Introduction
Prenatal ethanol exposure (PEE) is considered a
significant risk factor underlying several functional and
structural disorders, including stillbirth, spontaneous abortion,
premature birth, intrauterine growth retardation and low birth
weight, as well as developmental abnormalities of the central
nervous system (CNS), which are collectively referred to as fetal
alcohol spectrum disorders (1,2). These
deficits may result in lifelong behavioral abnormalities. In the
human fetus, PEE-induced abnormalities exhibit several
characteristics, including facial abnormalities, growth deficits,
lowered intelligence and microcephaly (3,4). In
several animal models, including mouse (5), rat (6), macaque (7,8),
zebrafish (9) and chick models
(10), PEE causes similar birth
defects and permanent neurodevelopmental disorders. Although
alcohol usage is preventable and a well-established hazard to the
fetus, the global consumption of alcohol during pregnancy is still
prevalent (9.8%, 95% CI 8.9–11.1) (1,2,11). As
previously reported, alcohol usage is an urgent public health issue
in China (12). A population-based
Chinese study reported that pregnant women in China had a high
prevalence rate of alcohol consumption (6.2% in rural and 10.3% in
urban areas), although women in China are discouraged from alcohol
consumption (13). Therefore, it is
important to investigate the mechanisms underlying the effects of
prenatal ethanol or alcohol exposure on brain disorders.
Microcephaly is one of the major developmental brain
defects associated with maternal ethanol exposure (14). PEE can result in a range of cortical
dysfunctions, consisting of abnormal neuronal migration (15), reduced cell viability (16), changes to cortical thickness
(17,18) and loss of neurons (8). Several factors are involved in the
process of embryonic cortex development, such as morphogens,
including fibroblast growth factor (19), bone morphogenetic protein 7
(20) and transforming growth
factor β1 (21,22), as well as Notch and Wnt signaling
(23,24), radial glial scaffolds for neuronal
migration (25,26) and cell-cell communications between
cortical neurons and cortical precursor cells (27,28).
Among these, several studies have revealed that alterations of
radial glial development contribute to the effects of alcohol on
the development of the cortex (29–31).
In vivo experimental results indicated that ethanol exposure
decreases glial fibrillary acidic protein (GFAP) mRNA expression
levels in the brains of pups from ethanol-fed Sprague Dawley or
Wistar rat mothers and impairs the morphology of radial glial cells
(RGCs) (29,31). In vitro studies also revealed
that ethanol could affect the distribution and content of the
radial glial cytoskeletal proteins GFAP, Vimentin and Nestin
(32). Thus, investigation of the
effects of prenatal alcohol exposure on radial glial fiber
proteins, such as Vimentin, Nestin and GFAP, may shed light on the
mechanisms involved in alcohol-induced cortical malformation.
Previous studies have reported that neurogenesis
occurs in two primary sites in the mammalian brain where the neural
stem cells (NSCs) reside: The subventricular zone (SVZ) of the
lateral ventricle and the subgranular zone (SGZ) of the hippocampal
dentate gyrus (33). RGCs derived
from neuroepithelial cells are ubiquitously present in the
developing brain from the start of fetal neurogenesis until their
final transformation into astrocytes in certain regions of the
brain (34,35). It has been confirmed that RGCs are
specialized cells that can generate both neurons and astrocytes
(34). In addition to the
generation of neurons and intermediate progenitors, RGCs also
provide radial glial scaffolding for neuronal migration via a long
radial process (36,37). As suggested by their name, RGCs have
two characteristics: A long radial process, which spans the entire
thickness of the wall of the neural tube and an (astro) glial
property, which is indicated by the expression of glial proteins,
including GFAP (38). Previous
studies observed that GFAP-expressing neural progenitor cells are
the major source of continuous neurogenesis (39–41).
RGCs are ultrastructurally similar to astrocytes with regards to
the expression of the filament protein GFAP and glycogen
accumulation (42). During early
cortical neurogenesis, GFAP expression is specifically found in
RGCs, where co-localization with Vimentin and Nestin is also
observed. However, GFAP positive cells exhibit a more
lineage-restricted phenotype (34).
Hence, the investigation of GFAP and the dynamic changes of its
major variants, such as GFAPα and GFAPδ, during the developmental
process may elucidate the mechanisms underlying how GFAP regulates
fetal cortical development.
In the present study, the developmental profile of
GFAPδ was investigated in the cortex in the control and
ethanol-exposed mice, with the aim of characterizing the roles of
GFAPδ in alcohol-induced cortex maldevelopment.
Materials and methods
Animals
Adult C57BL/6J mice (age, 8–12 weeks; mean body
weight, 24.0±1.8 g) were purchased from Hunan Silaike Jingda
Laboratory Animal Co., Ltd. A single male mouse was housed with
five female mice per standard polycarbonate cage. Mice were housed
in a temperature- and humidity-controlled environment (22°C; 50%
humidity) with a 12-h light/dark cycle. Before the initiation of
the experiments, the mice were maintained with ad libitum
access to standard laboratory chow and water. A total of 46 adults
(male 6 and female 40) and 215 fetal mice were used in the present
study. There were 14 dams and 102 fetuses in the ethanol (EtOH)
group, and 14 dams and 113 fetuses in the Control group. There were
6 dams in the EtOH group and 6 dams in the Control group that were
used specifically for a blood alcohol assessment. All animal
maintenance and experimental procedures were approved by the
Institutional Animal Care and Use Committee of Xi'an Medical
University and animal experiments were performed in accordance with
the National Institute of Health Guide for the Care and Use of
Laboratory Animals (NIH Publications no. 80-23) (43).
Animal maintenance and ethanol
acclimation
Before ethanol administration, the mice were
acclimated for ≥1 week after arrival. For the generation of the
prenatal alcohol exposure models, female mice were fed on a liquid
diet containing ethanol. The rodent liquid diet (Lieber-DeCarli
′82, Bio-Serv 14-726-629 and 14-726-630) was purchased from Thermo
Fisher Scientific, Inc. Control mice were maintained for the same
period on the same diet without ethanol, but instead supplemented
with maltodextrin to provide the appropriate amount of
ethanol-derived calories. For alcohol exposure, the female mice
were provided a Lieber-DeCarli liquid diet containing 2.4% (v/v)
ethanol for 2 days, then the ethanol concentration in the diet was
increased to 4.8% (v/v) and maintained for a further 14 days; 5%
sucrose to Lieber-DeCarli liquid diet was added to increase
palatability as described previously (44,45).
During alcohol exposure, the liquid diet was changed every day with
a freshly prepared diet. After the acclimation/acquisition period
for 16 days, the liquid diet was changed into normal laboratory
chow and water for an ethanol deprivation period, during which the
mice could recover from any potential alcohol withdrawal symptoms.
This ethanol deprivation period averaged 16 days. During this
ethanol deprivation period, the mice were bred by placing two
females in a cage with one male for 1 h during the dark cycle and
then were assessed for the presence of a copulation plug. If the
plug was present, the time at the initiation of the 1-h mating
period was considered as the gestational day (GD) 0. On GD7.0 at
the beginning of the dark cycle, the mice were provided a liquid
diet containing ethanol 4.8% (v/v, ethanol group) or a control
liquid diet. The control liquid diet, which contained an isocaloric
amount of maltodextrin instead of ethanol to meet the calories
derived from ethanol, served as the pair-fed control. Maternal
bodies were weighed, and diet consumptions were recorded based on
the previous 24 h change of liquid diet from 30-ml graduated
screw-cap tubes. Hence, the control and ethanol groups received the
same amount of diet (ml) based on the ml diet per g body weight
(g). This matching of calorific content and diet feeding was to
reduce the possible effects of any potential nutritional
differences (44). Subsequently,
the mice were maintained on this diet for 48 h through GDs 7 and 8
and returned to the standard laboratory chow and water until the
fetuses were collected on GD14.
Maternal blood alcohol assessment
A set of dams was used to assess blood alcohol
concentration (BAC). A total of 30 µl tail blood was collected into
heparinized capillary tubes from mice with sperm plugs on the 8th
day of pregnancy (at 21:00 and 23:00 h). BACs were measured using
an Analox alcohol analyzer (Model AM1; Analox Instruments USA,
Inc.). For each time point, ≥3 mice were sampled and analyzed, the
total samples of each mouse were controlled for no more than three
times. None of the fetuses from these animals were used for
histological analysis or western blot analysis.
Collection of the embryos
On the morning of GD 14.0, the embryos of pregnant
dams were collected after sacrifice using CO2 inhalation
according to the NIEHS euthanasia methods for rodent fetuses and
neonates (44). For CO2
asphyxiation, a clear acrylic chamber with a gasket lid (VWR
International, LLC) was used. The total chamber volume was 2 l (0.5
gal). The gas flow was set to 400 ml/min to achieve a 20% per min
volume displacement rate. The death of the pregnant female mice was
confirmed by the cessation of blood circulation (absence of
heartbeat) and the absence of a breathing and pedal reflex. Fetal
death was confirmed using ultrasound determination of the absence
of a heartbeat. Subsequently, embryos were isolated from the uterus
and extra-embryonic membranes under a dissecting microscope. The
embryos were immersed in ice-cold 0.1 M 1X PBS. Tissues were fixed
in 4% PFA in 1X Dulbecco's (D)PBS overnight at 4°C with gentle
shaking for immunohistochemistry. The dissected cortical tissues
were collected and pooled from each dam for western blot
analysis.
Protein extraction and western blot
analysis
For protein extraction, the cortical tissues from
mice embryos (n=5–10 samples per litter) were pooled and dissected,
meninges were removed under a stereomicroscope and dissolved in 1X
lysis buffer (Cell Signaling Technology, Inc.; cat. no. 9803)
supplemented with 1 mM PMSF. Subsequently, the tissues were
mechanically homogenized with a tissue homogenizer and incubated on
ice for 15 min, followed by centrifugation at 12,000 × g for 15 min
at 4°C to obtain the cleared supernatants. The supernatants were
collected, and protein concentrations were measured using a Pierce™
BCA Protein assay kit (Thermo Fisher Scientific, Inc.; cat. no.
23225). The samples were then adjusted and dissolved in 2X Laemmli
Sample buffer (Bio-Rad Laboratories, Inc.; cat. no. 1610737).
Subsequently, the samples were boiled at 95°C for 5 min, loaded on
a 12% SDS-gel and resolved using SDS-PAGE; a BLUEstain™ Protein
ladder (Goldbio; cat. no. P007-1500) was used to determine the
molecular weight. The gels were blotted on PVDF membranes
(Immobilon; EMD Millipore) at 200 mA for 1.5 h at 4°C. A total of
15 µg protein lysate was loaded into each well of the 12% SDS gel.
After transferring and washing three times with PBS with 0.1%
Tween® detergent (PBST), 5 min per wash, the membranes
were blocked using 3% bovine serum albumin (BSA; cat. no. ab64009,
Abcam) in 1X PBST for 1 h at room temperature. Primary antibodies
were diluted in 3% BSA-PBST buffer and incubated at 4°C on a
shaker. After three washes (10 min each), the secondary horseradish
peroxidase-conjugated anti-rabbit (1:2,000; cat. no. ab6721; Abcam)
or anti-mouse antibodies (1:2,000; cat. no. ab6789; Abcam) were
incubated with the membrane at room temperature for 1 h. After
three washes (10 min each), the membranes were developed using
SuperSignal™ West Femto Maximum Sensitivity substrate (Thermo
Fisher Scientific, Inc.; cat. no. 34096) and imaged using Image
Lab™ Software Version 6.0.1 (Bio-Rad Laboratories, Inc.). For
normalization, β-actin was used as the loading control. The
following antibodies were used: Mouse anti-GFAP antibody
(Sigma-Aldrich; Merck KGaA; cat. no. G3893; 1:2,000), rabbit
anti-GFAPδ antibody (Abcam; cat. no. ab93251; 1:2,000) or β-actin
antibody (Abcam; cat. no. ab8227; 1:5,000). Densitometry analysis
was performed using ImageJ version 1.6.0 (National Institutes of
Health).
Embryo collections, fixation,
sectioning and immunohistochemistry
On E14.5, control and ethanol-exposed mice were
euthanized by CO2 inhalation, and cervical dissociation
was performed. The maternal abdomen was opened and the fetuses were
removed from the uterus. The number of fetuses was recorded and
imaged for each litter using a digital camera (Micropublisher 5.0;
Qimaging Corp.), which was mounted on an Olympus binocular
dissecting microscope (Olympus America, Inc.).
For the histological analysis, the fetuses were
collected from individual extraembryonic membranes and fixed in 4%
paraformaldehyde (PFA, cat. no. 158127, Sigma-Aldrich; Merck KGaA)
in 1XDPBS overnight at 4°C. After immersion in 30% sucrose in 1X
DPBS, 20-µm-thick coronal sections were cut on a cryostat microtome
(Leica CM3050 S; Leica Microsystems GmbH). The sections were washed
with PBS three times (each for 10 min), then blocked using 10%
donkey or goat serum (Abcam) in DPBS supplemented with 0.25% Triton
X-100 (Sigma-Aldrich; Merck KGaA) for 2 h at room temperature.
Sections were subsequently incubated with primary antibodies at 4°C
overnight. The following antibodies were used: Rabbit anti-SOX2
(Cell Signalling Technology, Inc.; cat. no. 23064; 1:200), mouse
anti-Glial Fibrillary Acidic Protein (GFAP) antibody (Sigma; cat.
no. G3893, 1:200), Mouse Anti-SOX2 antibody [9-9-3] (1:200; cat.
no. ab79351; Abcam), rabbit anti-GFAPδ antibody (1:200; cat. no.
ab93251; Abcam), mouse anti-Hop antibody (E-1) (HOPX; Santa Cruz
Biotechnology, Inc.; cat. no. sc-30216; 1:200), rabbit anti-Nestin
antibody (1:200 cat. no. ab92391; Abcam), rabbit anti-Vimentin
antibody (1:200; cat. no. ab92547; Abcam), mouse anti-S100β
(β-Subunit; Sigma-Aldrich; Merck KGaA; cat. no. S2532; 1:200),
mouse anti-Ki67 antibody (Sigma-Aldrich; Merck KGaA; cat. no.
MAB4190; 1:200) mouse anti-phospho-histone H3 (Ser10; 6G3) (pHH3;
Cell Signaling Technology, Inc.; cat. no. 9706; 1:200), Rat
anti-Ctip2 (1:200; cat. no. ab18465; Abcam). After washing three
times with 1X DPBS, the fluorophore-conjugated secondary antibodies
were used at a dilution of 1:200 and incubated for 2 h at room
temperature. The secondary antibodies used were all purchased from
Thermo Fisher Scientific, Inc. and were: Alexa Fluor 488 goat
anti-rabbit IgG (H+L) Cross-Adsorbed Secondary antibody (cat. no.
A11008) and Alexa Fluor 594 donkey anti-mouse IgG (H+L) Highly
Cross-Adsorbed Secondary antibody (cat. no. A21203). Cell nuclei
were counterstained at room temperature for 10 min with DAPI
(Sigma-Aldrich; Merck KGaA; cat. no. D9542; 1:2,000). Subsequently,
the slides were sealed and images were captured using a Nikon A1R
confocal microscope (Nikon Corporation) with ×200 magnification
with the appropriate laser channels. Images were processed using
ImageJ 1.50i (National Institutes of Health). The ventricular zone
(VZ), SVZ, intermediate zone (IZ), subcortical plate (SP) and
cortical plate (CP) layers were delineated using DAPI staining.
Quantification of the proliferative or
mitotic GFAPδ+ RGCs
The number of Ki67+ GFAPδ+ or
pHH3+ GFAPδ+ double-labeled cells, as well as
total GFAPδ+ RGCs, were quantified using ImageJ 1.50i
with the co-localization finder plugin (46). For each brain region, six
representative coronal sections were acquired using the Nikon A1R
confocal microscope with ×200 magnification. Of these images, three
images were captured at the level of the ventricular surface,
including the VZ and the inner SVZ. The other three images were
captured above the initial images, including outer the SVZ, IZ and
CP. Hence, cells were counted in ~6 images per brain region (6
images per region × 2 brain regions per section × 4 sections × 10
embryonic brains). Furthermore, the ratio of Ki67+
GFAPδ+ or pHH3+ GFAPδ+ cells to
total GFAPδ+ cells was calculated.
Statistical analysis
For quantification of immunostaining, the images
were processed in parallel and were captured using the same
settings and laser power for confocal microscopy. The cell numbers
were manually counted using the ImageJ 1.50i cell counter plugin
(National Institutes of Health). The fluorescence intensity was
measured using ImageJ 1.51i and normalized to the size of the
aggregates. All quantitative results are presented as the mean ±
SEM. To determine the statistical difference, an unpaired Student's
t-test was performed. P<0.05 was considered to indicate a
statistically significant difference. Experiments were repeated at
least three times.
Results
Ethanol intake and blood concentration
measurements
The aforementioned ethanol exposure protocol
resulted in an average ethanol intake on GD8 of 25.1 g/kg/day. This
mean amount of ethanol intake resulted in BACs of 168–216 mg/dl
(mean; 178.4 mg/dl).
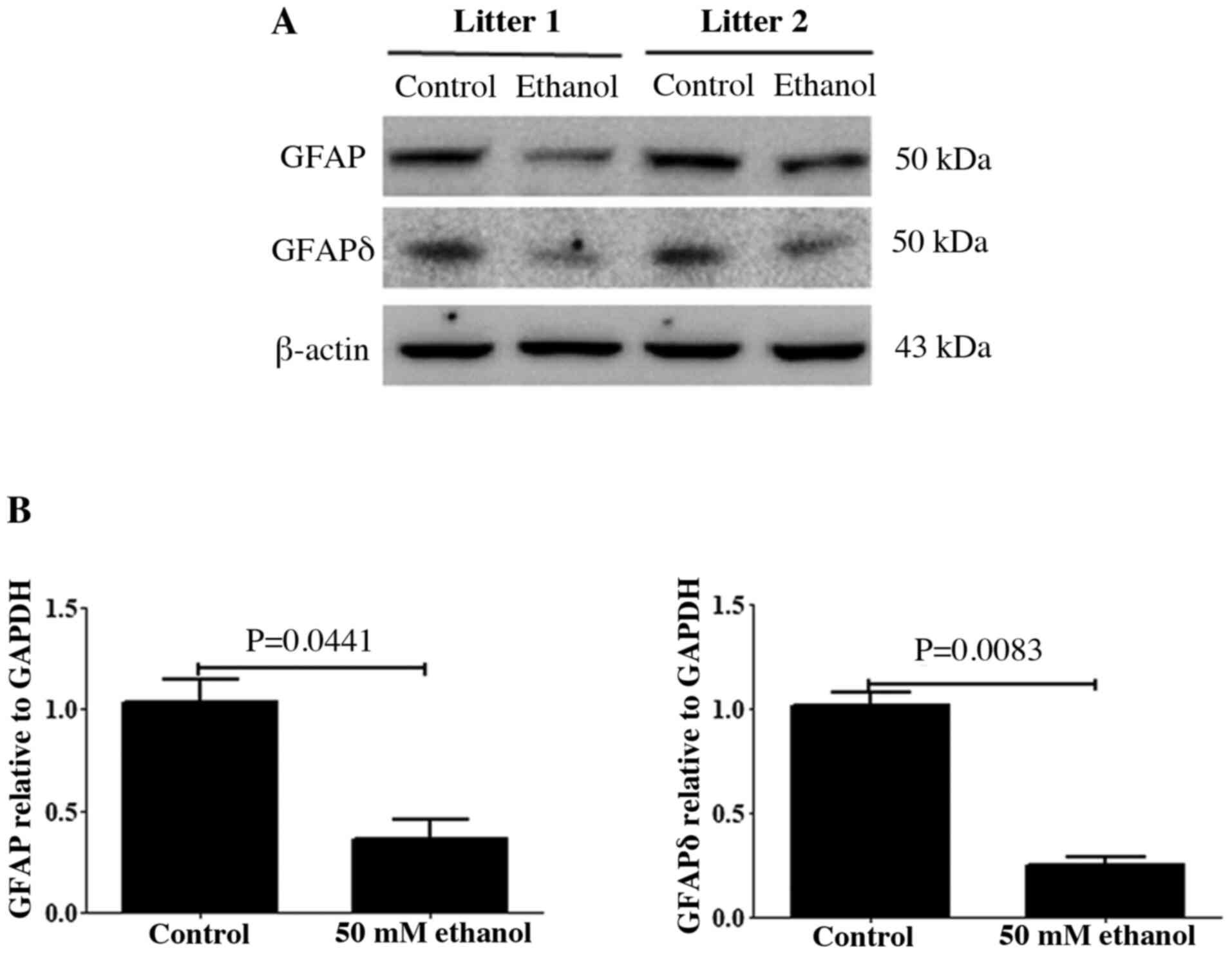
Western blot analysis of GFAP and
GFAPδ expression levels in the cortex
GFAP and its variant GFAPδ are two major components
of radial glial fibers, and both proteins are expressed in RGCs
(41). The relative expression
levels of GFAP and GFAPδ in the control and PEE brain on E14.5 were
analyzed using western blotting. As presented in Fig. 1A, immunoblots of GFAP and GFAPδ
showed a single band of ~50 kDa, which corresponded to the
molecular mass of GFAP and GFAPδ. After PEE, the relative densities
of GFAP and GFAPδ were significantly decreased compared with those
of the controls (Fig. 1B).
Effects of PEE on the expression of
GFAP in RGCs
To assess the effects of ethanol exposure on the
distribution of GFAP, cortical sections were fixed and
co-immunostained for SOX2 and GFAP using the corresponding
antibodies. The results demonstrated that SOX2+ cells
were predominantly present in the VZ and SVZ, where almost all
cells were SOX2+, and only a few SOX2+ cells
in the IZ were present (Fig. 2).
Moreover, few cells were SOX2+ in the SP/CP.
SOX2+ cells also co-express GFAP. At E14.5,
GFAP+ cells were observed in all layers. Some
ventricular GFAP+ cells showed the characteristic
morphologies of RGCs with an endfoot contacting the ventricular
surface, a cell body within the ventricular zone and a long basal
process extending toward the pia. Following ethanol treatment, the
fluorescence intensities of both SOX2 and GFAP were significantly
decreased (Fig. 2B and C),
suggesting that ethanol exposure decreased the stem cell potential
of RGCs.
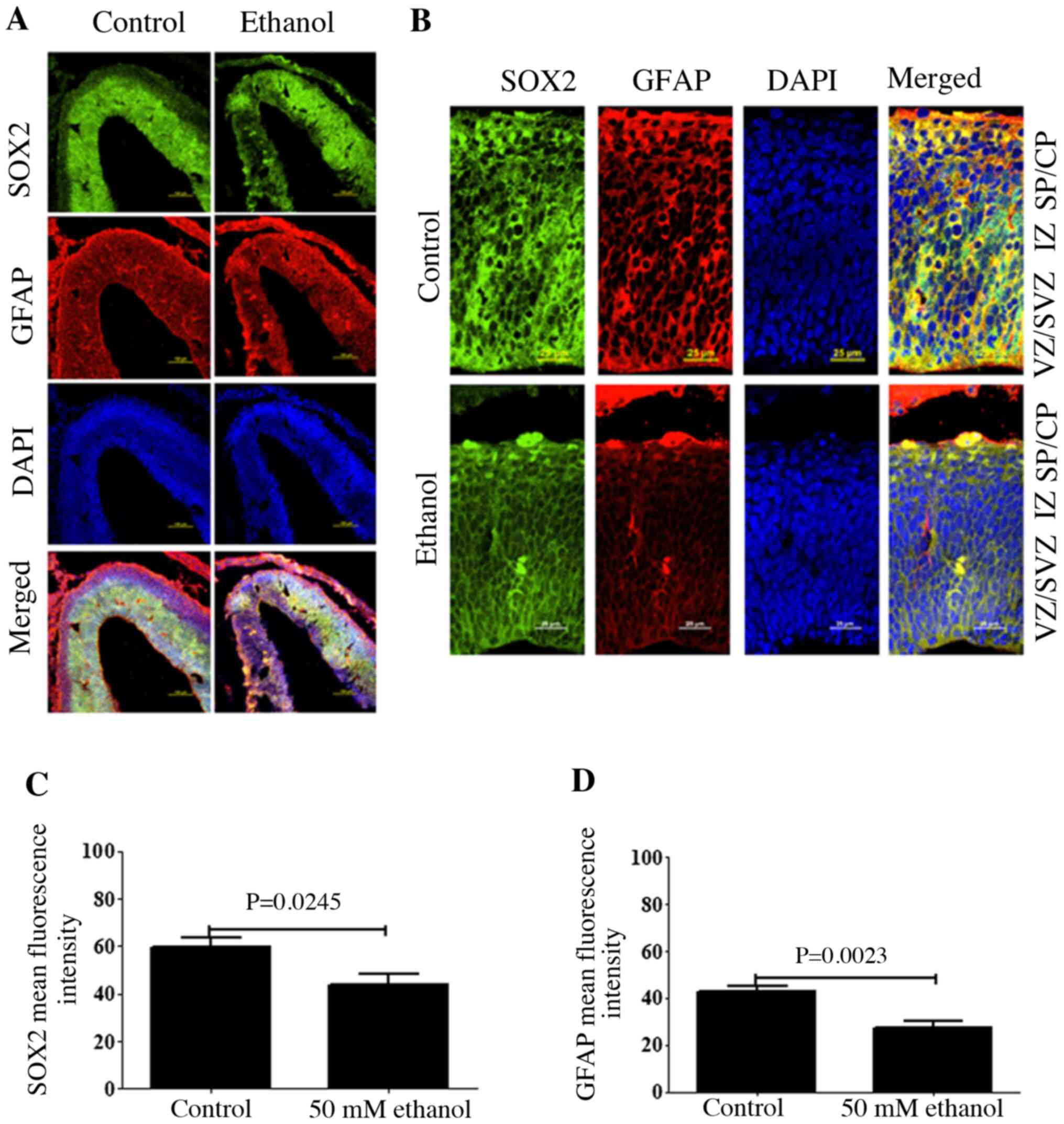 | Figure 2.Effects of PEE on the expression
level of GFAP in radial glial cells. (A) Coronal sections of
control and PEE treated brain was stained with DAPI (blue), SOX2
(green), and GFAP (red). Immunostaining revealed the distributions
of SOX2-positive neural progenitors and GFAP-positive cells in
control and PEE-treated cortices. (B) The distribution of SOX2 and
GFAP in cortical layers. Serial sections of embryonic cortices were
double immunostained with the antibodies against SOX2 (green) and
GFAP (red). Images indicated that GFAP was expressed in the whole
embryonic cortex layers and colocalized with SOX2 in control group.
PEE decreased the colocalization of SOX2 and GFAP. Quantification
of mean fluorescence intensity of (C) SOX2 and (D) GFAP. PEE
significantly decreased the numbers of
SOX2+GFAP+ radial glial cells. Scale bars,
100 µm (A) and 25 µm (B). The P-value was obtained from an unpaired
Student's t-test. PEE, prenatal ethanol exposure; GFAP, glial
fibrillary acidic protein; SP, subcortical plate; CP, cortical
plate; IZ, intermediate zone; VZ, ventricular zone; SVZ,
subventricular zone. |
Effect of PEE on the expression of
GFAPδ in RGCs
As GFAPδ expression was specifically limited to RGCs
in the neurogenic regions, whether GFAPδ was co-expressed with the
well-defined neural stem cell marker SOX2 and outer RGC (oRG)
marker, HOPX expression was next determined. To confirmed the
specificity of GFAPδ, GFAPδ and CTIP2 were co-immunostained and it
was found that GFAPδ was not expressed in CTIP2-positive neurons
(Fig. S1). The results
demonstrated that GFAPδ had similar expression patterns with SOX2
and HOPX (Fig. 3). As a marker of
oRG, HOPX was predominantly expressed in the VZ, SVZ and CP as
indicated by the arrow in Fig. 3C.
Double staining identified that GFAPδ expression strongly
overlapped with both SOX2 and HOPX. Furthermore, the quantitative
data indicated that ethanol treatment could significantly decrease
the fluorescence intensity of GFAPδ and HOPX (Fig. 3B and D), suggesting that PEE could
disrupt the assembly of oRG fibers.
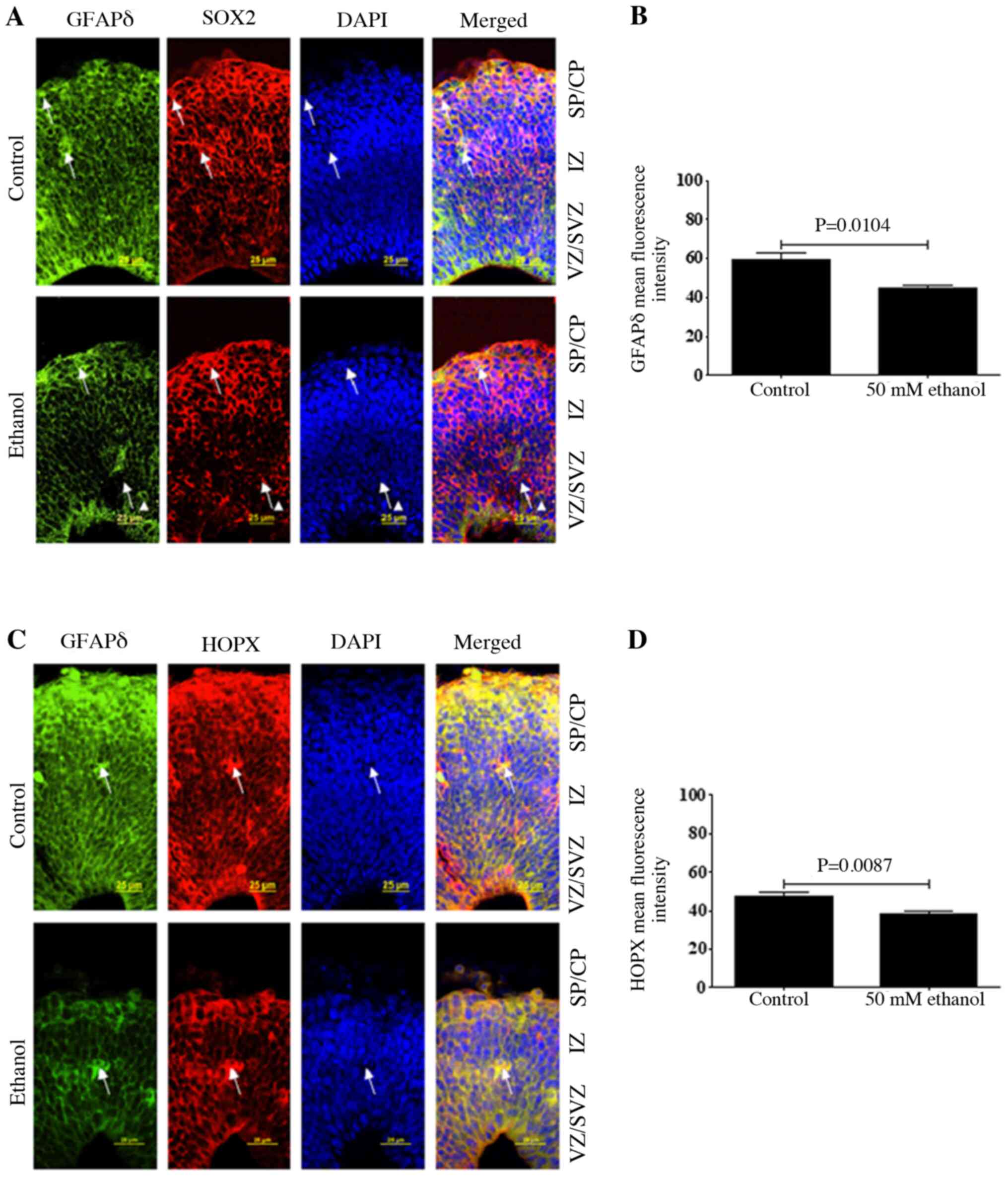 | Figure 3.Effects of PEE on the expression
level of GFAPδ in radial glial cells. Serial sections of embryonic
cortices were subjected to the antibodies against GFAPδ and (A)
radial glial marker SOX2 or (C) outer radial glial marker HOPX. At
embryonic day 14.5, cells with overlapping expression levels of
GFAPδ and SOX2 were shown in yellow. The nuclear stain was in blue.
The data indicated that GFAPδ+ cells were also
co-labeling with SOX2. Arrows pointed to representative
SOX2+ cells with long process labeling with GFAPδ. In
control mice, double labeling of GFAPδ with HOPX suggested that
GFAPδ can also be expressed in outer radial glial cells. PEE
decreases the intensity of GFAPδ in (B) SOX2 and (D) HOPX positive
cells as shown in the right panels, indicating that PEE decreases
the stem cell potential of radial glial cells. Scale bars, 25 µm.
The P-value was obtained from an unpaired Student's t-test. PEE,
prenatal ethanol exposure; GFAP, glial fibrillary acidic protein;
SP, subcortical plate; CP, cortical plate; IZ, intermediate zone;
VZ, ventricular zone; SVZ, subventricular zone; HOPX, Hop antibody
(E-1). |
Effect of PEE on co-localization of
GFAPδ with Nestin or Vimentin in RGCs
To determine whether PEE could impair other
intermediate filament proteins, the expression distribution of
Nestin and Vimentin was also assessed. The results demonstrated
that Nestin was expressed on both the apical and basal surface
processes of the RGCs, and its expression was co-localized with
GFAPδ and surrounded the nuclei, similar to GFAPδ. Ethanol
treatment significantly decreased both Nestin and GFAPδ expression
(Fig. 4A and B). In the SVZ and IZ
layers, Nestin expression, based on fluorescence intensity, was
significantly decreased following ethanol exposure (Fig. 4B). Similar to Nestin, Vimentin
expression was also co-localized with GFAPδ in the apical and basal
surfaces of the process from the RGCs (Fig. 4C). Several RGCs in VZ showed strong
Vimentin-positive apical processes. The majority of
Vimentin-immunostaining encircled the nuclei of the cells and
labeled the fibers of RGCs. At E14.5, Vimentin, similar to GFAPδ,
was observed across almost the entire cortical wall. Compared with
the staining of GFAPδ around the cell nucleus, Vimentin was more
concentrated in the process of the RGCs. Ethanol exposure decreased
the expression of Vimentin, as indicated by the presence of
GFAPδ+ Vimentin− cells (Fig. 4C). Although ethanol exposure
resulted in a slight decrease in the fluorescence intensity of
Vimentin, there was no significant difference compared with control
(Fig. 4C).
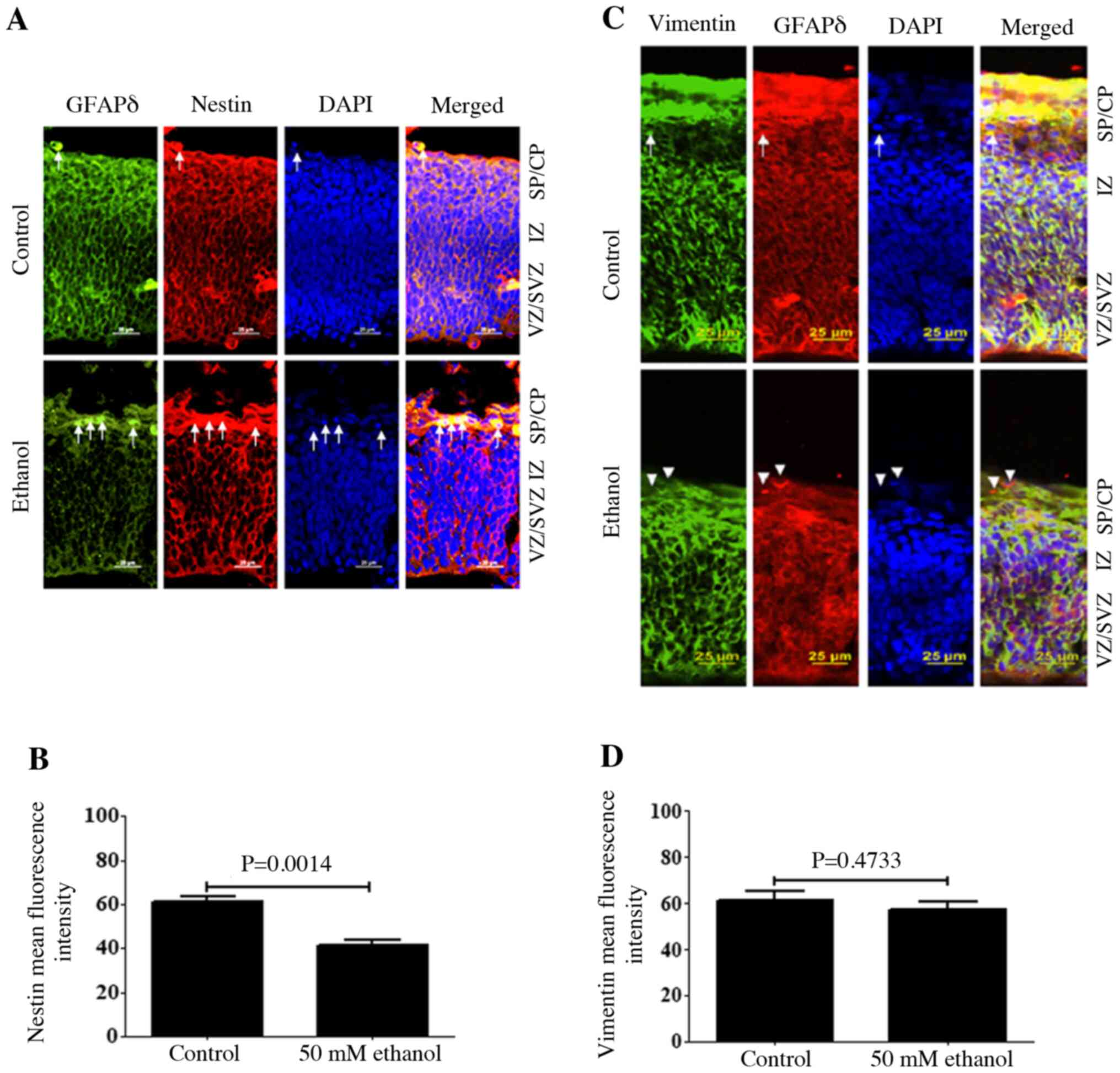 | Figure 4.Effects of PEE on the
co-immunostaining of Nestin or Vimentin in GFAPδ+ cells.
Serial sections of embryonic cortices were subjected to the
antibodies against GFAPδ and intermediate filament protein (A)
Nestin or (C) Vimentin. At embryonic day 14.5, cells with
overlapping expression of GFAPδ and Nestin or Vimentin were shown
in yellow. The nuclear stain was blue. The present data indicated
that Nestin immunostaining had similar distribution as GFAPδ, which
was surrounding the nuclear. In control mice, double
immunostainings of GFAPδ and Nestin or Vimentin indicated
colocalization of two immunoreactivities in most of the cells, as
indicated by arrows. Quantification of mean fluorescence intensity
of (B) Nestin and (D) Vimentin. PEE promoted the early appearance
of GFAPδ+ astrocytes in the SP, as shown by single GFAPδ
staining indicated by white arrowheads. It was indicated that PEE
impaired the intermediate filamentous components of radial glial
fibers and promoted the terminal differentiation of radial glial
cells. Scale bars, 25 µm. The P-value was obtained from an unpaired
Student's t-test. PEE, prenatal ethanol exposure; GFAPδ, glial
fibrillary acidic protein δ; SP, subcortical plate; CP, cortical
plate; IZ, intermediate zone; VZ, ventricular zone; SVZ,
subventricular zone. |
PEE decreases GFAP and GFAPδ protein
expression levels in RGCs and promotes their maturation
According to previous reports, GFAP is a defined
marker protein of astrocytes (40,47).
To determine whether GFAPδ expression was similar to the GFAP
expression in the developing cortex, double staining of GFAPδ
together with GFAP was performed. The results demonstrated that
both GFAP and GFAPδ were detected in the cortex at E14.5. These
exhibited similar expression patterns in the developing VZ and SVZ,
as well as the superficial layers, IZ and CP. GFAPδ-positive cells
had a bipolar morphology with the soma in the VZ, short processes
toward the ventricles and longer processes extending toward the CP
(Fig. 5). Moreover, GFAPδ filaments
were tangentially oriented, exhibiting unipolar or bipolar
morphologies. From the staining, it was identified that GFAPδ
filaments localized to the cell body and peri-nuclear regions.
Similarly, GFAP was also observed both in the somata and processes
of RGCs, which overlapped with the pattern of GFAPδ expression.
Both GFAP and GFAPδ could be detected in the superficial layers at
E14.5. Some GFAP+ RGCs exhibited only the basal
processes, resembling basal RGCs of the neocortex (arrow; Fig. 5A). After ethanol treatment, the
expression levels of both GFAP and GFAPδ were significantly
decreased. In the SVZ region after ethanol exposure, there were
some GFAP+ GFAPδ+ cells without apical and
basal processes, and these would have eventually become astrocytes.
Apical radial glia in the VZ had notable
GFAP+GFAPδ+ staining compared with that of
the control, which suggested ethanol treatment significantly
influenced the formation of radial glial fibers. Thus, altering the
expression of GFAP and GFAPδ may be how ethanol consumption during
fetal cortical development exerts its detrimental effects.
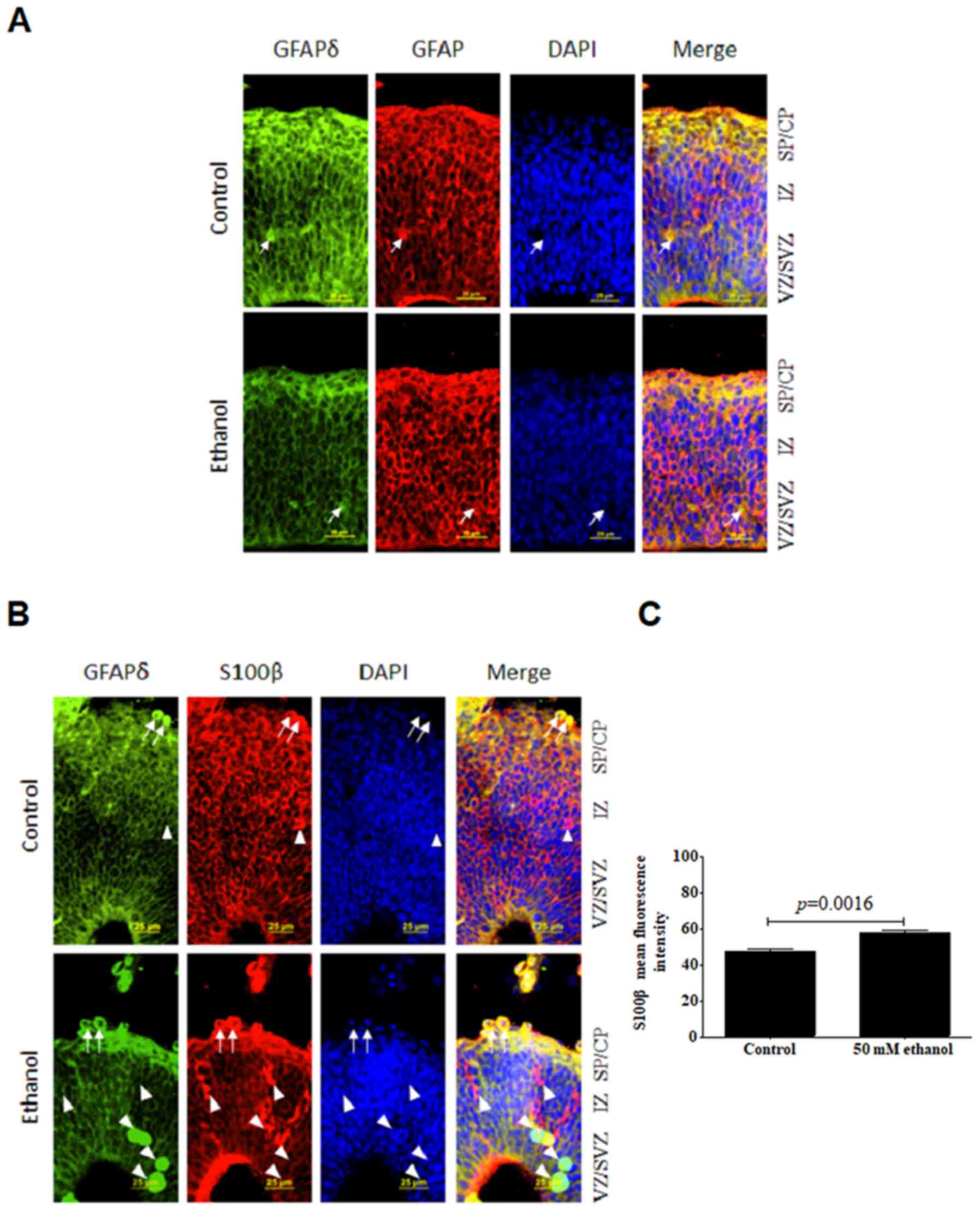 | Figure 5.Effects of PEE on the
co-immunostaining of GFAPδ and GFAP or S100β in radial glial cells.
Serial sections of embryonic cortices were double immunostained
with the antibodies against GFAPδ and GFAP or S100β. At embryonic
day 14.5, cells with overlapping expression of GFAPδ and (A) GFAP
or (B) S100β are shown in yellow. The nuclear stain is blue. The
results suggested that GFAP+ cells exhibited long
processes in control group. In control mice, double labelling of
GFAPδ and GFAP or S100β revealed the colocalization of the two
immunoreactivities. PEE notably decreased the expression of GFAPδ
in GFAP-positive cells and increased the expression of S100β in
GFAPδ-positive cells. Arrows indicated cells that have double
staining of GFAPδ and GFAP or S100β in both the control and PEE
cortex. Arrowheads showed cells had only express S100β or GFAPδ. It
was indicated that PEE enhanced the maturation of GFAPδ+
radial glial cells. (C) Quantification of mean fluorescence
intensity of S100β in control and ethanol groups. Scale bar, 25 µm.
The P-value was obtained from an unpaired Student's t-test. PEE,
prenatal ethanol exposure; GFAP, glial fibrillary acidic protein;
SP, subcortical plate; CP, cortical plate; IZ, intermediate zone;
VZ, ventricular zone; SVZ, subventricular zone. |
It was hypothesized that loss of GFAP and GFAPδ may
promote terminal differentiation of RGCs. S100β can be considered
as a marker of the progression of neural progenitors. To evaluate
whether GFAPδ could colocalize with S100β, double staining of GFAPδ
and S100β was performed. Interestingly, S100β exhibited similar
distribution patterns through all the layers of cortex with that of
GFAPδ (Fig. 5B), and its expression
was localized to the perinuclear area. The ethanol-exposed brain
showed a significantly increased intensity of cytoplasmic staining
compared with the control (Fig.
5C). However, there were more concentrated particles in RGCs in
the ethanol group. Unlike GFAP immunostaining, S100β is a protein
that can reliably define the maturation stage of GFAP+
cells (48). Increased expression
of S100β in the developmental RGCs indicated the loss of stem cell
differentiation potential by ethanol exposure. Quantification
analysis demonstrated that ethanol resulted in a significant
upregulation of S100β in the VZ and CP. Thus, it was suggested that
GFAPδ+ RGCs gradually lost their stem cell potential and
became more restricted following ethanol treatment.
Effects of PEE on the proliferative or
mitotic abilities of GFAPδ-positive cells
To determine whether GFAPδ positive cells were
proliferative cells or dividing cells, the colocalization of GFAPδ
with Ki67 (Fig. 6A and B) or pHH3
(Fig. 7A and B) was detected. The
results demonstrated that GFAPδ-expressing cells expressed the
proliferation marker Ki67 or mitotic marker pHH3 in both the
control and ethanol groups. PEE remarkably inhibited the
proliferative and mitotic activities of GFAPδ-positive cells
compared with that of the control group. Furthermore,
GFAPδ+ proliferative cells were distributed in all the
layers. The proliferative cells had both basal and apical processes
in the VZ region, suggesting these cells were apical RGCs. In the
SVZ, the Ki67+ proliferative cells had their basal
processes extending to the CP, which is one of the characteristic
morphologies of oRG cells (Fig.
6A). Hence, the GFAPδ+Ki67+ cells were
considered active RGCs.
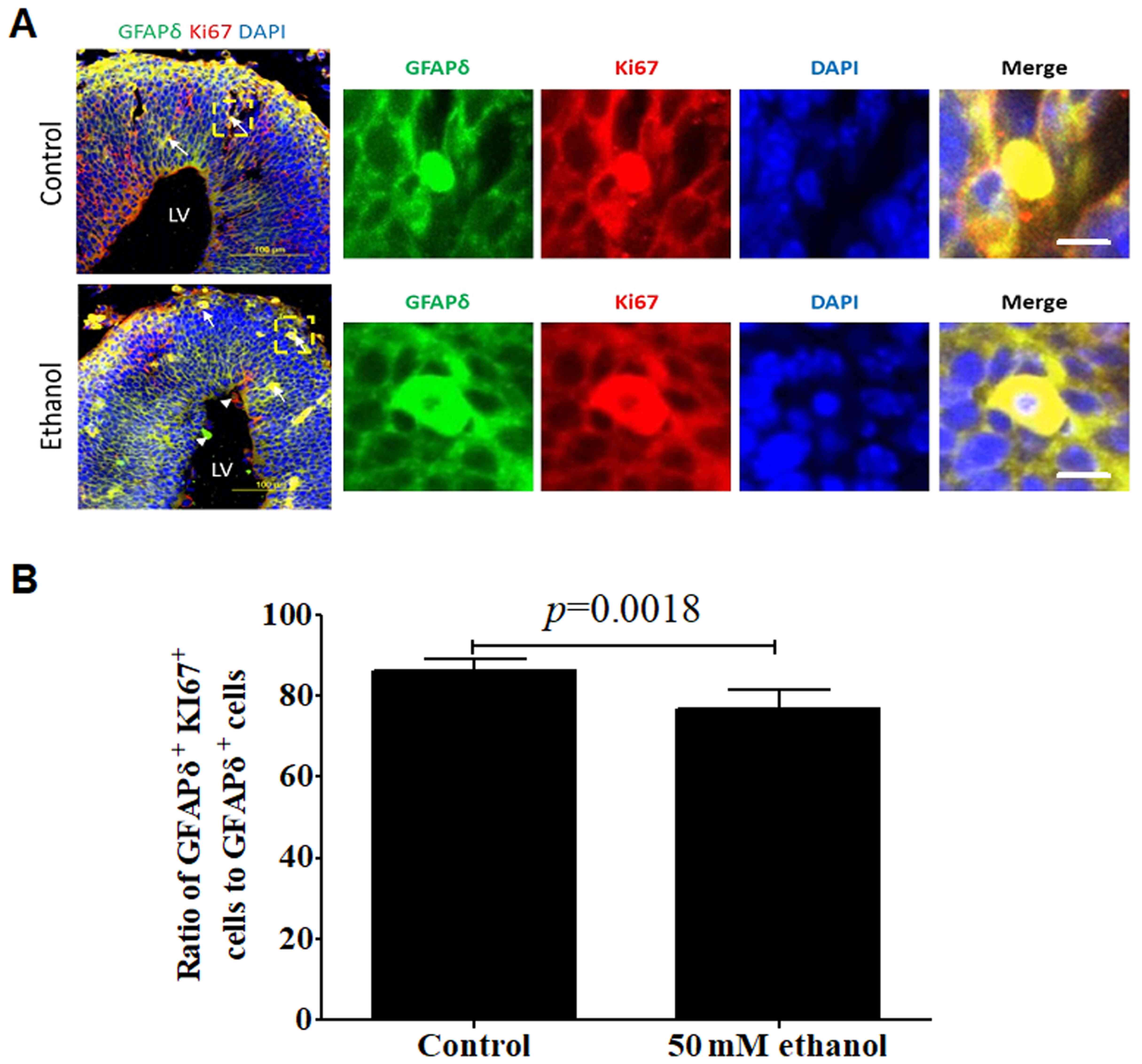 | Figure 6.Effect of PEE on the proliferative
activities in GFAPδ+ radial glial cells. (A) In
embryonic day 14.5 cortices, co-labeling for GFAPδ and Ki67
demonstrated that GFAPδ+ cells were proliferating RGCs
in the VZ, SVZ, IZ or CP. PEE significantly influenced the
distribution of Ki67+ cells. In the ethanol cortex,
radial glial cells were less abundant for
GFAPδ+Ki67+ cells. The arrows indicate
examples of GFAPδ+Ki67+ proliferative cells
located in the control and ethanol-treated cortex. Arrowheads show
the single labeling cells by GFAPδ or Ki67 in the ethanol-treated
embryonic cortex. Immunostainings for representative
GFAPδ+Ki67+ proliferative cells in control
and ethanol groups are showed in the right magnification images.
(B) Quantification of the ratio of
GFAPδ+Ki67+ positive cells to total
GFAPδ+ cells in the cortical layers. Scale bar, 100 µm
(Left image) and 10 µm (right magnification images). The P-value
was obtained from an unpaired Student's t-test. PEE, prenatal
ethanol exposure; GFAP, glial fibrillary acidic protein; SP,
subcortical plate; CP, cortical plate; IZ, intermediate zone; VZ,
ventricular zone; SVZ, subventricular zone; LV, lateral
ventricle. |
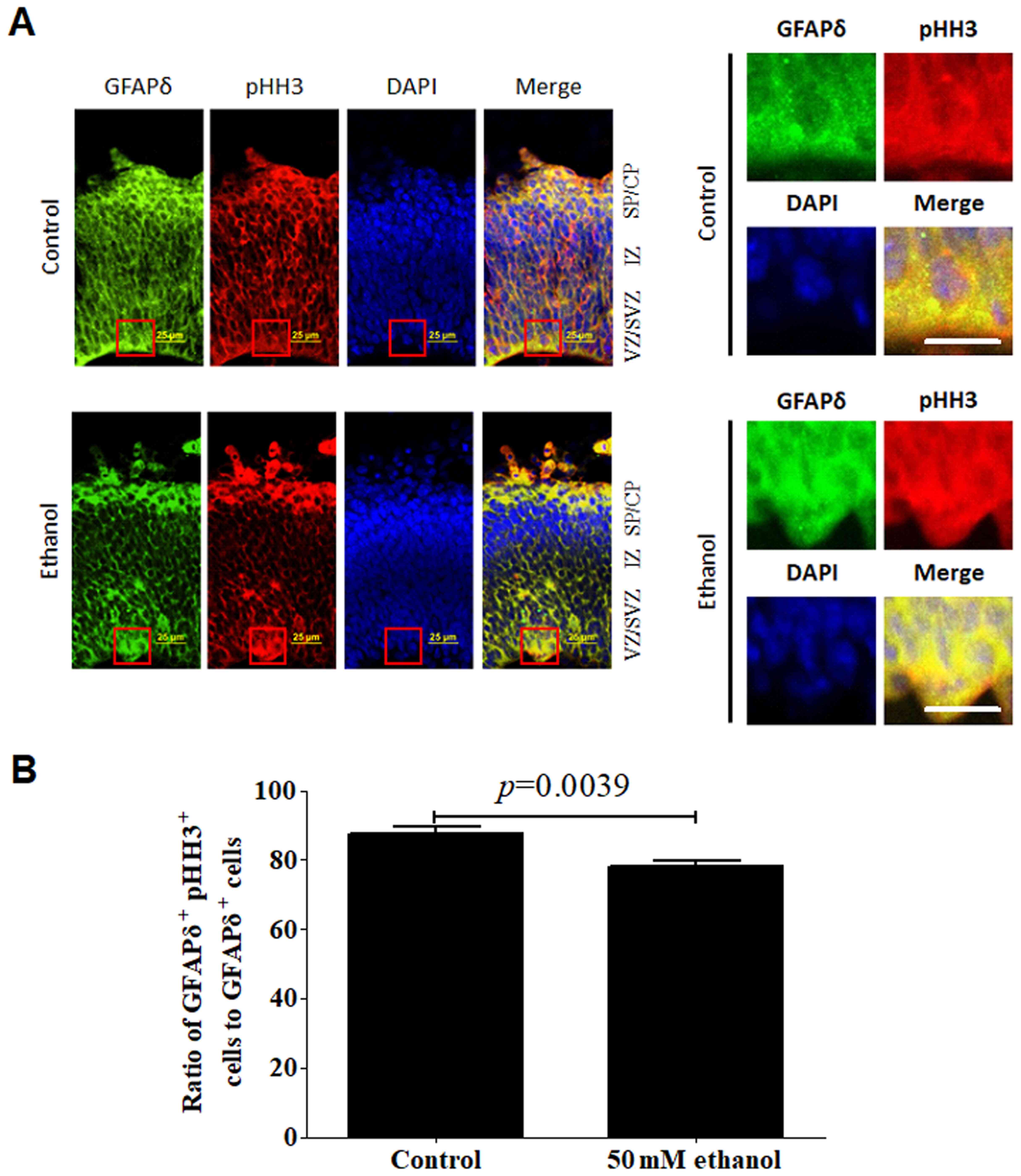 | Figure 7.Effect of PEE on the mitotic
activities in GFAPδ+ radial glial cells. (A) In
embryonic day 14.5 cortices, co-labeling for GFAPδ and pHH3
indicated that GFAPδ+ cells were mitotic RGCs in the VZ,
SVZ, IZ or CP. (B) PEE significantly influenced the distribution of
GFAPδ+ pHH3+ cells. In the ethanol cortex,
radial glial cells were less abundant for
GFAPδ+pHH3+ cells, especially in SVZ and IZ
layers. The representative mitotic cells by co-immunostaining of
GFAPδ and pHH3 in the control and ethanol-treated cortex are showed
in the right magnified images. Scale bar, 25 µm (left image) and 20
µm (right magnification images). The P-value was obtained from an
unpaired Student's t-test. PEE, prenatal ethanol exposure; GFAP,
glial fibrillary acidic protein; SP, subcortical plate; CP,
cortical plate; IZ, intermediate zone; VZ, ventricular zone; SVZ,
subventricular zone; LV, lateral ventricle; pHH3, phospho-histone
H3. |
Regarding the co-expression of GFAPδ and pHH3, most
of the GFAPδ+ cells exhibited active mitotic activities.
pHH3+ mitotic cells were distributed in almost all the
layers including the VZ, SVZ and CP. Ethanol treatment
significantly affected the colocalization of GFAPδ and pHH3,
particularly in the SVZ region (Fig.
7A). Following alcohol treatment, the ratio of co-localization
of GFAPδ+Ki67+ or
GFAPδ+pHH3+ to total GFAPδ+ cells
was significantly decreased (Figs.
6B and 7B).
Discussion
During the developmental process of the dorsal
telencephalon, RGCs and intermediate progenitor cells are the
primary sources of excitatory neurons (49). SOX2 is a Sry-related HMG-box-type
transcription factor, which is important for the maintenance of
stem cell differentiation potential (50). SOX2 is a well-known and key
regulator of cell fate decisions during neurogenesis, and it is
important for the maintenance of the proliferative potential and
the supervision of the generation of subpopulations of cells with
varying phenotypes in the developing neocortex (51). A high expression level of SOX2
autonomously maintains stem cell potential, inhibits neuronal
differentiation and maintains progenitor characteristics (49). Moreover, inhibition of SOX2 results
in a loss of expression of progenitor markers and enhances early
neuronal differentiation (50).
Using conditional deletion, it has been revealed that SOX2 deletion
could significantly influence the embryonic development of the
ventral telencephalon (52). Thus,
SOX2 is a key regulator between the stem cell and the dividing
progenitors (53,54). It was previously confirmed that SOX2
is specifically expressed in neural epithelial cells and RGCs, but
not in intermediate progenitor cells (49), thus it is considered a radial glial
marker protein. Following the process of differentiation, SOX2
expression is decreased during the final fate decision of stem
cells. During cortical development, SOX2 can be expressed together
with GFAP in RGCs within the neurogenic regions SVZ and SGZ
(39). In the present study, the
findings demonstrated that prenatal ethanol exposure resulted in a
significant decrease in both the number of SOX2+ cells
and the intensity of GFAPδ and GFAP expression in SOX2+
cells.
Radial glial fibers are highly dynamic and are cell
specific, providing the scaffold for neuroblast migration during
normal neurodevelopment (25,26).
Radial glial fibers promote the postnatal
ventricular-subventricular zone (v-svz)-derived neuroblast to
migrate toward to the lesion sites (36). GFAP is one of the major intermediate
filament proteins in RGCs and astrocytes, which can colocalize with
several other intermediate filaments, such as Vimentin and Nestin
(41,55). Previous studies have confirmed that
increased expression of intermediate filament proteins is an
important step in the formation of reactive astrocytes (47,56,57).
Dividing GFAP-expressing cells with bipolar or unipolar
morphologies are considered RGCs in the human forebrain (58). In rodent models, GFAP expression is
only detected in the astrocytic lineage after neurogenesis has
ended and Vimentin expression is no longer detectable (58). GFAP is a member of class III
intermediate filament proteins. There are 10 different transcripts
of the GFAP gene and these can generate 10 differential splicing
variants: GFAPα, GFAPβ, GFAPγ, GFAPδ, GFAPκ, GFAPζ, GFAPD135,
GFAPD164, GFAPDexon6 and GFAPDexon7 (56,59).
All these variants are similar, and the differences primarily
reside in the C-terminal tail region. GFAPα is the most widely
expressed protein. Moreover, GFAPδ expression is limited to
specialized astrocyte populations and is more abundant in
neurogenic regions, such as the SVZ of the lateral ventricle, the
rostral migratory stream, the SGZ of the hippocampus and the
subpial astrocytes (39,41,55,56).
GFAPδ positive cells are specifically restricted to these
neurogenic areas, where expression of Nestin and Vimentin is also
observed, suggesting that GFAPδ may be a RGC-specific filament
protein that is required for their substrate functions and may be a
determining factor associated with neurogenesis (55,60).
The ratio of GFAPα and GFAPδ is an important factor in the
formation of the intermediate filament network, which is closely
associated with cellular functions (such as neurogenesis) or
cellular morphology (such as gliosis) (56). The changes in the expression levels
of the GFAP splice variants could significantly influence the
intermediate filament network, which is closely associated with
cell morphology and cell motility (59).
GFAPδ and the GFAP expression are first observed at
around gestational week 13 in the human fetal brain (41). In 2010, Middeldorp et al
(41) first confirmed that GFAPδ
was specifically found in radial glia and SVZ neural progenitors
during human brain development. The GFAP transcript is first
detectable at E9.5 in mice embryonic cortex, earlier than the
corresponding time for a human (55). During the developmental process,
GFAP expression continuously increases until a plateau arrives in
the adult brain. In the second trimester, differentiated astrocytes
from radial glia are observed in the germinal layers, corpus
callosum and cavum septum pellucidum (61). However, it remains unknown what
roles GFAPδ serves in the transition of RGCs into astrocytes. The
present study found that PEE could markedly decrease the
intensities of GFAP and GFAPδ, indicating that ethanol might impair
the development of radial glial fibres and influence the supportive
role for the migrating neuroblasts.
Nestin is a class VI intermediate filament protein
that is expressed in the majority of mitotically active neural
progenitors, including RGCs, which produce both neurons and
astrocytes in the developing CNS (62). After differentiation, Nestin
expression is downregulated. Using immunostaining, the present
study identified that the majority of Nestin labeled RGCs were
confined to the VZ and SVZ. Hence, Nestin was considered a marker
of NSCs. Using Nestin–/– mice, Wilhelmsson et al
(63) confirmed that Nestin
negatively regulated neuronal differentiation and survival through
its role in Notch signaling. Further experiments demonstrated that
Nestin was co-expressed with GFAP during the transitional
differentiation process of the neural progenitors into astrocytes
(64) or neurons (65). The increased colocalization of GFAP
and Nestin is a feature of reactive astrocytes (64). NSCs expressing both GFAP and Nestin
were termed stem astrocytes (63).
The Nestin+GFAP+ double labeling of neural
progenitors may assist in determining the status of neural
progenitors and their differentiation direction. In the present
study, ethanol treatment significantly decreased both Nestin and
GFAPδ expression levels in the SVZ. The co-expression of Nestin and
GFAPδ expression was also decreased by ethanol treatment. These
results indicated that ethanol treatment influenced the stem cell
potential of oRG and their morphologies.
Vimentin (also known as nanofilament) is another
intermediate filament protein expressed in neural stem/progenitor
cells and astrocytes (41,55). Its phosphorylation is an important
step required for the organization of the intermediate filament
network and the subcellular distribution of its proteins (66–68).
Mutations of Vimentin not only alter the cell morphology,
proliferative abilities and motility but are also closely
associated with alterations in neuronal differentiation of neural
progenitor cells and neurogenesis (67). In astrocytes and RGCs, Vimentin
forms an intermediate filament complex with GFAP and Nestin. The
absence of GFAP and Vimentin could decrease Notch signaling and
further increase neuronal differentiation (67,69). A
recent report by Cunningham et al (70) revealed that all mitotic cells at the
surface of the ventricle in the human and non-human primate dorsal
telencephalon express GFAP and Vimentin. Following differentiation
into astrocytes, RGCs stop expressing Vimentin and continually
upregulate GFAP expression, which suggests that intermediate
filament proteins can be transformed during the process of
differentiation (67). In the
present study, ethanol treatment accelerated the downregulation of
Vimentin and promoted the appearance of GFAPβ+
cells.
HOPX is the smallest Homeodomain-only protein, which
was identified as a temporally and spatially restricted gene in the
neurogenic regions in the developing medial cortex and cortical
hemisphere at a certain period during the process of neurogenesis
(71). Using in situ
hybridization, the HOPX gene was first detected at E12.5 forebrains
in the most medial part of the cortex. From E14.5 to E16.5, HOPX
expression in the forebrain increased from a low level at E14.5 to
its peak at E16.5 (71). Thus, HOPX
expression exhibits an expression gradient at a high level in the
cortical VZ and a low level in the lateral caudal forebrain. In the
dorsal forebrain, HOPX expression begins at E12.5 and reaches a
peak at ~E16.5 with a rostromedial to the caudolateral gradient
(71). At the same timepoint, RGCs
are gradually produced. Several molecular markers including GFAP,
SOX2 and Nestin are expressed in quiescent NSCs of this sub-region
(71). Moreover, HOPX can be
specifically expressed in slow-cycling or quiescent NSCs together
with GFAP, Nestin and SOX2, but is not co-expressed with
transit-amplifying cell progenitor markers, such as T-box brain
protein 2 or the neuroblast marker Doublecortin (72). The aforementioned studies suggested
that HOPX may modulate neurogenesis and self-renewal of neural stem
cells. Furthermore, the present study demonstrated that ethanol
treatment reduced the intensity of HOPX staining in
GFAPδ+ cells.
The S100β protein is an acidic calcium-binding
protein that can be expressed both in differentiated and
undifferentiated proliferating glial cells (73). Additionally, S100β is regarded as a
classical astrocyte marker, which suggests that S100β expression
characterizes a terminal maturation stage (48). During embryonic neurodevelopment,
the dynamic expression of S100β in NSCs and RGCs is associated with
the proliferative potential and the migration of neural progenitors
(73). A previous study reported
that the SVZ microenvironment can repress S100β expression
(48). Using transgenic S100β-EGFP
cells, Raponi et al (48)
confirmed that S100β expression defined a late neurodevelopmental
stage coinciding with the loss of their stem cell potency. RGCs, as
a type of neural stem cell, share certain similar characteristics
with immature and mature astrocytes and one of these
characteristics is the expression of the intermediate filament GFAP
(38). Hence, RGCs have both
neurogenic and gliogenic abilities during the development of the
embryonic forebrain (74). The
onset of S100β expression can be regarded as the acquisition of a
more mature developmental stage-related to spatiotemporal
maturation and the loss of neural stem cell potential (48). Amongst its multiple functions, S100β
has been identified as a promoter of cytoskeletal stabilization,
which serves an important role in normal corticogenesis (75). Decreased GFAP expression may impair
the formation of filament complexes together with Nestin and
further disrupt the supportive roles in the process of neuronal
migration. Co-expression of Nestin and S100β or Nestin and GFAP may
indicate that S100β+ or GFAP+ RGCs exhibit
more restricted developmental potential, and are capable of
differentiating into glial cells directed by factors in the local
environment (65). In the present
study, GFAPδ+ cells exhibited a strong intensity of
S100β expression following ethanol treatment, which indicated that
ethanol promoted the transformation of RGCs into astrocytes.
Ki67 is a nuclear protein, which is used as a marker
for identifying dividing cells. It can label cells in the late
G1, S, G2 and M phases, but not the
G0 phase of the cell cycle (76,77).
pHH3 is a proliferation marker that is closely associated with
mitotic chromatin condensation in the late G2 and M
phases of the cell cycle and HH3 is not phosphorylated during
apoptosis (78). Compared with
Ki67, PHH3 is a more reproducible proliferation and
mitosis-specific marker (77). The
results of the present study demonstrated that ethanol treatment
notably inhibited the proliferative and mitotic activities of
GFAPδ+ cells, which contributed to the decreased
production of neurons or intermediate progenitor cells and impaired
cortical thickness.
In summary, to the best of our knowledge, the
present study was the first to identify the effects of prenatal
alcohol exposure on the expression and distribution of GFAPδ during
early neurogenesis. The impairment in GFAPδ+ RGCs
contributed to prenatal alcohol exposure-induced abnormal cortical
development. Although the mechanisms have not fully been
elucidated, the results suggested that the regulation of GFAPδ may
highlight a novel direction for the treatment and management of
prenatal alcohol exposure associated with abnormal cortical
development.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the Key
Research and Development Foundation of Shaanxi Province, China
(grant no. 2018SF-029), National Natural Science Foundation of
China (grant no. 81760441; Principal Investigator, XLD) and Xi'an
Science and Technology Plan Project [grant no. 20YXYJ0009(11)].
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
YL, FYX and HZ designed the experimental plan. YL,
LNZ, LC and YL performed the experiments. YL analysed the data and
wrote the manuscript. FYX, HZ and XLD was the principal
investigator and revised the manuscript. All authors read and
approved the final version of the manuscript and agree to take
responsibility for the published article.
Ethics approval and consent to
participate
All animal maintenance and experimental procedures
were approved by the Institutional Animal Care and Use Committee of
Xi'an Medical University and animal experiments were performed in
accordance with the National Institute of Health Guide for the Care
and Use of Laboratory Animals (NIH Publications No. 80-23).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
GFAP
|
glial fibrillary acidic protein
|
|
RGCs
|
radial glial cells
|
|
oRGs
|
outer radial glial cells
|
|
NSCs
|
neural stem cells
|
|
PEE
|
prenatal ethanol exposure
|
|
BACs
|
blood alcohol concentrations
|
|
VZ
|
ventricular zone
|
|
SVZ
|
subventricular zone
|
|
IZ
|
intermediate zone
|
|
SP
|
subcortical plate
|
|
CP
|
cortical plate
|
|
CNS
|
central nervous system
|
References
|
1
|
Popova S, Lange S, Probst C, Gmel G and
Rehm J: Estimation of national, regional, and global prevalence of
alcohol use during pregnancy and fetal alcohol syndrome: A
systematic review and meta-analysis. Lancet Global health.
5:e290–e299. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Popova S, Lange S, Probst C, Parunashvili
N and Rehm J: Prevalence of alcohol consumption during pregnancy
and Fetal Alcohol Spectrum Disorders among the general and
Aboriginal populations in Canada and the United States. Eur J Med
Genet. 60:32–48. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Riley EP, Infante MA and Warren KR: Fetal
alcohol spectrum disorders: An overview. Neuropsychol Rev.
21:73–80. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wilhoit LF, Scott DA and Simecka BA: Fetal
alcohol spectrum disorders: Characteristics, complications, and
treatment. Community Ment Health J. 53:711–718. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Petrelli B, Weinberg J and Hicks GG:
Effects of prenatal alcohol exposure (PAE): Insights into FASD
using mouse models of PAE. Biochem Cell Biol. 96:131–147. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Davis-Anderson KL, Wesseling H, Siebert
LM, Lunde-Young ER, Naik VD, Steen H and Ramadoss J: Fetal regional
brain protein signature in FASD rat model. Reprod Toxicol.
76:84–92. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Creeley CE, Dikranian KT, Johnson SA,
Farber NB and Olney JW: Alcohol-induced apoptosis of
oligodendrocytes in the fetal macaque brain. Acta Neuropathol
Commun. 1:232013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Farber NB, Creeley CE and Olney JW:
Alcohol-induced neuroapoptosis in the fetal macaque brain.
Neurobiol Dis. 40:200–206. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gerlai R: Embryonic alcohol exposure:
Towards the development of a zebrafish model of fetal alcohol
spectrum disorders. Dev Psychobiol. 57:787–798. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Flentke GR and Smith SM: The avian embryo
as a model for fetal alcohol spectrum disorder. Biochem Cell Biol.
96:98–106. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Popova S, Lange S, Probst C, Gmel G and
Rehm J: Global prevalence of alcohol use and binge drinking during
pregnancy, and fetal alcohol spectrum disorder. Biochem Cell Biol.
96:237–240. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cheng HG, Deng F, Xiong W and Phillips MR:
Prevalence of alcohol use disorders in mainland China: A systematic
review. Addiction. 110:761–774. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang YY and D'Amato RC: Understanding
fetal alcohol spectrum disorders in China. J Pediatr Neuropsychol.
3:53–60. 2017. View Article : Google Scholar
|
|
14
|
Clarren SK, Alvord EC Jr, Sumi SM,
Streissguth AP and Smith DW: Brain malformations related to
prenatal exposure to ethanol. J Pediatr. 92:64–67. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Miller MW: Migration of cortical neurons
is altered by gestational exposure to ethanol. Alcohol Clin Exp
Res. 17:304–314. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lotfullina N and Khazipov R: Ethanol and
the developing brain: Inhibition of neuronal activity and
neuroapoptosis. Neuroscientist. 24:130–141. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sowell ER, Mattson SN, Kan E, Thompson PM,
Riley EP and Toga AW: Abnormal cortical thickness and
brain-behavior correlation patterns in individuals with heavy
prenatal alcohol exposure. Cereb Cortex. 18:136–144. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhou D, Lebel C, Lepage C, Rasmussen C,
Evans A, Wyper K, Pei J, Andrew G, Massey A, Massey D and Beaulieu
C: Developmental cortical thinning in fetal alcohol spectrum
disorders. NeuroImage. 58:16–25. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kang W, Wong LC, Shi SH and Hébert JM: The
transition from radial glial to intermediate progenitor cell is
inhibited by FGF signaling during corticogenesis. J Neurosci.
29:14571–14580. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Segklia A, Seuntjens E, Elkouris M,
Tsalavos S, Stappers E, Mitsiadis TA, Huylebroeck D, Remboutsika E
and Graf D: Bmp7 regulates the survival, proliferation, and
neurogenic properties of neural progenitor cells during
corticogenesis in the mouse. PLoS One. 7:e340882012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Choe Y, Huynh T and Pleasure SJ: Migration
of oligodendrocyte progenitor cells is controlled by transforming
growth factor β family proteins during corticogenesis. J Neurosci.
34:14973–14983. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tiberi L, Vanderhaeghen P and van den
Ameele J: Cortical neurogenesis and morphogens: Diversity of cues,
sources and functions. Curr Opin Cell Biol. 24:269–276. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bengoa-Vergniory N and Kypta RM: Canonical
and noncanonical Wnt signaling in neural stem/progenitor cells.
Cell Mol Life Sci. 72:4157–4172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Inestrosa NC and Varela-Nallar L: Wnt
signalling in neuronal differentiation and development. Cell Tissue
Res. 359:215–223. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Razavi MJ, Zhang T, Chen H, Li Y, Platt S,
Zhao Y, Guo L, Hu X, Wang X and Liu T: Radial structure scaffolds
convolution patterns of developing cerebral cortex. Front Comput
Neurosci. 11:762017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamamoto H, Mandai K, Konno D, Maruo T,
Matsuzaki F and Takai Y: Impairment of radial glial
scaffold-dependent neuronal migration and formation of double
cortex by genetic ablation of afadin. Brain Res. 1620:139–152.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Falk S and Götz M: Glial control of
neurogenesis. Curr Opin Neurobiol. 47:188–195. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yuzwa SA and Miller FD: Deciphering
cell-cell communication in the developing mammalian brain.
Neurogenesis (Austin). 4:e12864252017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Miller MW and Robertson S: Prenatal
exposure to ethanol alters the postnatal development and
transformation of radial glia to astrocytes in the cortex. J Comp
Neurol. 337:253–266. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rubert G, Miñana R, Pascual M and Guerri
C: Ethanol exposure during embryogenesis decreases the radial glial
progenitorpool and affects the generation of neurons and
astrocytes. J Neurosci Res. 84:483–496. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vallés S, Pitarch J, Renau-Piqueras J and
Guerri C: Ethanol exposure affects glial fibrillary acidic protein
gene expression and transcription during rat brain development. J
Neurochem. 69:2484–2493. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nash R, Krishnamoorthy M, Jenkins A and
Csete M: Human embryonic stem cell model of ethanol-mediated early
developmental toxicity. Exp Neurol. 234:127–135. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Johansson PA, Cappello S and Götz M: Stem
cells niches during development-lessons from the cerebral cortex.
Curr Opin Neurobiol. 20:400–407. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Götz M, Hartfuss E and Malatesta P: Radial
glial cells as neuronal precursors: A new perspective on the
correlation of morphology and lineage restriction in the developing
cerebral cortex of mice. Brain Res Bull. 57:777–788. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Malatesta P, Appolloni I and Calzolari F:
Radial glia and neural stem cells. Cell Tissue Res. 331:165–178.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jinnou H, Sawada M, Kawase K, Kaneko N,
Herranz-Pérez V, Miyamoto T, Kawaue T, Miyata T, Tabata Y, Akaike
T, et al: Radial glial fibers promote neuronal migration and
functional recovery after neonatal brain injury. Cell Stem Cell.
22:128–137.e9. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nowakowski TJ, Pollen AA,
Sandoval-Espinosa C and Kriegstein AR: Transformation of the radial
glia scaffold demarcates two stages of human cerebral cortex
development. Neuron. 91:1219–1227. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Barry DS, Pakan JM and McDermott KW:
Radial glial cells: Key organisers in CNS development. Int J
Biochem Cell Biol. 46:76–79. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Garcia AD, Doan NB, Imura T, Bush TG and
Sofroniew MV: GFAP-expressing progenitors are the principal source
of constitutive neurogenesis in adult mouse forebrain. Nat
Neurosci. 7:1233–1241. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Johnson K, Barragan J, Bashiruddin S,
Smith CJ, Tyrrell C, Parsons MJ, Doris R, Kucenas S, Downes GB,
Velez CM, et al: Gfap-positive radial glial cells are an essential
progenitor population for later-born neurons and glia in the
zebrafish spinal cord. Glia. 64:1170–1189. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Middeldorp J, Boer K, Sluijs JA, De
Filippis L, Encha-Razavi F, Vescovi AL, Swaab DF, Aronica E and Hol
EM: GFAPdelta in radial glia and subventricular zone progenitors in
the developing human cortex. Development. 137:313–321. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gotoh H, Nomura T and Ono K: Glycogen
serves as an energy source that maintains astrocyte cell
proliferation in the neonatal telencephalon. J Cereb Blood Flow
Metab. 37:2294–2307. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
National Research Council, . Guide for the
Care and Use of Laboratory Animals. 8th edition. The National
Academies Press; Washington, DC: 2011
|
|
44
|
Parnell SE, Chen SY, Charness ME, Hodge
CW, Dehart DB and Sulik KK: Concurrent dietary administration of
D-SAL and ethanol diminishes ethanol's teratogenesis. Alcohol Clin
Exp Res. 31:2059–2064. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Parnell SE, Dehart DB, Wills TA, Chen SY,
Hodge CW, Besheer J, Waage-Baudet HG, Charness ME and Sulik KK:
Maternal oral intake mouse model for fetal alcohol spectrum
disorders: Ocular defects as a measure of effect. Alcohol Clin Exp
Res. 30:1791–1798. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Dunn KW, Kamocka MM and McDonald JH: A
practical guide to evaluating colocalization in biological
microscopy. Am J Physiol Cell Physiol. 300:C723–C742. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Pekny M and Pekna M: Astrocyte
intermediate filaments in CNS pathologies and regeneration. J
Pathol. 204:428–437. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Raponi E, Agenes F, Delphin C, Assard N,
Baudier J, Legraverend C and Deloulme JC: S100B expression defines
a state in which GFAP-expressing cells lose their neural stem cell
potential and acquire a more mature developmental stage. Glia.
55:165–177. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hutton SR and Pevny LH: SOX2 expression
levels distinguish between neural progenitor populations of the
developing dorsal telencephalon. Dev Biol. 352:40–47. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kamachi Y and Kondoh H: Sox proteins:
Regulators of cell fate specification and differentiation.
Development. 140:4129–4144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Bani-Yaghoub M, Tremblay RG, Lei JX, Zhang
D, Zurakowski B, Sandhu JK, Smith B, Ribecco-Lutkiewicz M, Kennedy
J, Walker PR and Sikorska M: Role of Sox2 in the development of the
mouse neocortex. Dev Biol. 295:52–66. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ferri A, Favaro R, Beccari L, Bertolini J,
Mercurio S, Nieto-Lopez F, Verzeroli C, La Regina F, De Pietri
Tonelli D, Ottolenghi S, et al: Sox2 is required for embryonic
development of the ventral telencephalon through the activation of
the ventral determinants Nkx2.1 and Shh. Development.
140:1250–1261. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Hagey DW and Muhr J: Sox2 acts in a
dose-dependent fashion to regulate proliferation of cortical
progenitors. Cell Rep. 9:1908–1920. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wang S, Chandler-Militello D, Lu G, Roy
NS, Zielke A, Auvergne R, Stanwood N, Geschwind D, Coppola G,
Nicolis SK, et al: Prospective identification, isolation, and
profiling of a telomerase-expressing subpopulation of human neural
stem cells, using sox2 enhancer-directed fluorescence-activated
cell sorting. J Neurosci. 30:14635–14648. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mamber C, Kamphuis W, Haring NL, Peprah N,
Middeldorp J and Hol EM: GFAPδ expression in glia of the
developmental and adolescent mouse brain. PLoS One. 7:e526592012.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kamphuis W, Mamber C, Moeton M, Kooijman
L, Sluijs JA, Jansen AH, Verveer M, de Groot LR, Smith VD,
Rangarajan S, et al: GFAP isoforms in adult mouse brain with a
focus on neurogenic astrocytes and reactive astrogliosis in mouse
models of Alzheimer disease. PLoS One. 7:e428232012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yang Z and Wang KK: Glial fibrillary
acidic protein: From intermediate filament assembly and gliosis to
neurobiomarker. Trends Neurosci. 38:364–374. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Howard BM, Zhicheng M, Filipovic R, Moore
AR, Antic SD and Zecevic N: Radial glia cells in the developing
human brain. Neuroscientist. 14:459–473. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Sullivan SM: GFAP variants in health and
disease: Stars of the brain… and gut. J Neurochem. 130:729–732.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zalfa C, Grasselli C, Santilli G, Ferrari
D, Lamorte G, Vescovi AL and De Filippis L: GFAP delta as divergent
marker of human glial progenitors. J Stem Cell Res Ther. 8:92018.
View Article : Google Scholar
|
|
61
|
Rezaie P, Ulfig N and Male D: Distribution
and morphology of GFAP-positive astrocytes in the human fetal brain
at second trimester. Neuroembryology. 2:50–63. 2003. View Article : Google Scholar
|
|
62
|
Sunabori T, Tokunaga A, Nagai T, Sawamoto
K, Okabe M, Miyawaki A, Matsuzaki Y, Miyata T and Okano H:
Cell-cycle-specific nestin expression coordinates with
morphological changes in embryonic cortical neural progenitors. J
Cell Sci. 121:1204–1212. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wilhelmsson U, Lebkuechner I, Leke R,
Marasek P, Yang X, Antfolk D, Chen M, Mohseni P, Lasič E, Bobnar
ST, et al: Nestin regulates neurogenesis in mice through notch
signaling from astrocytes to neural stem cells. Cereb Cortex.
29:4050–4066. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Messam CA, Hou J and Major EO:
Coexpression of nestin in neural and glial cells in the developing
human CNS defined by a human-specific anti-nestin antibody. Exp
Neurol. 161:585–596. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Vukojevic K, Petrovic D and Saraga-Babic
M: Nestin expression in glial and neuronal progenitors of the
developing human spinal ganglia. Gene Expr Patterns. 10:144–151.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Eriksson JE, He T, Trejo-Skalli AV,
Härmälä-Braskén AS, Hellman J, Chou YH and Goldman RD: Specific in
vivo phosphorylation sites determine the assembly dynamics of
vimentin intermediate filaments. J Cell Sci. 117:919–932. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Chen M, Puschmann TB, Marasek P, Inagaki
M, Pekna M, Wilhelmsson U and Pekny M: Increased neuronal
differentiation of neural progenitor cells derived from
phosphovimentin-deficient mice. Mol Neurobiol. 55:5478–5489. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chou YH, Khuon S, Herrmann H and Goldman
RD: Nestin promotes the phosphorylation-dependent disassembly of
vimentin intermediate filaments during mitosis. Mol Biol Cell.
14:1468–1478. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Widestrand A, Faijerson J, Wilhelmsson U,
Smith PL, Li L, Sihlbom C, Eriksson PS and Pekny M: Increased
neurogenesis and astrogenesis from neural progenitor cells grafted
in the hippocampus of GFAP−/− Vim−/− mice.
Stem Cells. 25:2619–2627. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Cunningham CL, Martinez-Cerdeno V and
Noctor SC: Diversity of neural precursor cell types in the prenatal
macaque cerebral cortex exists largely within the astroglial cell
lineage. PLoS One. 8:e638482013. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li D, Takeda N, Jain R, Manderfield LJ,
Liu F, Li L, Anderson SA and Epstein JA: Hopx distinguishes
hippocampal from lateral ventricle neural stem cells. Stem Cell
Res. 15:522–529. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Braun SM and Jessberger S: Adult
neurogenesis: Mechanisms and functional significance. Development.
141:1983–1986. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Patro N, Naik A and Patro IK: Differential
temporal expression of S100β in developing rat brain. Front Cell
Neurosci. 9:872015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Docampo-Seara A, Santos-Durán GN, Candal E
and Rodríguez Díaz MÁ: Expression of radial glial markers (GFAP,
BLBP and GS) during telencephalic development in the catshark
(Scyliorhinus canicula). Brain Struct Funct. 224:33–56. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Aronne MP, Guadagnoli T, Fontanet P,
Evrard SG and Brusco A: Effects of prenatal ethanol exposure on rat
brain radial glia and neuroblast migration. Exp Neurol.
229:364–371. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Malik S, Vinukonda G, Vose LR, Diamond D,
Bhimavarapu BB, Hu F, Zia MT, Hevner R, Zecevic N and Ballabh P:
Neurogenesis continues in the third trimester of pregnancy and is
suppressed by premature birth. J Neurosci. 33:411–423. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang J and Jiao J: Molecular biomarkers
for embryonic and adult neural stem cell and neurogenesis. Biomed
Res Int. 2015:7275422015.PubMed/NCBI
|
|
78
|
van den Berge SA, van Strien ME, Korecka
JA, Dijkstra AA, Sluijs JA, Kooijman L, Eggers R, De Filippis L,
Vescovi AL, Verhaagen J, et al: The proliferative capacity of the
subventricular zone is maintained in the parkinsonian brain. Brain.
134:3249–3263. 2011. View Article : Google Scholar : PubMed/NCBI
|