Introduction
Alzheimer's disease (AD), which is pathologically
manifested as amyloid β (Aβ) deposition and the formation of
neurofibrillary tangles (NFTs) (1),
accompanied by the decrease of memory, judgment, attention and
cognitive ability, is a neurodegenerative disorder (2). It has a serious effect on the quality
of life of patients. Thus far, there are no effective means to
prevent or treat AD (3).
Ginsenoside Rg1 (Rg1) exhibits anti-inflammatory,
antioxidant, antitumor and anti-apoptosis effects (4,5). The
effects of Rg1 on memory enhancement and neuroprotection have
attracted wide attention (6,7). Rg1,
which inhibits Aβ plaques and has anti-inflammatory and
anti-oxidation effects, can improve the learning and memory ability
of AD mice (6). In vitro
experiments, by accelerating the activation and nuclear
translocation and transcription of NF-κB, Rg1 increased the binding
of NF-κB with the corresponding DNA sites at the promoter region of
the β secretase gene, inhibiting the transcription and translation
of β secretase and thus reducing the production of Aβ (7).
The microbiota of the large intestine is a complex
micro-ecosystem in the host and its stability is closely related to
the health of the body (8).
Intestinal microbial composition and the communication between
microbes and the brain may change with age and the microbial
composition of those >65 years old is significantly different
from those <9 years old (9–11).
Intestinal symptoms have been observed in patients with
neurodegenerative diseases and mental disorders, including
Parkinsons disease, autism, schizophrenia, Alzheimer's disease and
depression (12–15).
Previous studies have confirmed that AD is closely
associated with the composition of intestinal microflora. For
example, bacterial metabolites, such as short-chain fatty acids,
may mediate the maturation of microglia (16) and further result in the clearance of
Aβ and the protection of neurons (17). When rats aged 22.5–25 months were
exposed to E. coli able to produce curly amyloid fibrils,
the deposition of Aβ in the gut and brain neurons increased and the
proliferation of glial cells and astrocytes was enhanced (18). Antibiotics, which interfere with the
intestinal microbial diversity of AD mice, significantly reduce the
deposition of Aβ and glial cells hyperplasia (19). 16S rRNA sequencing of fecal samples
of amyloid precursor protein (APP)/presenilin 1 transgenic mice and
wild-type mice demonstrates a significant difference in the
composition of intestinal flora between the two groups of mice
(20). Zhang et al (8) demonstrated that, compared with the
wild-type mice, AD mice had more diversified intestinal bacteria, a
lower ratio of Firmicutes to Bacteroides and more
Proteus and Verrucous.
The effect of traditional Chinese medicine Rg1 on
intestinal bacteria has become a focus of medical research. Poria
cocos water-insoluble polysaccharide can adjust intestinal
flora in mice, increasing Helicobacter and
Clostridium and further improving hyperglycemia and
hyperlipidemia (21). Following
long-term use of ginseng extract, the number of probiotics such as
Bifidobacterium, Lactobacillus and Clostridium was
significantly increased in the intestinal flora of rats, suggesting
that it can promote the growth of probiotics (22). Tiansi solution, a traditional
Chinese herbal medicine, which increases the relative abundance of
Ruminococcaceae, Lactococcus and Lactobacillus,
significantly improves the exploratory behavior of depressed rats
(23).
The Chinese tree shrew (Tupaia belangeri
chinensis) is considered a ‘low-level primate’ based on its
relation to primates (24). There
is a genetic basis for using tree shrew as a viable neurological
disease model (25). The tree shrew
model of AD has been successfully constructed in our study
(26). A recent study has suggested
that the neocortex development of the tree shrew is closer to
primates than to rodents (27).
This makes the tree shrew a good experimental animal in
neurological diseases.
The present study investigated the influence of Rg1
on the expression of Aβ-related proteins, phosphorylated (p-)tau
and apoptosis-related protein including Bax, Bcl-2,
microtubule-associated protein 2 (MAP2) and Fox-3 (NeuN), in AD
tree shrews and further analyzed the microbiota of their large
intestine.
Materials and methods
Ethics statement
The present study was performed in strict accordance
with the Health Guide for the Care and Use of Laboratory Animals
(28). The protocol was approved by
the Committee on the Ethics of Animal Experiments of Kunming
Medical University (approval number KMMU2018004).
Animals
Tree shrews were purchased from the Department of
Experimental Animals of Kunming Medical University. As female tree
shrews menstruate, only males were used to avoid the influence of
menstruation. A total of 15 adult male tree shrews were used (10–12
months; weight range, 130–150 g). The tree shrews were housed in
independent ventilation cages at an ambient temperature of 24±2°C
and a humidity of 40–60% under a 12-h light/dark cycle with free
access to food and water at Kunming Medical University. All the
animals were acclimated to their new habitat for a week prior to
the experiments.
The tree shrews were divided into the model group
(D+A), the control group (CT) and the treatment group (D+A+Rg1),
with five tree shrews in each group. The D+A group received
one-time lateral ventricle injection (0.8 mm posterior to the
bregma, 1.5 mm lateral to the sagittal suture 3.6 mm beneath the
surface of the brain; the endocranium was exposed by drilling the
skull, and a micro syringe inserted vertically for injection.
Aβ25–35 (10 µg/5 µl; cat. no. A9810; Sigma-Aldrich; Merck KGaA) and
D-galactose (60 mg/kg/day; cat. no. A-G5388; Sigma-Aldrich; Merck
KGaA) were injected subcutaneously, using a syringe for 6 weeks
(29). Ginsenoside Rg1 (Kunming
Pharmaceutical Group Co., Ltd.) was administered to the treatment
group (30 mg/kg/day) following the basis of the administration
modeling method in the D+A group. The control group was given an
equal amounts of saline and the injection site was the same as for
Aβ25–35 in the model group. Following the Morris water maze tests,
all shrews were sacrificed by exsanguination and the cerebral
hemisphere was obtained; one hemisphere was perfused with 4%
paraformaldehyde (4°C) overnight and the hippocampus and cortex
taken for immunohistochemistry. The other hemisphere was frozen at
−80°C for western blotting. The content of the large intestine was
obtained and snap-frozen in liquid nitrogen and stored at −80°C for
16SrRNA analysis.
Morris water maze
A black circular pool of 160 cm diameter with a
platform in its center was used. Water was poured into the pool to
a depth of ~40 cm, then milk powder was scattered to make the water
opaque. The water temperature was maintained at 22±1°C and the
platform height at ~11 cm. A tracking camera located directly above
the water was used to record the position, swimming time and paths
of the tree shrews. The first day was adaptation training, with the
water maze arranged with a visible platform. Tree shrews were held
to face the wall of the pool and dropped down from a height of 10
cm above the water surface. On the first days adaptation training,
if the platform was not found after 30 sec, the tree shrew would be
guided to the platform and then be kept there for 10 sec. The tree
shrew was taken out and wiped dry, and the procedure repeated for
two rounds over 2–6 days for the positioning cruise experiment. In
order to hide the platform, it was placed 1 cm below the water
surface. The positioning cruise experiment was performed for 2–6
days. The same operation as the first day was performed to record
how long it took the tree shrew to find the platform. In the
spatial search experiment (7th day), the platform which was used in
the experiment in the previous 7 days was removed, and the number
of times that the tree shrews crossed the platform were recorded.
Software (Morris water maze: cat. no. XR-XM101; Shanghai Xin-Xin
soft Information Technology Co., Ltd.) recorded the escape latency
and the frequency of crossing the platform of tree shrews. GraphPad
Prism 8.0.2 (GraphPad Software, Inc.) was used for analysis.
Western blot analysis
Total proteins of hippocampal tissues were extracted
by using a protein extraction kit (cat. no. R0020; Beijing Solarbio
Science & Technology Co., Ltd.). The lysate was added
proportionally as 200 µl lysate per 20 mg of tissue and seahorse
tissue was homogenized with glass homogenizer until fully lysed.
The lysate was centrifuged for 3–5 min at 5,000 × g and −4°C and
the supernatant removed. Hippocampal protein concentration was
determined by the BCA protein concentration assay kit (cat. no.
PC0020; Beijing Solarbio Science & Technology Co., Ltd.). The
supernatant of the protein sample was heated at 95°C for 10 min.
Sodium dodecyl sulfate-polyacrylamide gel (10%) was prepared by a
SDS-page gel preparation kit with a sample amount of 40 µg, and
then electrophoresed for 30 min at 80 V and 1.5 h at 120 V. Protein
bands transferred onto polyvinylidene fluoride membranes and
blocked with 5% skimmed milk at room temperature for 1 h. The
membranes were incubated overnight at 4°C with antibodies against
MAP2 (1:1,000; Cell Signaling Technology, Inc.; cat. no. 8707),
NeuN (1:1,000; Cell Signaling Technology, Inc.; cat. no. 24307),
β-secretase 1 (BACE1; 1:500; Abcam; cat. no. ab183612) and β-actin
(1:1,000; Elabscience, Inc.; cat. no. 20031). The secondary
antibody [peroxidase-conjugated goat anti-rabbit IgG (H+L); 1:5000;
OriGene Technologies, Inc.; cat. no. ZB-2301] was incubated for 1 h
and then developed with ECL reagent (ECL Plus Ultra Sensitive kit;
Phygene Life Sciences; cat. no. PH0353). Chemiluminescence signals
were imaged by a Gel Imaging System (Bio-Rad Laboratories, Inc.)
and protein band signals are quantified by ImageJ software (v1.8.0;
National Institutes of Health).
Immunohistochemical analysis of
hippocampus and cortex
The cerebral hemisphere was fixed in 4%
paraformaldehyde for 48 h at room temperature, rinsed under tap
water for 1.5 h, changed to 70% alcohol and soaked for 2 h, changed
to 80% alcohol and soaked for 2 h, changed to 90% alcohol and
soaked for 1.5 h, changed to 95% alcohol and soaked for 1.5 h,
changed to 100% alcohol and soaked for 45 min, changed to another
100% alcohol and soaked for 45 min, changed to xylene and soaked
for 15 min, changed to another 100% xylene and soaked for 15 min,
changed to paraffin and soaked for 4 h before being cut into
coronal sections at 5 µm thickness and warmed for 30 min. The
sections were dewaxed (xylene 1 for 10 min, xylene 2 for 10 min,
100% ethanol 1 for 3 min, 100% ethanol 2 for 3 min, 95% ethanol 3
min, 90% ethanol 3 min, 80% ethanol 3 min, 70% ethanol 3 min and
distilled water 3 min). Following heating with citric acid antigen
repair solution in a pressure cooker (100°C) for 2 min the sections
were washed with PBS for three times, 3 min per time. The sections
were blocked with 3% hydrogen peroxide for 10 min at room
temperature and washed with PBS for three times, 3 min per time.
Primary antibodies Aβ1–42 (1:200; cat. no. ab2539; Abcam), APP
(1:4,000; Sigma-Aldrich; Merck KGaA; cat. no. SAB4300464), p-tau
(1:1,000; cat. no. ab92676; Abcam), Bax (1:1,000; cat. no. 5023;
Cell Signaling Technology, Inc.), Bcl-2 (1:1,000; cat. no. 4223;
Cell Signaling Technology, Inc.) were added and the sections placed
into a 4°C refrigerator overnight. Taken out of the 4°C
refrigerator, the sections were rinsed with PBS for three times, 3
min per time and secondary antibody (goat anti-rabbit IgG; 1:5,000;
cat. no. AP132; Sigma-Aldrich; Merck KGaA) added and incubated for
15 min at room temperature. PBS was used to rinse for 3 times at 3
min per time. DAB was used for staining for 4 min and hematoxylin
counterstaining for 2 min at room temperature. After washing in
water for several seconds the sections were differentiated in
hydrochloric acid and alcohol. The sections were dehydrated (a
gradient of 70% ethanol, 80% ethanol, 90% ethanol, 95% ethanol,
100% ethanol 1, and 100% ethanol for 1 min, and xylene 1 and xylene
2 for 10 min), sealed and allowed to dry. A total of three sections
were taken randomly from each animal, five animals in each group.
The staining intensity over the entire surface area of the slide
was calculated using ImageJ software (v.1.51j8; National Institutes
of Health).
Analysis microbiota of large
intestine
The contents of the large intestine were extracted
by using a DNA extraction kit (E.Z.N.A.® Stool DNA kit;
cat. no. D4015-01; Omega Bio-Tek, Inc.) and the bacterial genome
was extracted according to the manufacturers protocols. The
bacterial genomic DNA of large intestine contents among groups D+A,
CT and D+A+Rg1 were detected by 1% agarose gel electrophoresis. The
sequencing primers 341F (5′-CCTAYGGGRBGCASCAG-3′) and 806R
(5′-GGACTACNNGGGTATCTAAT-3′) were used to amplify V3-V4 region of
bacterial 16S rDNA gene sequence using the sequencing kit HiSeq
Rapid SBS Kit v2 that performed in accordance with the
manufacturers protocols. (cat. no. FC-402-4022; Illumina,
Inc.).
PCR products were quantified by QuantiFluor™-ST blue
fluorescence quantitative system (Promega Corporation). An Illumina
PE250 library was built (Illumina, Inc.): ‘Y’ sticky joints were
connected, self-connecting segments were removed by magnetic bead
screening, the template libraries were enriched by PCR, and the
single-stranded DNA fragments were produced by sodium hydroxide.
The database, obtained from the Illumina PE250 platform (Illumina,
Inc.), was uploaded to NCBI Sequence Read Archive (SRP:
227299).
In order to get corresponding species classification
information from each operational taxonomic unit (OTU), the present
study used the RDP Classifier (v 2.2; http://sourceforge.net/projects/rdp-classifier/)
Bayesian algorithm on the 97% similarity level OTU to characterize
taxonomy sequence analysis, at each classification level (domain,
kingdom, phylum, class, order, family, genus and species) for the
community composition of each sample.
The Chao, Shannon and diversity indices were
analyzed based on Mothur (v.1.30.1 http://www.mothur.org). Linear discriminant analysis
effect size (LEfSe) uses linear discriminant analysis (LDA) to
estimate the effect of abundance of each component (species) on
different effects. LDA is performed according to different grouping
conditions according to taxonomic composition to identify the
communities or species that have a significant influence on sample
division.
Shannon: One of the microbial diversity indexes used
to estimate microbial diversity in samples. It is often used with
the Simpson diversity index to reflect the Alpha diversity index.
Higher Shannon values indicate higher community diversity.
Hshannon=-∑i=1SobsniNlnniN
Sobs = the actual number of OTUs
measured; ni= the sequence number contained in
the ith OTU; N = the number of sequences.
Chao: This is an index to estimate the number of OTU
in a sample by using the chao1 algorithm. Chao1 is often used in
ecology to estimate the total number of species and was first
proposed by Chao (30). The
calculation formula used in this analysis is as follows:
Schao1=Sobs+n1(n1-1)2(n2+1)
Schao1= Estimated OTU number;
Sobs= actual observed OTU number;
n1= number of OTUs containing only one sequence
(e.g. ‘Singletons’); n2= number of OTUs containing only
two sequences (e.g. ‘Doubletons’).
Statistical analysis
All data are expressed as mean ± standard deviation
or arithmetical mean. Differences among groups were analyzed by
one-way analysis of variance (ANOVA) followed by Tukeys post hoc
test. Statistical analyses were performed using SPSS v21.0 software
(IBM Corp.) except for Fig. 1A,
which was analyzed by two-way ANOVA followed by Tukeys post hoc
test. GraphPad Prism 8.0.2 (GraphPad Software, Inc.) software was
used for the statistical graphs. Metastats (http://metastats.cbcb.umd.edu/) difference analysis
based on the data of community abundance detected the differences
of the two groups of microbial communities in abundance, and
performed multiple hypothesis tests analysis, assess the
significance of the observed differences. P<0.05 was considered
to indicate a statistically significant difference.
Results
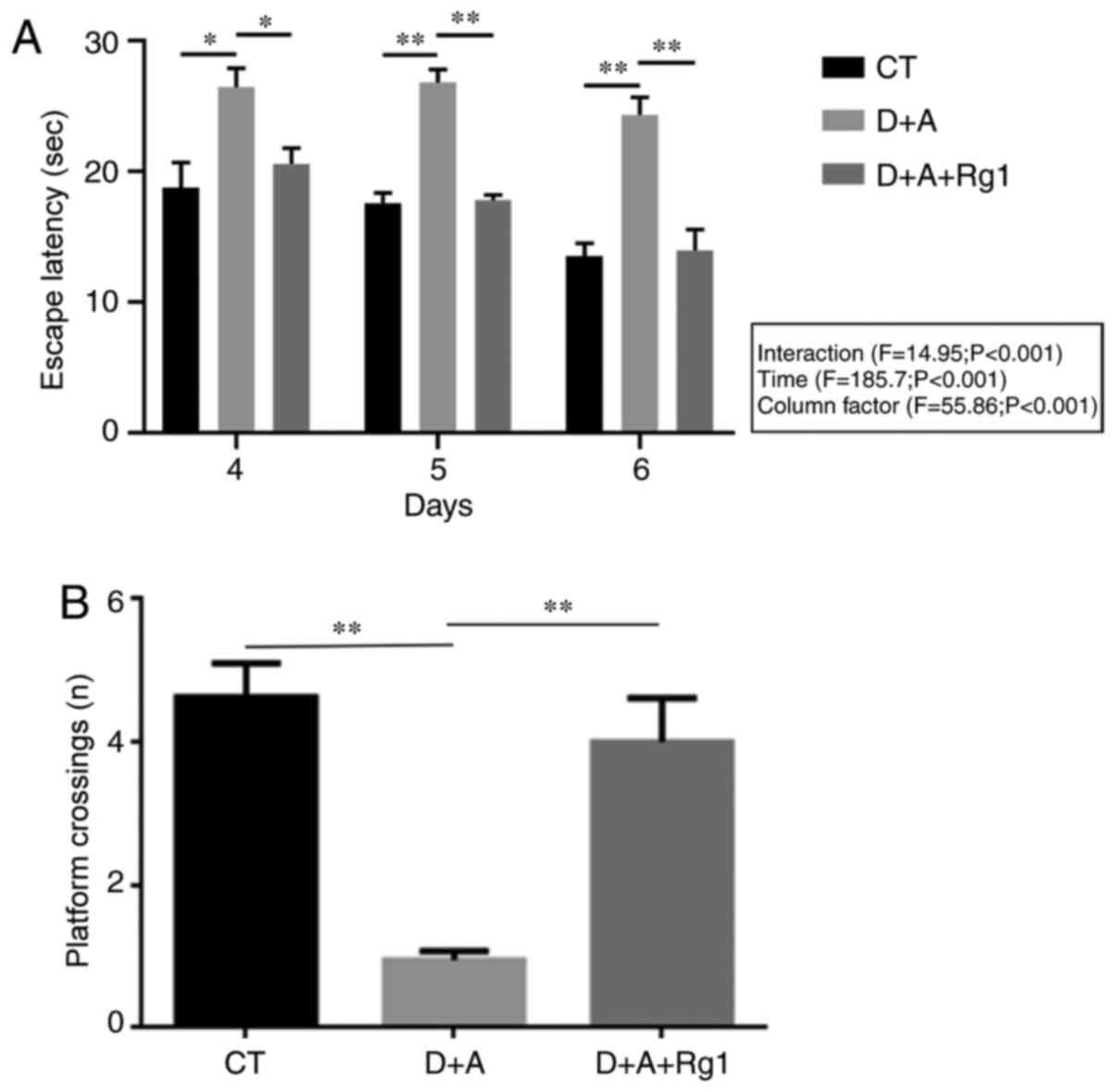
Rg1 improves learning and memory
capacity in AD tree shrews
A Morris water maze was used to detect the escape
latency period (2–6 days) and the times of crossing the platform
(7th day) of tree shrews in the 3 groups. The results demonstrated
that the evasive incubation period of group D+A+Rg1 was
significantly lower compared with group D+A (P<0.01) on days 4–6
(Fig. 1A) and no significantly
difference was observed among the individual groups on days 4–6.
The number of times of crossing the platform of group D+A+Rg1 was
significantly greater compared with group D+A (P<0.01; Fig. 1B).
Rg1 reduces production of Aβ1–42 and
phosphorylation of tau
Immunohistochemistry demonstrated no significant
difference in the expression of APP protein in the hippocampus and
cortex (Fig. 2A and C). Western
blot analysis demonstrated that the expression of BACE1 protein was
significantly increased in the hippocampus of the D+A group
compared with the CT group (P<0.01; Fig. 2B and D). The expression of BACE1
protein was significantly lower in the hippocampus of group D+A+Rg1
compared with group D+A (P<0.01; Fig. 2B and D). The expression of Aβ1–42 in
the hippocampus and cortex of group D+A+Rg1 was significantly lower
compared with group D+A (P<0.01; Fig. 2E and G). Immunohistochemistry
demonstrated that the expression of tau phosphorylated at serine
404 (pS404-tau) in the hippocampus and cortex of group D+A was
significantly higher compared with the CT group (P<0.01). Rg1
significantly inhibited the phosphorylation of pS404-tau in the
hippocampus and cortex (P<0.01; Fig.
2F and H).
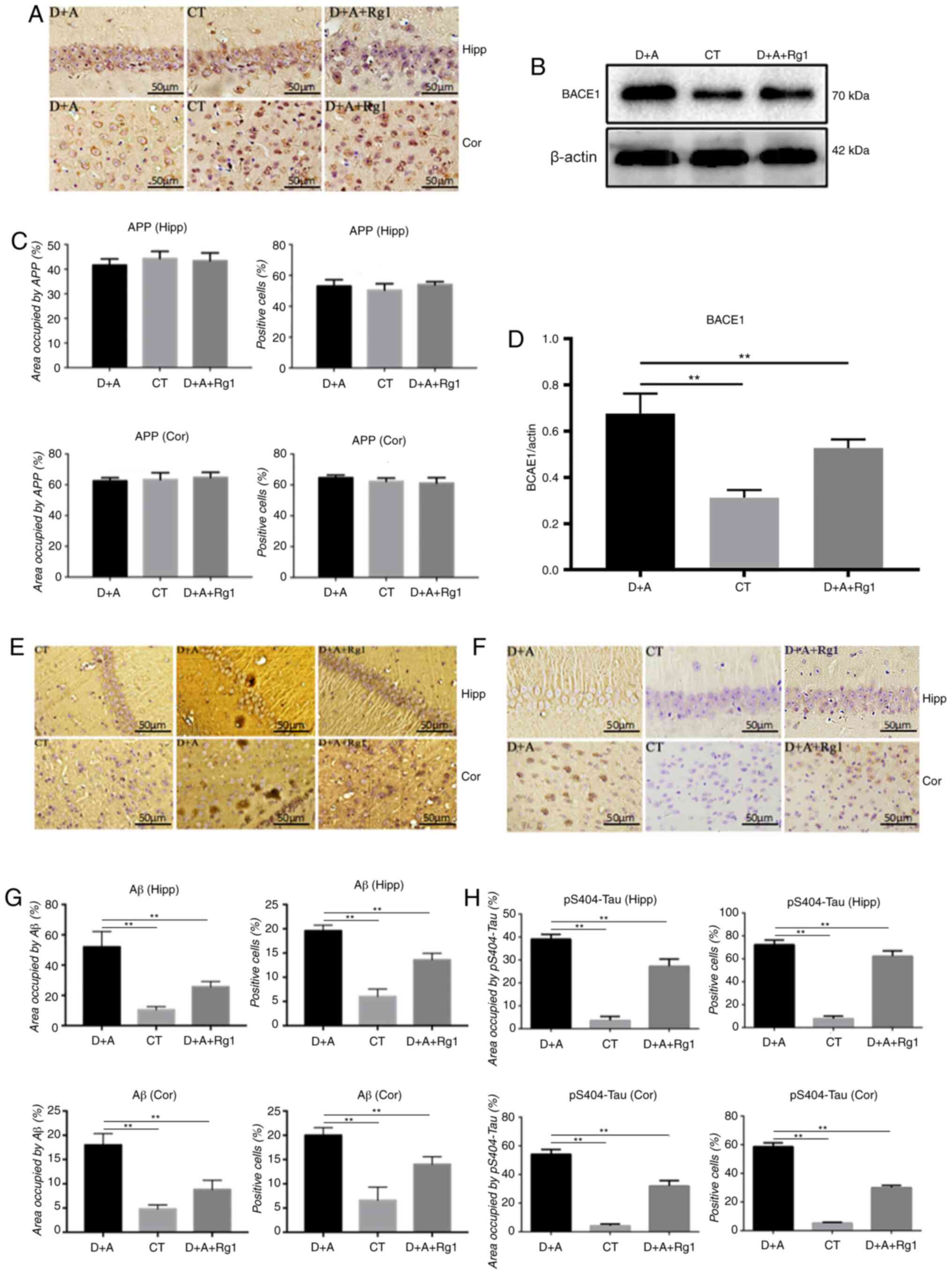 | Figure 2.Effect of ginsenoside Rg1 on Aβ1–42
associated protein and pS404-tau expression in AD tree shrews.
Immunohistochemical analysis of (A and C) APP, (E and G) Aβ1–42 and
(F and H) pS404-tau in the hippocampus and cortex. Western blot
analysis of (B and D) BACE1 in the hippocampus and cortex.
**P<0.01. Aβ, amyloid β; pS404-tau, tau phosphorylated at serine
404; AD, Alzheimer's disease; APP, amyloid precursor protein; CT,
control group; D+A, model group; D+A+Rg1, treatment group; Rg1,
ginsenoside Rg1; Hipp, hippocampus; cor, cortex; BACE1, β-secretase
1. |
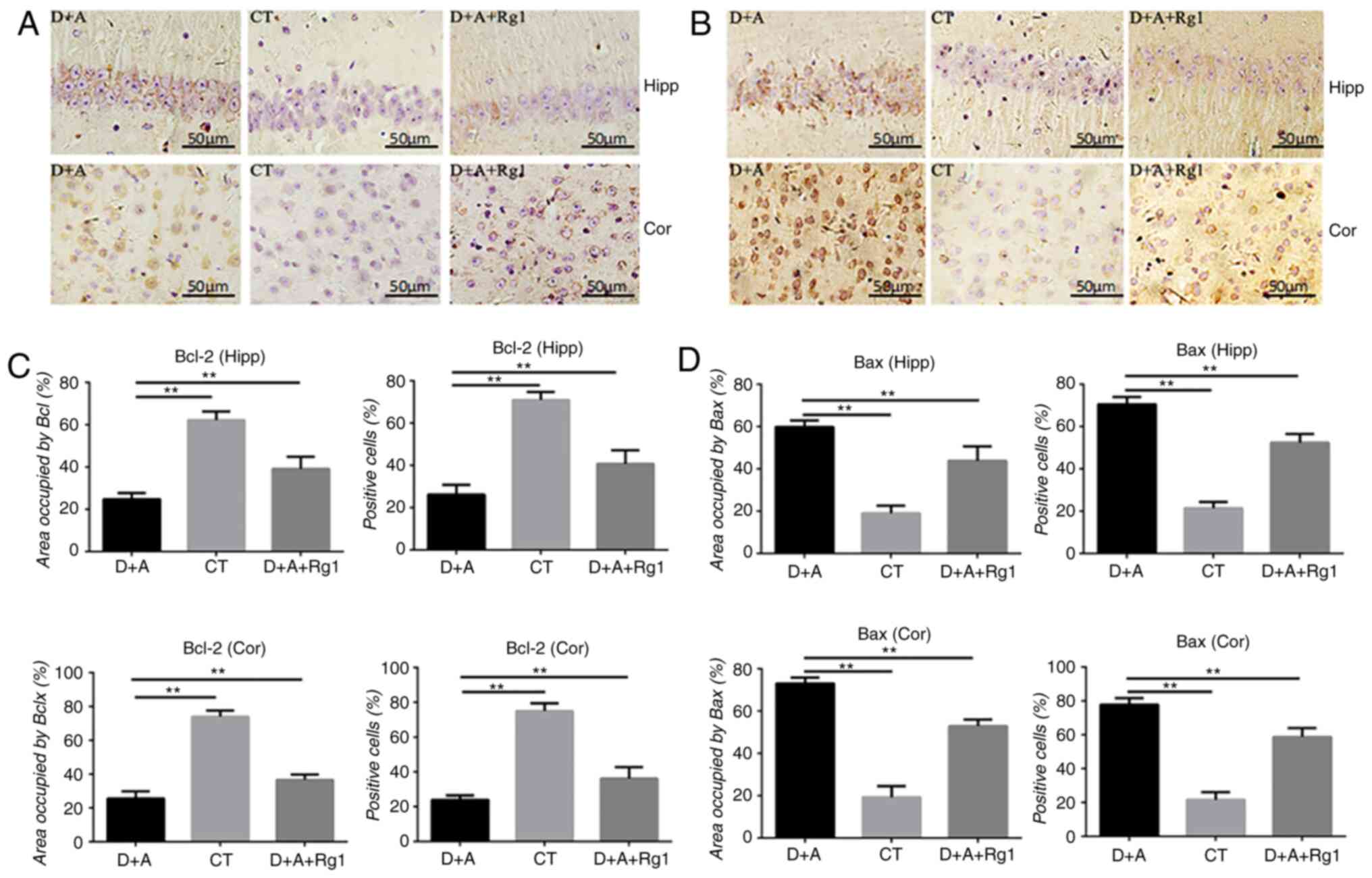
Rg1 increases the expression of
anti-apoptotic factor Bcl-2 and decreases the expression of
pro-apoptotic factor Bax
Immunohistochemistry of the hippocampus and cortex
demonstrated that the expression of Bcl-2 protein in the D+A group
was significantly decreased compared with the CT group (P<0.01).
Compared with the D+A group (P<0.01), Rg1 significantly
increased the expression of Bcl-2 protein in the D+A+Rg1 group
(P<0.01; Fig. 3A and C), while
it significantly decreased the expression of Bax protein in the
D+A+Rg1 group (P<0.01; Fig. 3B and
D).
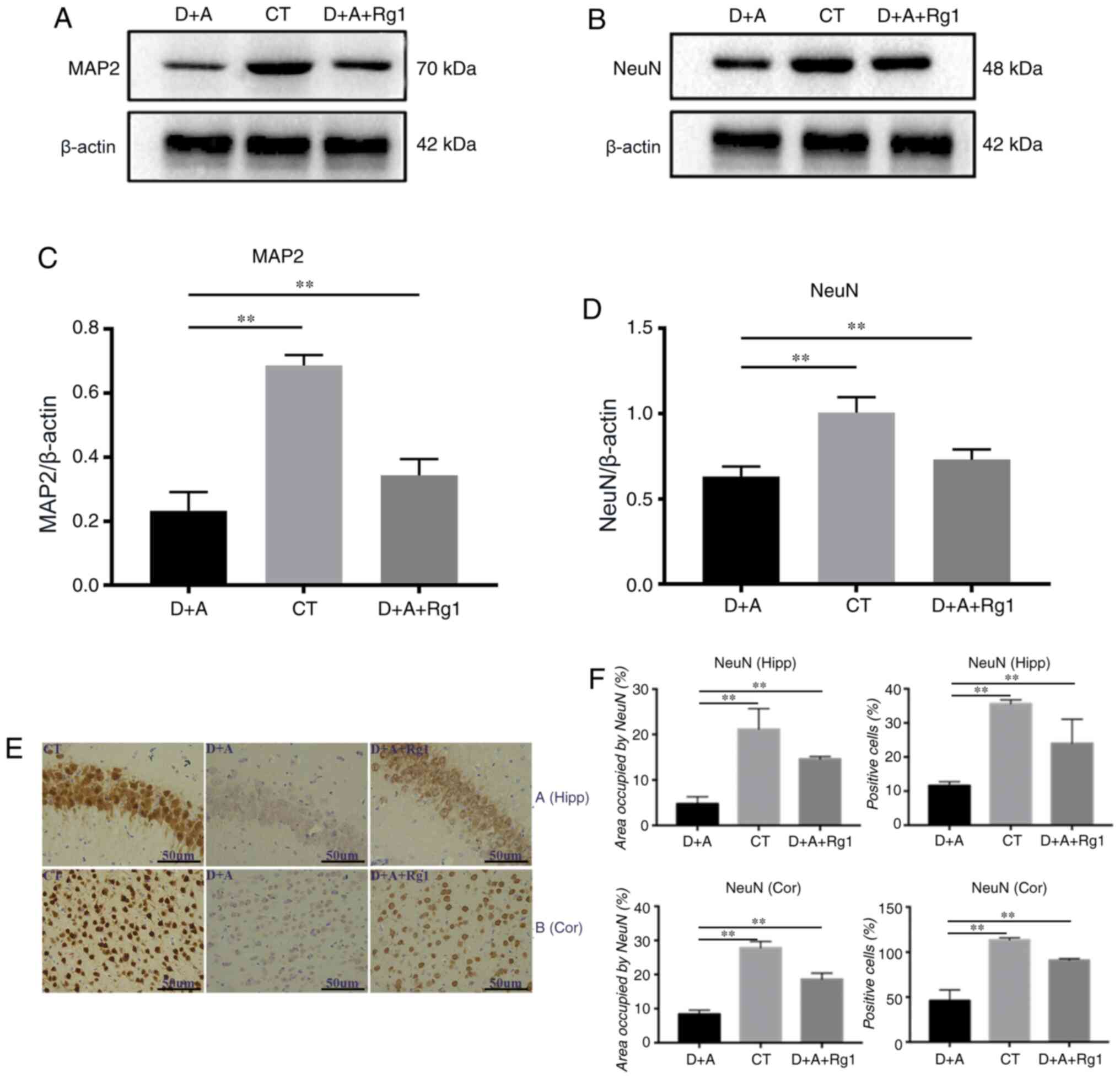
The neuron effects of Rg1 on the
hippocampus and cortex
Western blot analysis demonstrated that the
expression of MAP2 and NeuN in tree shrews were similar. The
expression of MAP2 (P<0.01) and NeuN (P<0.01) in the D+A
group were significantly decreased compared with the CT group, Rg1
significantly increased expression of MAP2 (P<0.01; Fig. 4A and C) and NeuN (P<0.01;
Fig. 4B and D).
Immunohistochemistry demonstrated that NeuN expression in the
cortex and hippocampus demonstrated that the positive cell rate and
area in the D+A+Rg1 group were significantly increased compared
with the D+A group (P<0.01; Fig. 4E
and F).
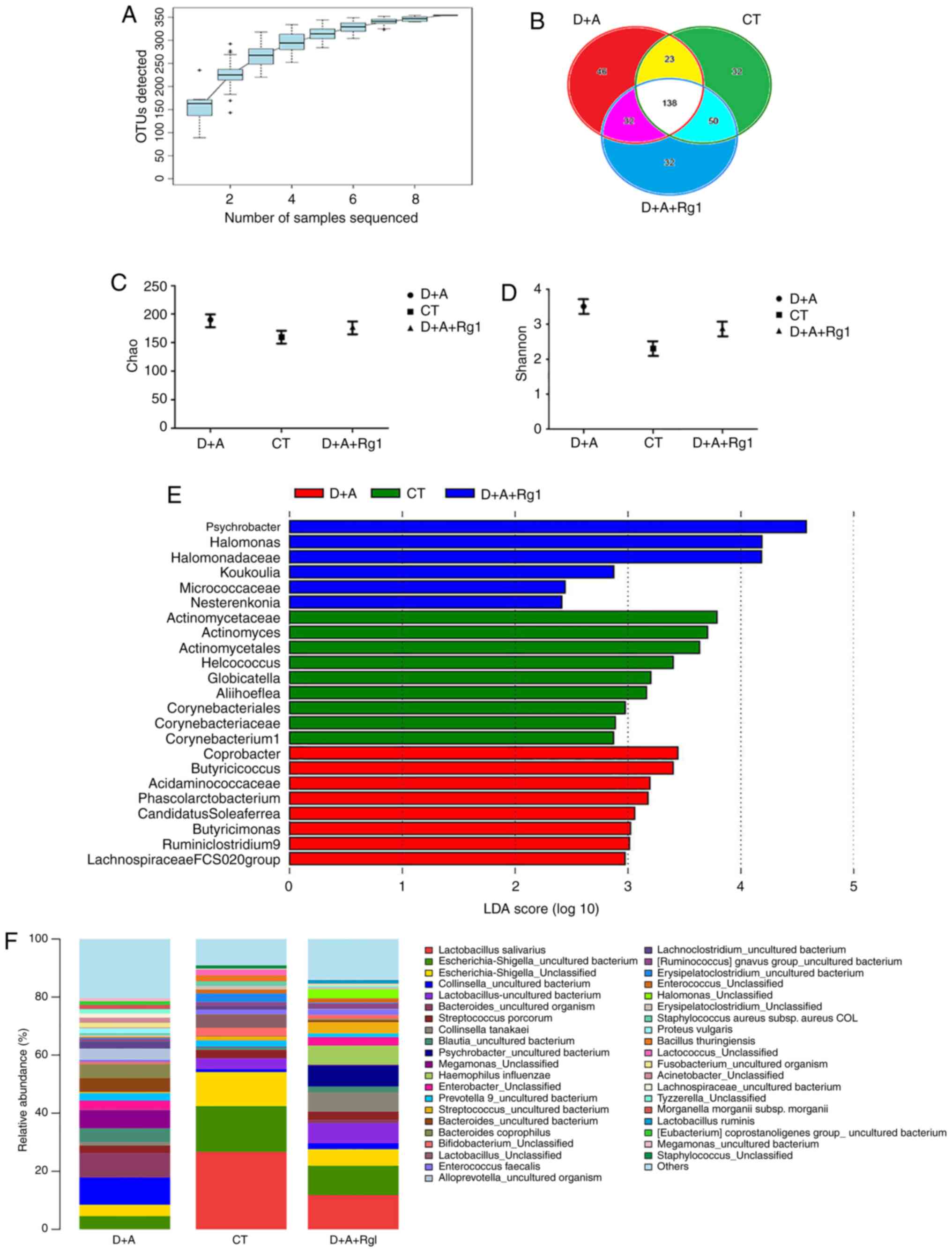
Rg1 changes the microbiota of large
intestine
A total of 398,382 sequences were acquired in the
microbiota of large intestine contents samples. OTUs were mapped at
97% similarity. The coverage index of nine samples was >0.999,
which meant that the sequencing results contained >99.9% of the
bacterial diversity in the large intestine (Fig. 5A). A Venn diagram demonstrated that,
among all 353 OTUs, all samples shared 138 OTUs; the CT group was
32 OTUs, the D+A group was 46 OTUs and the D+A+Rg1 was 32 OTUs
(Fig. 5B). These results suggested
that the bacteria species in D+A+Rg1 group were similar to the CT
group. The larger the Chao value, the larger total number of
species; the larger Shannon value, the higher community diversity
(Fig. 5C and D). No significant
difference was observed between the Chao and Shannon indexes among
the three groups (P>0.05). The D+A group exhibited higher Chao
and Shannon indexes compared with the CT group, while the results
from the D+A+Rg1 group were closer to the CT group. LEfSe analysis
revealed nine genera that were differentially represented in the CT
group compared with other groups; and there were 8 genera in the
D+A group and six genera in the D+A+Rg1 group (P<0.05; Fig. 5E). The abundance of Lactobacillus
salivarius in the D+A group was the lowest among the groups and
highest in the CT group. The abundance of Lactobacillus
salivarius in the D+A+Rg1 group was higher than D+A group,
which indicated that Rg1 could increase the abundance of
Lactobacillus salivarius in the intestinal flora of AD tree
shrews (Fig. 5F).
Meta Stat difference analysis at the phylum level
demonstrated that the abundance of Bacteroidetes in the D+A
group was significantly higher compared with the D+A+Rg1 group
(P<0.01); no significant difference was observed between the
D+A+Rg1 and CT groups (Table I).
The Firmicutes/Bacteroides ratio of the D+A and CT group was
1.3:24.7 and the Firmicutes/Bacteroides ratio in the D+A and
D+A+Rg1 group was 1.3:9. The ratio of
Firmicutes/Bacteroidetes in tree shrews in the D+A group was
only 0.0526 times higher than the CT group. Following Rg1
treatment, the Firmicutes/Bacteroides ratio in the
intestinal flora of tree shrews increased to 6.923 times that of
the D+A group. At the class level, the abundance of Bacteroidia in
group D+A was significantly higher compared with the CT group
(P<0.01) and the D+A+Rg1 group (P<0.01), while no significant
difference was observed between groups D+A+Rg1 and CT (Table I). The abundance of
Negativicutes in the D+A group was higher in the CT
(P<0.05) and the D+A+Rg1 groups (P<0.05), but no significant
difference was observed between the D+A+Rg1 and CT groups (Table I).
 | Table I.Composition of intestinal flora in
three groups of tree shrews at the level of phylum and class. |
Table I.
Composition of intestinal flora in
three groups of tree shrews at the level of phylum and class.
| Groups |
Bacteroidetes |
Betaproteobacteria |
Negativicutes |
|---|
| D+A |
0.2867±0.1381a |
0.2866±0.1380a |
0.0807±0.0502b |
| CT | 0.0255±0.0306 | 0.0246±0.0307 | 0.0033±0.0031 |
| D+A+Rg1 | 0.0486±0.0428 | 0.0471±0.0449 | 0.0045±0.0027 |
Discussion
Rg1 may improve the cognitive function
of AD tree shrews by interfering with Aβ1–42 formation and reducing
tau phosphorylation
In the present study, the escape latency decreased
and the number of times the platform was crossed increased
following treatment with Rg1. Therefore, Rg1 enhanced learning and
memory ability in AD tree shrews. The spatial memory ability of
mice following the Rg1 treatment group was improved and the
aggregation level of Aβ was significantly reduced (31).
The pathological changes of AD are extracellular
senile plaques formed by the deposition of Aβ and intracellular
neurofibrillary tangles formed by excessive phosphorylation of tau
protein (19). APP is produced by
the shear of BACE1 and γ-secretase (32). The majority of patients with AD have
elevated levels of BACE1, which leads to the production of
β-amyloid. BACE1 may be a potential biomarker and therapeutic
target for AD (33,34). Du et al (31) found that mice treated with Rg1
possess significantly reduced levels of Aβ aggregation and tau
phosphorylation. In general, Rg1 can prevent AD by reducing the
production of Aβ and the phosphorylation of tau (35).
Rg1 exhibits a neuroprotective effect
on AD tree shrews by inhibiting the expression of pro-apoptotic
proteins
MAP2 is an important component of the neuronal
cytoskeleton, serving an important role in different stages of
nervous system development, formation and regeneration (36). Jiang et al (37) demonstrated that the decrease of MAP2
in dendrites shows that neurodegeneration leads to neuron death.
Neurodegeneration and neuron loss are features of the hippocampus
and cortex in AD, making them potential targets for AD treatment
strategies (38). Kandimalla et
al (39) demonstrated that the
accumulation of phosphorylated tau in the hippocampus results in
abnormal mitochondrial dynamics and reduces the dendritic protein
MAP2, resulting in learning and memory impairment (40).
NeuN is often used to detect the survival rate of
cerebral cortex neurons (36).
Leplus et al (41) suggested
that increased expression of NeuN decreases neuronal loss in
transgenic AD rats. Intestinal microbiota inducing the low growth
phenotype is associated with decreases in the NeuN of humanized
gnotobiotic mice (42). The results
of the present study demonstrated that Rg1 had protective effects
on hippocampal and cortical neurons.
Bcl-2 and Bax are apoptosis-related proteins and the
main mechanism of AD causing neuron loss is apoptosis. Bcl-2 is an
anti-apoptotic protein, while Bax is a pro-apoptotic protein that
enhances programmed cell death (43). Hu et al (44) found that Bax is significantly
downregulated and Bcl-2 expression is upregulated following Rg1
treatment in rats. When damaged, the intestinal villi of a hindlimb
unloading rat model upregulates the expression of Bax while
downregulating Bcl-2, influencing the expansion of
Bacteroidetes and the decrease of Firmicutes in
intestinal microbiota (45).
Therefore, the expression of Bax and Bcl-2 are associated with
Firmicutes and Bacteroidetes.
Rg1 alters the microbiota abundance of
the large intestine in tree shrews
Previous studies have identified differences in the
intestinal microbial composition of neurodegenerative diseases and
microbial ecological imbalance may lead to disease pathogenesis
(46). The abundance of
Bacteroidetes is increased in patients with AD (47). Changes in the microbial composition
may alter signals from microorganisms to the central nervous system
(48). In the present study, the
number of Bacteroidetes in group D+A was significantly
higher compared with the CT group (P<0.01), which was consistent
with the patients with AD (9).
However, Rg1 significantly reduced the abundance of
Bacteroidetes in the intestinal tract of tree shrews and
there were no significant differences compared with the CT group.
These results indicated that Rg1 can regulate intestinal flora
disorder in AD tree shrews.
Vogt et al (47) identified that the abundance of
Firmicutes is decreased and Bacteroidetes is
increased in patients with AD. The Firmicutes/Bacteroidetes
ratio is one of the energy output indicators of microbial
fermentation (49); the higher the
ratio, the higher the energy output. The proportion of
Bacteroidetes decreases and Firmicutes increases in
newly malnourished mice at the growth stage after supplementary
feeding (50). Furthermore, the
Firmicutes/Bacteroidetes ratio increases in the intestinal
flora of obese mice (49),
increasing the ability to obtain energy from the diet, and the
amount of body fat. Zhang et al (8) noted that compared with normal mice,
the Firmicutes/Bacteroidetes ratio in the intestinal flora
of AD mice of all ages decreased. In the present study, the
Firmicutes/Bacteroidetes ratio in the tree shrews of group
D+A was only 0.0526 times that of the CT group. Following Rg1
treatment, the Firmicutes/Bacteroidetes ratio increased to
6.923 times that of the D+A group. This suggested that the lower
Firmicutes/Bacteroidetes ratio of AD tree shrews was related
to the reduced energy requirements of AD tree shrews.
Rezaeiasl et al (51) suggested that Lactobacillus
supplementation may positively effect learning capacity in rats
with AD. Lactobacillus improves the gut microbiota of a
Drosophila melanogaster AD model (52). Lactobacillus salivarius, a
gram-positive bacterium, is a type of beneficial bacteria widely
existing in the intestinal tract of human beings and animals
(53). Feeding with
Lactobacillus salivarius can extend the average life span of
Caenorhabditis elegans by 11.9% and increase the activity of
superoxide dismutase (53). In the
present study, the abundance of Lactobacillus salivarius in
the CT group was 24 times higher compared with the D+A group, while
that in the treatment group was 10 times higher than the D+A group.
It indicates that Rg1 may improve cognitive impairment by
increasing the population of Lactobacillus salivarius.
In conclusion, Rg1 can inhibit the expression of
pro-apoptotic proteins and protect neurons in a tree shrew AD
model. Rg1 also reduced the production of Aβ and tau
phosphorylation, improved cognitive impairment and the microbiota
imbalance of the large intestine. Although the mechanisms of Rg1 in
tree shrew AD models remain to be elucidated, the present study
provided clues to further elucidate the improvement of cognitive
impairment.
Acknowledgements
Not applicable.
Funding
This study was supported by the Applied Basic
Research Foundation of Yunnan Province [grant nos. 2017FE468-014,
2017FE468-016 and 2019FE001(−026)], the National Natural Science
Foundation of China (grant no. 81460647), the Scientific Research
Foundation of Yunnan Education Bureau (grant no. 2018Y048) and the
Innovation Project of Kunming Medical University (grant no.
2019S093).
Availability of data and materials
The 16S rRNA database analyzed during the current
study are available in the NCBI repository (SRP: 227299; https://www.ncbi.nlm.nih.gov/sra.
Authors contributions
YG and LW performed the experiments, and were major
contributors in writing the original draft. JL and YY analyzed the
data. JJ and HZ helped with data analysis and designed and produced
the graphs and edited the manuscript. ZL and HZ conceived and
designed the study. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The animal experiment protocol was approved by the
Committee on the Ethics of Animal Experiments of Kunming Medical
University (approval number KMMU2018004).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Quigley EMM: Microbiota-brain-gut axis and
neurodegenerative diseases. Curr Neurol Neurosci Rep. 17:942017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lecouvey G, Morand A, Gonneaud J, Piolino
P, Orriols E, Pélerin A, Ferreira Da Silva L, de La Sayette V,
Eustache F and Desgranges B: An impairment of prospective memory in
mild Alzheimer's disease: A ride in a virtual town. Front Psychol.
10:241. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Manso-Calderón R, Cacabelos-Pérez P,
Sevillano-García MD, Herrero-Prieto ME and González-Sarmiento R:
The impact of vascular burden on behavioural and psychological
symptoms in older adults with dementia: The BEVASDE study. Neurol
Sci. 41:165–174. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ratno Budiarto B and Chan WH: Oxidative
stresses-mediated apoptotic effects of ginsenoside Rb1 on pre- and
post-implantation mouse embryos in vitro and in vivo. Environ
Toxicol. 32:1990–2003. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tian W, Chen L, Zhang L, Wang B, Li XB,
Fan KR, Ai CH, Xia X, Li SD and Li Y: Effects of ginsenoside Rg1 on
glucose metabolism and liver injury in streptozotocin-induced type
2 diabetic rats. Genet Mol Res. 16:March 30–2017, https://doi.org/10.4238/gmr16019463
View Article : Google Scholar
|
|
6
|
Ong WY, Farooqui T, Koh HL, Farooqui AA
and Ling EA: Protective effects of ginseng on neurological
disorders. Front Aging Neurosci. 7:1292015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Chen LM, Lin N, Zhang J, Zhu YG and Chen
XC: Mechanism of ginsenoside Rg1 regulating the activity of β
secretase in N2a/APP695 cells. Zhonghua Yi Xue Za Zhi. 92:330–335.
2012.(In Chinese). PubMed/NCBI
|
|
8
|
Zhang L, Wang Y, Xiayu X, Shi C, Chen W,
Song N, Fu X, Zhou R, Xu YF, Huang L, et al: Altered gut microbiota
in a mouse model of Alzheimer's disease. J Alzheimer's Dis.
60:1241–1257. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Claesson MJ, Cusack S, OSullivan O,
Greene-Diniz R, de Weerd H, Flannery E, Marchesi JR, Falush D,
Dinan T, Fitzgerald G, et al: Composition, variability, and
temporal stability of the intestinal microbiota of the elderly.
Proc Natl Acad Sci USA. 108 (Suppl 1):4586–4591. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Claesson MJ, Jeffery IB, Conde S, Power
SE, OConnor EM, Cusack S, Harris HMB, Coakley M, Lakshminarayanan
B, OSullivan O, et al: Gut microbiota composition correlates with
diet and health in the elderly. Nature. 488:178–184. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Odamaki T, Kato K, Sugahara H, Hashikura
N, Takahashi S, Xiao JZ, Abe F and Osawa R: Age-related changes in
gut microbiota composition from newborn to centenarian: A
cross-sectional study. BMC Microbiol. 16:902016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McElhanon BO, McCracken C, Karpen S and
Sharp WG: Gastrointestinal symptoms in autism spectrum disorder: A
meta-analysis. Pediatrics. 133:872–883. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pfeiffer RF: Gastrointestinal dysfunction
in Parkinsons disease. Lancet Neurol. 2:107–116. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Severance EG, Yolken RH and Eaton WW:
Autoimmune diseases, gastrointestinal disorders and the microbiome
in schizophrenia: More than a gut feeling. Schizophr Res.
176:23–35. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Doraiswamy PM, Leon J, Cummings JL, Marin
D and Neumann PJ: Prevalence and impact of medical comorbidity in
Alzheimer's disease. J Gerontol A Biol Sci Med Sci. 57:M173–M177.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kowalski K and Mulak A:
Brain-gut-microbiota axis in Alzheimer's disease. J
Neurogastroenterol Motil. 25:48–60. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mosher KI and Wyss-Coray T: Microglial
dysfunction in brain aging and Alzheimer's disease. Biochem
Pharmacol. 88:594–604. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen SG, Stribinskis V, Rane MJ, Demuth
DR, Gozal E, Roberts AM, Jagadapillai R, Liu R, Choe K, Shivakumar
B, et al: Exposure to the functional bacterial amyloid protein
curli enhances alpha-synuclein aggregation in aged fischer 344 rats
and Caenorhabditis elegans. Sci Rep. 6:34477. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Minter MR, Zhang C, Leone V, Ringus DL,
Zhang X, Oyler-Castrillo P, Musch MW, Liao F, Ward JF, Holtzman DM,
et al: Antibiotic-induced perturbations in gut microbial diversity
influences neuro-inflammation and amyloidosis in a murine model of
Alzheimer's disease. Sci Rep. 6:30028. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Harach T, Marungruang N, Duthilleul N,
Cheatham V, Mc Coy KD, Frisoni G, Neher JJ, Fåk F, Jucker M, Lasser
T, et al: Reduction of Abeta amyloid pathology in APPPS1 transgenic
mice in the absence of gut microbiota. Sci Rep. 7:41802. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang CH, Sheng JQ, Sarsaiya S, Shu FX,
Liu TT, Tu XY, Ma GQ, Xu GL, Zheng HX and Zhou LF: The
anti-diabetic activities, gut microbiota composition, the
anti-inflammatory effects of Scutellaria-coptis herb couple against
insulin resistance-model of diabetes involving the toll-like
receptor 4 signaling pathway. J Ethnopharmacol. 237:202–214. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun YF, Zhang X, Wang XY and Jia W: Effect
of long-term intake of ginseng extracts on gut microbiota in rats.
Zhongguo Zhong Yao Za Zhi. 43:3927–3932. 2018.(In Chinese).
PubMed/NCBI
|
|
23
|
Cheng D, Chang H, Ma S, Guo J, She G,
Zhang F, Li L, Li X and Lu Y: Tiansi liquid modulates gut
microbiota composition and tryptophan-kynurenine metabolism in rats
with hydrocortisone-induced depression. Molecules. 23:28322018.
View Article : Google Scholar
|
|
24
|
Xiao J, Liu R and Chen CS: Tree shrew
(Tupaia belangeri) as a novel laboratory disease animal
model. Zool Res. 38:127–137. 2017.PubMed/NCBI
|
|
25
|
Fan Y, Luo R, Su LY, Xiang Q, Yu D, Xu L,
Chen JQ, Bi R, Wu DD, Zheng P, et al: Does the genetic feature of
the Chinese Tree Shrew (Tupaia belangeri chinensis) support
its potential as a viable model for Alzheimer's disease research? J
Alzheimer's Dis. 61:1015–1028. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zheng H, Niu S, Zhao H, Li S and Jiao J:
Donepezil improves the cognitive impairment in a tree shrew model
of Alzheimer's disease induced by amyloid-β1–40 via activating the
BDNF/TrkB signal pathway. Metab Brain Dis. 33:1961–1974. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Römer S, Bender H, Knabe W, Zimmermann E,
Rübsamen R, Seeger J and Fietz SA: Neural progenitors in the
developing Neocortex of the Northern Tree Shrew (Tupaia
belangeri) show a closer relationship to gyrencephalic primates
than to lissencephalic rodents. Front Neuroanat. 12:292018.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Health N: Guide for the care and use of
laboratory animals. NIH contract No No1-RR-2-2135. 1985.11–28
|
|
29
|
Ghumatkar PJ, Patil SP, Peshattiwar V,
Vijaykumar T, Dighe V, Vanage G and Sathaye S: The modulatory role
of phloretin in Aβ25–35 induced sporadic Alzheimer's disease in rat
model. Naunyn Schmiedebergs Arch Pharmacol. 392:327–339. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chao A: Non-parametric estimation of the
classes in a population. Scand J Stat. 11:265–270. 1984.
|
|
31
|
Du H, Guo L, Zhang W, Rydzewska M and Yan
S: Cyclophilin D deficiency improves mitochondrial function and
learning/memory in aging Alzheimer disease mouse model. Neurobiol
Aging. 32:398–406. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Feng L, Liao YT, He JC, Xie CL, Chen SY,
Fan HH, Su ZP and Wang Z: Plasma long non-coding RNA BACE1 as a
novel biomarker for diagnosis of Alzheimer disease. BMC Neurol.
18:42018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mulder SD, van der Flier WM, Verheijen JH,
Mulder C, Scheltens P, Blankenstein MA, Hack CE and Veerhuis R:
BACE1 activity in cerebrospinal fluid and its relation to markers
of AD pathology. J Alzheimer's Dis. 20:253–260. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dash R, Emran TB, Uddin MMN, Islam A and
Junaid M: Molecular docking of fisetin with AD associated AChE,
ABAD and BACE1 proteins. Bioinformation. 10:562–568. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Song XY, Hu JF, Chu SF, Zhang Z, Xu S,
Yuan YH, Han N, Liu Y, Niu F, He X, et al: Ginsenoside Rg1
attenuates okadaic acid induced spatial memory impairment by the
GSK3β/tau signaling pathway and the Aβ formation prevention in
rats. Eur J Pharmacol. 710:29–38. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Manczak M, Kandimalla R, Yin X and Reddy
PH: Hippocampal mutant APP and amyloid beta-induced cognitive
decline, dendritic spine loss, defective autophagy, mitophagy and
mitochondrial abnormalities in a mouse model of Alzheimer's
disease. Hum Mol Genet. 27:1332–1342. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang S, Nandy P, Wang W, Ma X, Hsia J,
Wang C, Wang Z, Niu M, Siedlak SL, Torres S, et al: Mfn2 ablation
causes an oxidative stress response and eventual neuronal death in
the hippocampus and cortex. Mol Neurodegener. 13:52018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lax N, Fainstein N, Nishri Y, Ben-Zvi A
and Ben-Hur T: Systemic microbial TLR2 agonists induce
neurodegeneration in Alzheimer's disease mice. J Neuroinflammation.
17:552020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kandimalla R, Manczak M, Yin X, Wang R and
Reddy PH: Hippocampal phosphorylated tau induced cognitive decline,
dendritic spine loss and mitochondrial abnormalities in a mouse
model of Alzheimer's disease. Hum Mol Genet. 27:30–40. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Koo YS, Kim H, Park JH, Kim MJ, Shin YI,
Choi BT, Lee SY and Shin HK: Indoleamine 2,3-dioxygenase-dependent
neurotoxic kynurenine metabolism contributes to poststroke
depression Induced in mice by ischemic stroke along with spatial
restraint stress. Oxid Med Cell Longev. 2018:24138412018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Leplus A, Lauritzen I, Melon C,
Kerkerian-Le Goff L, Fontaine D and Checler F: Chronic fornix deep
brain stimulation in a transgenic Alzheimer's rat model reduces
amyloid burden, inflammation, and neuronal loss. Brain Struct
Funct. 224:363–372. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lu J, Lu L, Yu Y, Cluette-Brown J, Martin
CR and Claud EC: Effects of intestinal microbiota on brain
development in humanized gnotobiotic mice. Sci Rep. 8:54432018.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhong L, Tong Y, Chuan J, Bai L, Shi J and
Zhu Y: Protective effect of ethyl vanillin against Aβ-induced
neurotoxicity in PC12 cells via the reduction of oxidative stress
and apoptosis. Exp Ther Med. 17:2666–2674. 2019.PubMed/NCBI
|
|
44
|
Hu J, Gu Y and Fan W: Rg1 protects rat
bone marrow stem cells against hydrogen peroxide-induced cell
apoptosis through the PI3K/Akt pathway. Mol Med Rep. 14:406–412.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Jin M, Zhang H, Zhao K, Xu C, Shao D,
Huang Q, Shi J and Yang H: Responses of intestinal iucosal barrier
functions of rats to simulated eeightlessness. Front Physiol.
9:7292018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sharon G, Sampson TR, Geschwind DH and
Mazmanian SK: The central nervous system and the gut microbiome.
Cell. 167:915–932. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Vogt NM, Kerby RL, Dill-McFarland KA,
Harding SJ, Merluzzi AP, Johnson SC, Carlsson CM, Asthana S,
Zetterberg H, Blennow K, et al: Gut microbiome alterations in
Alzheimer's disease. Sci Rep. 7:13537. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Heiss CN and Olofsson LE: The role of the
gut microbiota in development, function and disorders of the
central nervous system and the enteric nervous system. J
Neuroendocrinol. 31:e126842019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini
V, Mardis ER and Gordon JI: An obesity-associated gut microbiome
with increased capacity for energy harvest. Nature. 444:1027–1031.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Preidis GA, Ajami NJ, Wong MC, Bessard BC,
Conner ME and Petrosino JF: Microbial-derived metabolites reflect
an altered intestinal microbiota during catch-up growth in
undernourished neonatal mice. J Nutr. 146:940–948. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Rezaeiasl Z, Salami M and Sepehri G: The
effects of probiotic Lactobacillus and
Bifidobacterium strains on memory and learning behavior,
long-term potentiation (LTP), and some biochemical parameters in
β-amyloid-induced rats model of Alzheimer's disease. Prev Nutr Food
Sci. 24:265–273. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Tan FHP, Liu G, Lau SA, Jaafar MH, Park
YH, Azzam G, Li Y and Liong MT: Lactobacillus probiotics improved
the gut microbiota profile of a Drosophila melanogaster
Alzheimer's disease model and alleviated neurodegeneration in the
eye. Benef Microbes. 11:79–89. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhao Y, Zhao L, Zheng X, Fu T, Guo H and
Ren F: Lactobacillus salivarius strain FDB89 induced
longevity in Caenorhabditis elegans by dietary restriction.
J Microbiol. 51:183–188. 2013. View Article : Google Scholar : PubMed/NCBI
|