Introduction
Male fertility relies on the continuous production
of sperm through a complex developmental process termed
spermatogenesis. This process involves the differentiation of
spermatogonia into spermatocytes, the development of spermatocytes
into round spermatids through two successive meiotic divisions and
the final elongation of spermatids (1,2). The
process of spermatid elongation is also known as spermiogenesis and
involves the differentiation of round spermatids into sperm via
marked morphological changes. Spermiogenesis includes chromatin
condensation, elongation of the nucleus, biogenesis of the
acrosome, extension of the flagellum and removal of excess
cytoplasm (3–5).
The acrosome is a unique cap-like organelle that is
derived from the Golgi apparatus and gradually expands to cover the
apical region of the sperm nucleus (6). Formation of the acrosome is one of the
most readily detectable changes in the seminiferous tubules
(7), and the process responsible
for acrosomal development has been well characterized. First,
proacrosomal granules originating from the trans-Golgi stacks
assemble in the concave region of the nuclear surface and fuse with
each other to form a single acrosomal granule. This subsequently
enlarges and spreads to form a cap over the nucleus. Finally, the
acrosome migrates over the surface of the sperm nucleus and forms
its characteristic shape (8–10).
Microtubule structure plays an important role in
spermiogenesis. For example, the manchette, a temporary
microtubular structure, first appears in step-8 spermatids but
subsequently disappears as spermiogenesis enters steps 13 and 14
(2). During the elongation of
spermatids, the manchette shapes the distal half of the head and
facilitates the intracellular transport of proteins to the
flagellum (11,12).
A previous study reported that >2,300 genes are
expressed predominantly in mouse testes (13), and a number of them are identified
as serving important roles in numerous events during spermiogenesis
through gene-editing technology, such as the biogenesis of the
acrosome, the morphology of the manchette and the assembly of the
flagellum. For example, mice lacking autophagy related 7, sirtuin
1, serine-rich single-pass membrane protein 1, vacuolar protein
sorting (Vps)13b, Vps54 and sperm acrosome associated 1
exhibit abnormal round-headed sperm as a result of aberrant
acrosome formation (8,9,14–16).
Leucine-rich repeats and guanylate kinase domain containing isoform
1, serine/threonine kinase 22, katanin-like 2, sperm flagellar 2,
hook microtubule tethering protein 1 and calcium and integrin
binding family member 4 are essential for the correct formation of
the manchette; deficiency in any of these genes can lead to
abnormal elongation of the manchette and thus, a malformed head or
flagellum (11,12,17–20).
In addition, the deletion of sperm associated antigen 16, kinesin
light chain 3, kinase suppressor of ras 2, testis-specific serine
kinase 4 or genes of the septin (Sep) family (Sept2,
Sept4 and Sept12) causes alterations in the
intra-flagellum structure and reduces sperm motility (21–25).
Despite the crucial function of these genes in spermiogenesis, the
localization and functional requirement of other genes requires
further characterization.
Testis-expressed protein 33 (TEX33), also known as
Ean57, 1700061J05Rik, cE81G9.2 and C22orf33, is evolutionarily
conserved. Orthologs of this protein have been identified in a
number of different species, including humans, mice, Norwegian
rats, zebrafish and Rhesus monkeys. A previous bioinformatics
analysis revealed that TEX33 was expressed abundantly in the testis
and associated with the formation of cilia (26). Therefore, we hypothesized that TEX33
may be a testis-enriched or specific protein that localizes to the
sperm flagellum and is potentially associated with
spermiogenesis.
The aim of the present study was to investigate the
function of TEX33 in mouse spermiogenesis and fertility using
CRISPR/Cas9 gene-editing. It is hoped that the results will enrich
our understanding of the molecular networks underlying
spermiogenesis and provide new insight into the genetics of male
fertility.
Materials and methods
Animals
A total of three male C57BL/6N mice aged 10 weeks
and two female C57BL/6N mice aged 8 weeks were purchased from
Cyagen Biosciences, Inc. The CRISPR/Cas9 system was used to
generate Tex33 knockout mice. Two guide RNAs (gRNAs) were designed
using the E-CRISPR database (http://www.e-crisp.org/E-CRISP/designcrispr.html)
to target exons 2–4 of Tex33: gRNA1, TACCAGAATCATCTAGTCCCTGG
(matching the forward strand of the gene) and gRNA2,
GCTAGCCAAGGCCAACACCTGGG (matching the reverse strand of the gene).
The two gRNAs and Cas9 NLS endonuclease (cat. no. M0646M; New
England BioLabs, Inc.) were microinjected into the cytoplasm of
fertilized eggs from C57BL/6N mice. These eggs were then implanted
into the oviducts of pseudo-pregnant female mice. Founder mice were
then crossed with C57BL/6N mice to generate
Tex33+/− offspring. Finally,
Tex33−/− mice were obtained by breeding from the
Tex33+/− mice.
All mice were maintained under specific
pathogen-free conditions with free access to standard mouse lab
pellet food and water, and housed at 20–22°C and 50–70% humidity
with a 12-h light/dark cycle. All animal experiments were approved
by the Animal Ethics Committee of Yangzhou University.
Reverse transcription-polymerase chain
reaction (RT-PCR)
For RNA extraction, heart, brain, spleen, liver,
lung, kidney and trachea were collected from adult wild-type mice
(n=3) and testes were harvested from wild-type mice at different
spermatogenesis stages (postnatal days 7, 14, 21, 28, 35 and 56;
n=3 each). Total RNAs were isolated on ice using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) and then
dissolved in RNase-free distilled water. The purity of the total
RNAs was determined using ultraviolet spectrophotometry (Eppendorf)
and agarose gel electrophoresis. Only RNA samples with a ratio of
absorbance at 260 and 280 nm of ≥1.8 were used for RT-PCR. For
reverse transcription, 500 ng total RNA in a final volume of 10 µl
was mixed with 5X PrimeScript RT Master Mix (Takara Bio, Inc.) and
RNase-free distilled water. For cDNA synthesis, the mixture was
incubated at 37°C for 15 min, and then at 85°C for 15 sec.
Tex33 transcript variants were then amplified by PCR using
the following primers: Tex33 transcript variant V1:
5′-GGTTCATTTGCCTTCTTCCC-3′ (forward), 5′-TCCTTGTCGTTCGGGTGTG-3′
(reverse); Tex33 transcript variant V2: 5′-ACCTGGGGAGA
GCTGCTGCTCG-3′ (forward), 5′-GTTGGTGGTAGAGGT GGAG-3′ (reverse);
Tex33 transcript variant V3: 5′-TGGGTCA GGAAGGAGTGTG-3′
(forward), 5′-GTTGGTGGTAG AGGTGGAG-3′ (reverse). Glyceraldehyde
3-phosphate dehydrogenase (GAPDH) was used as a positive
control using the following primer sequences: 5′-GACGGCCGCATCTTCT
TGT-3′ (forward) and 5′-ACACCGACCTTCACCATTTT GT-3′ (reverse). PCR
was performed using a Taq DNA polymerase kit (cat. no. M1661S;
Promega Corporation). Amplification conditions for Tex33
transcript variant V1, V2 and V3 were as follows: V1, initial
denaturation step at 95°C for 5 min followed by 35 cycles of at
95°C for 30 sec, an annealing step at 60°C for 45 sec, an extension
step at 72°C for 45 sec, and a final extension at 72°C for 10 min;
V2, initial denaturation at 95°C for 5 min, followed by 35 cycles
at 95°C for 30 sec, 58°C for 30 sec, 72°C for 45 sec, and
elongation at 72°C for 10 min; V3, an initial denaturation step of
5 min at 95°C, followed by 35 cycles 95°C for 30 sec, 52°C for 30
sec, 72°C for 45 sec, ended by a final elongation at 72°C for 10
min. PCR protocol for GAPDH included a 5 min denaturation
step at 95°C followed by 35 cycles of 50 sec at 95°C, 1 min
annealing step at 55°C and 1 min extension at 72°C and a final
elongation at 72°C for 10 min. PCR products were electrophoresed on
1.2% agarose gels using 1X Tris/Borate/EDTA buffer (TBE). Gels were
stained with SYBR Safe™ (Thermo Fisher Scientific, Inc.) and the
bands visualized using a gel imaging system (Tanon Science and
Technology Co., Ltd.).
Synthesis of an anti-TEX33
antibody
PCR was used to amplify the open reading fragment
(ORF) of Tex33 transcript variant V2 (encoding 266 amino
acids), which represents the predominant transcript according to
the GenBank database and shares the same ORF as V1. The following
primers were used: 5′-GGGAATTCATGGAGCTGAGCCACCGAC-3′ (forward) and
5′-GGAAGCTTGCTTGGCCTTGGATCTGCTC-3′ (reverse). The PCR product was
purified with a DNA gel extraction kit (Corning, Inc.), sequenced
by next generation sequencing (BGI Group) and finally subcloned
into a pET-30a (+) vector (Novagen; Merck KGaA). After induction
with isopropyl β-D-thiogalactoside for 12–14 h at 37°C, the
prokaryotic protein samples were analyzed by electrophoresis on 10%
sodium dodecyl sulfate (SDS) polyacrylamide gels using Coomassie
Brilliant Blue solution. The TEX33 prokaryotic protein was
successfully expressed in Escherichia coli BL21 (DE3) plysS
(Novagen; Merck KGaA). The resulting His-tag fusion protein was
purified using Ni-NTA affinity resin (RuiXiang Bio Inc.) in
accordance with the manufacturers instructions. The concentration
of the purified protein was determined using a bicinchoninic acid
(BCA) kit (CWBio).
A total of 5 male New Zealand white rabbits (~2 kg
body weight) aged 10 weeks were used for animal immunization. They
were purchased from Laboratory Animal Center of Shanghai Branch of
Chinese Academy of Science and maintained in individual cages at
22°C, 42% humidity, with a 12-h light/dark cycle and free access to
food and water. Purified protein (0.75 mg/ml) was mixed with an
equal volume of Freunds adjuvant (Sigma-Aldrich; Merck KGaA) and
500 µl purified protein/adjuvant mixture was subsequently injected
into the backs of adult male rabbits at 0, 2, 4, 6 and 8 weeks.
After the last immunization, blood was harvested from each rabbit
and the serum was separated. The antibody titer from each rabbit
was determined using an enzyme-linked immunosorbent assay (ELISA;
Fig. S1A).
For ELISA, 5 µg/ml purified TEX33 prokaryotic
protein was added to each well of a 96-well plate (Corning Inc.)
and incubated at 4°C overnight. After washing with 0.05% Tween-20
solution in phosphate-buffered saline (PBS-T), each well was
blocked with 1% bovine serum albumin (BSA); Beijing Solarbio
Science & Technology Co., Ltd.) for 1 h at room temperature.
After washing, 1:10, 1:100, 1:1,000, 1:10,000, 1:100,000, and
1:1,000,000 dilutions of serum were added and the plate was
incubated at 37°C for 1 h. The wells were then washed thrice with
PBS-T and incubated with goat-anti-rabbit IgG-HRP secondary
antibody (cat. no. ZB-2301; 1:2,500 dilution; OriGene Technologies,
Inc.) at 37°C for 1 h. TMB Two-Component Substrate solution
(Beijing Solarbio Science & Technology Co., Ltd.) was then
added and allowed to react for 10 min at room temperature in the
dark. The reaction was stopped with 1 mM hydrochloric acid and the
absorbance was measured at 450 nm using a Multiskan FC microplate
reader (Thermo Fisher Scientific, Inc.). The specificity of the
antibody was confirmed by western blot (Fig. S1B) and immunofluorescence analyses
(Fig. S1C-F).
Western blot analysis
For protein extraction, heart, brain, spleen, liver,
lung, kidney and trachea were collected from multiple tissues of
adult wild-type mice (n=3) and testes were harvested from wild-type
mice at different spermatogenesis stages (postnatal days 7, 14, 21,
28, 35 and 56; n=3 each). Tissues were lysed in RIPA buffer (CWBio)
on ice, followed by sonication on ice for 10 min. After
centrifugation at 14,000 × g for 10 min at 4°C, the supernatants
were collected. The protein concentration was determined using a
BCA kit. Protein samples (40 µg) were then separated on 10%
SDS-polyacrylamide gels and transferred onto polyvinylidene
fluoride membranes (Millipore; Merck KGaA). Non-specific antibody
binding was blocked using 5% non-fat milk in Tris-buffered saline
(TBS) for 1 h at room temperature. Membranes were then incubated at
4°C overnight with the following primary antibodies: TEX33 antibody
(1:1,000 dilution; generated in the present study), GAPDH antibody
(cat. no. CW0100M; 1:2,500 dilution; CWBio) and β-actin antibody
(cat. no. ET1701-80; 1:2,500 dilution; HuaBio). After washing with
0.05% TBS-T, membranes were incubated with goat-anti-rabbit IgG-HRP
secondary antibody (cat. no. ZB-2301; 1:2,500 dilution; OriGene
Technologies, Inc.) for 1 h at room temperature. After washing
three times with TBS-T, protein bands were visualized with enhanced
chemiluminescence reagent (New Cell and Molecular Biotech Co.,
Ltd.) using an automatic chemiluminescence image analysis system
(Tanon Science and Technology Co., Ltd.).
Immunofluorescence analysis
In order to analyze tissue sections by
immunofluorescence, testicular samples were collected from the mice
and fixed in 4% paraformaldehyde solution (pH 7.0) for 24–48 h at
4°C. The testicular samples were dehydrated in an ascending alcohol
gradient, then cleared in chloroform overnight. The cleared samples
were infiltrated in xylene, embedded in paraffin and cut into 4-µm
sections. The sections were rehydrated in a descending alcohol
gradient followed by antigen retrieval in 10 mM sodium citrate (pH
6.0) at 100°C for 10 min. After blocking with 1% BSA for 1 h at
room temperature, the sections were incubated with primary
antibodies at 4°C overnight. The following primary antibodies were
used: TEX33 antibody (1:200 dilution; generated in this study) and
mouse α-tubulin antibody (cat. no. AC012; 1:100 dilution; ABclonal
Biotech Co., Ltd.). After washing with PBS, sections were further
incubated with Alexa Fluor 488-conjugated donkey-anti-mouse
secondary antibody (cat. no. A32766TR; 1:1,000 dilution; Thermo
Fisher Scientific, Inc.) and Alexa Fluor 555-conjugated
donkey-anti-rabbit secondary antibody (cat. no. A-31572; 1:1,000
dilution; Thermo Fisher Scientific, Inc.) for 1 h at room
temperature, and then counterstained with
4,6-diamidino-2-phenylindole (DAPI) dye (cat. no. C0060; 1:1,000
dilution; Beijing Solarbio Science & Technology Co., Ltd.) for
10 min at room temperature. Fluorescence intensity was finally
detected by confocal laser-scanning microscopy (Carl Zeiss AG).
In order to investigate sperm using
immunocytochemistry, the cauda epididymis of each mouse was cut
into small pieces to allow motile sperm to swim out. After
filtration and centrifugation at 800 × g for 15 min at room
temperature, the sperm sample was rinsed with PBS, incubated in PBS
at 37°C for 15 min, smeared on slides and fixed with 4%
paraformaldehyde solution for 30 min at room temperature. The
slides were blocked with donkey serum (cat. no. SL050; 1:100
dilution; Beijing Solarbio Science & Technology Co., Ltd.) at
room temperature for 1 h, and then incubated with anti-TEX33
primary antibody (1:100 dilution; generated in the present study)
at 4°C overnight. This was followed by incubation with
FITC-conjugated peanut agglutinin (PNA-FITC; cat. no. L7381; 1:50
dilution; Sigma-Aldrich; Merck KGaA) and Alexa Fluor 555
donkey-anti-rabbit secondary antibody (cat. no. A-31572; 1:1,000
dilution). After washing three times with PBS, the nuclei were
stained with DAPI for 10 min at room temperature. Images were
captured by confocal laser-scanning microscopy.
Mouse genotyping
Mouse tail samples (0.2–0.3 cm) were obtained and
lysed at 55°C overnight in lysis buffer containing 1 mM Tris-HCl
(pH 8.0), 1 mM NaCl, 1 mM EDTA (pH 8.0), 0.5% SDS and 10 µg/µl
proteinase K (Sigma-Aldrich; Merck KGaA). Genomic DNA was extracted
from the lysate using phenol and chloroform (Beijing Solarbio
Science & Technology Co., Ltd.), precipitated with isopropanol
(Sinopharm Chemical Reagent Co., Ltd.) and dissolved in distilled
water at 60°C. Subsequent PCR was performed using a Taq DNA
polymerase kit (cat. no. M1661S; Promega Corporation) according to
the manufacturers instruction. Primers for genotyping were
designed: 5′-CAGCCGATGCTTCTATGACAAC-3′ (forward),
5′-TCCCACATTTTCCGCAGGTG-3′ (reverse 1), 5′-TCTGGGCTCCGTAATGTCTG-3′
(reverse 2). The PCR conditions were as follows: Denaturation at
94°C for 5 min, followed by 35 cycles at 94°C for 50 sec, 55°C for
1 min, 65°C for 50 sec, and a final extension at 65°C for 10 min.
PCR products were separated on 1.2% agarose gels by electrophoreses
in 1X TBE buffer. The products were visualized using a gel imaging
system (Tanon Science and Technology Co., Ltd.) after staining with
SYBR Safe™ (Thermo Fisher Scientific, Inc.).
Histological analysis
Paraffin sections (~4 µm) fixed in Bouins solution
(including testes, cauda epididymides and ovaries) were used for
both histological and morphological analyses. The sections were
dewaxed with xylene, and then rehydrated with 100, 95, 90, 80 and
70% alcohol for 2 min each. After washing with RNase-free distilled
water, the sections were used for hematoxylin and eosin (H&E)
or periodic acid-schiff (PAS) staining.
For H&E staining, the sections were first
stained with hematoxylin (Beyotime Institute of Biotechnology) for
2 min at room temperature, followed by 6 dips in 1% hydrochloric
alcohol and rinsing in RNase-free distilled water. The sections
were then stained with eosin (Beyotime Institute of Biotechnology)
for 3 sec at room temperature, followed by dehydration in 70, 90
and 100% alcohol for 1 min each. Finally, the sections were
immersed in xylene for 5 min. Images were visualized by light
microscopy (Nikon-70i; Nikon Corporation).
For PAS staining, a Schiffs kit (cat. no. G1360;
Beijing Solarbio Science & Technology Co., Ltd.) was used in
accordance with the manufacturers protocols. In brief, the sections
were pretreated with periodic acid for 5 min at room temperature,
slowly rinsed in RNase-free distilled water and then stained with
Schiffs solution for 10 min at room temperature in the dark. The
nuclei were stained with hematoxylin for 5 min at room temperature,
followed by 6 dips in 1% hydrochloric alcohol. After dehydration
with 70, 90 and 100% graded alcohol, the sections were immersed
twice in xylene for 10 min each. Images were captured using light
microscopy (Nikon-70i).
Analysis of sperm motility and
morphology
Cauda epididymides were collected and rinsed with
RNase-free distilled water, followed by the incubation of swim-out
sperm in 1 ml warm human tubal fluid (Wisent Biotechnology) at
37°C. The total sperm count was determined using a hemocytometer
and sperm motility was determined with a computer-assisted sperm
analysis system (CASA; Sperm Class Analyzer v. 4.0.0; Microptic
S.L., Barcelona, Spain). The sperm deformity rate was studied using
sperm smears and microscopy counting methods. Sperm smears were
stained with H&E and images were captured by light microscopy
(Nikon-70i). More than 1,000 sperm were observed and counted from
Tex33+/+ and Tex33−/− males
(n=3 mice/group). The numbers of sperm showing normal and abnormal
morphology were counted and analyzed.
Fertility testing
Tex33+/+ and
Tex33−/− male mice (10–12 weeks old) were
continuously mated with Tex33+/+ female mice
(8–10 weeks old) with demonstrable fertilizing ability (at a ratio
of 1:2) for 1 month. The Tex33+/+ female mice
were changed once a week. Plugs and the numbers of pups born were
recorded.
Statistical analysis
Sperm quality and average litter sizes are expressed
as mean ± standard error of mean and the Student's t-test was used
to identify significant differences. Plugs and pregnancies were
compared between Tex33+/+ and
Tex33−/− mice using the Chi-squared test.
P<0.05 was considered to indicate a statistically significant
result.
Results
Expression of Tex33 during
spermatogenesis in mice
The GenBank database revealed that the mouse
Tex33 gene is located in the E1 region of chromosome 15 and
contains seven exons (Fig. 1A).
There are three transcript variants: V1, V2 and V3 (Fig. 1B). V1 is the longest Tex33
transcript variant; it is 1,147 bp in size and lacks exon 3 as the
result of alternative splicing events (Fig. 1B). V2 is the predominant transcript
variant of the Tex33 gene according to GeneBank, and
exhibits alternative splice sites in exon 1 and exon 3, resulting
in an additional exon and the deletion of exon 3 (Fig. 1B). Another transcript variant, V3,
is the shortest variant and exhibits alternative splice sites in
exons 1 and 2 (Fig. 1B). The
translation of V1 and V2 begins from the same site in exon 2 and
these two variants share one stop codon in exon 7; thus, they both
encode the same isoform (isoform 1; Fig. 1C). However, V3, encoding isoform 2,
translates from exon 4 and lacks a portion of the 5 coding region;
this leads to a shorter N-terminus compared with isoform 1
(Fig. 1C).
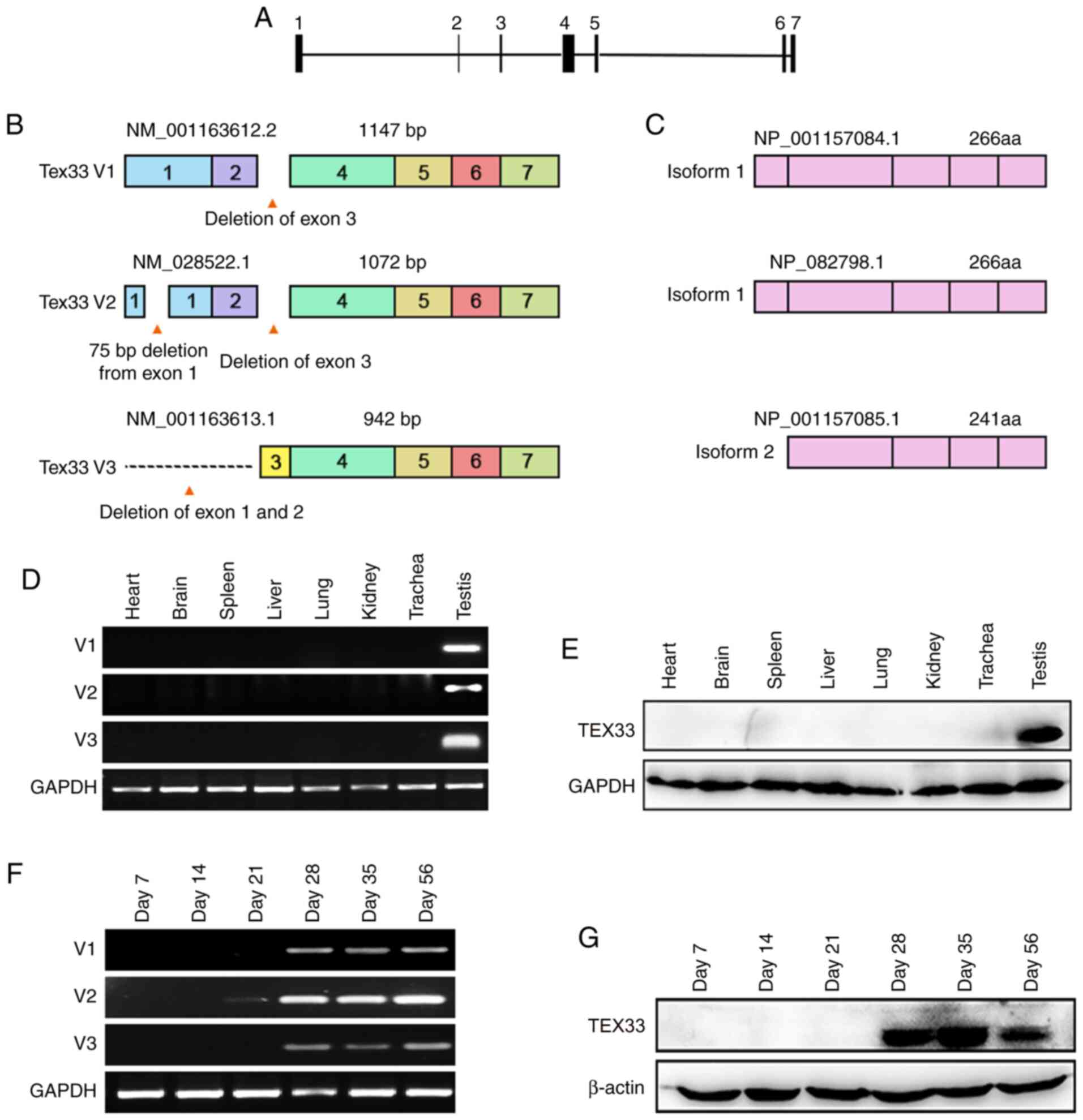 | Figure 1.Characterization of mouse
Tex33. (A) Analysis of the Tex33 gene in mice. The
Tex33 gene consists of seven exons. (B) Tex33
transcript variants in mice. Deleted regions reveal alternate
splicing events and different combinations of exons. Transcript
variant V1 contains six exons, transcript variant V2 has seven
exons and transcript variant V3 has five exons. (C) Two different
isoforms are encoded by transcript variants V1, V2 and V3. (D) mRNA
levels of Tex33 transcript variants in various mouse tissues
were analyzed by RT-PCR. Transcript variant V1, 220 bp; transcript
variant V2, 348 bp; transcript variant V3, 261 bp. (E) Expression
of TEX33 protein in various mouse tissues, as determined by western
blot analysis. (F) Expression of Tex33 transcript variants
during the first wave of spermatogenesis, as determined by RT-PCR.
(G) Expression of TEX33 protein during the first wave of
spermatogenesis, as determined by western blot analysis. TEX33,
testis-expressed protein 33; RT-PCR, reverse
transcription-polymerase chain reaction. |
To examine the mRNA levels of Tex33 in vivo,
specific primers were designed for the three transcript variants
and the expression profiles of the three transcript variants were
then determined by RT-PCR using cDNA samples synthesized from
various adult mouse organs, including the heart, brain, spleen,
liver, lung, kidney, trachea and testis. The results showed that
all transcript variants were only detected in the testes (Fig. 1D). To investigate the levels of
TEX33 protein in these tissues, a TEX33 antibody with high
efficiency was generated as described in our previous study
(27). Western blot analysis
performed using the TEX33 antibody revealed that TEX33 was
expressed exclusively in the testes, which is consistent with the
expression profiles of the transcript variants (Fig. 1E).
To determine the expression profile of Tex33
during the first wave of spermatogenesis, mouse testes were
obtained at postnatal day 7 (containing only spermatogonia), day 14
(containing spermatogonia and spermatocytes), day 21 (in which
round spermatids begin to appear), day 28 (in which spermatids
begin to elongate), day 35 (around which time the first wave of
spermatogenesis is completed) and day 56 (3,28). The
results of RT-PCR analysis suggested that V2 was expressed weakly
from day 21 in the mouse testes, peaked at day 28 and was expressed
into adulthood, while V1 and V3 were expressed from day 28 and
maintained peak expression levels into adulthood (Fig. 1F). Western blotting showed that
TEX33 protein first presented on day 28 and maintained high
expression levels into adulthood (Fig.
1G). The appearance of Tex33 transcript variants and
proteins coincides with the spermiogenesis phase, during which time
the appearance of round spermatids begins and spermiogenesis is
initiated (28).
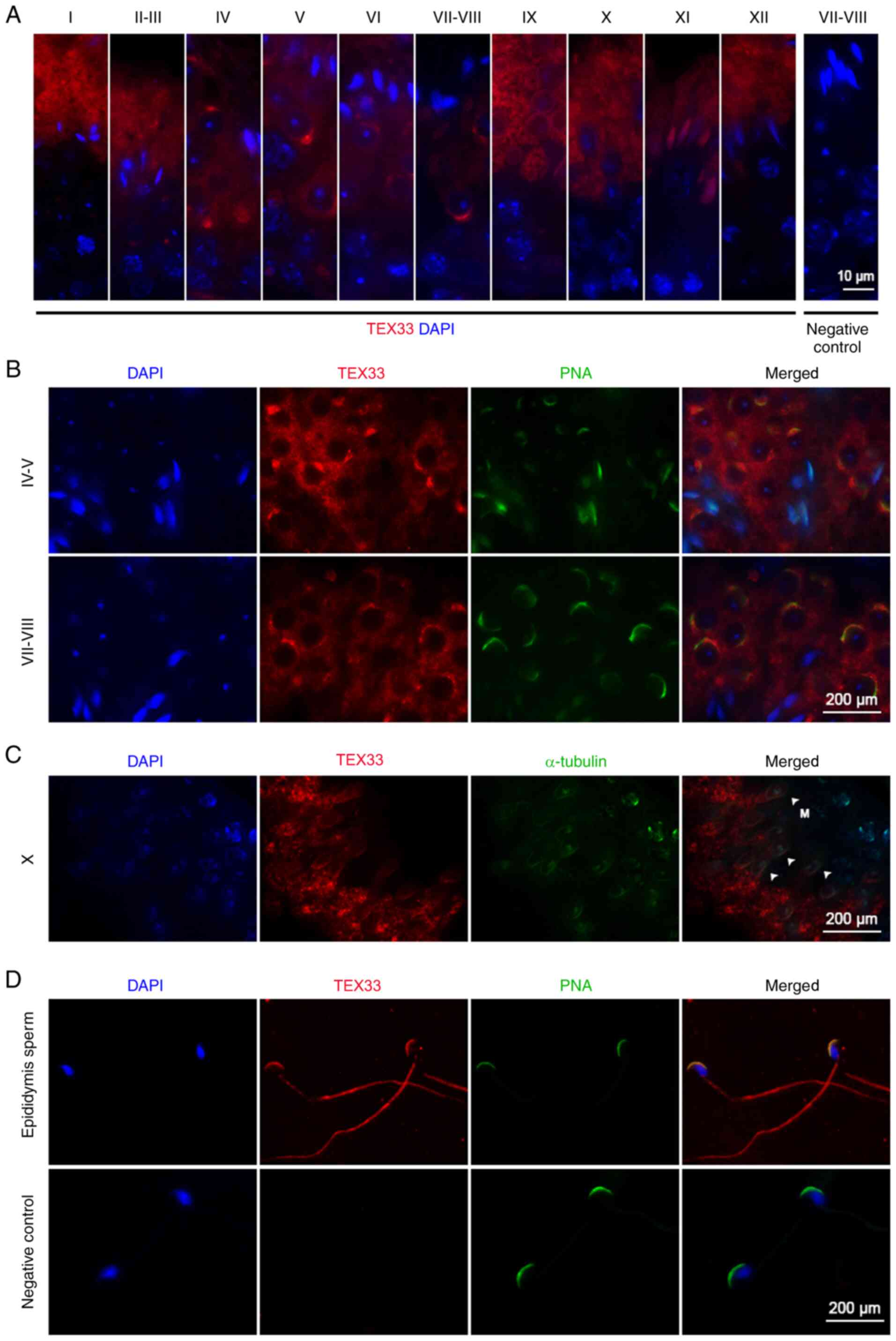
Immunofluorescence analysis demonstrated that TEX33
protein was localized in spermatids and testicular sperm within the
seminiferous tubules (Fig. 2A). In
spermatids, TEX33 was detected in Golgi-derived pro-acrosomal
granules, the cap-like acrosome and the fully formed acrosome from
spermatogenic stages I to XII (Fig.
2A). It has been reported that the acrosome could bind many
lectins in a variety of mammalian species (29,30),
such as peanut agglutinin (PNA), which reacts specifically with
acrosomal sugar components and is widely used to visualize
acrosomal formation (31–35). In the present study, TEX33 was
co-localized with PNA on the acrosomes during different stages of
spermatid development (Fig. 2B) and
on mature acrosomes (Fig. 2D).
TEX33 was also detected on the manchette of elongating spermatids
(spermatogenic stage X) via α-tubulin co-labeling (Fig. 2C), as well as the sperm flagellum
(Fig. 2D). These findings indicate
that TEX33 may be involved in the formation of the acrosome,
manchette and flagellum during spermiogenesis.
Deletion of Tex33 in mice
The initial data indicate that TEX33 is expressed at
high levels in mouse testes. On the basis of this, the biological
function of TEX33 during spermatogenesis was investigated.
CRISPR/Cas9-mediated genome engineering was used to create
Tex33 knockout mice. Exons 2, 3 and 4 were selected as the
target sites. Cas9 and gRNA were co-injected into fertilized eggs
in order to delete exons 2–4 (Fig.
3A). Mice were then genotyped by PCR using specific primers
(Fig. 3A and B). Western blot
analysis was performed on testis samples to validate the efficiency
of Tex33 removal. As shown in Fig. 3C, TEX33 protein was present in the
Tex33+/+ and Tex33+/− mice, but
absent from the Tex33−/− mice. Furthermore,
immunofluorescence analysis of the testis and sperm confirmed that
TEX33 expression was absent from the Tex33−/−
mice (Fig. 3D).
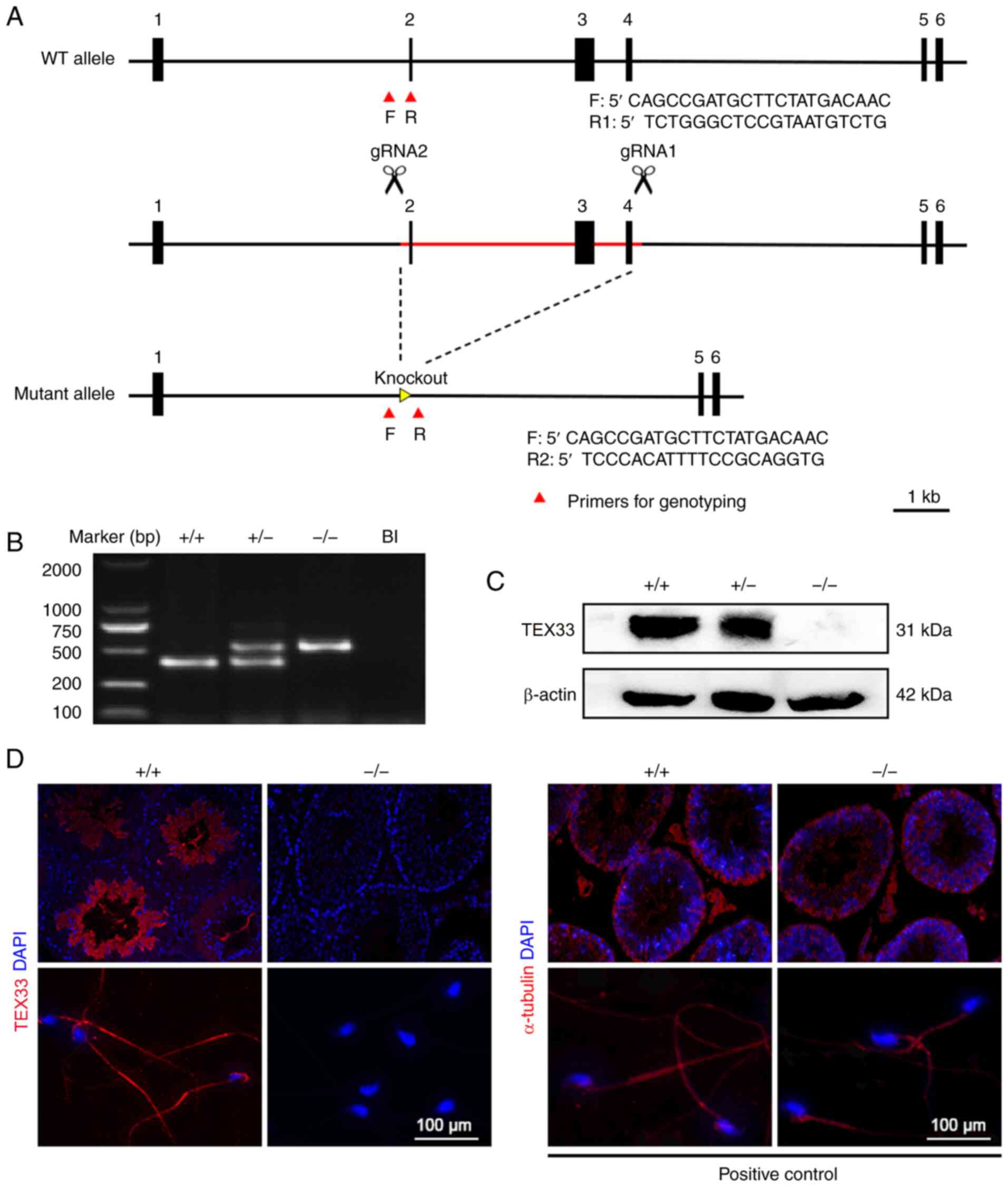 | Figure 3.Generation of Tex33 knockout
mice. (A) Schematic representation of the deletion of Tex33
exons 2–4 via Cas9 microinjection. Primers were designed for
genotyping. F targets intron 1, R1 targets exon 2 and R2 targets
intron 4. (B) Genotyping of Tex33 offspring by PCR assay.
(C) Comparison of TEX33 knockout efficiency between
Tex33+/+, Tex33+/− and
Tex33−/− mice by western blot analysis. (D)
Detection of TEX33 expression in testes (upper panels) and sperm
(lower panels) by immunofluorescence analysis. WT, wild-type; F,
forward; R, reverse; gRNA, guide RNA; TEX33, testis-expressed
protein 33; +/+, Tex33+/+ (wild-type) mice; +/−,
Tex33+/− (heterozygous) mice; −/−,
Tex33−/− (homozygote) mice; Bl, blank; DAPI,
4,6-diamidino-2-phenylindole. |
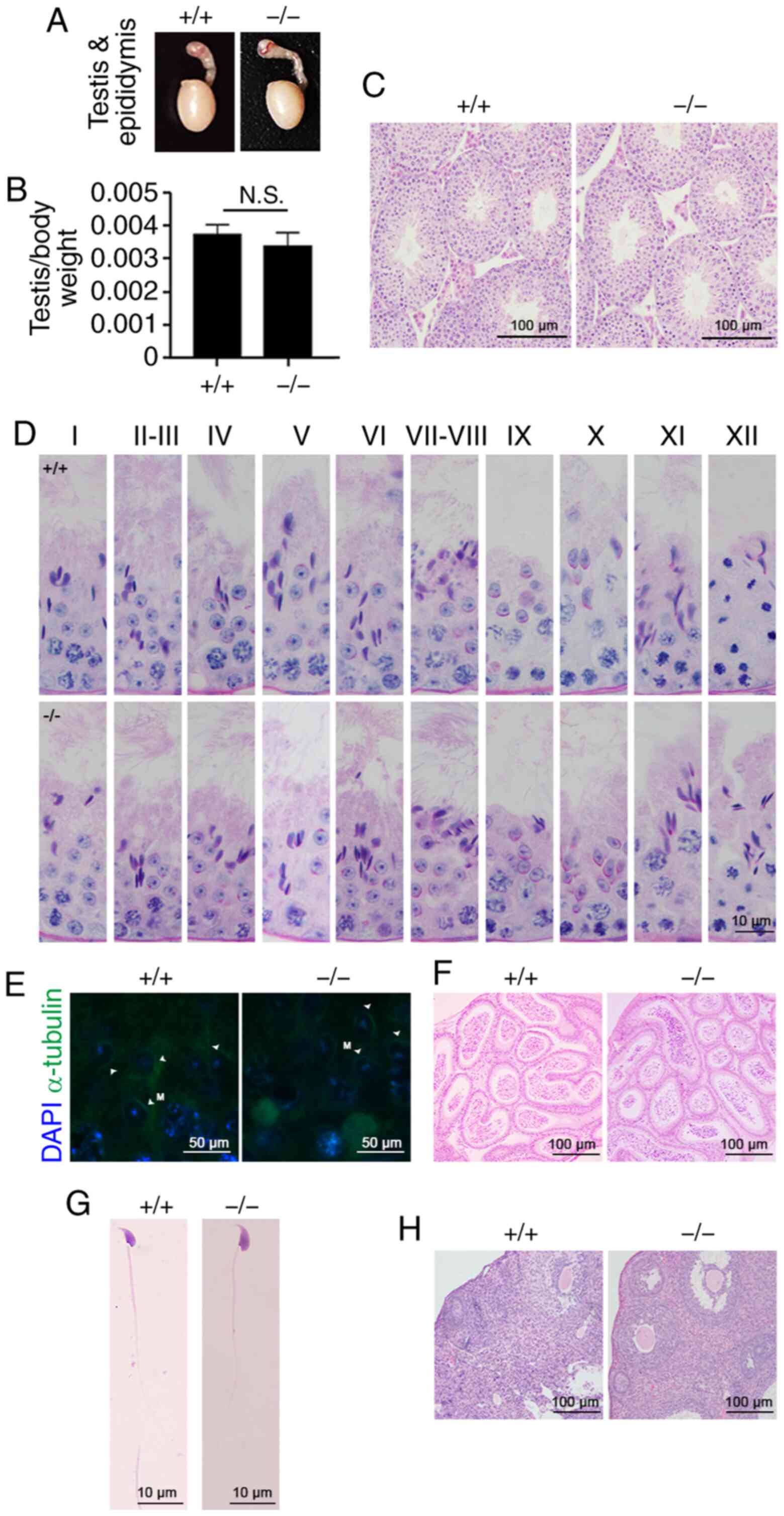
Tex33 deletion does not affect spermatogenesis and
fertility. To determine whether the deletion of Tex33 has an
effect on fertility, functional analyses were then carried out on
Tex33 knockout mice. As shown in Fig. 4A, the sizes of the testes and
epididymis were comparable between adult Tex33+/+
and Tex33−/− males. The testis-to-body weight
ratios were not significantly different between the
Tex33+/+ and Tex33−/− male mice
(Fig. 4B). The process of sperm
development in the testis was examined by H&E staining. The
results revealed that Tex33+/+ and
Tex33−/− mice exhibited seminiferous tubules with
normal morphology and spermatogenic cells (Fig. 4C).
PAS staining was then performed to analyze the
process of acrosome development during spermatogenesis. No apparent
morphological differences were observed between the testes of
Tex33+/+ and Tex33−/− mice in
spermatogenic stages I–XII (Fig.
4D). The morphology of the manchette was evaluated by α-tubulin
immunofluorescence staining and immunostaining was identified in
elongating spermatids in the seminiferous tubules of
Tex33+/+ and Tex33−/− male mice
at spermatogenic stage IX (Fig.
4E). H&E staining revealed the presence of mature sperm in
the cauda epididymides of Tex33+/+ and
Tex33−/− mice (Fig.
4F). In addition, the morphology of sperm exhibited no clear
differences between the Tex33+/+ and
Tex33−/− mice (Fig.
4G and Table I). Sperm were
counted and their motility assessed to reveal the effect of
Tex33 knockout on sperm quality. Sperm counts were
determined using a counting board while motility was determined by
a CASA analysis that provided total motility and progressive
motility data. These investigations revealed no detectable
differences in either sperm count or motility between the
Tex33+/+ and Tex33−/− mice
(Table I).
 | Table I.Analysis of sperm quality in
Tex33+/+ and Tex33−/− male
mice. |
Table I.
Analysis of sperm quality in
Tex33+/+ and Tex33−/− male
mice.
| Genotype | Sperm count
(×106/ml) | Total sperm
motility (%) | Progressive
motility (%) | Sperm deformation
(%) |
|---|
| +/+ (n=3) | 9.02±0.46 | 80.82±3.69 | 57.59±5.06 | 16.31±3.76 |
| −/− (n=3) | 8.98±0.35 | 77.96±4.51 | 58.26±3.47 | 18.12±4.08 |
| P-value | 0.91 | 0.47 | 0.86 | 0.60 |
A breeding assay was performed to evaluate the
fertility of the Tex33 knockout mice.
Tex33+/+ and Tex33−/− male mice
(10–12 weeks old) were mated separately with female mice (8–10
weeks old) for 1 month. The Tex33+/+ and
Tex33−/− male mice exhibited normal fertility
(Tables II–IV), and the litter sizes sired by
Tex33−/− mice were similar to those sired by
Tex33+/+ mice (Table
II). These results indicate that the fertilizing ability of
Tex33−/− male mice was not significantly
different from that of Tex33+/+ mice. No marked
differences in female fertility were observed between the two
groups of mice, as the Tex33−/− female mice
exhibited normal ovarian structures (Fig. 4H), underwent normal pregnancies and
produced equivalent litter sizes to the Tex33+/+
female mice (Tables II and
III).
| Table II.Fertility analysis of
Tex33+/+ and Tex33−/− mice. |
Table II.
Fertility analysis of
Tex33+/+ and Tex33−/− mice.
| Test | Males | Females | Average litter
size | P-value |
|---|
| Male fertility | +/+ (n=8) | +/+ (n=64) | 8.69±0.37 | 0.13 |
|
| −/− (n=8) | +/+ (n=64) | 8.41±0.34 |
|
| Female
fertility | +/+ (n=5) | +/+ (n=12) | 8.67±0.86 | 0.61 |
|
| +/+ (n=5) | −/− (n=12) | 8.92±0.80 |
|
| Table IV.Comparison of plugs in
Tex33+/+ and Tex33−/− mice. |
Table IV.
Comparison of plugs in
Tex33+/+ and Tex33−/− mice.
| Test | Males | Females | Plugs (%) | P-value |
|---|
| Male fertility | +/+ (n=8) | +/+ (n=64) | 93.75 | 0.35 |
|
| −/− (n=8) | +/+ (n=64) | 87.50 |
|
| Female
fertility | +/+ (n=5) | +/+ (n=12) | 91.67 | 0.54 |
|
| +/+ (n=5) | −/− (n=12) | 83.33 |
|
| Table III.Comparison of pregnancies in
Tex33+/+ and Tex33−/− mice. |
Table III.
Comparison of pregnancies in
Tex33+/+ and Tex33−/− mice.
| Test | Males | Females | Pregnancies
(%) | P-value |
|---|
| Male fertility | +/+ (n=8) | +/+ (n=64) | 90.00 | 0.23 |
|
| −/− (n=8) | +/+ (n=64) | 94.64 |
|
| Female
fertility | +/+ (n=5) | +/+ (n=12) | 81.81 | 0.59 |
|
| +/+ (n=5) | −/− (n=12) | 90.00 |
|
Discussion
Mature sperm is the end product of spermiogenesis
and exhibits highly specialized morphology. Spermiogenesis is
facilitated by an important microtubule structure referred to as
the manchette, which appears in elongating spermatids and helps to
create the shape of the sperm head and transport proteins to the
flagellum (2,11). Failure in spermiogenesis, commonly
results in subfertility or infertility due to sperm malformation
and/or dysfunction (36,37). A number of genes have been reported
to be responsible for spermiogenesis (38–42);
however, the expression profiles of other genes and their
respective roles in spermiogenesis remain unknown and require
further investigation. In the present study, Tex33, a gene
previously shown to be associated with cilia formation (26), was demonstrated for the first time,
to the best of our knowledge, to be testis-specific and associated
with spermiogenesis. Its role in spermatogenesis was investigated
further using a Tex33 knockout mouse model.
In mammals, primary RNA transcripts give rise to
different transcript variants through alternative splicing, thereby
resulting in different mRNA and protein isoforms that may have
distinct functions and localization patterns (43). Analysis of GenBank data revealed
that Tex33 pre-mRNA undergoes alternative splicing to
produce three transcripts. Exon 3 is present in V3, but lacking
from V1 and V2; this may be due to exon skipping. Similarly, exon
skipping may have contributed to the deletion of exons 1 and 2 from
V3. In addition, unlike exon 1 of V1, exon 1 of V2 has a 75-bp
deletion. The results of RT-PCR analysis revealed that all three
transcript variants are testis specific. Furthermore, TEX33 protein
was also detected only in the testis, indicating that the
Tex33 gene is testis specific and conserved from
transcription to RNA translation. However, the distribution of this
gene in the testis has not previously been elucidated. It has been
reported that Tex33 is associated with cilia formation;
therefore, we hypothesized that the TEX33 protein may localize on
the sperm flagellum. RT-PCR analysis revealed that the expression
of Tex33 transcript variants began during spermiogenesis,
between days 21 and 28, when round spermatids begin to appear and
undergo morphological changes. Consistent with this finding, the
expression of TEX33 protein was first detected at day 28, and
localized on the acrosome at different developmental stages,
including at maturity. Furthermore, it was also detected on the
manchette of elongating spermatids during spermatogenesis stages IX
and X, as well as on the sperm flagellum. These results confirm
that Tex33 is not only associated with cilia, as previously
reported, but is also testis-specific and associated with
spermiogenesis. Thus, Tex33 may serve a crucial role in
mouse spermiogenesis, particularly in terms of acrosome biogenesis,
manchette morphology and flagellum formation.
Considering the enrichment and cellular localization
of TEX33 in the testis, Tex33 knockout mice were generated
by application of the CRISPR/Cas9 system, as reported by Miyata
et al (44). The association
between TEX33 and spermatid elongation was then explored. Exons 2–4
were selected as targets to ensure the non-functionality of
Tex33 after knockout. The deletion was confirmed by
genotyping, western blotting and immunofluorescence analysis.
Recent studies have identified that numerous
testis-enriched/specific genes are necessary for male fertility,
including CFAP97 domain containing 1, kinesin family member 9,
intraflagellar transporter protein 140, protease serine 55 and
RIKEN cDNA 1700102P08 (39,40,45–47).
Mice lacking any of these genes have been shown to be sterile due
to a failure to complete spermatogenesis. However, the Tex33
knockout mice were found to be fertile. Further analyses of testis
and cauda epididymis histology, acrosome biogenesis, manchette
morphology and sperm quality also indicated that there were no
significant differences between Tex33−/− mice and
the controls. The possible reason for these knockout mice
exhibiting normal spermatogenesis and fertility may be functional
redundancy (48–50). Paralogous genes may compensate for
the loss of function of knockout genes when they contribute to
single function (50). Likewise, it
is plausible that Tex33 paralogs, such as Tex11 and
Tex14, have compensational functions in the absence of
Tex33.
The function of Tex33 during mouse
spermatogenesis has been preliminarily clarified using CRISPR/Cas9
system in the present study. However, considering the cellular and
subcellular localization of Tex33 on sperm and the acrosome,
further investigations using transmission electron microscopy or
in vitro fertilization experiments are recommended.
In summary, the present study preliminarily
describes the expression profile and role of TEX33 during
spermatogenesis in mice. Although Tex33 was testis-specific
and associated with spermiogenesis, deletion of this gene had no
effect on sperm development and reproduction in mice. This
indicates that Tex33 may regulate spermiogenesis in
association with cooperating factors; however, it is not required
for spermiogenesis and male fertility.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81871205), the
Postgraduate Research & Practice Innovation Program of Yangzhou
University (grant no. XKYCX18_129) and the Undergraduate Science
& Technology Innovation Program of Yangzhou University (grant
no. 20180744).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors contributions
MX, JX and CN designed and performed the
experiments. MX wrote the manuscript. YaZ collected and analyzed
the data. MX, TG and YD generated knockout mice and the TEX33
antibody. YiZ designed the study. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Animal
Ethics Committee of Yangzhou University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bao J, Tang C, Li J, Zhang Y, Bhetwal BP,
Zheng H and Yan W: RAN-binding protein 9 is involved in alternative
splicing and is critical for male germ cell development and male
fertility. PLoS Genet. 10:e10048252014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lehti MS and Sironen A: Formation and
function of the manchette and flagellum during spermatogenesis.
Reproduction. 151:R43–R54. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li RK, Tan JL, Chen LT, Feng JS, Liang WX,
Guo XJ, Liu P, Chen Z, Sha JH, Wang YF, et al: Iqcg is essential
for sperm flagellum formation in mice. PLoS One. 9:e980532014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nishimura H and LHernault SW:
Spermatogenesis. Curr Biol. 27:R988–R994. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kazarian E, Son H, Sapao P, Li W, Zhang Z,
Strauss JF III and Teves ME: SPAG17 is required for male germ cell
differentiation and fertility. Int J Mol Sci. 19:12522018.
View Article : Google Scholar
|
|
6
|
Bryant JM, Donahue G, Wang X, Meyer-Ficca
M, Luense LJ, Weller AH, Bartolomei MS, Blobel GA, Meyer RG, Garcia
BA, et al: Characterization of BRD4 during mammalian postmeiotic
sperm development. Mol Cell Biol. 35:1433–1448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kistler WS, Baas D, Lemeille S, Paschaki
M, Seguin-Estevez Q, Barras E, Ma W, Duteyrat JL, Morlé L, Durand
B, et al: RFX2 is a major transcriptional regulator of
spermiogenesis. PLoS Genet. 11:e10053682015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang H, Wan H, Li X, Liu W, Chen Q, Wang
Y, Yang L, Tang H, Zhang X, Duan E, et al: Atg7 is required for
acrosome biogenesis during spermatogenesis in mice. Cell Res.
24:852–869. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu C, Song Z, Wang L, Yu H, Liu W, Shang
Y, Xu Z, Zhao H, Gao F, Wen J, et al: Sirt1 regulates acrosome
biogenesis by modulating autophagic flux during spermiogenesis in
mice. Development. 144:441–451. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Khawar MB, Gao H and Li W: Mechanism of
acrosome biogenesis in mammals. Front Cell Dev Biol. 7:1952019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Okuda H, DeBoer K, OConnor AE, Merriner
DJ, Jamsai D and OBryan MK: LRGUK1 is part of a multiprotein
complex required for manchette function and male fertility. FASEB
J. 31:1141–1152. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Martins LR, Bung RK, Koch S, Richter K,
Schwarzmüller L, Terhardt D, Kurtulmus B, Niehrs C, Rouhi A,
Lohmann I, et al: Stk33 is required for spermatid differentiation
and male fertility in mice. Dev Biol. 433:84–93. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Schultz N, Hamra FK and Garbers DL: A
multitude of genes expressed solely in meiotic or postmeiotic
spermatogenic cells offers a myriad of contraceptive targets. Proc
Natl Acad Sci USA. 100:12201–12206. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nozawa K, Zhang Q, Miyata H, Devlin DJ, Yu
Z, Oura S, Koyano T, Matsuyama M, Ikawa M and Matzuk MM: Knockout
of serine-rich single-pass membrane protein 1 (Ssmem1) causes
globozoospermia and sterility in male mice. Biol Reprod.
103:244–253. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Da Costa R, Bordessoules M, Guilleman M,
Carmignac V, Lhussiez V, Courot H, Bataille A, Chlémaire A, Bruno
C, Fauque P, et al: Vps13b is required for acrosome biogenesis
through functions in Golgi dynamic and membrane trafficking. Cell
Mol Life Sci. 77:511–529. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Paiardi C, Pasini ME, Gioria M and Berruti
G: Failure of acrosome formation and globozoospermia in the wobbler
mouse, a Vps54 spontaneous recessive mutant. Spermatogenesis.
1:52–62. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dunleavy JEM, Okuda H, OConnor AE,
Merriner DJ, ODonnell L, Jamsai D, Bergmann M and OBryan MK:
Katanin-like 2 (KATNAL2) functions in multiple aspects of haploid
male germ cell development in the mouse. PLoS Genet.
13:e10070782017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lehti MS, Zhang FP, Kotaja N and Sironen
A: SPEF2 functions in microtubule-mediated transport in elongating
spermatids to ensure proper male germ cell differentiation.
Development. 144:2683–2693. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Schwarz T, Prieler B, Schmid JA, Grzmil P
and Neesen J: Ccdc181 is a microtubule-binding protein that
interacts with Hook1 in haploid male germ cells and localizes to
the sperm tail and motile cilia. Eur J Cell Biol. 96:276–288. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xu Z, Miyata H, Kaneda Y, Castaneda JM, Lu
Y, Morohoshi A, Yu Z, Matzuk MM and Ikawa M: CIB4 is essential for
the haploid phase of spermatogenesis in mice. Biol Reprod.
103:235–243. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang Z, Kostetskii I, Tang W,
Haig-Ladewig L, Sapiro R, Wei Z, Patel AM, Bennett J, Gerton GL,
Moss SB, et al: Deficiency of SPAG16L causes male infertility
associated with impaired sperm motility. Biol Reprod. 74:751–759.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Ou Y, Cheng M, Saadi HS,
Thundathil JC and van der Hoorn FA: KLC3 is involved in sperm tail
midpiece formation and sperm function. Dev Biol. 366:101–110. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kuo YC, Shen YR, Chen HI, Lin YH, Wang YY,
Chen YR, Wang CY and Kuo PL: SEPT12 orchestrates the formation of
mammalian sperm annulus by organizing core octameric complexes with
other SEPT proteins. J Cell Sci. 128:923–934. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moretti E, Collodel G, Mazzi L, Russo I
and Giurisato E: Ultrastructural study of spermatogenesis in KSR2
deficient mice. Transgenic Res. 24:741–751. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang X, Wei Y, Fu G, Li H, Saiyin H, Lin
G, Wang Z, Chen S and Yu L: Tssk4 is essential for maintaining the
structural integrity of sperm flagellum. Mol Hum Reprod.
21:136–145. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
McClintock TS, Glasser CE, Bose SC and
Bergman DA: Tissue expression patterns identify mouse cilia genes.
Physiol Genomics. 32:198–206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xia M, Xia J, Yang D, Liu M, Niu C, Shen
X, Sun H and Zheng Y: Preparation and application of rabbit
polyclonal antibody against mouse Tex33. Xi Bao Yu Fen Zi Mian Yi
Xue Za Zhi. 34:643–649. 2018.(In Chinese). PubMed/NCBI
|
|
28
|
Busada JT, Velte EK, Serra N, Cook K,
Niedenberger BA, Willis WD, Goulding EH, Eddy EM and Geyer CB:
Rhox13 is required for a quantitatively normal first wave of
spermatogenesis in mice. Reproduction. 152:379–388. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee MC and Damjanov I: Lectin binding
sites on human sperm and spermatogenic cells. Anat Rec.
212:282–287. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Arya M and Vanha-Perttula T: Comparison of
lectin-staining pattern in testis and epididymis of gerbil, guinea
pig, mouse, and nutria. Am J Anat. 175:449–469. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wakayama T, Nakata H, Kumchantuek T,
Gewaily MS and Iseki S: Identification of
5-bromo-2-deoxyuridine-labeled cells during mouse spermatogenesis
by heat-induced antigen retrieval in lectin staining and
immunohistochemistry. J Histochem Cytochem. 63:190–205. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fàbrega A, Puigmulé M, Dacheux JL, Bonet S
and Pinart E: Glycocalyx characterisation and glycoprotein
expression of Sus domesticus epididymal sperm surface samples.
Reprod Fertil Dev. 24:619–630. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu Y, Zhong A, Zheng H, Jiang M, Xia Z, Yu
J, Chen L and Huang X: Expression of flotilin-2 and acrosome
biogenesis are regulated by MiR-124 during spermatogenesis. PLoS
One. 10:e01366712015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhong L, Yang M, Zou X, Du T, Xu H and Sun
J: Human umbilical cord multipotent mesenchymal stromal cells
alleviate acute ischemia-reperfusion injury of spermatogenic cells
via reducing inflammatory response and oxidative stress. Stem Cell
Res Ther. 11:2942020. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang Q, Liu H, Zeng J, Li W, Zhang S,
Zhang L, Song S, Zhou T, Sutovsky M, Sutovsky P, et al: COP9
signalosome complex subunit 5, an IFT20 binding partner, is
essential to maintain male germ cell survival and acrosome
biogenesis. Biol Reprod. 102:233–247. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Anawalt BD: Approach to male infertility
and induction of spermatogenesis. J Clin Endocrinol Metab.
98:3532–3542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
García-Vázquez FA, Gadea J, Matás C and
Holt WV: Importance of sperm morphology during sperm transport and
fertilization in mammals. Asian J Androl. 18:844–850.
2016.PubMed/NCBI
|
|
38
|
Morohoshi A, Miyata H, Shimada K, Nozawa
K, Matsumura T, Yanase R, Shiba K, Inaba K and Ikawa M:
Nexin-Dynein regulatory complex component DRC7 but not FBXL13 is
required for sperm flagellum formation and male fertility in mice.
PLoS Genet. 16:e10085852020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Oura S, Kazi S, Savolainen A, Nozawa K,
Castañeda J, Yu Z, Miyata H, Matzuk RM, Hansen JN, Wachten D, et
al: Cfap97d1 is important for flagellar axoneme maintenance and
male mouse fertility. PLoS Genet. 16:e10089542020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Miyata H, Shimada K, Morohoshi A, Oura S,
Matsumura T, Xu Z, Oyama Y and Ikawa M: Testis-enriched kinesin
KIF9 is important for progressive motility in mouse spermatozoa.
FASEB J. 34:5389–5400. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang L, Zhen J, Huang Q, Liu H, Li W,
Zhang S, Min J, Li Y, Shi L, Woods J, et al: Mouse
spermatogenesis-associated protein 1 (SPATA1), an IFT20 binding
partner, is an acrosomal protein. Dev Dyn. 249:543–555. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lehti MS and Sironen A: Formation and
function of sperm tail structures in association with sperm
motility defects. Biol Reprod. 97:522–536. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang ET, Sandberg R, Luo S, Khrebtukova I,
Zhang L, Mayr C, Kingsmore SF, Schroth GP and Burge CB: Alternative
isoform regulation in human tissue transcriptomes. Nature.
456:470–476. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Miyata H, Castaneda JM, Fujihara Y, Yu Z,
Archambeault DR, Isotani A, Kiyozumi D, Kriseman ML, Mashiko D,
Matsumura T, et al: Genome engineering uncovers 54 evolutionarily
conserved and testis-enriched genes that are not required for male
fertility in mice. Proc Natl Acad Sci USA. 113:7704–7710. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang Y, Liu H, Li W, Zhang Z, Zhang S,
Teves ME, Stevens C, Foster JA, Campbell GE, Windle JJ, et al:
Intraflagellar transporter protein 140 (IFT140), a component of
IFT-A complex, is essential for male fertility and spermiogenesis
in mice. Cytoskeleton (Hoboken). 75:70–84. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kobayashi K, Endo T, Matsumura T, Lu Y, Yu
Z, Matzuk MM and Ikawa M: Prss55 but not Prss51 is required for
male fertility in mice. Biol Reprod. 103:223–234. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wu XL, Yun DM, Gao S, Liang AJ, Duan ZZ,
Wang HS, Wang GS and Sun F: The testis-specific gene 1700102P08Rik
is essential for male fertility. Mol Reprod Dev. 87:231–240. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Krakauer DC and Plotkin JB: Redundancy,
antiredundancy, and the robustness of genomes. Proc Natl Acad Sci
USA. 99:1405–1409. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Khan M, Jabeen N, Khan T, Hussain HMJ, Ali
A, Khan R, Jiang L, Li T, Tao Q, Zhang X, et al: The evolutionarily
conserved genes: Tex37, Ccdc73, Prss55 and Nxt2 are dispensable for
fertility in mice. Sci Rep. 8:49752018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Holcomb RJ, Oura S, Nozawa K, Kent K, Yu
Z, Robertson MJ, Coarfa C, Matzuk MM, Ikawa M and Garcia TX: The
testis-specific serine proteases PRSS44, PRSS46, and PRSS54 are
dispensable for male mouse fertility. Biol Reprod. 102:84–91.
2020.PubMed/NCBI
|