received humane care during the study.
All rats were housed at 18–23°C with 40–60% humidity, 12 h
light/dark cycles, and free access to food and water. Animal health
and behavior were monitored once a week.
Measurement of the right ventricular
systolic pressure (RVSP) and right ventricular hypertrophy index
(RVHI) in rats
The rats were anesthetized by the intraperitoneal
injection of 10% chloral hydrate (400 mg/kg) and fixed on an
experimental operating table. No signs of peritonitis were observed
after the administration of 10% chloral hydrate. The right external
jugular vein was exposed and dissociated. After the vessel was cut,
a prefilled copper heparin PE-50 catheter was quickly inserted into
the right ventricle via the vessel along the incision. The other
end of the catheter was connected to a multi-lead physiological
recording instrument by a pressure sensor. The pulmonary artery
systolic pressure was indirectly reflected by recording the RVSP.
Subsequently, rats were sacrificed directly by cervical dislocation
while unconsciousness. After cessation of breathing and heartbeat,
lung and heart samples were separated. The right ventricular (RV)
free wall was cut along the interventricular septum and the left
ventricle and ventricular septum (LV + S) were separated. Finally,
the RVHI was calculated after weighing based on the following
formula: RVHI = RV/(LV + S).
Morphological investigation
Lung tissues were obtained from anesthetized rats
and sagittal sections of right lungs were fixed in 4%
paraformaldehyde solution for 72 h, embedded in paraffin and
serially sectioned at 4 µm. Following dehydration using alcohol
dehydration (70, 80, 90, 100 and 100% alcohol; 30 min per step) and
clearing with benzene, sections were deparaffinized as follows:
Benzene for 5 min; 100% alcohol for 5 min; 90% alcohol for 2 min;
80% alcohol for 2 min; 70% alcohol for 2 min; and ddH2O
for 2 min. Tissue sections were then stained with hematoxylin for
10 min at room temperature, washed with water for 15 min and then
stained with eosin for 1 min at room temperature. Pulmonary
arteries (n=5; external diameters 50–100 µm) were randomly chosen
from each rat and observed using an optical light microscope
(magnification, ×200). The images were shot and analyzed with
Image-Pro Plus, Version 6.0 (Media Cybernetics, Inc.). The degree
of pulmonary artery remodeling was assessed by calculating the
percentage of the medial wall thickness (WT%) and medial wall area
(WA%) according to the following equations: WT% = 2 × WT/external
diameter) ×100; WA% = (medial WA/total vessel area) ×100. The
pulmonary arteries were isolated from the remaining lung tissues
and further used for western blot analyses.
Cell experiments
HPASMCs (ScienCell Research Laboratories, Inc.) were
cultured in a smooth muscle cell culture medium with fetal bovine
serum (FBS; Lonza Group, Ltd.) In the normal group, the cells were
cultured at 37°C under the condition of 21% O2, 5%
CO2 and 74% N2. In the hypoxia group, the
cells were cultured at 37°C under the condition of 3%
O2, 5% CO2 and 92% N2. After
exposure to hypoxia for 6, 12 and 24 h, the mRNA expression levels
of miR-153, ROCK1 and NFATc3 in HPASMCs were evaluated by reverse
transcription-quantitative (RT-q)PCR. Transfection efficiency was
estimated by evaluation of green fluorescent protein expression
under a fluorescence microscope.
Cell transfection
miR-153 (miR-153 mimic), negative control miRNA
(mimic control), miR-153 inhibitor (miR-153 inhibitor) and negative
control (inhibitor control) were purchased from Shanghai GenePharma
Co., Ltd. The sequences of the transfection vectors were as
follows: miR-153 mimics, 5′-UUGCAUAGUCACAAAAGUGAUC-3′; mimic
control, 5′-UUCUCCGAACGUGUCACGU-3′; miR-153 inhibitor,
5′-GAUCACUUUUGUGACUAUGCAA-3′; inhibitor control,
5′-CAGUACUUUUGUGUAGUACAA-3′. Subsequently, 100 nM miR-153 mimic,
mimic control, miR-153 inhibitor or inhibitor control were
transfected into HPASMCs (1×106 cells/well) using the
GP-siRNA-Mate kit (Shanghai GenePharma Co., Ltd.) according to the
manufacturer's instructions. The transfection controls were
non-targeting controls. At 48 h post-transfection, cells were used
for subsequent experiments.
RNA isolation and RT-qPCR
Total RNA was isolated from HPASMCs using a
TRIzol® reagent kit according to the manufacturer's
protocol (Thermo Fisher Scientific, Inc.). RNA extraction, cDNA
synthesis (total 20 µl) and quantitative analysis (total 20 µl) was
performed using the Hairpin-it miRNA qPCR quantification kit
(Shanghai GenePharma Co., Ltd.) according to the manufacturer's
protocol. U6 was used as an internal reference. The reverse
transcription procedure for U6 small nuclear RNA and miRNA-153 was:
25°C for 30 min, 42°C for 30 min, 85°C for 5 min and storage at
4°C. The reverse transcription procedure for GAPDH, ROCK1 and
NFATc3 was: 42°C for 45 min, 85°C for 5 min and storage at 4°C. The
PCR reaction program was as follows: Pre denaturation at 95°C for 5
min, followed by 40 cycles of 95°C denaturation for 12 sec and
annealing at 62°C for 40 sec. The primer sequences are listed in
Table I. miRNA levels was
quantified using the 2−ΔΔCq method (26) and expression value was normalized to
the internal control gene U6. The assay was performed in
triplicate.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) | Length of product
(bp) |
|---|
| miR-153 |
|
|
|
| F:
AGCCGCTTGCATAGTCACA | 78 |
|
| R:
AGAGCAGGGTCCGAGGAT | 78 |
| U6 |
|
|
|
| F:
CGCTTCGGCAGCACATATAC | 81 |
|
| R:
TTCACGAATTTGCGTGTCATC | 81 |
| ROCK1 |
|
|
|
| F:
CTGCAACTGGAACTCAACCAAGAA | 167 |
|
| R:
TTAGCACGCAATTGCTCAATATCAC | 167 |
| NFATC3 |
|
|
|
| F:
TCCACCTCCATCTACTTTAACCA | 162 |
|
| R:
TTGGGACCACCTAATGGGCT | 162 |
| GAPDH |
|
|
|
| F:
CATGAGAAGTATGACAACAGCCT | 113 |
|
| R:
AGTCCTTCCACGATACCAAAGT | 113 |
Cell proliferation assay
Cell proliferation was assessed using a cell
counting kit (CCK-8; Dojindo Molecular Technologies, Inc.).
Transfected HPASMCs were seeded in a 96-well plate at
5×103 cells/well. After the cells were incubated for 12
h under hypoxia, 10 µl of CCK-8 was added to each well and the
cells were incubated at 37°C for 2 h. The absorbance (optical
density value) was measured at 450 nm.
Wound healing (cell migration)
assay
Cell migration ability was measured by a wound
healing assay. In brief, miR-153 mimic, mimic control, miR-153
inhibitor and inhibitor control were transfected into HPASMCs.
Cells, which were cultured in medium containing 2% FBS, were plated
in 24-well cell culture plates at a density of 4×105
cells/well and cultured to confluence. Subsequently, a 200 µl
sterile pipette tip was used to scratch a straight line in the
monolayer of cells. After washing three times with PBS, the
remaining cells were cultured in serum-free medium for another 12
h. Images were captured at 0 and 12 h using an inverted light
microscope (IX71; Olympus Corporation; magnification, ×100). The
wounded area was manually outlined based on images acquired
immediately after wounding and 12 h later. Only wounds with areas
between 1.2 and 2.5 mm2 were analyzed to ensure that
data would be comparable. Finally, four fields of view were
observed for each sample. The experiments were repeated three
times.
Transwell cell migration assay
Cell migration was evaluated using 8-µm pore
Transwell chambers (Corning Life Sciences). The cells were
suspended in a serum-free medium at a density of 1×105
cells/ml. Then, 200 µl cell suspension was added into the upper
chamber and 600 µl medium containing 5% FBS was added into the
lower chambers. After 12 h incubation at 37°C under hypoxic
conditions, the cells had migrated to the lower chamber.
Non-migratory cells were removed using a cotton swab. Cells were
fixed with 95% ethanol for 15 min at room temperature and washed
three times with PBS (5 min per wash). Subsequently, cells were
stained with 0.1% crystal violet (Sigma-Aldrich; Merck KGaA) for 10
min at room temperature. Following washing with tap water,
migratory cells were observed using an inverted light microscope
(magnification, ×200).
Western blot analysis
ProteinExt Mammalian Total Protein Extraction kit
(TransGen Biotech Co., Ltd.) was used to isolate total proteins.
Protein concentrations were determined using a Pierce bicinchoninic
acid protein assay kit (Pierce; Thermo Fisher Scientific, Inc.) and
equal amounts of the protein (~50 µg) were isolated by 10% SDS-PAGE
and transferred onto polyvinylidene fluoride membranes. Membranes
were then blocked for 1 h at room temperature with 5% BSA in TBST
(0.1% Tween-20). The membranes were then incubated overnight at 4°C
with primary antibodies for detection of the following: ROCK1 (cat.
no. ab134181; 1:1,000; Abcam), PCNA (cat. no. K200030M; 1:1,000;
Beijing Solarbio Science & Technology Co., Ltd.), matrix
metalloproteinase (MMP)-2 (cat. no. A00286; 1:300; Wuhan Boster
Biological Technology, Ltd.), NFATc3 (cat. no. sc-8405; 1:200;
Santa Cruz Biotechnology, Inc.) and β-actin (cat. no. ab8227;
1:1,000; Abcam). The membranes were washed three times and were
then incubated for 1 h with horseradish peroxidase secondary
detection antibody (cat. nos. A0208 and A0216; 1:10,000; Beyotime
Institute of Biotechnology); β-actin was used as internal controls.
The signals of bands were measured using enhanced chemiluminescence
(ECL) detection kit (Beyotime Institute of Biotechnology) and
visualized on a commercial X-ray film. Relative protein levels were
semi-quantified using ImageJ software (version 1.41; National
Institutes of Health).
Immunofluorescence experiments
The transfected HPASMCs were seeded on cover slips
in a 24-well plate at a density of 4×104 cells/ml. After
48 h, the cells were fixed with 4% paraformaldehyde for 30 min and
washed three times with phosphate-buffered saline. Triton X-100
(1%; 200 µl; Sigma-Aldrich; Merck KGaA) was added into each well
for 30 min at room temperature. After the samples were washed three
times with PBS, HPASMCs were blocked with goat serum (1:500;
Beijing Solarbio Science & Technology Co., Ltd.) at room
temperature for 30 min. The cells were then incubated overnight at
4°C with the primary antibody for detection of NFATc3 (cat. no.
sc-8405; 1:200; Santa Cruz Biotechnology, Inc.). The cells were
then incubated for 2 h at room temperature with an Alexa Fluor 647
fluorescently labeled anti-mouse secondary antibody (cat. no.
A0473; 1:100; Beyotime Institute of Biotechnology). Cells were
finally double-stained with DAPI (1:1,000, Invitrogen; Thermo
Fisher Scientific, Inc.) for 5 min at room temperature and then
viewed by fluorescence microscopy.
Dual-luciferase reporter assay
Luciferase reporter assays were performed to verify
that miR-153 directly interacted with the 3′-UTR of ROCK1 and
NFATc3. The target association between miR-153 and ROCK1 or NFATc3
was verified by the TargetScan (www.targetscan.org/vert_72; version 7.0) database
(27). The wild-type (WT)
miR-153-ROCK1 or miR-153-NFATc3 (WT ROCK1 or WT NFATc3) and mutant
miR-153-ROCK1 or miR-153-NFATc3 (Mut ROCK1 or Mut NFATc3) plasmids
were designed and constructed according to the database of
TargetScan and miRanda (www.mirbase.org). The 3′-UTR of 3′-UTR/NFATc3 or ROCK1
containing the complementary binding site of miR-153 was directly
synthesized by Hanbio Biotechnology Co., Ltd. into the psiCHECK-2
vector (Promega Corporation) to construct the ROCK1 or NFATc3 WT
luciferase reporter vector (ROCK1-WT or NFATc3-WT). To establish
the ROCK1 mutant luciferase reporter vector (ROCK1-MUT), the
site-specific mutagenesis system (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to mutate the complementary binding site
(CUAUGCA) in the ROCK1-WT vector to GUCUCCC. In order to establish
the NFATc3 mutant luciferase reporter vector NFATc3 (-MUT), the
mutated complementary binding site (AAUGUGA) located in the
NFATc3-WT vector was mutated to CGGACUA. Then, 293T cells were
transfected with 100 ng ROCK1-WT or NFATc3-WT or ROCK1-MUT or
NFATc3-MUT vector and 500 ng miR-153 mimic for 48 h at 37°C.
Luciferase activity was measured with a Lumat LB9508 luminometer
(Berthold, Bad Wildbad, Germany). Relative luciferase intensity was
normalized to Renilla luciferase activity.
Statistical analysis
The data collected were statistically evaluated and
analyzed with SPSS v18.0 (SPSS, Inc.). All data are expressed as
mean ± standard deviation. Comparisons between the two unpaired
groups were performed using an unpaired Student's t-test.
Differences between multiple groups were tested by one-way analysis
of variance, followed by a Dunnett's post-hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of chronic hypoxia on RVSP,
RVHI, WA% and WT% in rats
As shown in Fig. 1A and
B, the RVSP and RVHI values were significantly increased from 2
and 4 weeks after the rats were exposed to hypoxia (P<0.01),
respectively. The WA% (Fig. 1C) and
WT% (Fig. 1D) values in rats under
hypoxia were significantly elevated from 4 weeks onwards
(P<0.05). After 6 weeks of hypoxia, the WT% value in hypoxic
rats was 1.9-fold higher than the level in normoxic control rats
and the WA% values were 1.7-fold higher compared with the level in
the control group. Hematoxylin and eosin staining demonstrated
marked thickening of the smooth muscle layer of pulmonary vessels
in the hypoxic group, with narrowing of the lumen and inflammatory
cell infiltration (Fig. 1E).
Morphological investigation showed the proliferation and migration
of PASMCs.
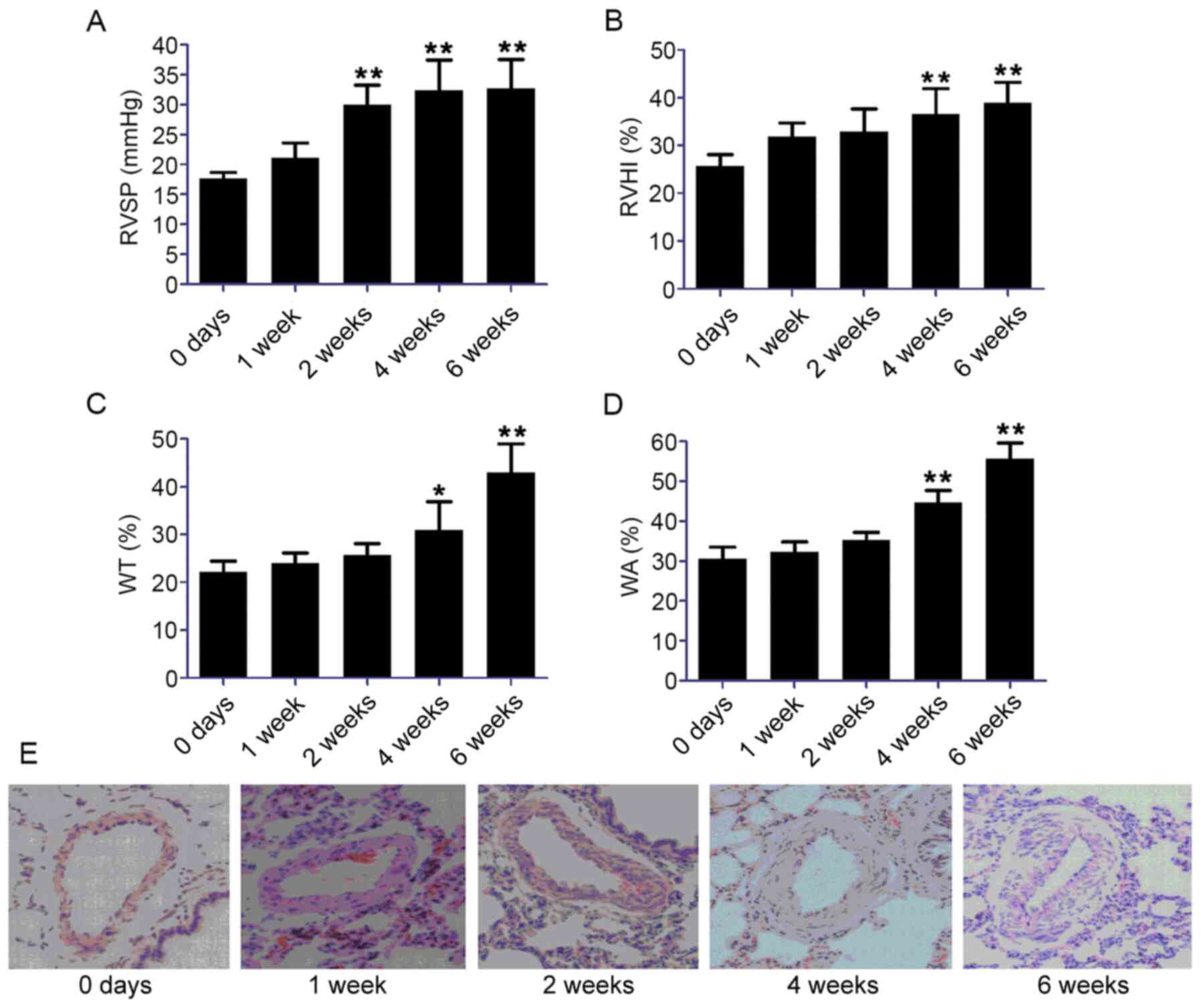 | Figure 1.Establishment of a rat model of HPH
and identification of rat pulmonary artery smooth muscle cells. (A)
RVSP and (B) RVHI, calculated as the weight ratio between the RV
and LV plus S: RV/LV+S; (C) WT and (D) WA of pulmonary arteries of
50–100 µm in diameter. (E) Hematoxylin and eosin staining of the
smooth muscle layer of pulmonary vessels in the hypoxic condition.
Scale bar, 100 µm. Data represent the mean ± standard deviation,
n=3. Comparisons were performed using one-way ANOVA followed by
Dunnett's post-hoc test. *P<0.05 and **P<0.01 vs. control at
day 0. HPH, hypoxia-induced pulmonary hypertension; RVSP, right
ventricular systolic pressure; RVHI, right ventricular hypertrophy
index; RV, right ventricle; LV, left ventricle; S, septum; WT,
medial wall thickness; WA, medial wall area. |
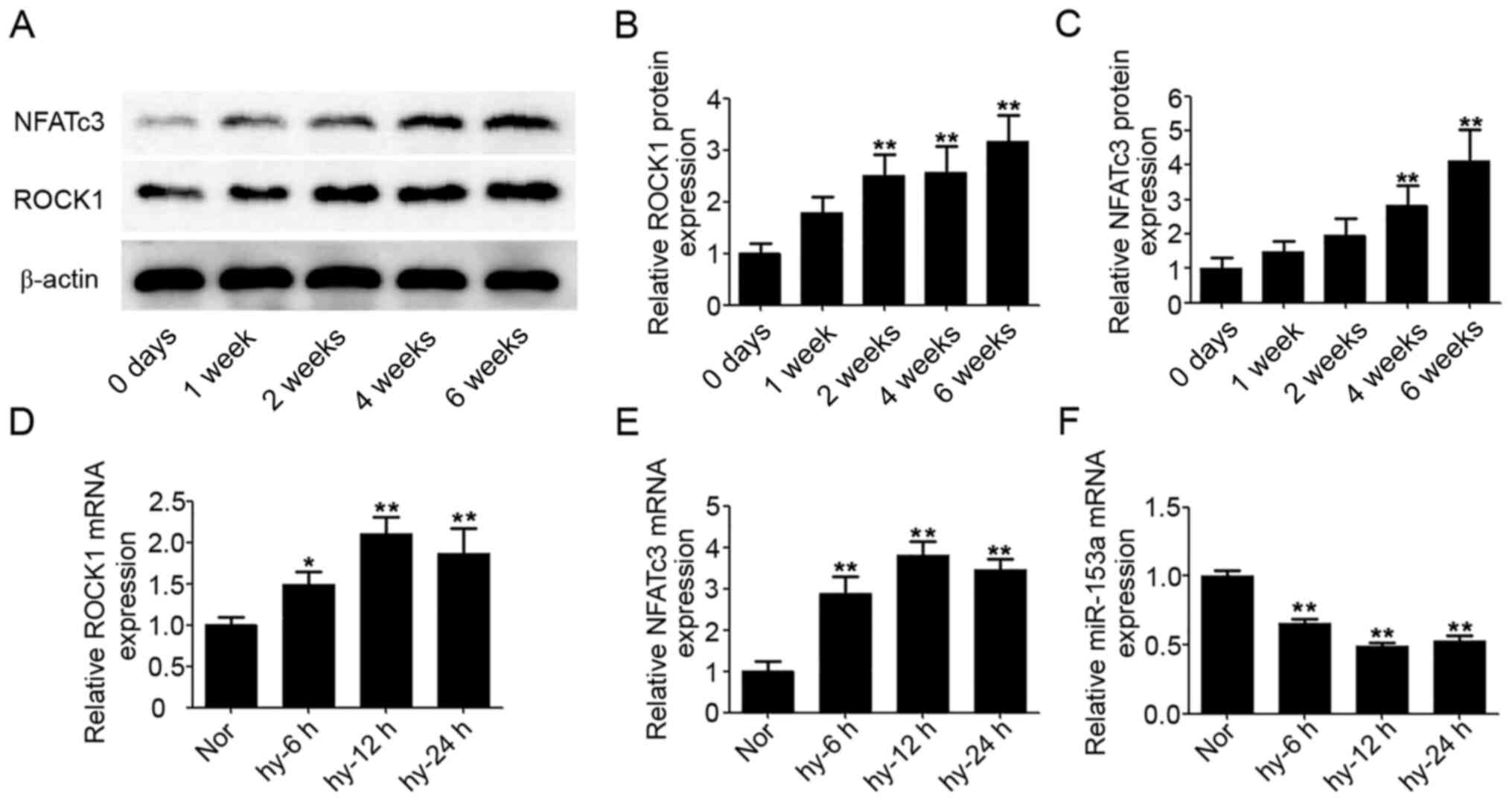
Upregulated expression levels of ROCK1
and NFATc3 proteins in hypoxic rats
At five time-points (0, 1, 2, 4 and 6 weeks) after
exposure to hypoxia, six rats in each group were sacrificed and the
pulmonary arteries removed. Western blotting was performed to
confirm the expression levels of ROCK1 and NFATc3 proteins in
pulmonary arteries; β-actin was used as the internal reference for
ROCK1 and NFATc3. As shown in Fig.
2A, the expression levels of ROCK1 and NFATc3 proteins were
increased from 2 and 4 weeks after exposure to hypoxia,
respectively. After 6 weeks, the expression levels of ROCK1 and
NFATc3 proteins were upregulated (2.1- and 2.3-fold, respectively)
compared with the level in the control group at day 0 (Fig. 2B and C).
miR-153, ROCK1 and NFATc3 expressions
are regulated by hypoxia in HPASMCs
From the results of rat models, the protein
expression levels of ROCK1 and NFATc3 were significantly increased
from 2 week after the rats were exposed to hypoxia (P<0.05).
Given the rapid changes of ROCK1 and NFATc3 protein expressions
under hypoxia, the gene changes within 24 h were determined using
HPASMCs under hypoxic conditions. After exposure of HPASMCs to
hypoxic conditions from 0 to 24 h, mRNA expressions of ROCK1 and
NFATc3 were clearly increased (Fig. 2D
and E). At the same time, the mRNA expression of miR-153 was
significantly decreased (Fig.
2F).
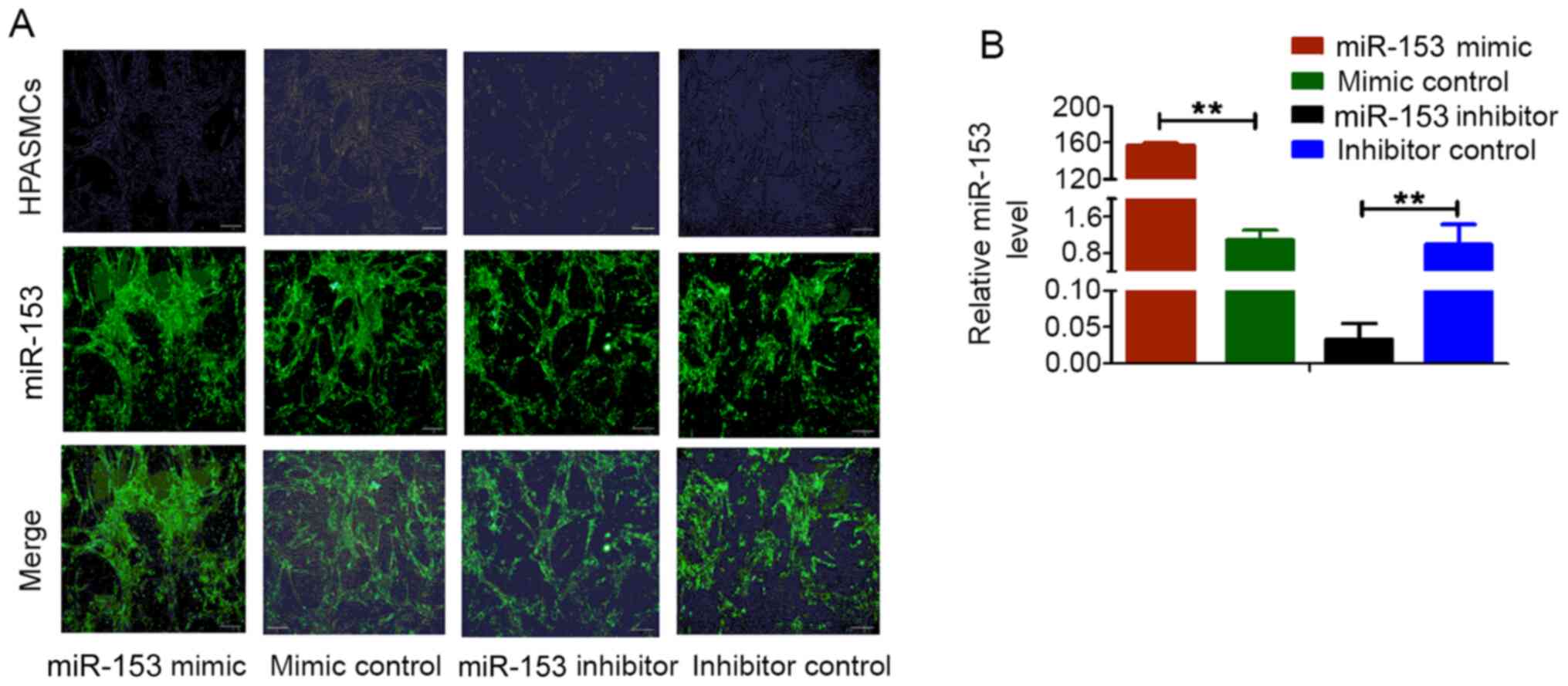
Validation of miR-153 mimic, mimic
control, miR-153 inhibitor and inhibitor control transfection into
HPASMCs
The immunofluorescence intensity of miR-153 in
HPASMs was increased after transfection of miR-153 mimic compared
with transfection with mimic control. RT-qPCR was used to detect
the expression level of miR-153 in HPASMCs following transfection;
the relative expression was normalized to that of U6, with that of
inhibitor control set as 1. Compared with control cells, miR-153
expression was markedly increased in cells transfected with the
miRNA-153 mimic and was decreased in cells transfected with the
miRNA-153 inhibitor (Fig. 3). These
observations demonstrated that miR-153 mimic, mimic control,
miR-153 inhibitor and inhibitor control were successfully
transfected into HPASMCs and could be used in subsequent
experiments.
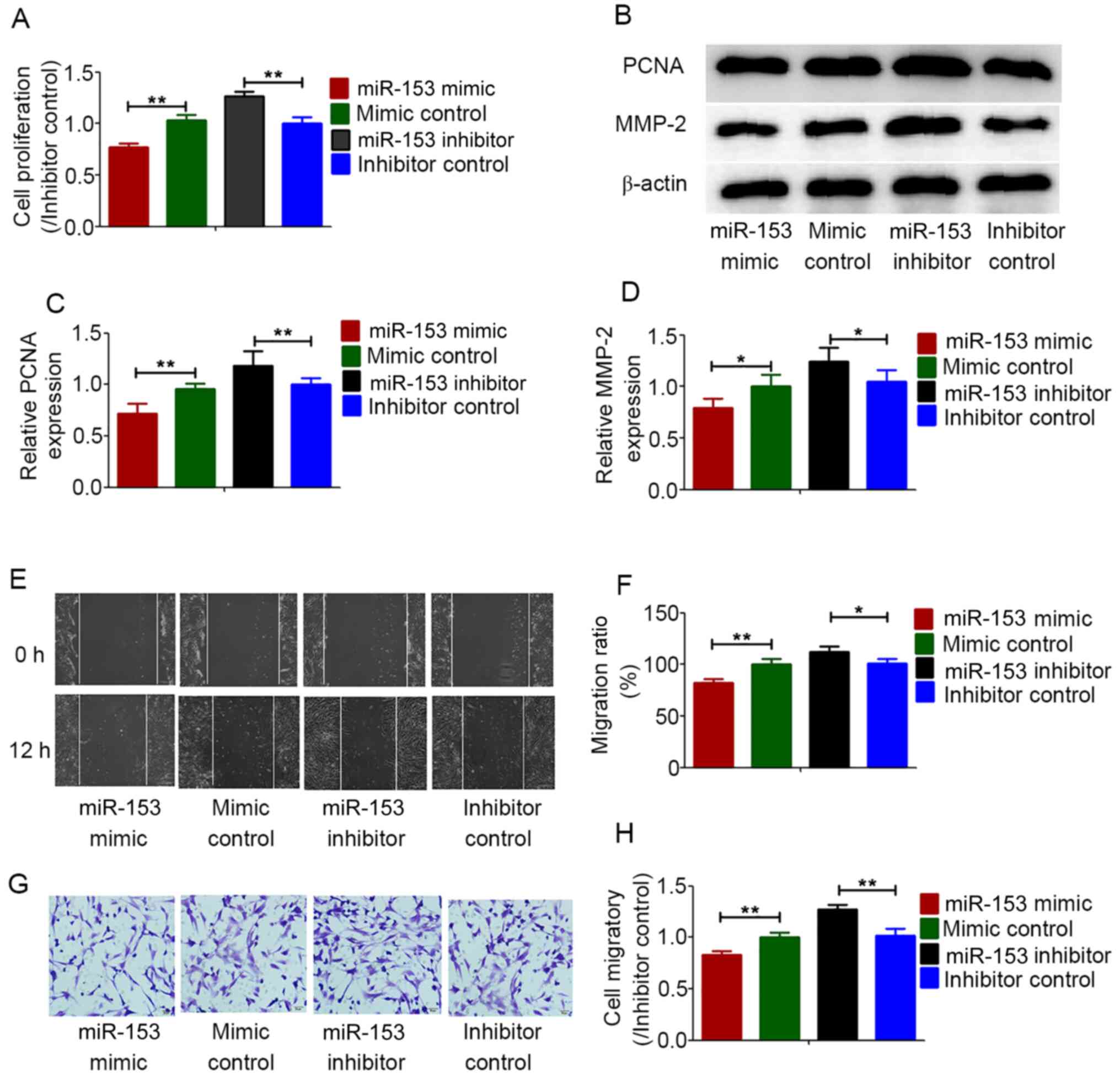
Effects of miR-153 on the
proliferation and migration of HPASMCs under hypoxic
conditions
The proliferation and migration of HPASMCs are
considered to be the cause of pulmonary vascular remodeling induced
by chronic hypoxia (28). To
determine the regulatory effect of miR-153 on HPASMCs under hypoxic
conditions, miR-153 was overexpressed in HPASMCs and then cell
proliferation and migration were analyzed under anoxic conditions.
Cell proliferation was determined using CCK-8 assays. MMP-2 and
PCNA expression was measured by western blot analysis. All the
results demonstrated that overexpression of miR-153 inhibited the
proliferation of HPASMCs under hypoxic conditions, while the
miR-153 inhibitor enhanced the proliferation of HPASMCs (Fig. 4A-C). Cell migration abilities were
determined using wound healing and Transwell assays. Under hypoxic
conditions, overexpression of miR-153 significantly inhibited the
migration abilities of HPASMCs, while the miR-153 inhibitor
enhanced the migration abilities of HPASMCs (Fig. 4B and D-H).
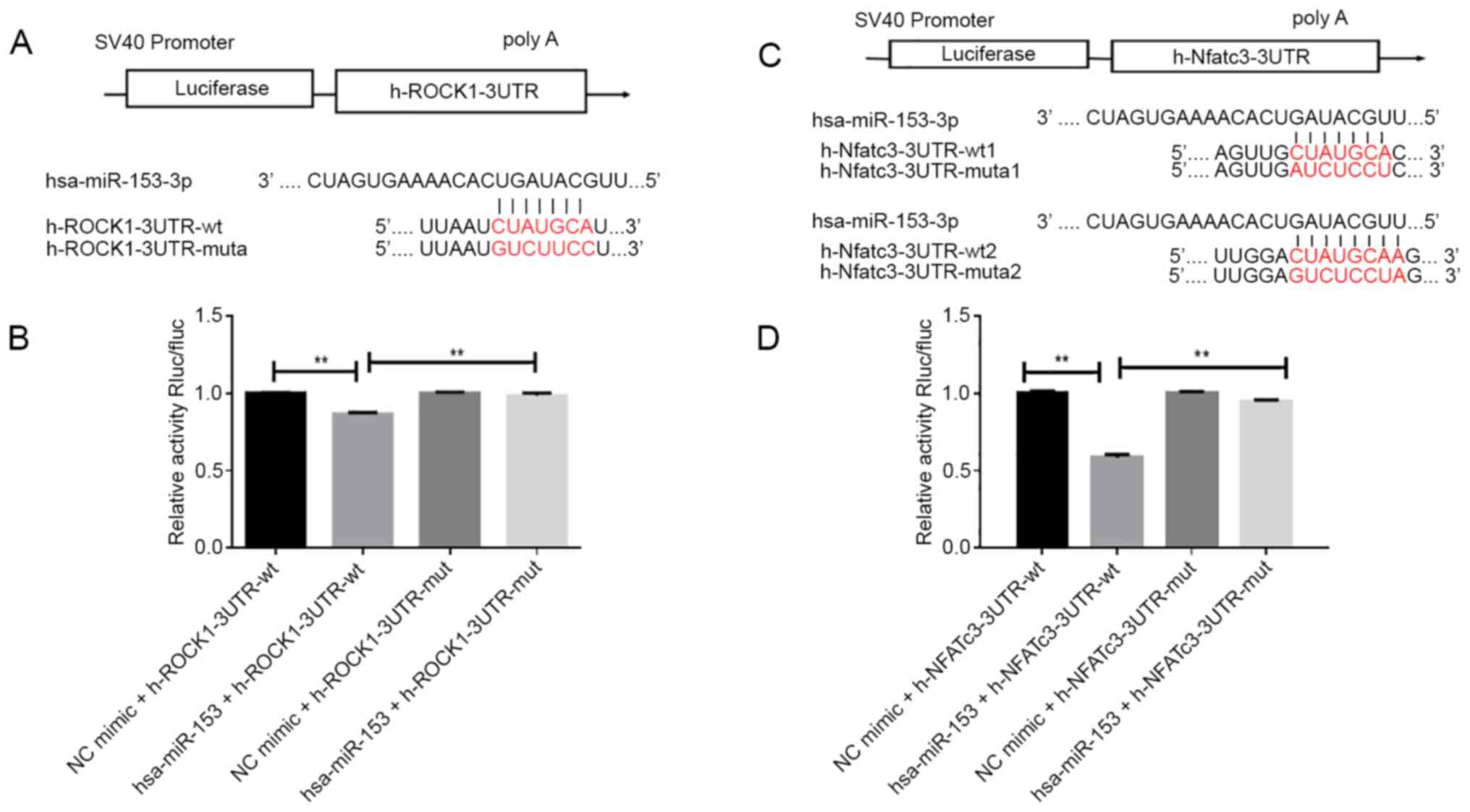
miR-153 directly targets ROCK1 or
NFATc3 in 293T cells
To further explore the molecular mechanism by which
miR-153 regulated the proliferation and migration of HPASMCs under
hypoxic conditions, TargetScan and miRanda were used to predict the
potential target sites of miR-153 in ROCK1 or NFATc3 mRNA. A region
in the ROCK1 mRNA was predicted to be a miR-153 binding site
(Fig. 5A) and two regions in the
NFATc3 mRNA were predicted to be miR-153 ROCK1 3′-UTR or
NFATc3-3′-UTR mutant plasmids (Fig.
5C).
Dual luciferase assays were then performed to
confirm the binding association between miR-153 and ROCK1 or
NFATc3. As shown in Fig. 5B and D,
the luciferase activity of the reporter was inhibited in 293T cells
co-transfected with miR-153 mimic compared with those
co-transfected the mimic control. However, there was no significant
change in the fluorescence intensity after mutation of the binding
site, suggesting that miR-153 interacted directly with ROCK1 or
NFATc3 at this binding site.
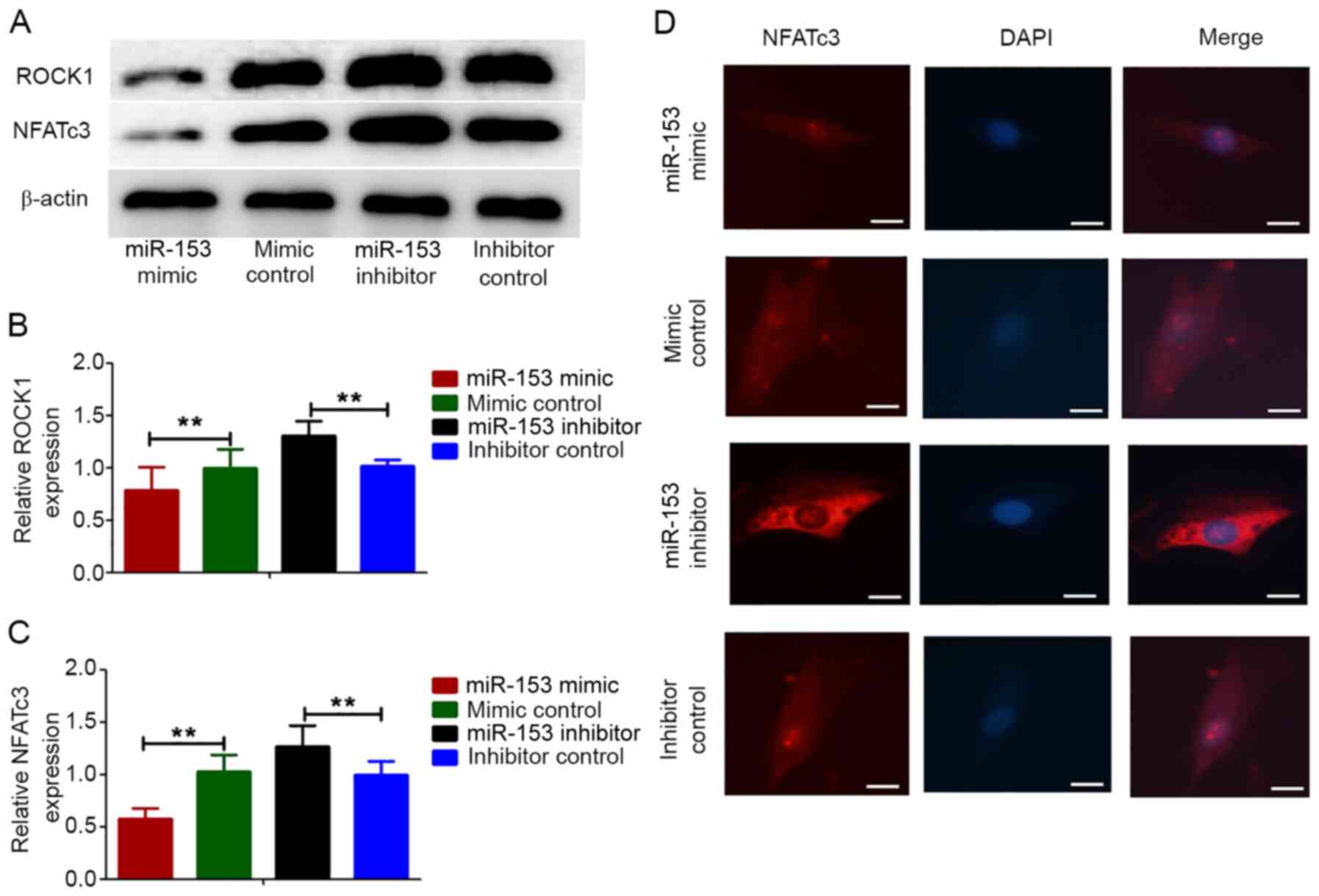
Effect of miR-153 on the expression of
ROCK1 and NFATc3 proteins in HPASMCs under hypoxic conditions
It has been reported that ROCK1 and NFATc3
expressions are related to PAH (29,30).
The present study investigated whether miR-153 regulated the
proliferation and migration of HPASMCs induced by hypoxia by
targeting ROCK1 and NFATc3. HPASMCs were transfected with
lentivirus vectors expressing the miRNA-153 mimic, miRNA-153
inhibitor and non-specific control and then they were treated with
hypoxia for 12 h. Compared with the control cells, the expression
levels of ROCK1 and NFATc3 proteins were markedly decreased in
cells transfected with the miRNA-153 mimic and were increased in
cells transfected with the miRNA-153 inhibitor (Fig. 6A-C). This result was consistent with
immunofluorescence experiment in that the expression of NFATc3
increased after inhibition of miRNA-153, while the expression of
NFATc3 decreases after overexpression of miRNA-153 (Fig. 6D).
Discussion
Chronic hypoxia is known to cause pulmonary
hypertension (31). The major
pathological changes associated with hypoxia-induced pulmonary
hypertension (HPH) are pulmonary vasoconstriction and remodeling
resulting from PAMSCs proliferation (32). The present study successfully
established a model of HPH. The RVSP, RVHI, WA% and WT% values of
rats in the model group exposed to hypoxia were significantly
increased compared with those of normal rats, suggesting that
hypoxia causes RV failure and pulmonary vascular remodeling.
Electron microscopic evaluation of the pulmonary vascular structure
revealed abnormal proliferation and migration of PAMSCs. In
addition, hypoxia significantly increased the thickness of lung
vessels and the proportion of muscularized pulmonary vessels. In
vitro studies were performed to confirm the effects of hypoxia on
PAMSCs.
ROCK1 and NFATc3 are involved in the regulation of
pulmonary vascular remodeling induced by hypoxia (30,33).
In the present study, expressions of ROCK1 and NFATc3 proteins in
the pulmonary vasculature were determined after rats were exposed
to hypoxia for 0, 1, 2, 4 and 6 weeks. The results demonstrated
significantly increased expression of ROCK1 and NFATc3 proteins
during the period of exposure to hypoxia. ROCK1 and NFATc3 mRNA
levels were evaluated after HPASMCs were cultured under hypoxia for
0, 6, 12 and 24 h and ROCK1 and NFATc3 expressions were
significantly upregulated. Given the important roles of ROCK1 and
NFATc3 in the formation of HPH (34,35),
ROCK1 and NFATc3 were related to the pathological process of HPH
and may even participate in this process.
Recently, a series of studies demonstrated that
hypoxic modulation of cellular function could be mediated by miRNAs
during the development of cancer (36–38).
HPH and cancer have similar phenotypes because both diseases are
characterized by increased cell proliferation and migration
(39,40). The present study chose miR-153 as
the research target because previous studies have reported its
antitumor function in different types of cancer including papillary
thyroid and breast cancer (41,42).
However, little is known about its expression and function in
HPASMCs in hypoxia. The present study revealed that miR-153
expression decreased in hypoxia-treated HPASMCs. It was also found
that miR-153 inhibitor increased HPASMC proliferation and
migration, while the miR-153 mimic reversed the increased
proliferation and migration of HPASMCs induced by hypoxia, which is
consistent with previous studies in cancer cells (43–45).
All these data suggested that miR-153 regulated hypoxia-induced
cell proliferation and migration.
The above data confirmed that miR-153 exhibited
preventive potential in the proliferation and migration of HPASMCs
induced by hypoxia, although the targets of miR-153 remained
elusive. ROCK1 and NFATc3 are important therapeutic targets for PAH
and they are activated and involved in the modulation of cell
proliferation and migration in PAH (46,47).
Therefore, an in-depth analysis was conducted using TargetScan to
determine whether NFATc3 and ROCK1 are the target genes of miR-153.
The dual-luciferase experiments verified that miR-153 directly
targeted NFATc3 and ROCK1. Using 293T cells, it was confirmed that
miR-153 interacted with the 3′-UTR of ROCK1 and NFATc3. Western
blotting and immunofluorescence analysis demonstrated that miR-153
significantly decreased the protein expressions of ROCK1 and
NFATc3. Thus, it was hypothesized that miR-153 directly targets
ROCK1 and NFATc3 to inhibit HPH. The translocation of
dephosphorylated NFATc3 from the cytoplasm to the nucleus is
essential for the subsequent activation of its target genes
(24). Future studies will
investigate the effect of miR-153 on nuclear translocation of
NFATc3. In addition, an HPH model of rats will be used to explore
the miR-153 expression in hypoxic pulmonary arteries with different
exposure time and confirm the effect of miR-153 on HPH.
In summary, the present study is the first, to the
best of the authors' knowledge, to provide evidence that miR-153
inhibited hypoxia-induced proliferation and migration of HPASMCs,
at least partly, through targeting ROCK1 and NFATc3. The present
study highlighted the biological significance of the miR-153/ROCK1
and miR-153/NFATc3 axes in HPASMCs under hypoxia. Further in vivo
studies should be performed to confirm that miR-153 is a promising
candidate for PAH therapy.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Nature Science Foundation of China (grant no. 81700050).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
MZ, LS and WW participated in the design of the
paper and experiment. MZ, YL and LS participated in the preliminary
experiment and the processing of experimental results. DK and NW
prepared the figures and conducted data analysis. MZ and YL were
involved in drafting the manuscript. All authors read and approved
the final manuscript.
Ethics approval and consent to
participate
The present study adhered to the Guide for the Care
and Use of Laboratory Animals published by the US National
Institutes of Health (NIH Publication No. 85-23, revised 1996) and
all experimental protocols were approved by the Animal
Experimentation Ethics Committee of Jinzhou Medical University
(approval no. 2019065).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kahraman BO, Savci S, Ozsoy I, Baran A,
Acar S, Ozpelit E, Balci A, Sevinc C and Akdeniz B: Effects of
neuromuscular electrical stimulation in patients with pulmonary
arterial hypertension: A randomized controlled pilot study. J
Cardiol. 75:702–708. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hiraide T, Kataoka M, Suzuki H, Aimi Y,
Chiba T, Isobe S, Katsumata Y, Goto S, Kanekura K, Yamada Y, et al:
Poor outcomes in carriers of the RNF213 variant (p.Arg4810Lys) with
pulmonary arterial hypertension. J Heart Lung Transplant.
39:103–112. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang L, Wang Y, Wu G, Rao L, Wei Y, Yue
H, Yuan T, Yang P, Xiong F, Zhang S, et al: Blockade of JAK2
protects mice against hypoxia-induced pulmonary arterial
hypertension by repressing pulmonary arterial smooth muscle cell
proliferation. Cell Prolif. 53:e127422020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Luo F, Wang X, Luo X, Li B, Zhu D, Sun H
and Tang Y: Invasive hemodynamic assessment for the right
ventricular system and hypoxia-induced pulmonary arterial
hypertension in mice. J Vis Exp. 152:e600902019.
|
|
5
|
Zhou W, Negash S, Liu J and Raj JU:
Modulation of pulmonary vascular smooth muscle cell phenotype in
hypoxia: Role of cGMP-dependent protein kinase and myocardin. Am J
Physiol Lung Cell Mol Physiol. 296:L780–L789. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Humbert M, Sitbon O, Chaouat A, Bertocchi
M, Habib G, Gressin V, Yaïci A, Weitzenblum E, Cordier JF, Chabot
F, et al: Survival in patients with idiopathic, familial, and
anorexigen-associated pulmonary arterial hypertension in the modern
management era. Circulation. 122:156–163. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ren ZP, Hou XB, Tian XD, Guo JT, Zhang LB,
Xue ZQ, Deng JQ, Zhang SW, Pan JY and Chu XY: Identification of
nine microRNAs as potential biomarkers for lung adenocarcinoma.
FEBS Open Bio. 9:315–327. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Peng XX, Yu R, Wu X, Wu SY, Pi C, Chen ZH,
Zhang XC, Gao CY, Shao YW, Liu L, et al: Correlation of plasma
exosomal microRNAs with the efficacy of immunotherapy in EGFR/ALK
wild-type advanced non-small cell lung cancer. J Immunother Cancer.
8:e0003762020.Erratum in: J Immunother Cancer 8: e000376corr1,
2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim D and Kim Y and Kim Y: Effects of
β-carotene on expression of selected micrornas, histone
acetylation, and DNA methylation in colon cancer stem cells. J
Cancer Prev. 24:224–232. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moloney BM, Gilligan KE, Joyce DP, O'Neill
CP, O'Brien KP, Khan S, Glynn CL, Waldron RM, Maguire CM, Holian E,
et al: Investigating the potential and pitfalls of EV-encapsulated
microRNAs as circulating biomarkers of breast cancer. Cells.
9:1412020. View Article : Google Scholar
|
|
11
|
Zhao L, Shan Y, Liu B, Li Y and Jia L:
Functional screen analysis reveals miR-3142 as central regulator in
chemoresistance and proliferation through activation of the
PTEN-AKT pathway in CML. Cell Death Dis. 8:e28302017.Retraction in:
Cell Death Dis 11: 121, 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhu TT, Zhang WF, Yin YL, Liu YH, Song P,
Xu J, Zhang MX and Li P: MicroRNA-140-5p targeting tumor necrosis
factor-α prevents pulmonary arterial hypertension. J Cell Physiol.
234:9535–9550. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Babicheva A, Ayon RJ, Zhao T, Ek Vitorin
JF, Pohl NM, Yamamura A, Yamamura H, Quinton BA, Ba M, Wu L, et al:
MicroRNA-mediated downregulation of K+ channels in pulmonary
arterial hypertension. Am J Physiol Lung Cell Mol Physiol.
318:L10–L26. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mohsenin V: The emerging role of microRNAs
in hypoxia-induced pulmonary hypertension. Sleep Breath.
20:1059–1067. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang P, Xu J, Hou Z, Wang F, Song Y, Wang
J, Zhu H and Jin H: miRNA-34a promotes proliferation of human
pulmonary artery smooth muscle cells by targeting PDGFRA. Cell
Prolif. 49:484–493. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tang BI, Tang MM, Xu QM, Guo JL, Xuan L,
Zhou J, Wang XJ, Zhang H and Kang PF: MicroRNA-143-5p modulates
pulmonary artery smooth muscle cells functions in hypoxic pulmonary
hypertension through targeting HIF-1α. J Biosci. 45:372020.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shan N, Shen L, Wang J, He D and Duan C:
MiR-153 inhibits migration and invasion of human non-small-cell
lung cancer by targeting ADAM19. Biochem Biophys Res Commun.
456:385–391. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang L, Lv X, Fu X, Su L, Yang T and Xu P:
MiR-153 inhibits the resistance of lung cancer to gefitinib via
modulating expression of ABCE1. Cancer Biomark. 25:361–369. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liang C, Li X, Zhang L, Cui D, Quan X and
Yang W: The anti-fibrotic effects of microRNA-153 by targeting
TGFBR-2 in pulmonary fibrosis. Exp Mol Pathol. 99:279–285. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liang H, Xiao J, Zhou Z, Wu J, Ge F, Li Z,
Zhang H, Sun J, Li F, Liu R, et al: Hypoxia induces miR-153 through
the IRE1α-XBP1 pathway to fine tune the HIF1α/VEGFA axis in breast
cancer angiogenesis. Oncogene. 37:1961–1975. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu WH, Xu XH, Luo Q, Zhang HL, Wang Y, Xi
QY, Zhao ZH and Liu ZH: Inhibition of the RhoA/Rho-associated,
coiled-coil-containing protein kinase-1 pathway is involved in the
therapeutic effects of simvastatin on pulmonary arterial
hypertension. Clin Exp Hypertens. 40:224–230. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang M, Chang Z, Zhang P, Jing Z, Yan L,
Feng J, Hu Z, Xu Q, Zhou W, Ma P, et al: Protective effects of
18β-glycyrrhetinic acid on pulmonary arterial hypertension via
regulation of Rho A/Rho kinsase pathway. Chem Biol Interact.
311:1087492019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhu J, Liu C, Wang B, Tong X and Li Z:
Qibai Pingfei capsule medicated serum inhibits the proliferation of
hypoxia-induced pulmonary arterial smooth muscle cells via the
Ca2+/calcineurin/nuclear factor of activated T-cells 3 pathway. J
Tradit Chin Med. 37:466–474. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kang K, Peng X, Zhang X, Wang Y, Zhang L,
Gao L, Weng T, Zhang H, Ramchandran R, Raj JU, et al: MicroRNA-124
suppresses the transactivation of nuclear factor of activated T
cells by targeting multiple genes and inhibits the proliferation of
pulmonary artery smooth muscle cells. J Biol Chem. 288:25414–25427.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hasnat AK, van der Velde ET, Hon JKF and
Yacoub MH: Reproducible model of post-infarction left ventricular
dysfunction: haemodynamic characterization by conductance catheter.
Eur J Cardiothorac Surg. 24:98–104. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:e050052015. View Article : Google Scholar
|
|
28
|
Zhao M, Chen N, Li X, Lin L and Chen X:
miR-19a modulates hypoxia-mediated cell proliferation and migration
via repressing PTEN in human pulmonary arterial smooth muscle. Life
Sci. 239:1169282019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Siddique MAH, Satoh K, Kurosawa R, Kikuchi
N, Elias-Al-Mamun M, Omura J, Satoh T, Nogi M, Sunamura S, Miyata
S, et al: Identification of emetine as a therapeutic agent for
pulmonary arterial hypertension: novel effects of an old drug.
Arterioscler Thromb Vasc Biol. 39:2367–2385. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Han J, Tian H, Liu Y and Fan F:
Sarpogrelate attenuates pulmonary arterial hypertension via
calcium/calcineurin axis. Front Biosci. 24:607–615. 2019.
View Article : Google Scholar
|
|
31
|
Yu L, Quinn DA, Garg HG and Hales CA:
Deficiency of the NHE1 gene prevents hypoxia-induced pulmonary
hypertension and vascular remodeling. Am J Respir Crit Care Med.
177:1276–1284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu J, Pan W, Wang C, Dong H, Xing L, Hou
J, Fang S, Li H, Yang F and Yu B: H2S attenuates
endoplasmic reticulum stress in hypoxia-induced pulmonary artery
hypertension. Biosci Rep. 39:BSR201903042019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu W, Li Y and Xu DQ: Role of ROS/Kv/HIF
Axis in the development of hypoxia-induced pulmonary hypertension.
Chin Med Sci J. 32:253–259. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Y, Duo D, Yan Y, He R and Wu X:
Magnesium lithospermate B ameliorates hypobaric hypoxia-induced
pulmonary arterial hypertension by inhibiting
endothelial-to-mesenchymal transition and its potential targets.
Biomed Pharmacother. 130:1105602020. View Article : Google Scholar
|
|
35
|
Bierer R, Nitta CH, Friedman J, Codianni
S, de Frutos S, Dominguez-Bautista JA, Howard TA, Resta TC and Bosc
LV: NFATc3 is required for chronic hypoxia-induced pulmonary
hypertension in adult and neonatal mice. Am J Physiol Lung Cell Mol
Physiol. 301:L872–L880. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shao H, Dong D and Shao F: Long non-coding
RNA TUG1-mediated down-regulation of KLF4 contributes to metastasis
and the epithelial-to-mesenchymal transition of colorectal cancer
by miR-153-1. Cancer Manag Res. 11:8699–8710. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xiao L, He Y, Peng F, Yang J and Yuan C:
Endometrial cancer cells promote M2-like macrophage polarization by
delivering exosomal miRNA-21 under hypoxia condition. J Immunol
Res. 2020:97310492020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yang Y, Qu A, Wu Q, Zhang X, Wang L, Li C,
Dong Z, Du L and Wang C: Prognostic value of a hypoxia-related
microRNA signature in patients with colorectal cancer. Aging
(Albany NY). 12:35–52. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Miao R, Liu W, Qi C, Song Y, Zhang Y, Fu
Y, Liu W, Lang Y, Zhang Y and Zhang Z: miR-18a-5p contributes to
enhanced proliferation and migration of PASMCs via targeting Notch2
in pulmonary arterial hypertension. Life Sci. 257:1179192020.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Peng Y and Wang HH: Cir-ITCH inhibits
gastric cancer migration, invasion and proliferation by regulating
the Wnt/β-catenin pathway. Sci Rep. 10:174432020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Luo Y, Teng X, Zhang L, Chen J, Liu Z,
Chen X, Zhao S, Yang S, Feng J and Yan X: CD146-HIF-1α hypoxic
reprogramming drives vascular remodeling and pulmonary arterial
hypertension. Nat Commun. 10:35512019.Erratum in: Nat Commun 10:
4098, 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cui Z, Luo Z, Lin Z, Shi L, Hong Y and Yan
C: Long non-coding RNA TTN-AS1 facilitates tumorigenesis of
papillary thyroid cancer through modulating the miR-153-3p/ZNRF2
axis. J Gene Med. 21:e30832019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zuo Z, Ye F, Liu Z, Huang J and Gong Y:
MicroRNA-153 inhibits cell proliferation, migration, invasion and
epithelial-mesenchymal transition in breast cancer via direct
targeting of RUNX2. Exp Ther Med. 17:4693–4702. 2019.PubMed/NCBI
|
|
44
|
Wang J, Liang S and Duan X: Molecular
mechanism of miR-153 inhibiting migration, invasion and
epithelial-mesenchymal transition of breast cancer by regulating
transforming growth factor beta (TGF-β) signaling pathway. J Cell
Biochem. 120:9539–9546. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ma H, Tian T, Liu X, Xia M, Chen C, Mai L,
Xie S and Yu L: Upregulated circ_0005576 facilitates cervical
cancer progression via the miR-153/KIF20A axis. Biomed
Pharmacother. 118:1093112019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
He RL, Wu ZJ, Liu XR, Gui LX, Wang RX and
Lin MJ: Calcineurin/NFAT signaling modulates pulmonary artery
smooth muscle cell proliferation, migration and apoptosis in
monocrotaline-induced pulmonary arterial hypertension rats. Cell
Physiol Biochem. 49:172–189. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Huetsch JC, Walker J, Undem C, Lade J, Yun
X, Baksh S, Jiang H, Lai N and Shimoda LA: Rho kinase and Na+/H+
exchanger mediate endothelin-1-induced pulmonary arterial smooth
muscle cell proliferation and migration. Physiol Rep. 6:e136982018.
View Article : Google Scholar : PubMed/NCBI
|