Introduction
Inflammatory bowel disease (IBD) most commonly
refers to ulcerative colitis and Crohn's disease, which are
conditions characterized by chronic gastrointestinal tract
inflammation (1). IBD has a
multifactorial etiology that involves the interplay of
environmental, genetic and immunological factors (2). Its common pathogenic feature is
disruption of the integrity of the intestinal epithelial barrier
(3). Under normal conditions,
several integral cellular proteins that maintain robust
intercellular connections between epithelial cells support the
intestinal mucosal barrier (4). The
barrier is mainly composed of intercellular junctional complexes
(5–7), which consist of tight junction (TJ)
proteins (occludin and claudin-1) interacting with the central
protein zona occludens (ZO)-1. TJ disruption leads to disturbances
in the paracellular barrier and an increase in intestinal
epithelial paracellular permeability. This alteration in
permeability causes potential harmful antigens and luminal bacteria
to penetrate the intestine, resulting in the initiation and
acceleration of the mucosal inflammation in IBD (8–11).
Thus, therapies that attenuate intestinal barrier dysfunction could
effectively treat IBD (10–12).
Natural products commonly used in Traditional
Chinese Medicine (TCM) have gained an increased medical interest
worldwide, due to their potent anti-inflammatory role (13–15).
One well-known traditional Chinese formula is Qing Hua Chang Yin
(QHCY), which is composed of Coptis chinensis Franch, Herba
et Gemma Agrimoniae, Radix Sanguisorbae, Magnolia
officinalis, Radix Paeoniae Rubra, Elettaria cardamomum,
Semen Coicis, Artemisia capillaris Thunb, Semen Dolichoris
Album, Herba Eupatorii Fortunei and Poria cocos. In TCM,
QHCY is considered beneficial in the treatment of UC; therefore,
QHCY has been used in the management of UC for several years in
China (16–21).
In mice, QHCY has been reported to alleviate the
clinical and histological manifestations of dextran sulfate
sodium-induced colitis (22–24);
this effect was revealed to be partly mediated by a reduction in
inflammatory cytokine release via the TLR4/NF-κB and IL-6/STAT3
signaling pathways. However, to the best of our knowledge, the
effect of QHCY on intestinal epithelial barrier function is
unknown. To elucidate the therapeutic mechanism underlying the
effects of QHCY, the present study investigated the in vitro
effects of QHCY on the intestinal epithelial barrier.
Materials and methods
Materials and reagents
Dulbecco's modified Eagle's medium (DMEM; cat. no.
C11995500BT), fetal bovine serum (FBS; cat. no. 10091148),
penicillin-streptomycin (cat. no. 15070063) and
trypsin-ethylenediaminetetraacetic acid (cat. no. 25200072) were
purchased from Gibco; Thermo Fisher Scientific, Inc.
Lipopolysaccharide (LPS; Escherichia coli serotype 055:B5;
cat. no. L6529) and fluorescein isothiocyanate-dextran
(FITC-dextran 4: FD4; cat. no. FD4) were acquired from
Sigma-Aldrich; Merck KGaA. The M-PER Mammalian Protein Extraction
Reagent (cat. no. 78501) and BCA assay kit (cat. no. 23227) were
purchased from Thermo Fisher Scientific, Inc. Anti-ZO-1 (cat. no.
GTX108592) was obtained from GeneTex, Inc., and anti-occludin (cat.
no. 13409-1-AP) and anti-claudin-1 (cat. no. 13050-1-AP) antibodies
were obtained from Proteintech Group, Inc. Moreover, anti-β-actin
antibody (cat. no. 4970) and horseradish peroxidase
(HRP)-conjugated secondary antibody (cat. no. 7074) were acquired
from Cell Signaling Technology, Inc. The human TNF-α ELISA kit
(cat. no. 430204) was obtained from BioLegend, Inc. RNAiso plus
reagent and the PrimeScriptRT reagent kit were obtained from Takara
Biotechnology Co., Ltd. Unless otherwise noted, all other reagents
were obtained from Sigma-Aldrich; Merck KGaA.
QHCY preparation
QHCY was prepared as previously described (23). Briefly, the following dehydrated
amounts of each component were used in the preparation: 33 g
Coptis chinensis Franch, 220 g Herba et Gemma Agrimoniae,
110 g Radix Paeoniae Rubra, 100 g Radix Sanguisorbae, 110 g
Magnolia officinalis, 56 g Elettaria cardamomum, 110
g Herba Eupatorii Fortunei, 110 g Artemisia capillaris
Thunb., 110 g Semen Dolichoris Album, 220 g Semen Coicis and 220 g
Poria cocos (obtained from the Department of Pharmacy,
Second People's Hospital Affiliated to Fujian University of
Traditional Chinese Medicine, Fuzhou, Fujian, China). The mixture
was extracted by boiling three times in 2 l distilled water. The
extracts were filtered and concentrated by boiling to a final
volume of 1 l. The stock concentration of QHCY was 1.4 g/ml.
Cell culture
Cells were cultured as previously described
(23). Human colon cancer Caco-2
cells (cat. no. HTB37) were purchased from the American Type
Culture Collection. The cells were cultured in DMEM supplemented
with 10% (v/v) FBS, glucose (1 g/l), penicillin (50 U/ml) and
streptomycin (50 µg/ml) in a humidified incubator containing 5%
CO2 at 37°C. Subsequently, cells were subcultured at
85–90% confluence and differentiated into enterocyte-like cells
18–20 days later, as described previously (22,23).
Fully differentiated cells were used for further experiments.
Cell Counting Kit-8 (CCK-8) assay
Cell viability was assessed using CCK-8 (Dojindo
Technologies, Inc.). Differentiated Caco-2 cells in 96-well plates
were treated with the indicated concentrations of LPS (0–80 µg/ml)
for 24 h at 37°C. Subsequently, 10 µl CCK-8 was added to each well
and incubated for 2 h at 37°C. Absorbance was measured at 405 nm
using a fluorescence plate reader (Model ELx80; BioTek
Corporation).
TNF-α ELISA assay
As described previously (23), differentiated Caco-2 cells in
24-well plates were incubated with QHCY (10 and 50 µg/ml) for 1 h
before stimulation with LPS (1 µg/ml) for 24 h at 37°C.
Subsequently, the supernatants were collected by centrifuging the
cell culture medium at 3,000 × g for 10 min at room temperature.
Using a human TNF-α ELISA kit, the production of TNF-α from Caco-2
cells was measured, according to the manufacturer's instructions.
Absorbance was read at 450 nm using a fluorescence plate reader.
All samples were assessed in triplicate.
Transepithelial electrical resistance
(TEER) measurement
Caco-2 cells (5×104 cells/well) were
seeded in the upper chamber of 24-well plates containing Transwell
inserts and cultured for 18–20 days before experimentation; the
culture medium was changed every other day. To determine TJ
formation, the TEER of the Caco-2 cell monolayer was measured using
a Millicell ERS (EMD Millipore). For the subsequent experiments,
monolayers with TEER between 400 and 500 Ω cm2 were
used, and were treated with QHCY (10 and 50 µg/ml) for 1 h before
LPS (1 µg/ml) stimulation for 24 h at 37°C. TEER was measured
before and after treatment. Moreover, triplet cell monolayers were
assessed for each experimental group. TEER changes during
experimental conditions were calculated as the percentage of
baseline levels.
Measurement of permeability
To evaluate paracellular permeability, the
fluorescently-labeled dextran 4 (FD4: cat. no. FD4; Sigma-Aldrich;
Merck KGaA) flux from apical to basolateral was measured as
previously described, with minor modifications (25). Briefly, Caco-2 cells
(5×104 cells/well) were seeded in the upper chamber of
24-well plates containing Transwell inserts and were cultured for
18–20 days before experimentation; the culture medium in the lower
chamber was changed every other day. The TEER of the Caco-2 cell
monolayer was measured using a Millicell ERS (EMD Millipore). For
the subsequent experiments, the cell monolayers with TEER between
400 and 500 Ω cm2 were used, and treated with QHCY and
LPS as aforementioned. The medium was then removed, and 200 µl FD4
(5 mg/ml) and 500 µl PBS was added to the apical and basolateral
compartments of each Transwell insert, respectively. After 2 h of
incubation at 37°C, 100 µl was removed from the basolateral
compartment and transferred to 96-well plates. Subsequently, FD4
concentration was determined using a fluorescence plate reader at
an excitation wavelength of 480/492 nm and an emission wavelength
of 520/525 nm. All samples were assessed three times.
Reverse transcription-quantitative PCR
(RT-qPCR) assay
Differentiated Caco-2 cells in 6-well plates were
incubated with QHCY and LPS as aforementioned. According to the
manufacturer's instructions, total cellular RNA was extracted using
RNAiso plus reagent (Takara Biotechnology Co., Ltd.) and RT was
conducted using the PrimeScript RT reagent kit (Takara
Biotechnology Co., Ltd.). The mRNA expression levels of ZO-1,
occludin and claudin-1 were determined by qPCR using SYBR green dye
(Thermo Fisher Scientific, Inc.) and the ABI 7500 fast sequence
detection system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). An initial denaturation step was performed at 95°C for 30
sec, followed by 40 cycles at 95°C for 3 sec and annealing at 60°C
for 30 sec. GAPDH served as an internal control. The sequences of
primers used for qPCR are provided in Table I. The mRNA expression levels of
genes of interest were quantified using 2−∆∆Cq analysis
(26). mRNA expression levels are
expressed as fold change relative to the control group.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene name | Sequences
(5′-3′) |
|---|
| ZO-1 | Forward:
AGCCTGCAAAGCCAGCTCA |
|
| Reverse:
AGTGGCCTGGATGGGTTCATAG |
| Claudin-1 | Forward:
GCATGAAGTGTATGAAGTGCTTGGA |
|
| Reverse:
CGATTCTATTGCCATACCATGCTG |
| Occludin | Forward:
CTTTGGCTACGGAGGTGGCTAT |
|
| Reverse:
CTTTGGCTGCTCTTGGGTCTG |
| GAPDH | Forward:
GCACCGTCAAGGCTGAGAAC |
|
| Reverse:
ATGGTGGTGAAGACGCCAGT |
Western blot analysis
Differentiated Caco-2 cells in 6-well plates were
incubated with QHCY (10 and 50 µg/ml) for 1 h before stimulation
with LPS (1 µg/ml) for 24 h at 37°C. A mammalian cell lysis buffer
containing a phosphatase inhibitor and a protease inhibitor
cocktail (EMD Millipore) (23) was
used to lyse the differentiated Caco-2 cells incubated with QHCY
and LPS for 24 h at 37°C. Lysed cells were then centrifuged at
12,000 × g for 15 min at 4°C and the supernatants were collected.
Protein concentrations were measured using the BCA protein assay
kit. Equal amounts of protein (50 µg) from each sample were
resolved on 12% Tris-glycine gels and transferred onto
nitrocellulose membranes. The membranes were blocked for 1 h at
room temperature with SuperBlock buffer (Thermo Fisher Scientific,
Inc.) and were then incubated with primary antibodies against ZO-1
(rabbit, polyclonal; 1:1,000), occludin (rabbit, polyclonal;
1:1,000), claudin-1 (rabbit, polyclonal; 1:1,000) and β-actin
(rabbit, polyclonal; 1:2,000) at 4°C overnight. Subsequently,
membranes were incubated with the appropriate anti-rabbit IgG,
HRP-linked secondary antibody (1:5,000) for 1 h at room
temperature. Thereafter, the membranes were visualized using
enhanced chemiluminescence using SuperSignal™ West Dura Extended
Duration Substrate (cat. no. 34076; Thermo Fisher Scientific,
Inc.). Images were captured using an imaging system (Bio-Rad
Laboratories, Inc.). The protein levels were analyzed with ImageJ
Software (National Institutes of Health). β-actin protein
expression was used as an internal control.
Statistical analysis
All data were analyzed using SPSS software (22.0)
for Windows (IBM Corp.). Data are presented as the mean ± SD of at
least three independent experiments. The significance of
differences among groups that conformed to a normal distribution
was determined by one-way ANOVA followed by Tukey's post hoc test.
Two-tailed P<0.05 was considered to indicate a statistically
significant difference.
Results
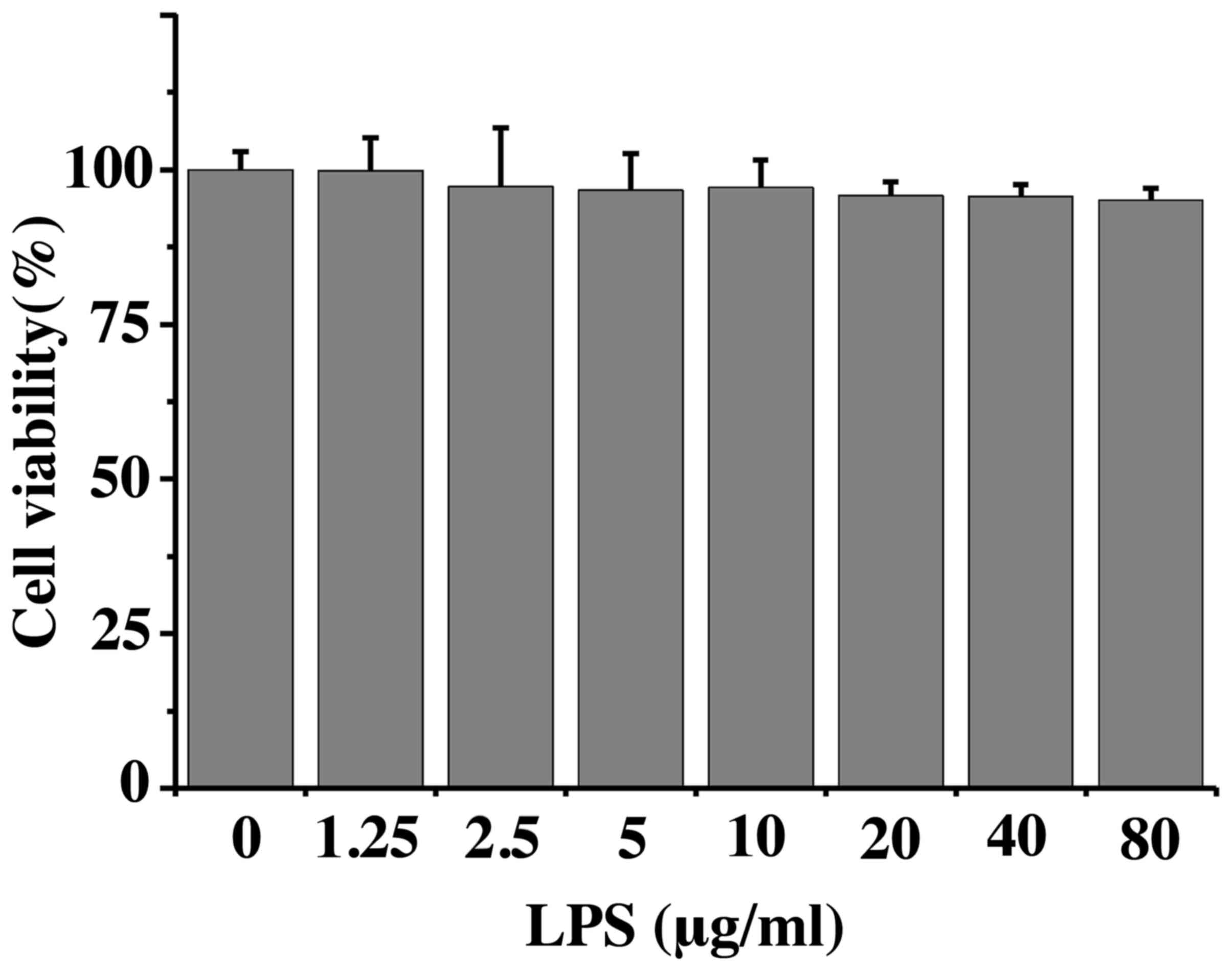
LPS does not affect Caco-2 cell
viability
Caco-2 cell viability was assessed following
exposure to LPS (Fig. 1); cell
viability was not affected by even the highest concentration of LPS
tested (80 µg/ml). These findings indicated that the observations
made in subsequent experiments were not caused by LPS-induced cell
death. A concentration of 10 µg/ml LPS was used for all further
experiments.
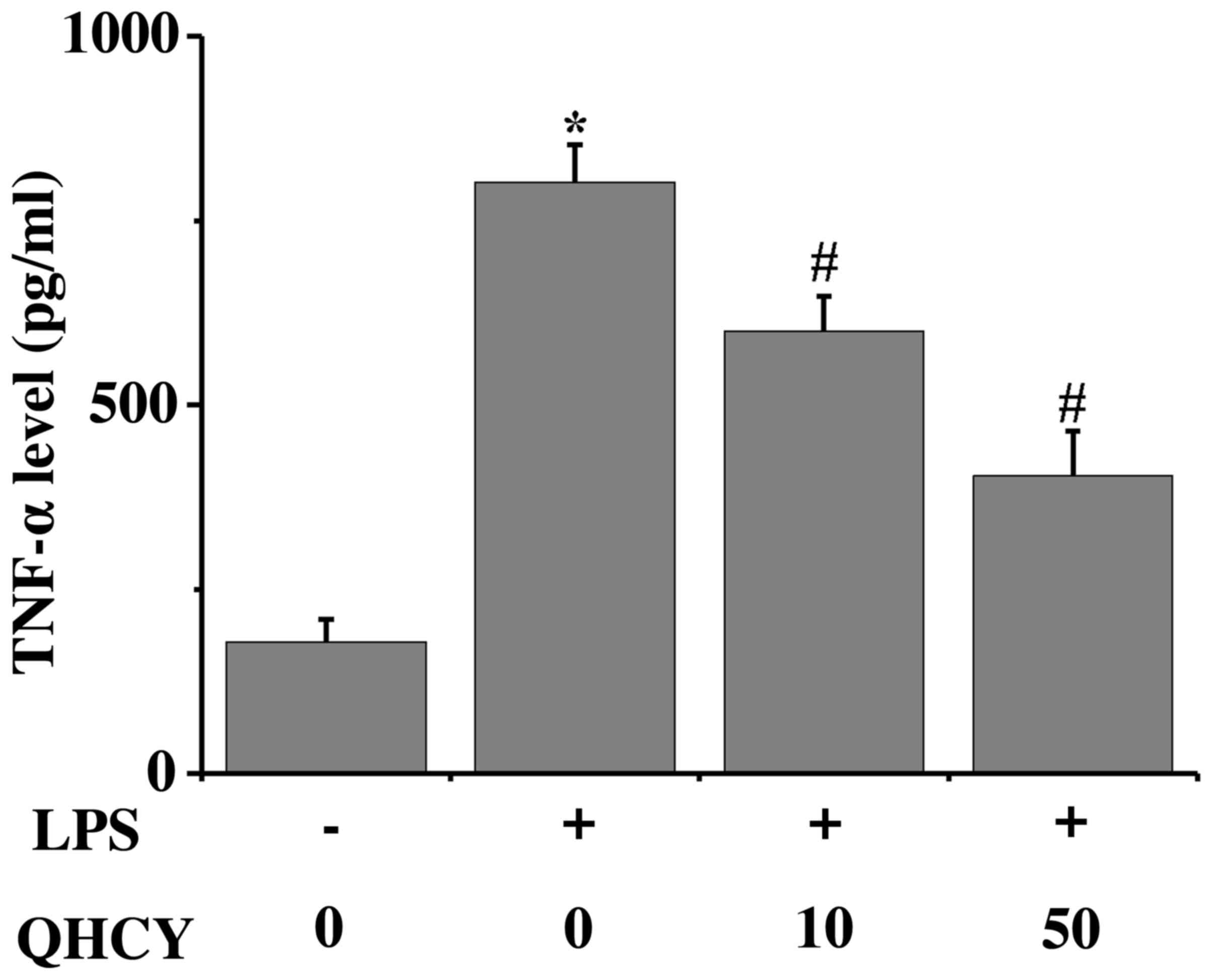
QHCY inhibits the LPS-induced
production of the proinflammatory cytokine TNF-α
TNF-α has been reported to participate in the
pathogenesis of various inflammatory disorders, including IBD
(26). When exposed to LPS, Caco-2
cells produced increased levels of TNF-α compared with in the
control group. To demonstrate the biological activity of the QHCY
preparation, the effects of QHCY on TNF-α production by LPS-exposed
Caco-2 cells were assessed (Fig.
2). As anticipated, LPS induced TNF-α secretion from the Caco-2
cells; however, QHCY significantly suppressed LPS-induced TNF-α
secretion, which is consistent with our previous observation
(24).
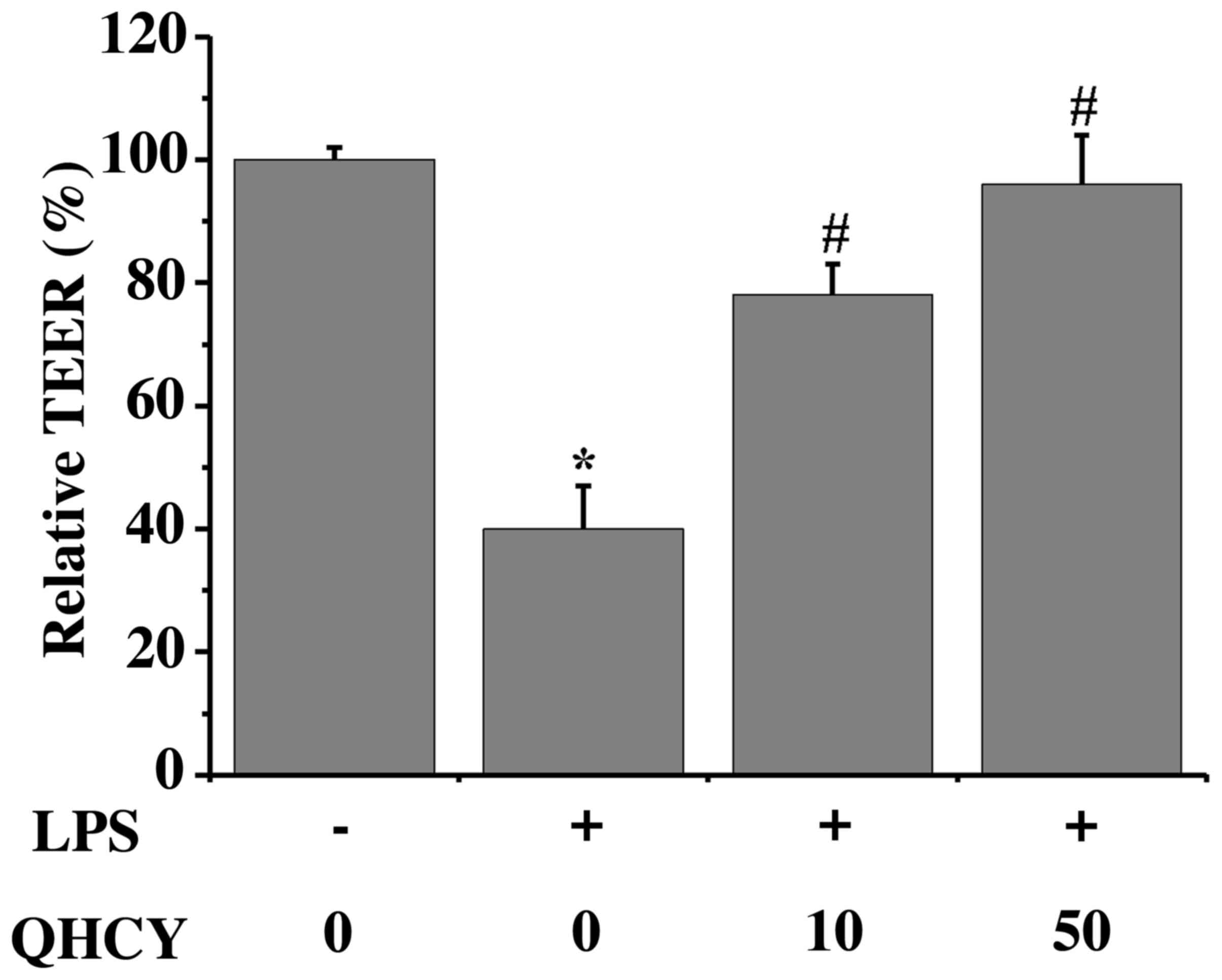
QHCY inhibits inflammation-induced
paracellular permeability in Caco-2 cell monolayers
To investigate whether QHCY can preserve the
intestinal epithelial barrier during inflammation, Caco-2 cell
monolayers were pretreated with QHCY before LPS exposure (Fig. 3). LPS significantly decreased the
TEER of the Caco-2 cell monolayers compared with that in the
control group (control, 100±2.3%; LPS, 39.8±7.5%), indicating an
increase in permeability. However, pretreatment with QHCY
ameliorated the LPS-induced decrease in TEER, demonstrating that
the normal barrier function was protected by QHCY treatment (10
µg/ml QHCY, 78.0±6.0%; 50 µg/ml QHCY, 96.4±7.6%). Moreover, as
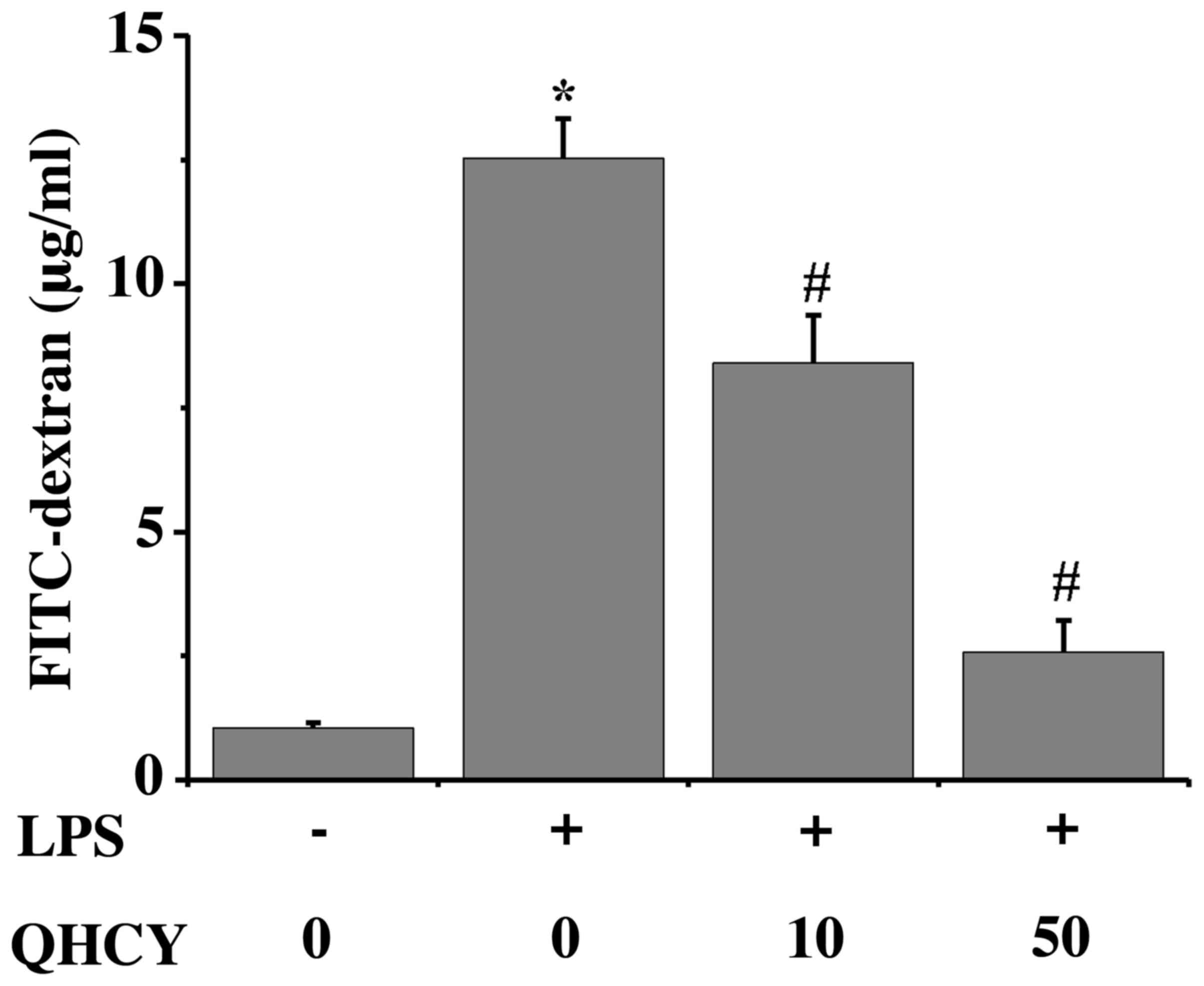
another measure of Caco-2 monolayer permeability, FD4 flux was
measured (Fig. 4). Consistent with
the TEER results, LPS significantly increased FD4 flux compared
with that in the control group (control, 1.04±0.11 µg/ml; LPS,
12.52±0.80 µg/ml). Pretreatment with QHCY provided protection
against the LPS-induced hyperpermeability of Caco-2 cell monolayers
(10 µg/ml QHCY, 8.40±0.97 µg/ml; 50 µg/ml QHCY, 2.57±0.64 µg/ml).
Taken together, these findings suggested that QHCY could contribute
to the maintenance of mucosal barrier integrity against
inflammation-induced permeability.
QHCY prevents LPS-induced disruption
of TJs in Caco-2 cell monolayers
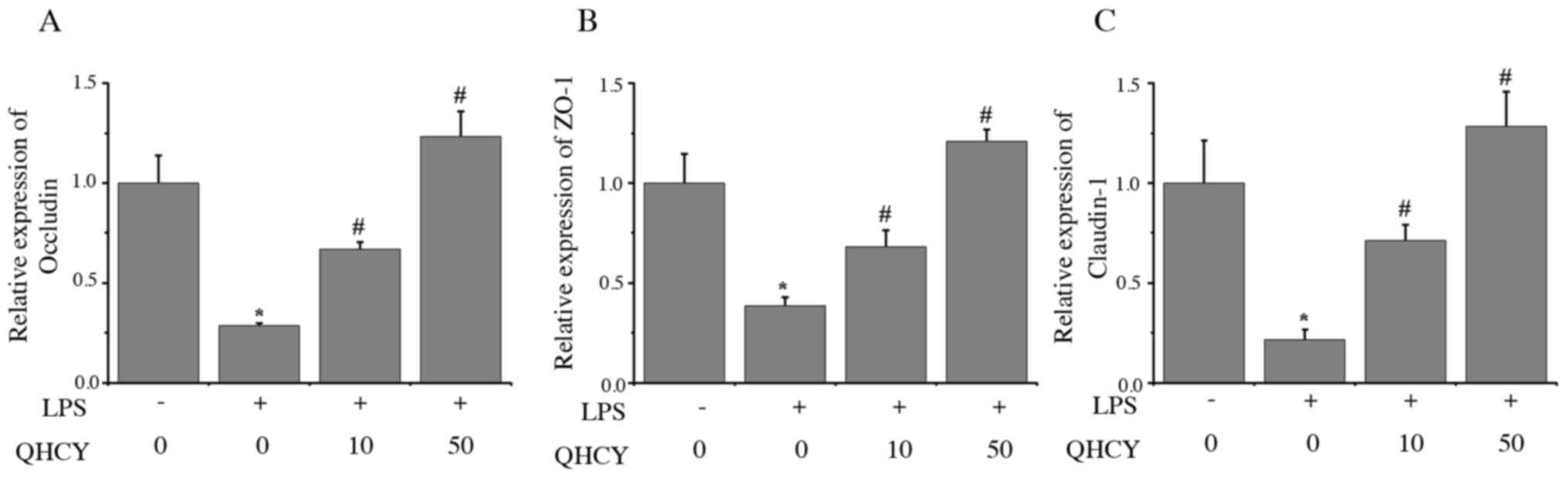
The present study investigated the expression levels
of TJ-associated factors by RT-qPCR and western blot analyses, in
order to determine the mechanisms underlying the protective effect
of QHCY on epithelial cell monolayer permeability. The effects of
QHCY on the mRNA expression levels of ZO-1, occludin and claudin-1
in epithelial cells were determined by RT-qPCR. As shown in
Fig. 5, the mRNA expression levels
of occludin (Fig. 5A), ZO-1
(Fig. 5B) and claudin-1 (Fig. 5C) were significantly decreased after
LPS treatment compared with those in the control group, whereas
treatment with QHCY significantly reversed the effect of LPS on the
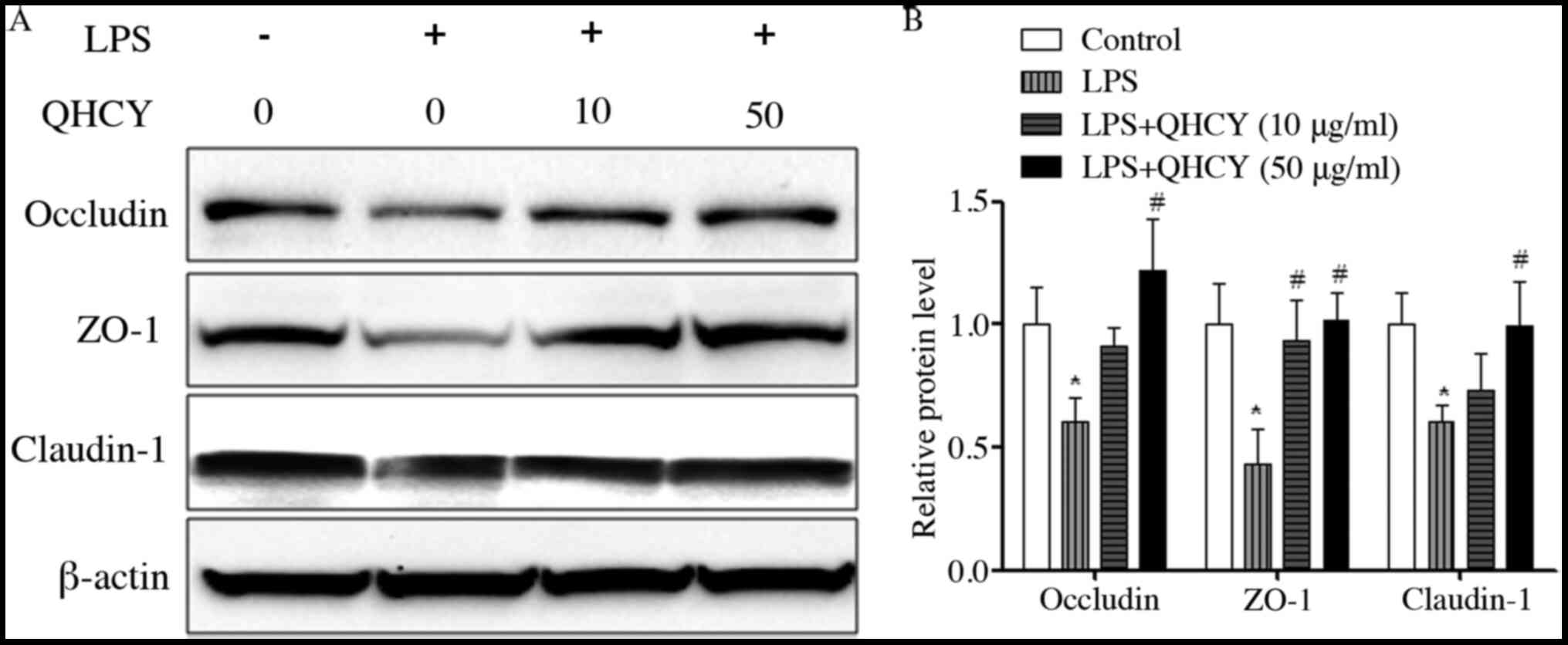
expression levels of ZO-1, occludin and claudin-1. Similarly, the
effects of QHCY on the protein expression levels of ZO-1, occludin
and claudin-1 in epithelial cells were determined using western
blot-ting. As shown in Fig. 6A and
B, compared with in the control group, treatment with LPS
induced a significant reduction in the protein expression levels of
TJ-associated factors, ZO-1, occludin and claudin-1, which was
reversed by the administration of QHCY. These findings suggested
that preventing the reduction in the expression levels of TJ
proteins may be the mechanism underlying the effects of QHCY on the
LPS-induced increase in epithelial cell monolayer permeability.
Discussion
Despite recent advances in therapy, patients with
IBD still suffer from disease manifestations stemming from
unremitting active inflammation. Moreover, current therapies are
related to significant potential adverse reactions, such as
systemic immunosuppression, headache, nausea and fatigue (27,28);
therefore, alternative effective therapies for IBD are required.
The anti-inflammatory effects of numerous natural products have
gained considerable attention due to their relatively low toxicity
(24). QHCY, which is a
well-recognized traditional Chinese formula, has been used in
clinical settings for treating several inflammatory disorders,
including arthritis, hepatitis, cholecystitis and IBD (29). However, the mechanisms underlying
the biological activity of QHCY remain to be elucidated. The
barrier function of colonic mucosa is maintained by TJs and their
molecular components, including ZO-1, occludin and claudin-1, and
an increase in intestinal permeability is often documented in
intestinal barrier dysfunction (30–32).
Thus, the present study examined the effects of QHCY on cell
permeability and TJ protein expression using an in vitro
model of intestinal epithelium that has been used in similar
studies (8,33–41).
Intestinal permeability serves a critical role in
host defense (34). Patients with
IBD exhibit increased intestinal mucosal permeability, which is
correlated with disease severity (8,35–37).
In the present study, QHCY significantly alleviated LPS-induced
increases in the permeability of epithelial cell monolayers. This
effect was observed by both TEER and FD4 flux assays. Thus, QHCY
may contribute to maintaining the integrity of the mucosal barrier
during inflammatory insults.
The intestinal epithelial barrier consists of
epithelial cells joined together by intercellular junctional
complexes, which include the TJ proteins ZO-1, occludin and
claudin-1 (5). These proteins are
crucial for barrier function maintenance, and their expression has
been shown to be reduced in response to inflammatory states,
including in IBD (36–39). In IBD, claudin-1 expression was
significantly decreased (42). As
previously demonstrated that QHCY significantly reversed the
DSS-induced downregulation of ZO-1, occludin, and claudin-1 in mRNA
and protein levels in the mice (30). The present study revealed that LPS
markedly suppressed the expression of ZO-1, occludin and claudin-1
at both the mRNA and protein expression levels in IEC, whereas QHCY
significantly reversed this effect. Therefore, QHCY may regulate TJ
expression to maintain mucosal barrier integrity. Possibly, the
effects of QHCY observed on the expression levels of TJ proteins
were indirect and caused by TNF-α activity suppression, since QHCY
could inhibit the LPS-induced production of the proinflammatory
cytokine TNF-α. Notably, QHCY may exert potential anti-inflammatory
effects and could be considered a useful therapy for the treatment
of IBD.
In conclusion, QHCY reduced the LPS-induced
secretion of TNF-α and reversed the increase in permeability of
Caco-2 cell monolayers, thereby preserving the expression levels of
ZO-1, occludin and claudin-1. Therefore, QHCY may be considered a
potential therapeutic agent for the treatment of IBD, due to its
direct suppressive effect on the secretion of proinflammatory
cytokines and the maintenance of epithelial barrier function.
However, further investigations should be conducted to determine
the underlying potential mechanism of QHCY treatment for IBD.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81673731) and the Natural
Science Foundation of Fujian Province (grant no. 2017J01302).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XK, TJS and JP acquired funding for the research.
WF, PZ, AS, YL, SS and XK conceived and designed the experiments.
WF, PZ, AS, LL, HC, SS and YC performed the experiments. WF, PZ, TS
and AS analyzed the data. YC, TJS and JP acquired, interpreted the
data and confirmed the authenticity of the raw data associated with
the preparation of the manuscript. JP, TJS and SS wrote the
manuscript. JP, YL, TJS and XK checked the manuscript. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
IBD
|
inflammatory bowel disease
|
|
QHCY
|
Qing Hua Chang Yin
|
|
TJ
|
tight junction
|
|
UC
|
ulcerative colitis
|
|
TCM
|
Traditional Chinese Medicine
|
References
|
1
|
Strober W, Fuss I and Mannon P: The
fundamental basis of inflammatory bowel disease. J Clin Invest.
117:514–521. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Talley NJ, Abreu MT, Achkar JP, Bernstein
CN, Dubinsky MC, Hanauer SB, Kane SV, Sandborn WJ, Ullman TA and
Moayyedi P; American College of Gastroenterology IBD Task Force, :
An evidence-based systematic review on medical therapies for
inflammatory bowel disease. Am J Gastroenterol. 106 (Suppl
1):S2–S25; quiz S26. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee JY, Wasinger VC, Yau YY, Chuang E,
Yajnik V and Leong RW: Molecular pathophysiology of epithelial
barrier dysfunction in inflammatory bowel diseases. Proteomes.
6:E172018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Camilleri M, Madsen K, Spiller R,
Greenwood-Van Meerveld B and Verne GN: Intestinal barrier function
in health and gastrointestinal disease. Neurogastroenterol Motil.
24:503–512. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma TY, Iwamoto GK, Hoa NT, Akotia V,
Pedram A, Boivin MA and Said HM: TNF-alpha-induced increase in
intestinal epithelial tight junction permeability requires NF-kappa
B activation. Am J Physiol Gastrointest Liver Physiol.
286:G367–G376. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ivanov AI, Nusrat A and Parkos CA: The
epithelium in inflammatory bowel disease: Potential role of
endocytosis of junctional proteins in barrier disruption. Novartis
Found Symp. 263:115–124; discussion 124–132, 211–218.
2004.PubMed/NCBI
|
|
7
|
Niessen CM: Tight junctions/adherens
junctions: Basic structure and function. J Invest Dermatol.
127:2525–2532. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Turner JR: Intestinal mucosal barrier
function in health and disease. Nat Rev Immunol. 9:799–809. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Laukoetter MG, Nava P, Lee WY, Severson
EA, Capaldo CT, Babbin BA, Williams IR, Koval M, Peatman E,
Campbell JA, et al: JAM-A regulates permeability and inflammation
in the intestine in vivo. J Exp Med. 204:3067–3076. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Arrieta MC, Madsen K, Doyle J and Meddings
J: Reducing small intestinal permeability attenuates colitis in the
IL10 gene-deficient mouse. Gut. 58:41–48. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Edelblum KL and Turner JR: The tight
junction in inflammatory disease: Communication breakdown. Curr
Opin Pharmacol. 9:715–720. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Arnott ID, Kingstone K and Ghosh S:
Abnormal intestinal permeability predicts relapse in inactive Crohn
disease. Scand J Gastroenterol. 35:1163–1169. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ahmad TB, Liu L, Kotiw M and Benkendorff
K: Review of anti-inflammatory, immune-modulatory and wound healing
properties of molluscs. J Ethnopharmacol. 210:156–178. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gao L, Jia C, Zhang H and Ma C: Wenjing
decoction (herbal medicine) for the treatment of primary
dysmenorrhea: A systematic review and meta-analysis. Arch Gynecol
Obstet. 296:679–689. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang LJ, Zhu JY, Sun MY, Song YN, Rahman
K, Peng C, Zhang M, Ye YM and Zhang H: Anti-inflammatory effect of
Man-Pen-Fang, a Chinese herbal compound, on chronic pelvic
inflammation in rats. J Ethnopharmacol. 208:57–65. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang XY and Tian DL: Etiological and
pathological characteristics of ulcerative colitis and TCM
differentiation and treatment. Beijing Zhong Yi Yao Da Xue Xue Bao.
30:554–559. 2007.(In Chinese).
|
|
17
|
Gong YP, Liu W, Ma GT, Hu HY, Xie JQ, Tang
ZP, Hao WW, Bian H, Zhu LY, et al: Randomized control study of
‘Qingchang Suppository’ on ulcerative colitis. Shanghai Zhong Yi
Yao Da Xue Xue Bao. 21:33–36. 2007.(In Chinese).
|
|
18
|
Fu NL and Huang JY: Progress of clinical
research of traditional Chinese medicine for the treatment of
ulcerative colitis. J Tradit Chin Med. 40:501–503. 1999.(In
Chinese).
|
|
19
|
Li QG: An idea about treatment of
ulcerative colitis by TCM methods. Beijing Zhong Yi. 23:149–150.
2004.(In Chinese).
|
|
20
|
Wang CH, Gao WY, Li YF, Chen SQ, Yang Z,
Lu YP, Gong Y and Liu Y: Study of Fufangkushen colon-release
capsule on ulcerative colitis of endo-retention of damp heat type.
Xian Dai Zhong Xi Yi Jie He Za Zh. 18:13–15. 2009.(In Chinese).
|
|
21
|
Chen JT, Ke X, Fu XY, Wang WR, Hu GH and
Yang CB: The clinical study of heat-clearing and damp-drying on the
treatment of damp-heat ulcerative colitis. Zhongguo Zhong Xi Yi Jie
He Xiao Hua Za Zh. 17:256–258. 2009.(In Chinese).
|
|
22
|
Ke X, Hu G, Fang W, Chen J, Zhang X, Yang
C, Peng J, Chen Y and Sferra TJ: Qing Hua Chang Yin inhibits the
LPS-induced activation of the IL-6/STAT3 signaling pathway in human
intestinal Caco-2 cells. Int J Mol Med. 35:1133–1137. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ke X, Chen J, Zhang X, Fang W, Yang C,
Peng J, Chen Y and Sferra TJ: Qing Hua Chang Yin attenuates
lipopolysaccharide-induced inflammatory response in human
intestinal cells by inhibiting NF-κB activation. Exp Ther Med.
6:189–193. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ke X, Zhou F, Gao Y, Xie B, Hu G, Fang W,
Peng J, Chen Y and Sferra TJ: Qing Hua Chang Yin exerts therapeutic
effects against ulcerative colitis through the inhibition of the
TLR4/NF-κB pathway. Int J Mol Med. 32:926–930. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Amasheh M, Grotjohann I, Amasheh S, Fromm
A, Söderholm JD, Zeitz M, Fromm M and Schulzke JD: Regulation of
mucosal structure and barrier function in rat colon exposed to
tumor necrosis factor alpha and interferon gamma in vitro: A novel
model for studying the pathomechanisms of inflammatory bowel
disease cytokines. Scand J Gastroenterol. 44:1226–1235. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Δ Δ C(T)) Method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Berends SE, Strik AS, Löwenberg M, D'Haens
GR and Mathôt RAA: Clinical Pharmacokinetic and Pharmacodynamic
Considerations in the Treatment of Ulcerative Colitis. Clin
Pharmacokinet. 58:15–37. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kondamudi PK, Malayandi R, Eaga C and
Aggarwal D: Drugs as causative agents and therapeutic agents in
inflammatory bowel disease. Acta Pharm Sin B. 3:289–296. 2013.
View Article : Google Scholar
|
|
29
|
Suzuki T: Regulation of intestinal
epithelial permeability by tight junctions. Cell Mol Life Sci.
70:631–659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ke X, Liu L, Zhao P, Chen Y, Peng J, Fang
W, Chen J, Hu G, Gao Y, Shen A, et al: The effects of Qing Hua
Chang Yin on the epithelial tight junctions of mice with
inflammatory bowel disease. Int J Clin Exp Med. 12:6864–6873.
2019.
|
|
31
|
Liu J, Lu XJ, Tian XM, Shen XP and Bao XP:
Antipyretic effects of Xinhuang Tablets on various animal fever
models. Drugs Clin. 4:375–379. 2015.(In Chinese).
|
|
32
|
Chen C, Bao XP, Qiu CX, Wang CF, Nan SH,
Huang WQ and Wang RG: Effects of Xinhuang tablets and its chinese
medicine components on high uric acid mice caused by hypoxanthine.
Strait Pharm J. 1:21–23. 2015.(In Chinese).
|
|
33
|
Putt KK, Pei R, White HM and Bolling BW:
Yogurt inhibits intestinal barrier dysfunction in Caco-2 cells by
increasing tight junctions. Food Funct. 8:406–414. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gasparetto M and Guariso G: Highlights in
IBD epidemiology and its natural history in the paediatric age.
Gastroenterol Res Pract. 2013:8290402013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Piche T, Barbara G, Aubert P, Bruley des
Varannes S, Dainese R, Nano JL, Cremon C, Stanghellini V, De
Giorgio R, Galmiche JP, et al: Impaired intestinal barrier
integrity in the colon of patients with irritable bowel syndrome:
Involvement of soluble mediators. Gut. 58:196–201. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhou Q, Zhang B and Verne GN: Intestinal
membrane permeability and hypersensitivity in the irritable bowel
syndrome. Pain. 146:41–46. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Camilleri M and Gorman H: Intestinal
permeability and irritable bowel syndrome. Neurogastroenterol
Motil. 19:545–552. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Van Itallie CM, Fanning AS, Bridges A and
Anderson JM: ZO-1 stabilizes the tight junction solute barrier
through coupling to the perijunctional cytoskeleton. Mol Biol Cell.
20:3930–3940. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fanning AS and Anderson JM: Zonula
occludens-1 and −2 are cytosolic scaffolds that regulate the
assembly of cellular junctions. Ann N Y Acad Sci. 1165:113–120.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hamada K, Shitara Y, Sekine S and Horie T:
Zonula Occludens-1 alterations and enhanced intestinal permeability
in methotrexate-treated rats. Cancer Chemother Pharmacol.
66:1031–1038. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Al-Sadi R, Khatib K, Guo S, Ye D, Youssef
M and Ma T: Occludin regulates macromolecule flux across the
intestinal epithelial tight junction barrier. Am J Physiol
Gastrointest Liver Physiol. 300:G1054–G1064. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zeissig S, Bürgel N, Günzel D, Richter J,
Mankertz J, Wahnschaffe U, Kroesen AJ, Zeitz M, Fromm M and
Schulzke JD: Changes in expression and distribution of claudin 2, 5
and 8 lead to discontinuous tight junctions and barrier dysfunction
in active Crohn's disease. Gut. 56:61–72. 2007. View Article : Google Scholar : PubMed/NCBI
|