Introduction
To improve the quality of life in patients with
dentition defects and delay the absorption of the alveolar bone,
osteointegrated dental implants have been developed and have been
widely used since the 1980s (1,2). With
the popularization of the dental implant, inflammation in the
mucosa and bone tissues around the implant has become a new problem
(3). The formation of the bacterial
biofilm, on the surface of the implant and in the peripheral
gingival crevicular fluid, stimulates the inflammatory reaction and
finally leads to alveolar bone absorption. In serious cases, the
implant has been found to loosen and fall off (3,4). A
meta-analysis based on 15 articles of 11 studies during 2006 to
2014, found that the incidence of peri-implantitis was ~22%
(5,6). At present, the primary clinical
treatment aim of peri-implantitis is to control the pathogens and
reduce the periodontal pockets around the implant, using
subgingival curettage, antibiotics and laser treatment (7,8).
However, it is difficult to reverse the process of bone absorption.
Therefore, it is important to identify a drug or material that can
inhibit the pathogenic bacteria-induced inflammation and improve
the bone regeneration capability.
Several of the major pathogens of peri-implantitis
belong to Gram-negative bacterium, which produce lipopolysaccharide
(LPS) in their cell wall (9). LPS
predominantly activates the Toll-like receptor 4 (TLR4)/NF-κB
signaling pathway and induces pro-inflammatory cytokine expression,
including TNF-α, IL-6, IL-1β and monocyte chemoattractant protein-1
(MCP-1) (10). Enhancement of
LPS-induced receptor activator of NF-κB ligand (RANKL) and
autophage activation was found to promote osteoclastogenesis
(11,12). Furthermore, LPS could adhere to the
surface of the titanium implant, which had no effect on cell
attachment but impaired osteoblast differentiation (13). Therefore, LPS is a contributing
factor in both soft-tissue destruction and alveolar bone
absorption, affecting the implant success rate (14).
As a key factor during the development of
peri-implantitis, the balance of alveolar bone absorption and
reconstruction was found to be damaged (15). Alveolar bone regeneration includes
two aspects: The osteoclastogenesis of monocyte macrophages and the
osteogenesis of mesenchymal stem cells (15). Bone marrow-derived mesenchymal stem
cells (BMSCs) are a type of multipotent cell, which are considered
to be common progenitors for osteoblasts and adipocytes (16). BMSCs have been applied for bone
repair, due to their osteogenic ability (17). Thus, in the current study, BMSCs
were used to investigate the regulation of osteogenesis. As
aforementioned, LPS was found to impair osteoblast differentiation;
in addition, a previous study has reported that LPS could affect
the potential differentiation tendency of BMSCs (18). Therefore, it is important to
investigate the regulation of drugs on LPS-induced
osteogenesis/adipogenesis imbalance and inflammation in BMSCs.
Berberine is a type of quaternary ammonium alkaloid,
extracted from varieties of plants species, such as Coptis
chinensis (19). Berberine
hydrochloride (BBR) is typically used in a clinical setting, due to
its numerous pharmacological activities, including anti-microbial,
glucose/cholesterol regulatory and immune modulatory properties
(19). Previous studies have
revealed that it is implicated in bone biology. For instance, BBR
inhibited adipogenesis and promoted osteogenesis in the C3H10T1/2
cell line (20). In addition,
berberine suppressed RANKL-induced osteoclast formation and
protected cell survival (21,22).
However, the role of BBR on osteogenesis and adipogenesis under
inflammatory conditions remains unknown. The aim of the present
study was to demonstrate the modulating effect of BBR on the
osteogenesis/adipogenesis balance in LPS-mediated inflammatory
response in BMSCs, which may provide novel evidence for
peri-implantitis medication.
Materials and methods
Materials
BBR, LPS (extracted from Escherichia coli
O111:B4 strain), L-ascorbic acid, β-glycerophosphate disodium,
dexamethasone (Dex), 3-isobutyl-1-methylxanthine (IBMX) and
indomethacin were purchased from Sigma-Aldrich (Merck KGaA).
Compound C was obtained from CSNpharm, Inc. The Cell Counting Kit
(CCK)-8 was purchased from Dojindo Molecular Technologies, Inc. The
alkaline phosphatase (ALP) content assay kit, the ALP stain kit,
BeyoECL Moon (ECL chemiluminescence kit), protease inhibitor
cocktail, phosphatase inhibitor cocktail and insulin were purchased
from Beyotime Institute of Biotechnology. The oil Red O Stain kit
was purchased from Cyagen Biosciences, Inc., and the PrimeScript™
RT kits was obtained from Takara Bio, Inc. The Kapa
SYBR® Fast QPCR Master Mix (2X) Universal kit was
purchased from Roche Diagnostics and the primers were synthesized
by Sangon Biotech Co., Ltd. The RIPA lysis buffer and
TRIzol® reagent were purchased from Thermo Fisher
Scientific, Inc., while the antibodies against phosphorylated
(p)-p65 (cat. no. 3033), total p65 (cat. no. 8242), (p)-IκBα (cat.
no. 2859), total IκBα (cat. no. 9243) and anti-rabbit IgG
HRP-linked (cat. no. 7074) were purchased from Cell Signaling
Technology, Inc. The anti-GAPDH antibody (cat. no. 10494-1-AP) was
purchased from ProteinTech Group, Inc. Basic α-modified Eagle
medium (α-MEM), FBS and penicillin-streptomycin antibiotics were
purchased from Thermo Fisher Scientific, Inc. Other reagents were
of analytical grade and used without further purification.
Primary cell isolation and
culture
Female Sprague-Dawley rats (age, 3 weeks; weight,
80-100 g; 20-26°; relative humidity 40-70%; 12-h light/dark cycle;
diet and drinking water ad libitum) were sacrificed using carbon
dioxide (2 l/min in a 10-l box). The femur and tibia of the rats
were isolated and the bone marrow cavity was washed with α-MEM to
obtain the primary BMSCs. The cells were collected and centrifuged
at 1,000 × g for 10 min at room temperature. After removing the
supernatant, the aspirates were re-suspended with BMSC growth
medium (α-MEM containing 10% FBS, 100 U/ml penicillin and 100 mg/ml
streptomycin) and seeded in a 10-cm dish for 3 days at 37°C, and
then the non-adherent cells were removed. Adherent BMSCs were
further cultured and expanded. Cells that passaged 3-6 times were
used for further experimentation.
Cell viability assay
The BMSCs cells were seeded into 96-well plates
(5,000 cells per well) and cultured overnight, then treated with
BBR at different concentrations (0, 1, 5, 10, 50 and 100 µM) at
37°C for 72 h. Cell viability was measured using the CCK-8 kit
according to the manufacturer's protocol. After removing the
supernatant, fresh growth medium containing CCK-8 (Vcck-8:Vmedium,
1:10) was added into each well and incubated at 37°C for 2 h. The
absorbance of the samples was measured at 450 nm using a Synergy 2
microplate reader (Agilent Technologies, Inc.).
RNA isolation and reverse
transcription-quantitative (RT-q) PCR analysis
Total RNA was extracted using TRIzol®,
and subsequently isolated with isopropanol and chloroform according
to the manufacturer's protocol. RT reactions and qPCR detections
were performed according to the manufacturer's instructions. The
reverse transcription reaction was: 37°C for 15 min, 85°C for 15
sec. The program of the qPCR thermocycling was: First step: 95°C
for 3 min; second: 95°C for 3 sec and 60°C for 30 sec, repeated for
40 cycles. The reactions were performed using Eppendorf
Mastercycler Pro and an ABI 7900 Real-time PCR system (Thermo
Fisher Scientific, Inc.). The relative mRNA expression levels were
normalized to GAPDH using the comparative 2−∆∆Cq method
(23). The PCR primer pairs are
shown in Table I.
 | Table I.Primer sequences for reverse
transcription-quantitative PCR. |
Table I.
Primer sequences for reverse
transcription-quantitative PCR.
| Gene | Forward primer
sequence (5′-3′) | Reverse primer
sequence (5′-3′) |
|---|
| ALP |
AACGTGGCCAAGAACATCATCA |
TGTCCATCTCCAGCCGTGTC |
| RUNX2 |
GCACCCAGCCCATAATAGA |
TTGGAGCAAGGAGAACCC |
| OCN |
TGAGGACCCTCTCTCTGCTC |
GGGCTCCAAGTCCATTGTT |
| SPP1 |
ATCTGAGTCCTTCACTG |
GGGATACTGTTCATCAGAAA |
| FABP4 |
GCGTAGAAGGGGACTTGGTC |
TTCCTGTCATCTGGGGTGATT |
| ADIPSIN |
CACGTGTGCGGTGGCACCCTG |
CCCCTGCAAGTGTCCCTGCGGT |
| PPARγ |
CCTTTACCACGGTTGATTTCTC |
GGCTCTACTTTGATCGCACTTT |
| MCP-1 |
TGCTGTCTCAGCCAGATGCAGTTA |
AGAAGTGCTTGAGGTGGTTGTGGA |
| TNF-α |
CCCAATCTGTGTCCTTCTAACT |
CAGCGTCTCGTGTGTTTCT |
| Il-6 |
GGTTTGCCGAGTAGACCTCA |
GTGGCTAAGGACCAAGACCA |
| Il-1β |
AAAGAAGGTGCTTGGGTCCT |
CAGGAAGGCAGTGTCACTCA |
| GAPDH |
CAGGGCTGCCTTCTCTTGT |
TCCCGTTGATGACCAGCTTC |
Western blot analysis
BMSCs were lysed with RIPA lysis buffer containing
protease inhibitor cocktail and phosphatase inhibitor cocktail.
Cell lysate was collected, and its total protein concentration was
determined using a BCA kit (Thermo Fisher Scientific, Inc.). Total
protein (~20 µg) was loaded in each lane and separated using
SDS-PAGE (12.5%) and then transferred onto PVDF membranes. The
membranes were blocked by 5% non-fat milk in PBST buffer (PBS
containing 0.1% Tween-20) for 1 h at room temperature. Then the
membranes were incubated with the corresponding primary antibodies
(the dilution of p-p65, t-p65, p-IκBα and t-IκBα antibodies,
1:1,000; GAPDH antibody, 1:5,000) at 4°C overnight. The membranes
were washed 3 times with PBST buffer, 10 min each time, and then
incubated with HRP-linked anti-Rabbit IgG (1:2,000) for 2 h at room
temperature. The membranes were then washed 3 times, as previously.
An ECL chemiluminescence kit (cat. no. P0018FS; Beyotime Institute
of Biotechnology) was used for HRP detection. GE Amersham Imager
600 (Cytiva) was used for signal collection. Semi-quantification of
band intensity was performed using ImageJ v1.44p software (National
Institute of Health). Data are presented as the mean ± SD from
three independent experiments.
Osteogenic and adipogenic
differentiation
BMSCs were seeded into 12-well plates at a density
of 2×104 cells/cm2 and cultured until
confluent. To identify the effects of drug administration, LPS (1
µg/ml), BBR (1, 5 and 10 µM) and compound C (1 µM) were used,
unless otherwise specified. For osteogenic differentiation, 10 nM
Dex, 100 mg/l L-ascorbic acid and 10 mM β-glycerophosphate disodium
were added to the BMSC growth media, co-incubated with given drug
simultaneously. The osteoblastic media with drugs was replaced
every 3 days. On day 7, the cells were harvested for ALP staining,
intracellular ALP content assay and gene expression level analysis.
For adipogenic differentiation, 1 µM Dex, 0.5 mM IBMX, 10 µg/ml
insulin and 100 µM indomethacin were added to the BMSC growth
media. The cells were incubated with the adipogenic media and
corresponding drugs at 37°C for 12 days, and the media was replaced
every 3 days. Then, the cells were harvested for oil red O
staining.
ALP stain and intracellular content
assay
Following treatment with osteogenic media and
corresponding drugs for 7 days, the BMSCs were washed with PBS and
fixed with 4% paraformaldehyde for 30 min at room temperature.
Following which, the cells were washed three times with PBS to
remove the fixing liquid. The ALP staining solution was mixed
according to the manufacturer's instructions, and added to the
wells of the culture plate. After 30-min incubation at room
temperature, the cells were washed with sterile water three times
and images were obtained using a light microscope at ×200
magnification (Leica Microsystems GmbH).
For the ALP content assay, cells were treated as
aforementioned, then washed with PBS and collected using a lysis
buffer. The concentration of the samples was detected using a BCA
kit (Thermo Fisher Scientific, Inc.). Each sample was diluted to 2
mg/ml, and the ALP content was measured according to the
manufacturer's instructions. The absorbance was measured at 405
nm.
Oil red O stain
The oil red O working solution was prepared by
diluting the stock solution with sterile water (Vstock:Vwater,
3:2). After differentiation for 12 days, the BMSCs were washed with
PBS, fixed with 4% formaldehyde at room temperature for 15 min and
then stained with freshly diluted oil red O working solution for 60
min at room temperature. Following which, the supernatant was
removed and the cells were washed with water three times. Images of
the oil red O-positive cells were collected using a bright-field
microscope at ×200 magnification.
Statistical analysis
Data are presented as the mean ± SD from three
separate experiments in each group. Statistical differences between
groups were analyzed using one-way ANOVA, multiple comparisons
between the groups were performed using Tukey's test with GraphPad
Prism version 6 (GraphPad Software, Inc.). P<0.05 was considered
to indicate a statistically significant difference.
Results
BBR reverses the LPS-induced decrease
in osteogenic gene expression and increase in adipogenic gene
expression levels
Firstly, to investigate the potential cytotoxicity
of BBR, BMSCs were treated with different concentrations of BBR for
3 days and the cell viability was measured. As shown in Fig. 1A, BBR had no significant cytotoxic
effect on BMSCs; therefore low concentrations, at 1-10 µM were
selected for subsequent experiments. Additionally, the cytotoxic
effects of LPS at different concentrations were also detected. At
the dose of 1 µg/ml, LPS had no significant cytotoxic effect;
however, 10 µg/ml LPS exhibited weak cytotoxicity (data not
shown).
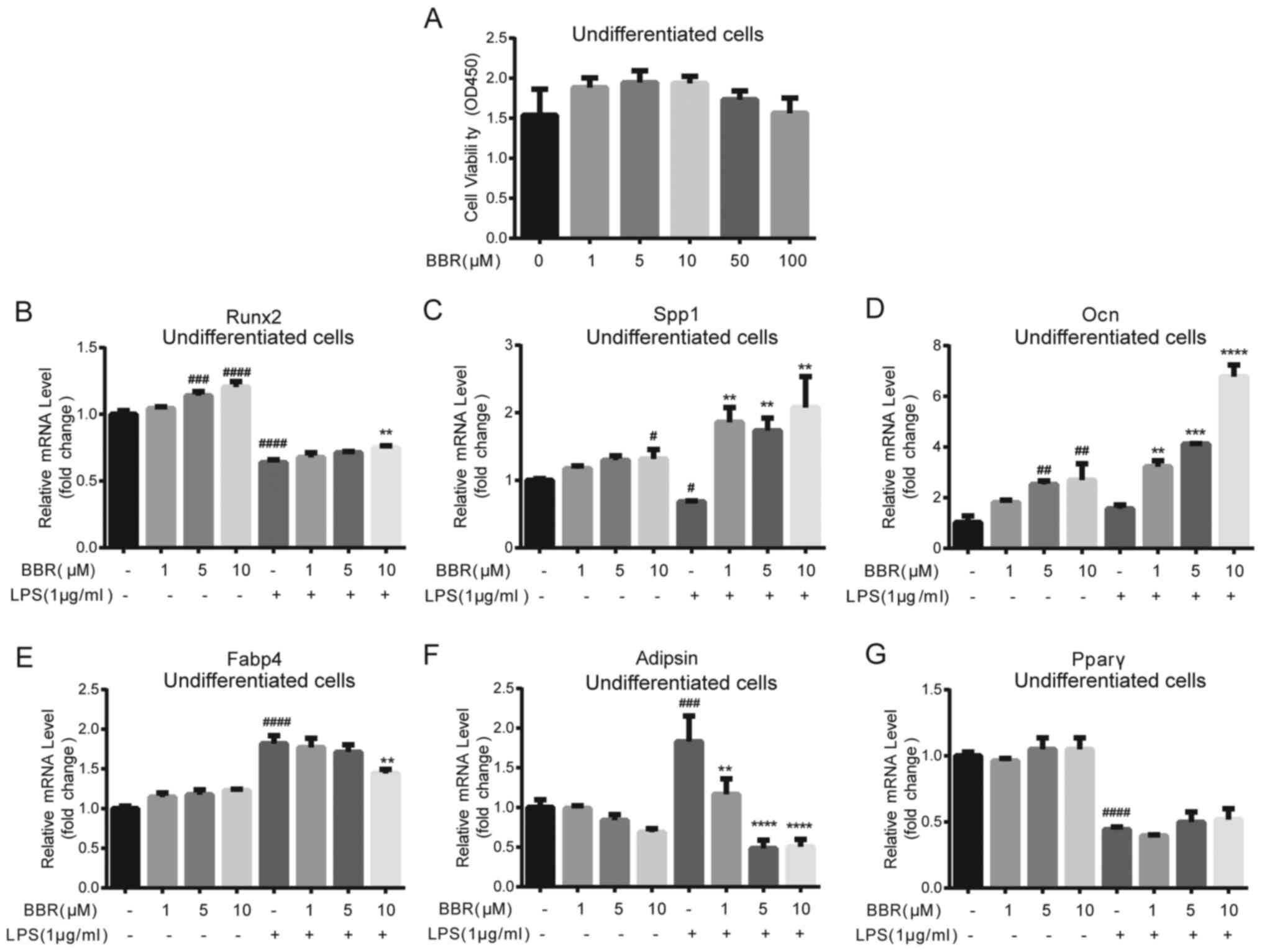 | Figure 1.BBR alters osteogenic and adipogenic
gene expression levels in undifferentiated bone marrow-derived
mesenchymal stem cells. (A) Cells were incubated with DMSO or BBR
(1, 5, 10, 50 or 100 µM) for 3 days, and then cell viability was
determined using a Cell Counting Kit-8 assay. The cells were
treated with BBR (1, 5 or 10 µM) alone or simultaneously with LPS
(1 µg/ml), and then collected for reverse
transcription-quantitative PCR analysis. The mRNA expression levels
of the osteogenic genes (B) Runx2, (C) Spp1 and (D) Ocn, and the
adipogenic genes (E) Fabp4, (F) Adipsin and (G) Pparγ were
measured. Their relative changes were normalized to the GAPDH
expression levels. The data are presented as the mean ± standard
deviation of three independent experiments. #P<0.05,
##P<0.01, ###P<0.001,
####P<0.0001 vs. control group; **P<0.01,
***P<0.001, ****P<0.0001 vs. LPS group. BBR, berberine
hydrochloride; LPS, lipopolysaccharide; Runx2, RUNX family
transcription factor 2; Ocn, osteocalcin; Spp1, secreted
phosphoprotein 1; Fabp4, fatty acid binding protein 4; Pparγ,
peroxisome proliferator-activated receptorγ. |
As LPS-related adverse effects on bone
reconstruction are a key factor in the development of
peri-implantitis (14), the
potential role of BBR on osteogenic and adipogenic gene expression
in undifferentiated BMSCs was investigated. The cells were
co-incubated with BBR at different concentrations (0, 1, 5 and 10
µM) and with or without 1 µg/ml LPS for 3 days. The results
demonstrated that BBR significantly promoted the mRNA expression
levels of osteogenic genes, RUNX family transcription factor 2
(Runx2), osteocalcin (Ocn) and secreted phosphoprotein 1 (Spp1), in
a dose-dependent manner (Fig.
1B-D). Notably, LPS significantly decreased the Runx2 and SPP1
mRNA expression levels, and these effects were partly or totally
reversed by BBR. The Ocn mRNA expression level was not changed by
LPS; however, it was increased after BBR treatment under the
LPS-stimulated condition. For the adipogenic genes, compared with
the negative control, LPS induced a two-fold increase in both fatty
acid binding protein 4 (Fabp4) and Adipsin gene expression levels
(Fig. 1E and F); however, the
peroxisome proliferator-activated receptorγ (Pparγ) expression
level decreased by half (Fig. 1G).
BBR itself had little effect on the adipogenic genes, except for
Adipsin, and a small reduction in the increase of Fabp4 expression
levels induced by LPS. Moreover, 10 µM BBR lowered Adipsin gene
expression level, and further dose-dependently reduced the increase
stimulated by LPS.
BBR promotes BMSC osteogenic
differentiation under LPS treatment
As aforementioned, it was determined that BBR could
ameliorate the LPS-induced inhibition of BMSCs osteogenic gene
expression. Considering that ALP is a key protein during the
differentiated process, particularly in the early and middle stages
(24), ALP protein expression was
determined using ALP staining. As shown in Fig. 2A (upper panel), cells treated with
BBR during the differentiation process exhibited increased staining
compared with that in the control cells, in a dose-dependent
manner. Furthermore, staining of cells treated with LPS was
markedly decreased compared with that in the control cells.
Notably, BBR reversed the LPS-induced reduction (Fig. 2A; lower panel). In addition,
intracellular Alp mRNA expression level exhibited a similar result
(Fig. 2B). Compared with Alp mRNA
expression in the control cells, there was a 13.7-fold increase
induced by BBR treatment and an 8.1-fold change induced by BBR and
LPS co-incubation. The gene expression level of Runx2, the key
transcriptional factor during osteogenesis (25), was higher in cells treated with BBR
and lower in LPS-treated cells. Furthermore, co-incubation with BBR
and LPS reversed the LPS-mediated reduction of Runx2 mRNA
expression level (Fig. 2C). Similar
results were also found for Ocn mRNA expression levels (Fig. 2D), which is one of the essential
osteogenic marker genes (26). The
regulation of BBR on SPP1 mRNA expression was similar with that on
Ocn mRNA expression; however, the result was not significantly
different (Fig. 2E). These findings
indicated that BBR could accelerate osteogenic differentiation and
assist with restricting LPS-induced adverse effects in BMSCs.
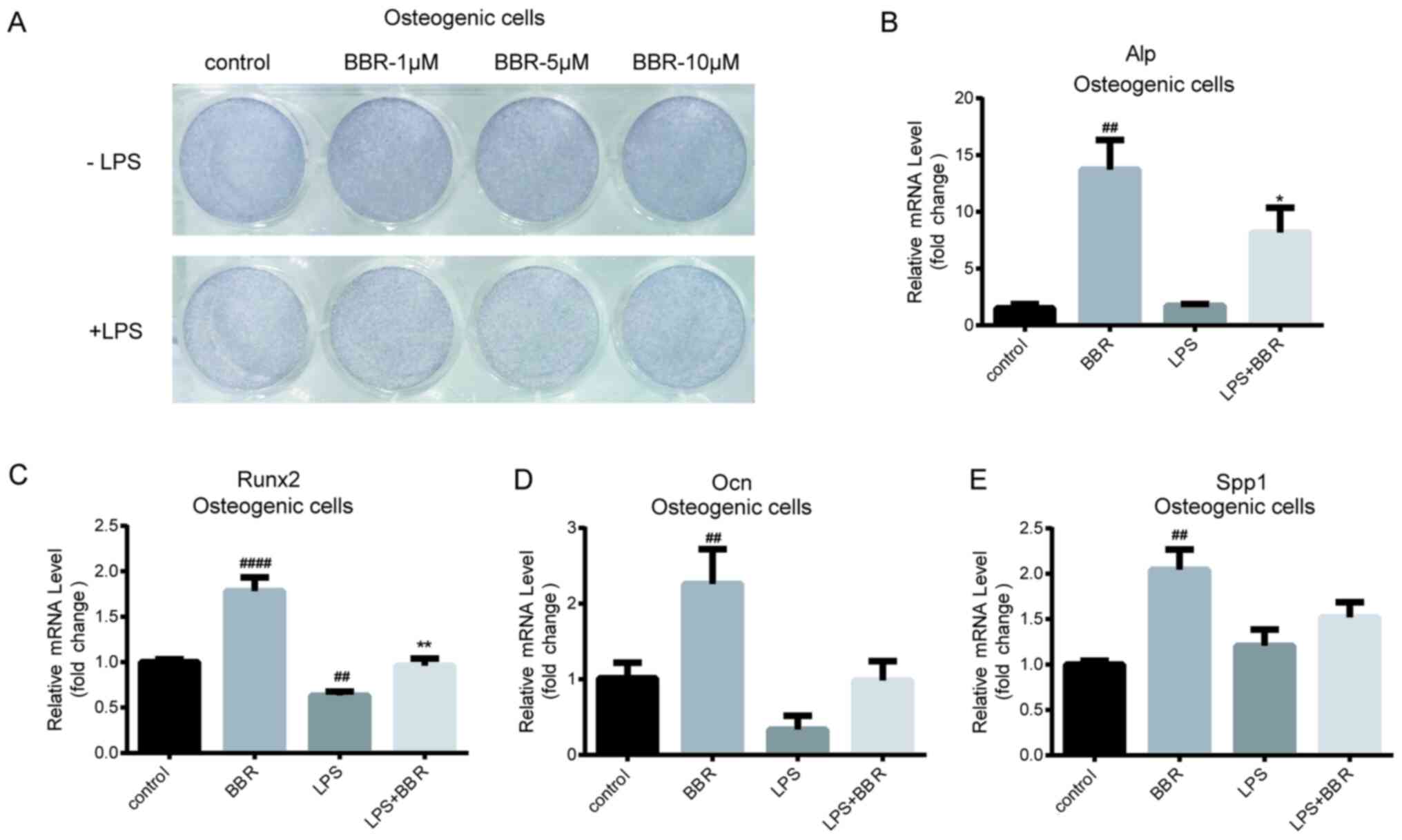 | Figure 2.BBR promotes osteogenesis in bone
marrow-derived mesenchymal stem cells. The cells were incubated
with BBR (0, 1, 5 or 10 µM) alone or simultaneously with LPS (1
µg/ml) in osteogenic media for 7 days. Then, the cells were
detected using ALP stain and reverse transcription-quantitative PCR
analysis. (A) Images of the ALP stained cells were obtained using a
camera. The mRNA expression levels of the (B) Alp, (C) Runx2, (D)
Ocn and (E) Spp1 osteogenic genes were measured. Their relative
changes were normalized to the GAPDH expression levels. The data
are presented as the mean ± standard deviation of three independent
experiments. ##P<0.01, ####P<0.0001 vs.
control group; *P<0.05, **P<0.01 vs. LPS group. BBR,
berberine hydrochloride; LPS, lipopolysaccharide; Runx2, RUNX
family transcription factor 2; Ocn, osteocalcin; Spp1, secreted
phosphoprotein 1; ALP, alkaline phosphatase. |
BBR reduces adipogenic differentiation
of BMSCs under LPS treatment
Basic effects of BBR on adipogenic gene expression
were weak in undifferentiated BMSCs; however, it was hypothesized
that BBR could serve a role in the adipogenic process. The maturity
of adipose-like cells, which were treated with different
concentrations of BBR in combination with or without LPS during the
differentiated process, was investigated. As illustrated in the oil
red O staining images, cells treated with a higher BBR dose
exhibited fewer positive stained cells (Fig. 3A; upper panel). LPS increased the
number of mature cells further, while BBR co-treatment reversed the
effect in a dose-dependent manner (Fig.
3A; lower panel). It was found that BMSCs administrated with
BBR combined with LPS had more positive cells compared with those
treated with BBR alone. This suggested that BBR partly blocked
differentiation.
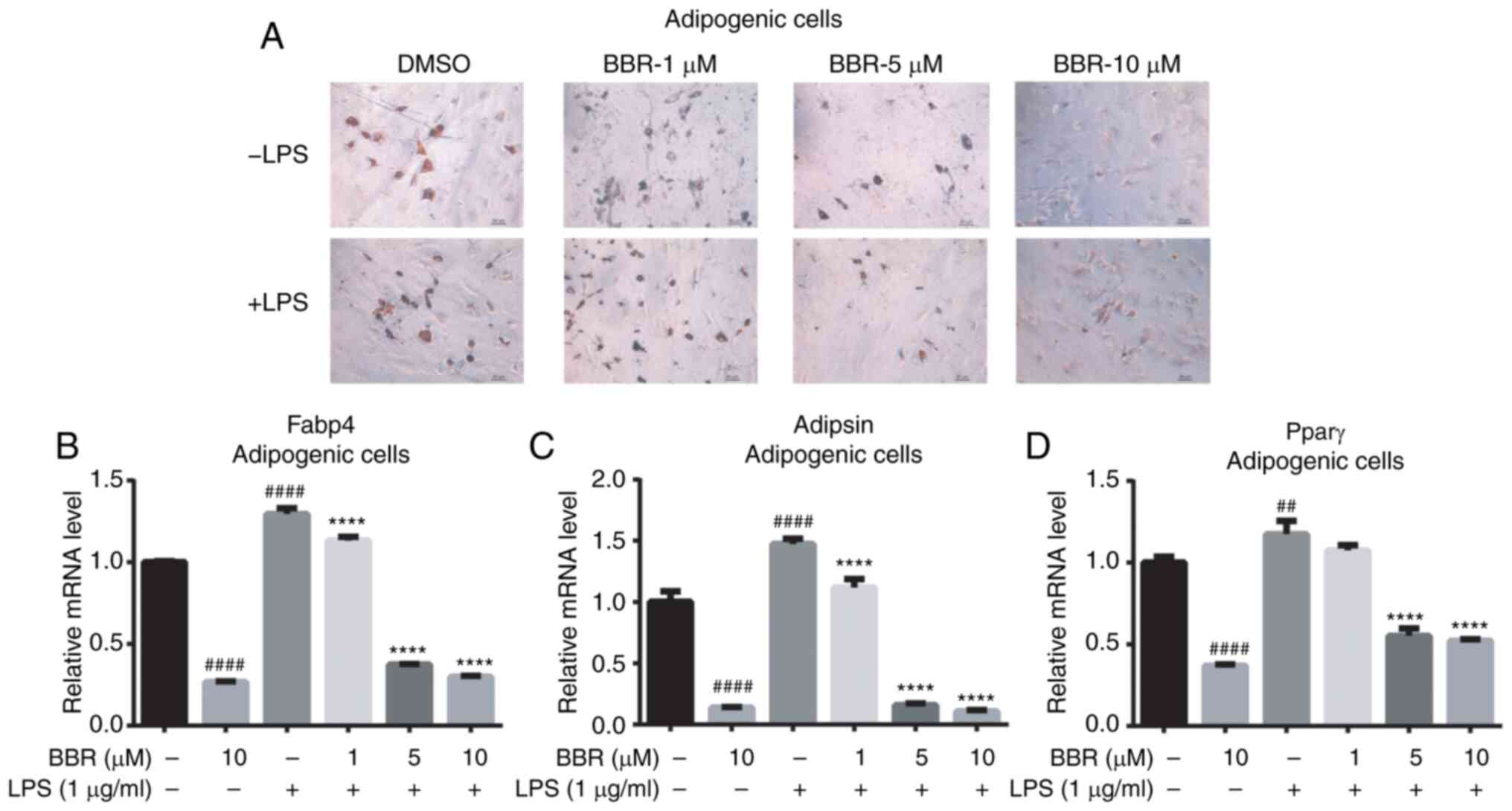 | Figure 3.BBR inhibits adipogenesis in bone
marrow-derived mesenchymal stem cells. The cells were incubated
with BBR (0, 1, 5 or 10 µM) alone or simultaneously with LPS (1
µg/ml) in adipogenic media for 12 days. Then, the cells were
detected using oil red O stain and reverse
transcription-quantitative PCR analysis. (A) Images of the ALP
stained cells were obtained using a microscope, at ×200
magnification. The mRNA expression levels of the (B) Fabp4, (C)
Adipsin and (D) Pparγ adipogenic genes were measured. Their
relative changes were normalized to the GAPDH expression levels.
The data are presented as the mean ± standard deviation of three
independent experiments. ##P<0.01,
####P<0.0001 vs. control group; ****P<0.0001 vs.
LPS group. BBR, berberine hydrochloride; LPS, lipopolysaccharide;
Fabp4, fatty acid binding protein 4; Pparγ, peroxisome
proliferator-activated receptorγ; ALP, alkaline phosphatase. |
The gene expression levels in differentiated BMSCs
were also detected. As presented in Fig. 3B-D, the mRNA expression levels of
all of the three adipogenic genes were decreased by BBR and
increased by LPS. Furthermore, BBR combined with LPS decreased the
LPS-stimulated Fabp4, Adipsin and Pparγ gene expression levels to a
lower level in a dose-dependent manner. Taken together, it was
found that BBR inhibited BMSCs adipogenesis and partly prevented
the LPS-mediated effect.
BBR ameliorates LPS-stimulated NF-κB
signaling activation and enhancement of pro-inflammatory factor
expression
As LPS activates cells and the expression of the
pro-inflammatory cytokines via the TLR4-NF-κB signaling pathway,
the effects of BBR on the primary proteins of the NF-κB signaling
pathway were investigated. It is known that p-IκBα and p-NF-κB p65
subunits are markers of TLR4/NF-κB pathway activation (27). BMSCs were cultured and treated with
BBR and LPS, alone or in combination, for another 2h. It was found
that the LPS-treated group exhibited darker p-p65 and p-IκBα bands
compared with those in the control group and in the BBR-treated
group (Fig. 4A). There was no
effect from BBR alone; however, BBR decreased the LPS-stimulated
activated protein expression levels. Densitometry analysis of the
bands is shown in Fig. 4B and C.
Notably, LPS treatment significantly increased the phosphorylation
of the NF-κB p65 subunit and IκBα, and these increments were
reversed when exposed to BBR simultaneously.
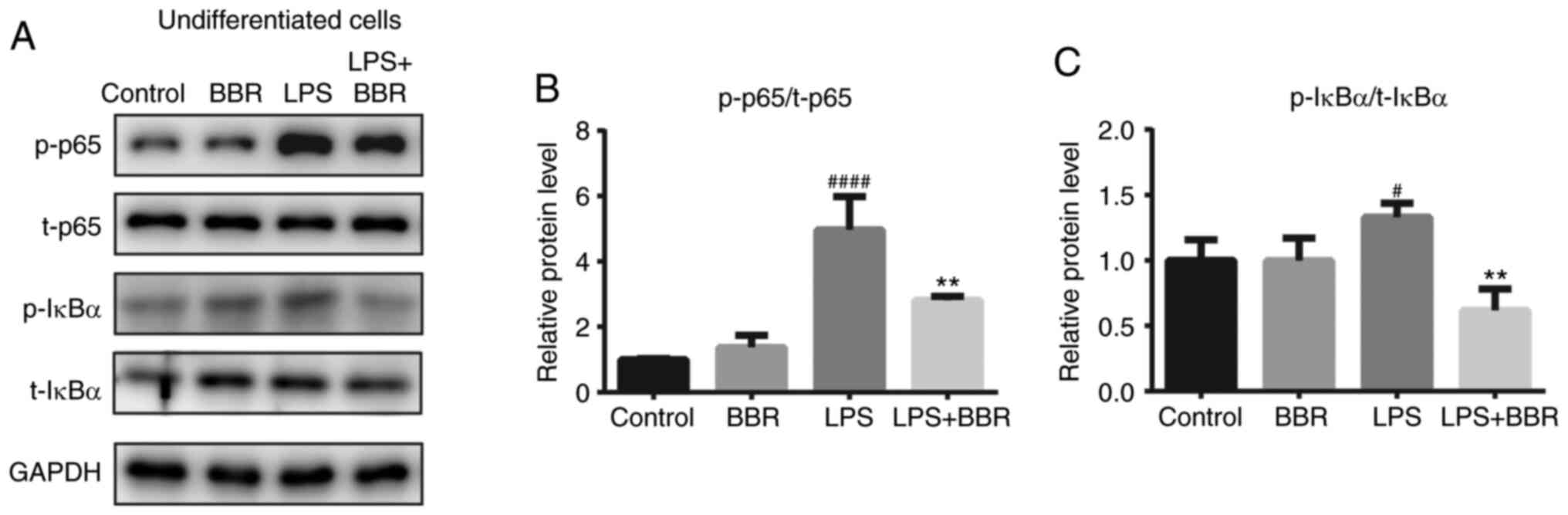 | Figure 4.BBR protects bone marrow-derived
mesenchymal stem cells from LPS-induced NF-κB activation under
undifferentiated condition. Cells were incubated with DMSO (as the
control), BBR (10 µM), LPS (1 µg/ml) or both BBR and LPS for 2h.
Then, the cells were harvested for western blot analysis. (A) Bands
of the p-and t- (B) NF-κB p65 subunit, (C) IκBα and GAPDH protein
expression levels are shown and the results were semi-quantified.
The data are presented as the mean ± standard deviation of three
independent experiments. #P<0.05,
####P<0.0001 vs. control group; **P<0.01 vs. LPS
group. T, total, p, phosphorylated; BBR, berberine hydrochloride;
LPS, lipopolysaccharide; p-, phosphorylated; t-, total. |
Subsequently, the product of the activated NF-κB
signaling pathway was investigated using RT-qPCR, including the
mRNA expression levels of TNF-α, MCP-1, Il-6 and Il-1β. BMSCs were
differentiated and treated as aforementioned, then mature
osteoblast cells and adipose-like cells were harvested and
detected. In osteogenic BMSCs (Fig.
5A-D), BBR alone had little effect on the pro-inflammatory
factors except MCP-1; however, LPS significantly increased TNF-α,
MCP-1, Il-6 and Il-1β gene expression to a great extent. Moreover,
the reversal effect of BBR against LPS was not strong. However, a
dose-dependent change was still found, particularly on MCP-1 mRNA
expression levels.
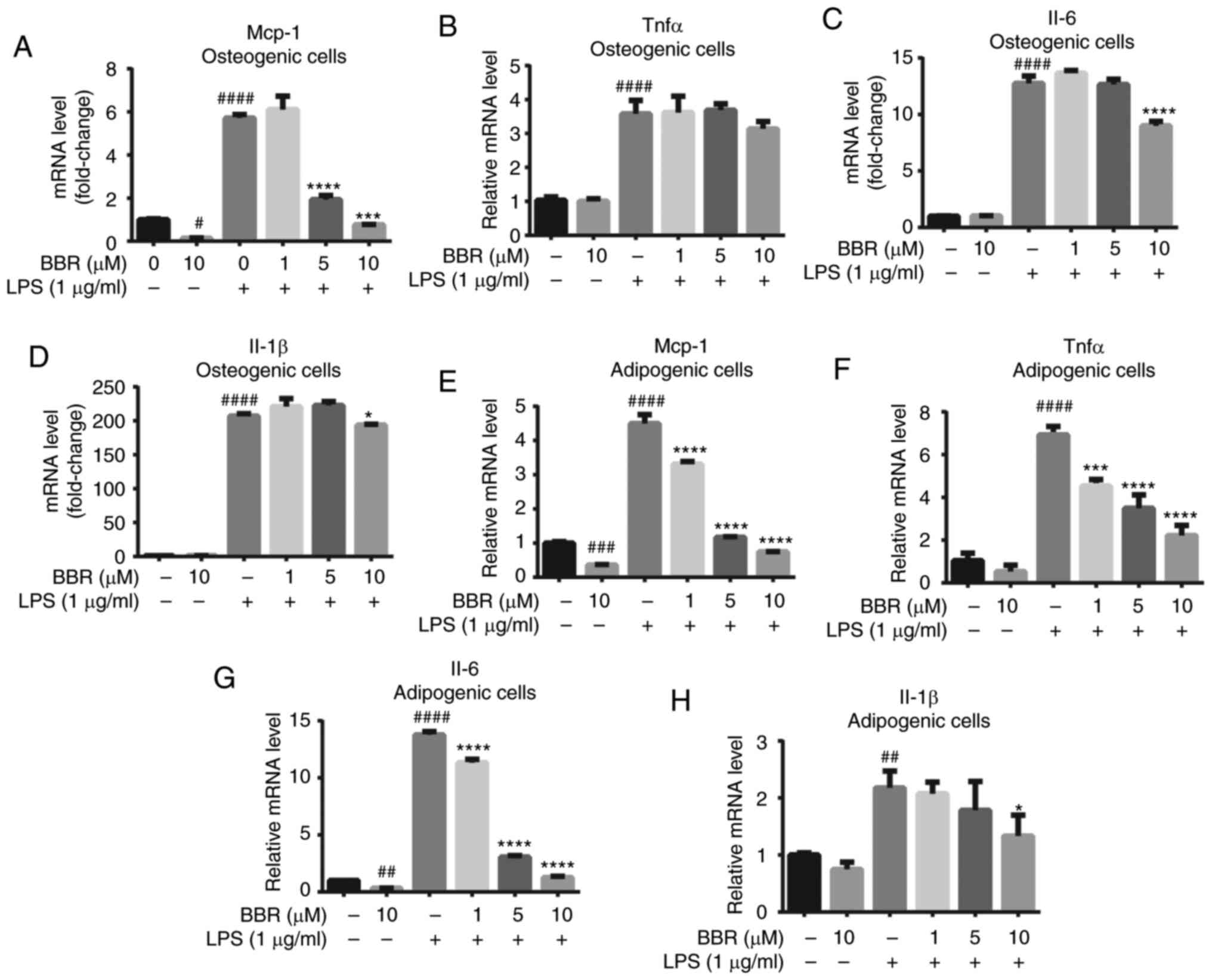 | Figure 5.BBR alleviates LPS-induced
inflammatory factor expression in both osteogenic and adipogenic
differentiated bone marrow-derived mesenchymal stem cells. The
cells were incubated with BBR (0, 1, 5 or 10 µM) or simultaneously
with LPS (1 µg/ml) in osteogenic media for 7 days or in adipogenic
media for 12 days. Then, the cells were harvested for reverse
transcription-quantitative PCR analysis. The mRNA expression levels
of the pro-inflammatory factors (A) MCP-1, (B) TNF-α, (C) Il-6 and
(D) Il-1β were measured in differentiated osteoblastic cells. (E)
MCP-1, (F) TNF-α, (G) Il-6 and (H) Il-1β expression levels were
also measured in adipogenic differentiated cells. Their relative
changes were normalized to GAPDH levels. The data are presented as
the mean ± standard deviation of three independent experiments.
#P<0.05, ##P<0.01,
###P<0.001, ####P<0.0001 vs. control
group; *P<0.05, ***P<0.001, ****P<0.0001 vs. LPS group.
BBR, berberine hydrochloride; LPS, lipopolysaccharide; MCP-1,
monocyte chemoattractant protein-1. |
The regulation of BBR was notable in adipogenic
BMSCs. The gene expression levels of MCP-1, Il-6 and Il-1β were
alleviated with BBR treatment. LPS stimulated a 4.5-, 6.9-, 13.7-
and 2.1-fold increase in the MCP-1, TNF-α, Il-6 and Il-1β mRNA
expression levels, respectively. Furthermore, BBR co-treatment
caused a significant and dose-dependent decrease in these
expression levels (Fig. 5E-H). At
10 µM concentration, BBR almost reversed the mRNA expression levels
of MCP-1 and Il-6 back to the basic level.
All the aforementioned results suggested that BBR
efficiently decreased the mRNA expression levels of
pro-inflammatory factors in both types of cells and may act more
sensitively in adipogenic cells.
BBR regulates the balance of
osteogenic/adipogenic differentiation partly via AMP-activated
protein kinase (AMPK) activation
As BBR typically acts as an adenosine AMPK agonist
(28), it was hypothesized whether
its regulation of osteogenesis/adipogenesis balance was due to AMPK
activation. The efficiency of BMSC osteogenic and adipogenic
differentiation with respective compound treatments was
subsequently investigated.
For osteogenic differentiation, compared with that
in the control cells, cells incubated with BBR stained darker,
while compound C (AMPK antagonist)-treated cells stained lighter
(Fig. 6A and B). The color of the
stain in BBR and compound C co-incubated cells was darker compared
with that in compound C only treated cells, and lighter compared
with that in cells treated with BBR only treated cells (Fig. 6A and B; upper panel). Similar
results were detected under the LPS-mediated inflammatory condition
(Fig. 6A and B; lower panel). In
addition, the intracellular ALP content was measured and the
results are presented in Fig. 6C.
The ALP content was weakly increased in the BBR group, while cells
treated with compound C and LPS had decreased ALP content. Compared
with that in the compound C group, the content of ALP in the BBR
+compound C group was significantly higher, suggesting the recovery
of ALP expression. Under inflammatory conditions, the LPS +
compound C group showed a lower ALP level compared with that in the
LPS, and LPS + BBR + compound C groups. Both the ALP staining and
intracellular content demonstrated that altered AMPK activation
affected osteogenic differentiation, and BBR could block the effect
of compound C.
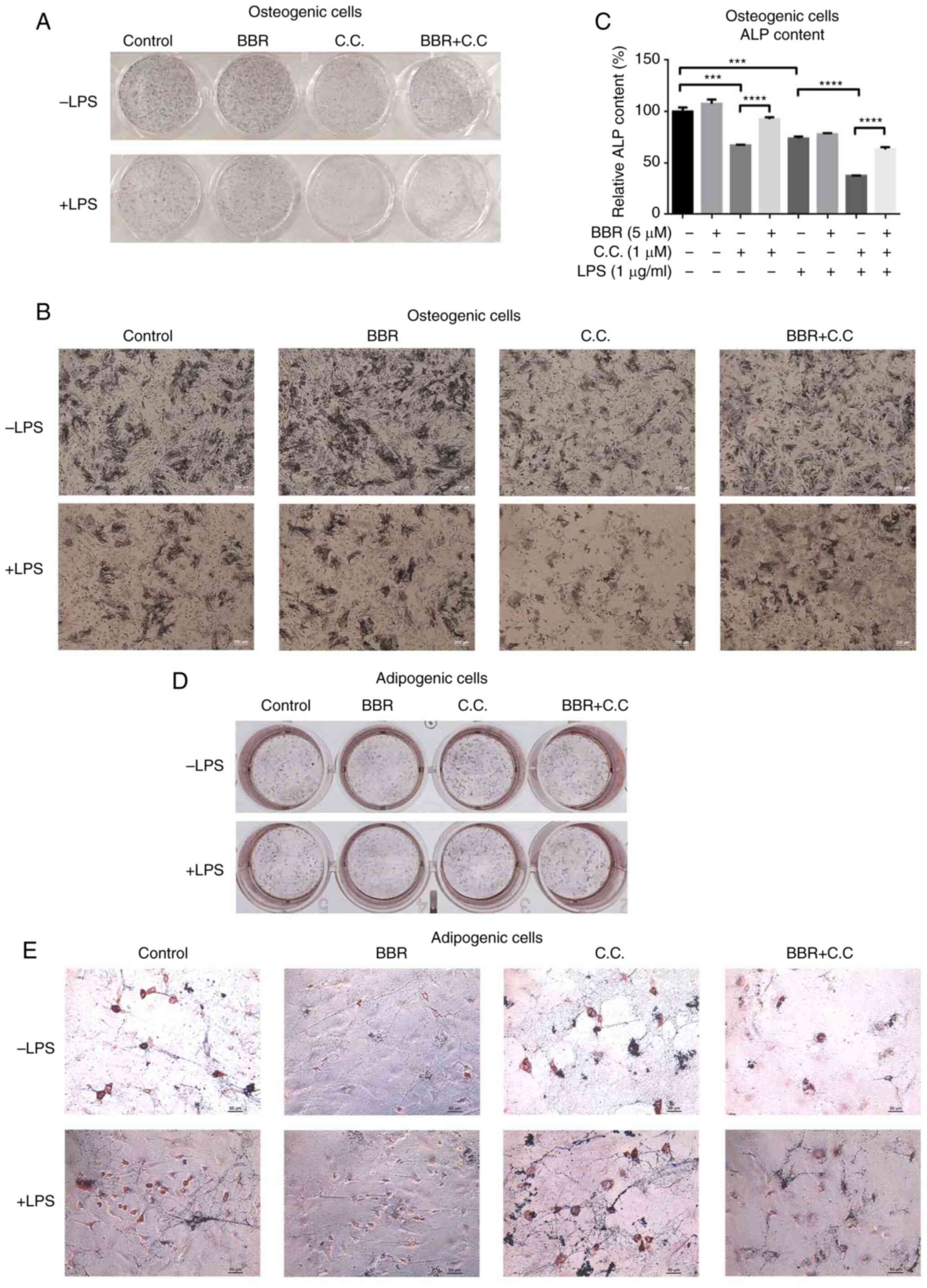 | Figure 6.BBR regulates osteogenic and
adipogenic differentiation via AMPK activation. The cells were
incubated with BBR (10 µM), C.C (1 µM) or both, in the presence or
absence of LPS (1 µg/ml), in osteogenic media for 7 days or in
adipogenic media for 12days. Then, the cells were collected for ALP
staining, ALP content assay or oil red O staining. The images of
the ALP stained cells were obtained using a (A) camera and (B)
microscope at ×200 magnification. (C) Relative ALP content levels
of osteogenic cells were measured and normalized to control levels.
The images of the oil red O stained cells were obtained using a (D)
camera and (E) microscope at ×200 magnification. The data are
presented as the mean ± standard deviation of three independent
experiments. ***P<0.001, ****P<0.0001. C.C, compound C; BBR,
berberine hydrochloride; LPS, lipopolysaccharide; ALP, alkaline
phosphatase. |
For adipogenic differentiation, the number of
positive cells in the BBR group was reduced compared with that in
the control group, while the compound C group had more positive
cells compared with that in the control group (Fig. 6D and E). The BBR + compound C group
had a higher number of positive cells compared with that in the
control group, but fewer compared with that in the compound C
group. On the other hand, compared with that in the control group,
LPS increased the number of positive cells, and the combination
group with compound C had even higher numbers (Fig. 6D and E). The LPS + BBR + compound C
group had less stained cells compared with that in the LPS +
compound C group. It was suggested that BBR reversed the
pro-adipogenic differentiation of LPS, and compound C disturbed the
anti-adipogenic function of BBR.
Discussion
Similar to periodontitis of teeth, peri-implantitis
of implanted teeth can occur. Inflammatory responses caused by
bacteria have been found to stimulate pro-inflammatory factor
upregulation and contribute to the pathogenesis of both
periodontitis and peri-implantitis (29). Therefore, controlling the
inflammatory reaction is an active strategy to solve the
peri-implant problem, by either reducing bacteria growth or
alleviating the accumulation of pro-inflammatory cytokines.
Notably, it would be beneficial to have both options. Previous
studies have reported that BBR was a broad-spectrum anti-bacterial
agent (30), and also exhibited
anti-inflammatory properties (28),
thus, presenting as a suitable candidate for peri-implantitis
therapy. Although BBR decreased the LPS-mediated inflammatory
response in several types of cells (28,31,32),
the anti-inflammatory effect of BBR in BMSCs is yet to be fully
elucidated. The present study demonstrated that BBR not only
attenuated the LPS-induced NF-κB p65 and IκBα activation in
undifferentiated BMSCs, but also downregulated the pro-inflammatory
factor expression levels in differentiated cells. These results
were consistent with the expected effect, confirming BBR as a
suitable candidate.
Besides anti-inflammation, bone regeneration is an
important aspect for peri-implantitis therapy. Thus, it should be
investigated whether a candidate would improve bone regeneration.
Firstly, previous studies have reported the pro-osteogenesis effect
of BBR (20,33); however, it was not clear whether the
action was sufficient to resist the effect by LPS. Secondly, LPS
could cause the imbalance of chondrogenesis and adipogenesis in
multi-potential BMSCs (18). The
function of LPS in regulating osteogenesis and adipogenesis balance
requires further investigation. In the present study, there was no
significant cytotoxic effect of LPS at 1 µg/ml concentration but
weak cytotoxicity was induced at the dose of 10 µg/ml. Notably, the
properties of LPS may vary at different concentrations. For
instance, LPS promoted adipogenesis at a dose of 10 µg/ml, but
attenuated adipogenic differentiation at a concentration of 100
ng/ml (18,34). Thus, in the current study, in order
to avoid the effect of cytotoxicity caused by high concentration of
LPS on cell differentiation, the dose of 1 µg/ml was selected for
subsequent experimentation. In a LPS-mediated inflammatory
condition, the regulation of BBR on the complex roles of
LPS-induced differentiation promotion or attenuation should also be
investigated.
To answer these questions, the changes in
undifferentiated and differentiated cells treated with BBR alone or
in combination with LPS were investigated. Understanding the basic
effect of BBR in undifferentiated cells may assist with predicting
osteogenic or adipogenic differentiation tendency in the complex
differentiated environment. In the present study, it was found that
BBR reversed the LPS-induced low mRNA expression levels of
osteogenic genes to a higher level (compared with that in the
control group) in undifferentiated cells; however, the changes in
the differentiated osteoblast were not as effective. Up to a 10 µM
concentration, BBR was not able to completely neutralize the
inhibition of LPS. Furthermore, the changes in the ALP protein
staining were moderate compared with that observed for the mRNA
expression level. Analogously, the resistant role of BBR against
LPS in regulating adipogenic gene expression level was weak in the
undifferentiated BMSCs; however, it was stronger in the mature
adipose-like cells. The oil red O stain assay further identified
the high efficiency of BBR. The extent of the variation maybe
different; however, the tendency of pro-osteogenic and
anti-adipogenic effects of BBR were found to be similar, regardless
of whether the BMSCs were differentiated or not. In other words,
LPS-mediated osteogenic and adipogenic differentiation was
rectified by BBR.
Given that BBR is an activator of AMPK, it was
hypothesized that BBR could modulate the balance of osteogenic and
adipogenic differentiation by activating AMPK. Therefore, an AMPK
inhibitor, compound C, was used. The present results identified
that compound C attenuated the effects of BBR; however, its strong
regulation of anti-osteogenesis and pro-adipogenesis was notable.
It was suggested that up- and down-regulation of AMPK could enhance
the pro-osteogenic and pro-adipogenic effects, accordingly. BBR has
been revealed to competitively bind with myeloid differentiation
protein (MD-2) and interfere with the binding of the TLR4/MD-2
complex with LPS (35,36); thus, BBR acts as a high-affinity LPS
antagonist. However, these molecular docking data were the result
of computer simulation, and not from experimental detection. In the
present study, BBR reversed the effects of LPS; however, this does
not suggest a direct interaction between BBR and LPS. Until now,
the mechanism of action was unknown, regarding whether it was
direct, indirect or mixed. Based on the aforementioned results,
further investigation is required to elucidate the direct
competitive antagonism. Elimination of LPS function by knockout or
mutation of the MD-2 gene may assist with identification or
exclusion of the direct method. Until this is identified, it can
only be hypothesized that BBR attenuated the LPS-mediated effects,
at least partly, via the activation of AMPK. Furthermore, BBR is a
multi-target molecule. For instance, ubiquitin-like with PHD and
RING Finger domains 1 is also a BBR target protein (37), which could negatively regulate Pparγ
in colorectal cancer (38). Whether
the ubiquitin-dependent proteasome system serves a role in the
osteogenic differentiated regulation in BMSCs also requires
confirmation.
In the present study, there was an interesting
finding that LPS stimuli decreased Pparγ mRNA expression level in
undifferentiated BMSCs but increased its gene expression level in
the adipogenic cells. Simonin et al (39) reported a decrease in Pparγ
expression induced by 10 µg/ml LPS in rat synovial fibroblasts.
Moreover, LPS (0.5 µg/ml) was found to downregulate Pparγ mRNA and
protein expression levels in vascular smooth muscle cells (40). The effects of LPS on Pparγ gene
expression in these cells were consistent with the current findings
in the undifferentiated BMSCs. On the other hand, similar to the
present results, another research team reported that LPS increased
Pparγ mRNA expression under the adipogenic condition (13). Sun et al (41) revealed that mineralization of ST2
cells is inhibited by PPARγ agonist troglitazone, while osteogenic
differentiation of BMSCs from osteoblast-targeted PPARγ knock-out
mice increases compared with wild type mice. Cortical bone
formation in the knock-out mice is also increased. In fact, Pparγ
is a protein with complex mechanism, which could inhibit certain
gene expression levels in untreated cells and enhance these in
LPS-stimulated cells (42). Thus,
it was suggested that the inhibition of Pparγ gene expression by
LPS in undifferentiated BMSCs may reflect a potential osteogenic
role. However, under adipogenic conditions, Pparγ expression and
activation was increased, and LPS further promoted Pparγ expression
via other mechanisms. This mechanism requires further study.
In summary, the present study demonstrated that BBR
ameliorated the LPS-induced imbalance of osteogenic and adipogenic
differentiation in BMSCs and promoted osteogenesis. The current
study provided a potential drug candidate, BBR, to solve the
peri-implantitis problem, with regards to its multiple role in
anti-bacterial capacity (43),
anti-inflammation action and osteogenesis.
Acknowledgements
Not applicable.
Funding
This work was supported by the Natural Science
Foundation of Shanghai (grant no. 19ZR1439600) and the Natural
Science Talent Cultivation Foundation of Shanghai Tenth People's
Hospital (grant no. 040318055).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YX, SQ, RZ and FC designed the research. RZ and HL
carried out the PCR analysis. FC and XZ performed the cell
differentiation and staining. XW isolated the primary cells. FY and
GS supervised the project and contributed to data analysis. YX, SQ
and RZ contributed to the writing of the manuscript. XZ and XW
helped with manuscript revision and SQ and YX confirmed the
authenticity of the data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All animal procedures performed in this work were
approved by the Animal Ethics Committee of Shanghai Tenth People's
Hospital, Shanghai, China (approval no. 2018-4487).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dreyer H, Grischke J, Tiede C, Eberhard J,
Schweitzer A, Toikkanen SE, Glöckner S, Krause G and Stiesch M:
Epidemiology and risk factors of peri-implantitis: A systematic
review. J Periodontal Res. 53:657–681. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Heitz-Mayfield LJ: Diagnosis and
management of peri-implant diseases. Aust Dent J. 53 (Suppl
1):S43–S48. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Daubert DM, Weinstein BF, Bordin S, Leroux
BG and Flemming TF: Prevalence and predictive factors for
peri-implant disease and implant failure: A cross-sectional
analysis. J Periodontol. 86:337–347. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mishler OP and Shiau HJ: Management of
peri-implant disease: A current appraisal. J Evid Based Dent Pract.
14 (Suppl):53–59. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Salvi GE, Cosgarea R and Sculean A:
Prevalence and mechanisms of peri-implant diseases. J Dent Res.
96:31–37. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Derks J and Tomasi C: Peri-implant health
and disease. A systematic review of current epidemiology. J Clin
Periodontol. 42 (Suppl 16):S158–S171. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Figuero E, Graziani F, Sanz I, Herrera D
and Sanz M: Management of peri-implant mucositis and
peri-implantitis. Periodontol 2000. 66:255–273. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hentenaar DFM, De Waal YCM, Van Winkelhoff
AJ, Meijer HJA and Raghoebar GM: Non-surgical peri-implantitis
treatment using a pocket irrigator device; clinical,
microbiological, radiographical and patient-centred outcomes-A
pilot study. Int J Dent Hyg. 18:403–412. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Takeuchi O, Hoshino K, Kawai T, Sanjo H,
Takada H, Ogawa T, Takeda K and Akira S: Differential roles of TLR2
and TLR4 in recognition of gram-negative and gram-positive
bacterial cell wall components. Immunity. 11:443–451. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cui L, Feng L, Zhang ZH and Jia XB: The
anti-inflammation effect of baicalin on experimental colitis
through inhibiting TLR4/NF-κB pathway activation. Int
Immunopharmacol. 23:294–303. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Park H, Noh AL, Kang JH, Sim JS, Lee DS
and Yim M: Peroxiredoxin II negatively regulates
lipopolysaccharide-induced osteoclast formation and bone loss via
JNK and STAT3. Antioxid Redox Signal. 22:63–77. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sul OJ, Park HJ, Son HJ and Choi HS:
Lipopolysaccharide (LPS)-induced autophagy is responsible for
enhanced osteoclastogenesis. Mol Cells. 40:880–887. 2017.PubMed/NCBI
|
|
13
|
Bonsignore LA, Anderson JR, Lee Z,
Goldberg VM and Greenfield EM: Adherent lipopolysaccharide inhibits
the osseointegration of orthopedic implants by impairing osteoblast
differentiation. Bone. 52:93–101. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nouneh RA, Wataha JC, Hanes PJ and
Lockwood PE: Effect of lipopolysaccharide contamination on the
attachment of osteoblast-like cells to titanium and titanium alloy
in vitro. J Oral Implantol. 27:174–179. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Insua A, Monje A, Wang HL and Miron RJ:
Basis of bone metabolism around dental implants during
osseointegration and peri-implant bone loss. J Biomed Mater Res A.
105:2075–2089. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen Q, Shou P, Zheng C, Jiang M, Cao G,
Yang Q, Cao J, Xie N, Velletri T, Zhang X, et al: Fate decision of
mesenchymal stem cells: Adipocytes or osteoblasts? Cell Death
Differ. 23:1128–1139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen KY, Chung CM, Chen YS, Bau DT and Yao
CH: Rat bone marrow stromal cells-seeded porous gelatin/tricalcium
phosphate/oligomeric proanthocyanidins composite scaffold for bone
repair. J Tissue Eng Regen Med. 7:708–719. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu J, Tang H, Zhang Z, Zhang Y, Qiu C,
Zhang L, Huang P and Li F: Kaempferol slows intervertebral disc
degeneration by modifying LPS-induced osteogenesis/adipogenesis
imbalance and inflammation response in BMSCs. Int Immunopharmacol.
43:236–242. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Pirillo A and Catapano AL: Berberine, a
plant alkaloid with lipid- and glucose-lowering properties: From in
vitro evidence to clinical studies. Atherosclerosis. 243:449–461.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee HW, Suh JH, Kim HN, Kim AY, Park SY,
Shin CS, Choi JY and Kim JB: Berberine promotes osteoblast
differentiation by Runx2 activation with p38 MAPK. J Bone Miner
Res. 23:1227–1237. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Han SY and Kim YK: Berberine Suppresses
RANKL-induced osteoclast differentiation by inhibiting c-Fos and
NFATc1 expression. Am J Chin Med. 47:439–455. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hu JP, Nishishita K, Sakai E, Yoshida H,
Kato Y, Tsukuba T and Okamoto K: Berberine inhibits RANKL-induced
osteoclast formation and survival through suppressing the NF-kappaB
and Akt pathways. Eur J Pharmacol. 580:70–79. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shanbhag S, Mohamed-Ahmed S, Lunde THF,
Suliman S, Bolstad AI, Hervig T and Mustafa K: Influence of
platelet storage time on human platelet lysates and platelet
lysate-expanded mesenchymal stromal cells for bone tissue
engineering. Stem Cell Res Ther. 11:3512020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang Y, Hou Q, Su H, Chen D, Luo Y and
Jiang T: miR-488 negatively regulates osteogenic differentiation of
bone marrow mesenchymal stem cells induced by psoralen by targeting
Runx2. Mol Med Rep. 20:3746–3754. 2019.PubMed/NCBI
|
|
26
|
Lee YC, Chan YH, Hsieh SC, Lew WZ and Feng
SW: Comparing the osteogenic potentials and bone regeneration
capacities of bone marrow and dental pulp mesenchymal stem cells in
a rabbit calvarial bone defect model. Int J Mol Sci. 20:50152019.
View Article : Google Scholar
|
|
27
|
Zhang J, Zheng Y, Luo Y, Du Y, Zhang X and
Fu J: Curcumin inhibits LPS-induced neuroinflammation by promoting
microglial M2 polarization via TREM2/ TLR4/ NF-κB pathways in BV2
cells. Mol Immunol. 116:29–37. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zou K, Li Z, Zhang Y, Zhang HY, Li B, Zhu
WL, Shi JY, Jia Q and Li YM: Advances in the study of berberine and
its derivatives: A focus on anti-inflammatory and anti-tumor
effects in the digestive system. Acta Pharmacol Sin. 38:157–167.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yan Q, Li Y, Cheng N, Sun W and Shi B:
Effect of retinoic acid on the function of
lipopolysaccharide-stimulated bone marrow stromal cells grown on
titanium surfaces. Inflamm Res. 64:63–70. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Stermitz FR, Lorenz P, Tawara JN, Zenewicz
LA and Lewis K: Synergy in a medicinal plant: Antimicrobial action
of berberine potentiated by 5′-methoxyhydnocarpin, a multidrug pump
inhibitor. Proc Natl Acad Sci USA. 97:1433–1437. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang H, Shan Y, Wu Y, Xu C, Yu X, Zhao J,
Yan J and Shang W: Berberine suppresses LPS-induced inflammation
through modulating Sirt1/NF-κB signaling pathway in RAW264.7 cells.
Int Immunopharmacol. 52:93–100. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liang Y, Fan C, Yan X, Lu X, Jiang H, Di
S, Ma Z, Feng Y, Zhang Z, Feng P, et al: Berberine ameliorates
lipopolysaccharide-induced acute lung injury via the PERK-mediated
Nrf2/HO-1 signaling axis. Phytother Res. 33:130–148. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tao K, Xiao D, Weng J, Xiong A, Kang B and
Zeng H: Berberine promotes bone marrow-derived mesenchymal stem
cells osteogenic differentiation via canonical Wnt/β-catenin
signaling pathway. Toxicol Lett. 240:68–80. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Fiedler T, Salamon A, Adam S, Herzmann N,
Taubenheim J and Peters K: Impact of bacteria and bacterial
components on osteogenic and adipogenic differentiation of
adipose-derived mesenchymal stem cells. Exp Cell Res.
319:2883–2892. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chu M, Ding R, Chu ZY, Zhang MB, Liu XY,
Xie SH, Zhai YJ and Wang YD: Role of berberine in anti-bacterial as
a high-affinity LPS antagonist binding to TLR4/MD-2 receptor. BMC
Complement Altern Med. 14:892014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nagai Y, Akashi S, Nagafuku M, Ogata M,
Iwakura Y, Akira S, Kitamura T, Kosugi A, Kimoto M and Miyake K:
Essential role of MD-2 in LPS responsiveness and TLR4 distribution.
Nat Immunol. 3:667–672. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gu C, Yin Z, Nie H, Liu Y, Yang J, Huang
G, Shen J, Chen L and Fei J: Identification of berberine as a novel
drug for the treatment of multiple myeloma via targeting UHRF1. BMC
Biol. 8:332020. View Article : Google Scholar
|
|
38
|
Sabatino L, Fucci A, Pancione M, Carafa V,
Nebbioso A, Pistore C, Babbio F, Votino C, Laudanna C, Ceccarelli
M, et al: UHRF1 coordinates peroxisome proliferator activated
receptor gamma (PPARG) epigenetic silencing and mediates colorectal
cancer progression. Oncogene. 31:5061–5072. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Simonin MA, Bordji K, Boyault S, Bianchi
A, Gouze E, Bécuwe P, Dauça M, Netter P and Terlain B: PPAR-gamma
ligands modulate effects of LPS in stimulated rat synovial
fibroblasts. Am J Physiol Cell Physiol. 282:C125–C133. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Qu R-N and Qu W: Metformin inhibits
LPS-induced inflammatory response in VSMCs by regulating TLR4 and
PPAR-γ. Eur Rev Med Pharmacol Sci. 23:4988–4995. 2019.PubMed/NCBI
|
|
41
|
Sun H, Kim JK, Mortensen R, Mutyaba LP,
Hankenson KD and Krebsbach PH: Osteoblast-targeted suppression of
PPARγ increases osteogenesis through activation of mTOR signaling.
Stem Cells. 31:2183–2192. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Sergeeva MG, Aleshin SE, Grabeklis S and
Reiser G: PPAR activation has dichotomous control on the expression
levels of cytosolic and secretory phospholipase A2 in astrocytes;
inhibition in naïve, untreated cells and enhancement in
LPS-stimulated cells. J Neurochem. 115:399–410. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huang X, Wang P, Li T, Tian X, Guo W, Xu
B, Huang G, Cai D, Zhou F, Zhang H, et al: Self-assemblies based on
traditional medicine berberine and cinnamic acid for
adhesion-induced inhibition multidrug-resistant Staphylococcus
aureus. ACS Appl Mater Interfaces. 12:227–237. 2020. View Article : Google Scholar : PubMed/NCBI
|














