Introduction
Ovarian cancer is one of the most common lethal
gynecological malignancies worldwide, with ~240,000 newly diagnosed
cases each year globally (1). In
2018, ovarian cancer was the fifth leading cause of
cancer-associated mortality in women, and the second most common
malignancy after breast cancer in women aged >40 years,
particularly in developed countries (2,3).
Although progress in surgical resection and aggressive treatment
with chemotherapy and radiotherapy has been achieved in recent
years, at present, there are no satisfactory treatments for
patients with late-stage disease; thus, the prognosis of ovarian
cancer remains poor (4). Therefore,
identifying the mechanisms underlying the progression of ovarian
cancer is important for the discovery of novel molecular targets
for diagnosis and treatment.
Kinesin (KIF) superfamily proteins are
microtubule-dependent molecular motors that exert multiple
functions in intracellular transport and cell division (5). Kinesin family member 4A (KIF4A), a KIF
protein, is an essential chromosome-associated molecular motor
encoding a 140-kDa protein (6).
Previous studies have revealed that KIF4A is involved in various
functions, including controlling chromosome condensation and
segregation, middle-spindle formation and cytokinesis during
mitotic division (7–9). Furthermore, previous studies have
demonstrated that KIF4A functions as an oncogene and serves crucial
roles in several malignancies, including colorectal (10), breast (11) and prostate cancer (12), as well as hepatocellular carcinoma
(13). Yang et al (14) identified abnormal expression levels
of KIF4A using ovarian cancer tissue data obtained from the Gene
Expression Omnibus database, and revealed that patients with
ovarian cancer with high KIF4A expression were more likely to
experience poor progression-free survival, indicating that KIF4A
may be involved in the pathogenesis of ovarian cancer. However, the
precise roles and mechanisms underlying KIF4A in ovarian cancer are
not completely understood.
Budding uninhibited by benzimidazoles 1 (BUB1) is a
mitotic checkpoint serine/threonine kinase that serves a critical
role in mitosis by phosphorylating members of the mitotic
checkpoint complex and activating the spindle checkpoint (15). Since BUB1 expression is upregulated
in several types of cancer, including gastric and prostate cancer,
and is closely associated with poor patient outcomes, emerging
studies have recognized BUB1 as an oncogene in tumors of diverse
origins (16,17). Notably, a potential interaction of
KIF4A and BUB1 has been predicted using Search Tool for the
Retrieval of Interacting Genes/Proteins (18). A bioinformatics analysis conducted
by Feng et al (19)
identified upregulated expression levels of BUB1 in ovarian cancer,
which were associated with poor prognosis. The aforementioned
results indicated that BUB1 might be involved in the progression of
ovarian cancer, which might also be associated with KIF4A.
The present study examined the expression levels of
KIF4A and BUB1, and their biological significance in ovarian cancer
cells, which provided a potential therapeutic target for ovarian
cancer.
Materials and methods
Bioinformatics analysis
Data from Gene Expression Profiling Interactive
Analysis (GEPIA; http://gepia2021.cancer-pku.cn/index.html) database,
which is a valuable and highly cited resource for gene expression
analysis based on tumor and normal samples from The Cancer Genome
Atlas and The Genotype-Tissue Expression databases, including 426
ovarian cancer tumor samples and 88 normal samples, were used to
analyze the expression levels of KIF4A.
Cell culture
The normal ovarian cell line (IOSE-80) was obtained
from Shanghai Yaji Biotechnology Co., Ltd. The human ovarian cancer
cell lines (ES-2, SKOV3, OVCAR3 and HEY-T30) were purchased from
American Type Culture Collection. A2780 cells were purchased from
BeNa Culture Collection (Beijing Beina Chunglian Biotechnology
Research Institute). IOSE-80, ES-2, HEY-T30 and SKOV3 cells were
cultured in RPMI-1640 (Biological Industries) supplemented with 10%
FBS (Gibco; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin. OVCAR3 and A2780 cells were cultured in
DMEM (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
FBS and 1% penicillin/streptomycin. All cells were cultured at 37°C
in a 5% CO2 incubator.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). RNA concentration and quality were measured at an absorbance
of 260/280 using a Nanodrop ND-100 spectrophotometer (Thermo Fisher
Scientific, Inc). Subsequently, RNA was reverse transcribed into
cDNA using the PrimeScript RT reagent kit (Takara Biotechnology
Co., Ltd.) according to the manufacturer's protocol. qPCR was
performed using SYBR Green PCR Master Mix (Applied Biosystems;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. The thermocycling conditions were as follows: 94°C for 5
min, 40 cycles of 94°C for 15 sec, 60°C for 25 sec and 72°C for 30
sec. The sequences of the gene primers were as follows: KIF4A
forward, 5′-CTGCAATTGGTTGGCGTCTC-3′ and reverse,
5′-CAGCGCCACTCTTACAGGAA-3′; BUB1 forward,
5′-TGGGAAAGATACATACAGTGGGT-3′ and reverse,
5′-AGGGGATGACAGGGTTCCAAT-3′; and β-actin forward,
5′-GGCACCCAGCAATGAA-3′ and reverse, 5′-TAGAAGCATTTGCGGTGG-3′. mRNA
expression levels were quantified using the 2−∆∆Cq
method (20) and normalized to the
internal reference gene β-actin.
Western blotting
Total protein was extracted from cells using RIPA
buffer containing protease inhibitor (Beyotime Institute of
Biotechnology). Following quantification using a BCA assay,
proteins (30 µg/lane) were separated via 12% SDS-PAGE, and
subsequently transferred to PVDF membranes. Following blocking with
5% skimmed milk at room temperature for 2 h, the membranes were
incubated at 4°C overnight with primary antibodies (all purchased
from Abcam) targeted against: KIF4A (1:1,000; cat. no. ab124903),
BUB1 (1:2,000; cat. no. ab9000), matrix metallopeptidase (MMP)2
(1:1,000; cat. no. ab92536), MMP9 (1:1,000; cat. no. ab38898),
Bcl-2 (1:1,000; cat. no. ab196495), Bax (1:1,000; cat. no.
ab182733), cleaved caspase3 (1:500; cat. no. ab32042), caspase3
(1:500; cat. no. ab13847) and GAPDH (1:1,000; cat. no. ab181603).
Subsequently, the membranes were incubated with HRP-conjugated goat
anti-rabbit (1:5,000; cat. no. sc-2004; Santa Cruz Biotechnology,
Inc.) secondary antibodies at room temperature for 2 h. Protein
bands were visualized using Pierce™ ECL Western Blotting Substrate
(Pierce; Thermo Fisher Scientific, Inc.) and semi-quantified using
ImageJ software (version 1.52; National Institutes of Health).
Cell transfection
The short hairpin RNA (shRNA) against KIF4A
(shRNA-KIF4A-1/2) and its negative control (shRNA-NC) were obtained
from Shanghai GenePharma Co., Ltd. cDNA encoding BUB1 was amplified
and inserted into the pcDNA3.1 vector (Shanghai GenePharma Co.,
Ltd.). An empty vector obtained from Shanghai GenePharma Co., Ltd.,
was used as the negative control (pcDNA-NC) for pcDNA-BUB1. OVCAR3
cells (5×105 cells/well) were transfected with 100 nM
shRNA-KIF4A1/2, pcDNA-BUB1 or the corresponding negative controls
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Following a 48-h incubation at 37°C, KIF4A or BUB1 expression
levels in transfected cells were determined via RT-qPCR and western
blotting.
Cell proliferation assay
OVCAR3 cells were seeded into 96-well plates
(2×103 cells/well) and cultured for 24, 48 or 72 h. At
each indicated time point, 10 µl Cell Counting Kit-8 (CCK-8)
solution (Dojindo Molecular Technologies, Inc.) was added into each
well and incubated for 2 h at 37°C. The optical density of each
well was measured at a wavelength of 450 nm using a microplate
reader.
Colony formation assay
OVCAR3 cells were plated into 6-well plates
(1×103 cells/well) and cultured for 10 days. The culture
medium was changed every 3 days. Cells were fixed in 4%
paraformaldehyde for 15 min at room temperature and stained with
0.5% crystal violet for 30 min at room temperature for
visualization. The number of clones (cell number >50
cells/colony) was counted with the naked eye or under a light
microscope (low magnification).
Wound healing assay
OVCAR3 cells were plated into 6-well plates
(1×105 cells/well). At 100% confluence, cell monolayers
were scratched using a sterile 10-µl pipette tip and washed with
PBS to remove debris. Subsequently, serum-free medium was added to
the plates. Images were captured at 0 and 48 h using a light
microscope (Leica Microsystems, Inc.). Cell migration was
calculated using the following formula: Migration rate (%) =
(migration distance/original distance) ×100%. The original distance
was defined as the width of the wound at 0 h, and the migration
distance was defined as the difference between the width of the
wound at 0 h and the width at 48 h. The relative migration rate was
presented as the fold-change relative to the control group.
TUNEL assay
The TUNEL assay was performed to assess cell
apoptosis. After washing with PBS, cells (4×103
cells/well) were fixed with 4% paraformaldehyde for 10 min at 4°C,
and treated with 1% Triton-100 for 30 min at room temperature and
3% H2O2 for 15 min at room temperature.
Subsequently, a TUNEL assay kit (Roche Diagnostics) was used to
detect apoptotic cells for 60 min at 37°C in the dark according to
the manufacturer's protocol, followed by an incubation with DAPI
for 10 min at room temperature in the dark. Apoptotic cells were
observed and imaged using fluorescence microscopy per unit area in
three fields.
Co-immunoprecipitation assay
Total protein was extracted from OVCAR3 cells using
RIPA buffer containing protease inhibitor (Beyotime Institute of
Biotechnology). Immunoprecipitation with specific antibodies
[anti-KIF4A (1:1,000; cat. no. ab124903; Abcam) and anti-BUB1
(1:2,000; cat. no. ab9000; Abcam)] was performed as previously
described (21). Immunoprecipitated
protein was subjected to western blotting as aforementioned, and
protein bands were visualized using Pierce™ ECL Western Blotting
Substrate (Pierce; Thermo Fisher Scientific, Inc.).
Immunofluorescence
OVCAR3 cells were seeded into 24-well plates
(2×104 cells/well). Following culture for 48 h, cells
were fixed in 4% paraformaldehyde at 4°C for 30 min and then
treated with 0.1% Triton-100 for 15 min at room temperature. Cells
were blocked with 5% BSA (Thermo Fisher Scientific, Inc.) at room
temperature for 1 h, followed by incubation with an anti-Ki67
primary antibody (1:200; cat. no. ab16667; Abcam) at 4°C overnight.
Subsequently, cells were incubated with Alexa Fluor®
488-conjugated secondary antibody (1:400; cat. no. A11008;
Molecular Probes; Thermo Fisher Scientific, Inc.) at room
temperature for 1 h, followed by incubation with DAPI for 3 min at
room temperature in the dark. Stained sections were observed and
imaged using fluorescence microscopy.
Flow cytometry assay
OVCAR3 cells were seeded into 6-well plates
(1×105 cells/well). At 48 h post-transfection, OVCAR3
cells were harvested in a centrifuge tube and fixed in 70% ethanol
at −20°C overnight. Following washing twice with PBS, cells were
incubated with 10 mg/ml RNase A and 1 mg/ml propidium iodide in the
dark at 37°C for 30 min. Cell cycle distribution was analyzed using
a FACSCalibur flow cytometer (BD Biosciences) with CellQuest Pro
software (version 5.1; Becton, Dickinson and Company).
Statistical analysis
Statistical analyses were performed using GraphPad
Prism (version 6.0; GraphPad Software, Inc.). Data are presented as
the mean ± SD from at least three experimental repeats. One-way or
two-way ANOVA followed by Tukey's post hoc test were used to
analyze comparisons among multiple groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
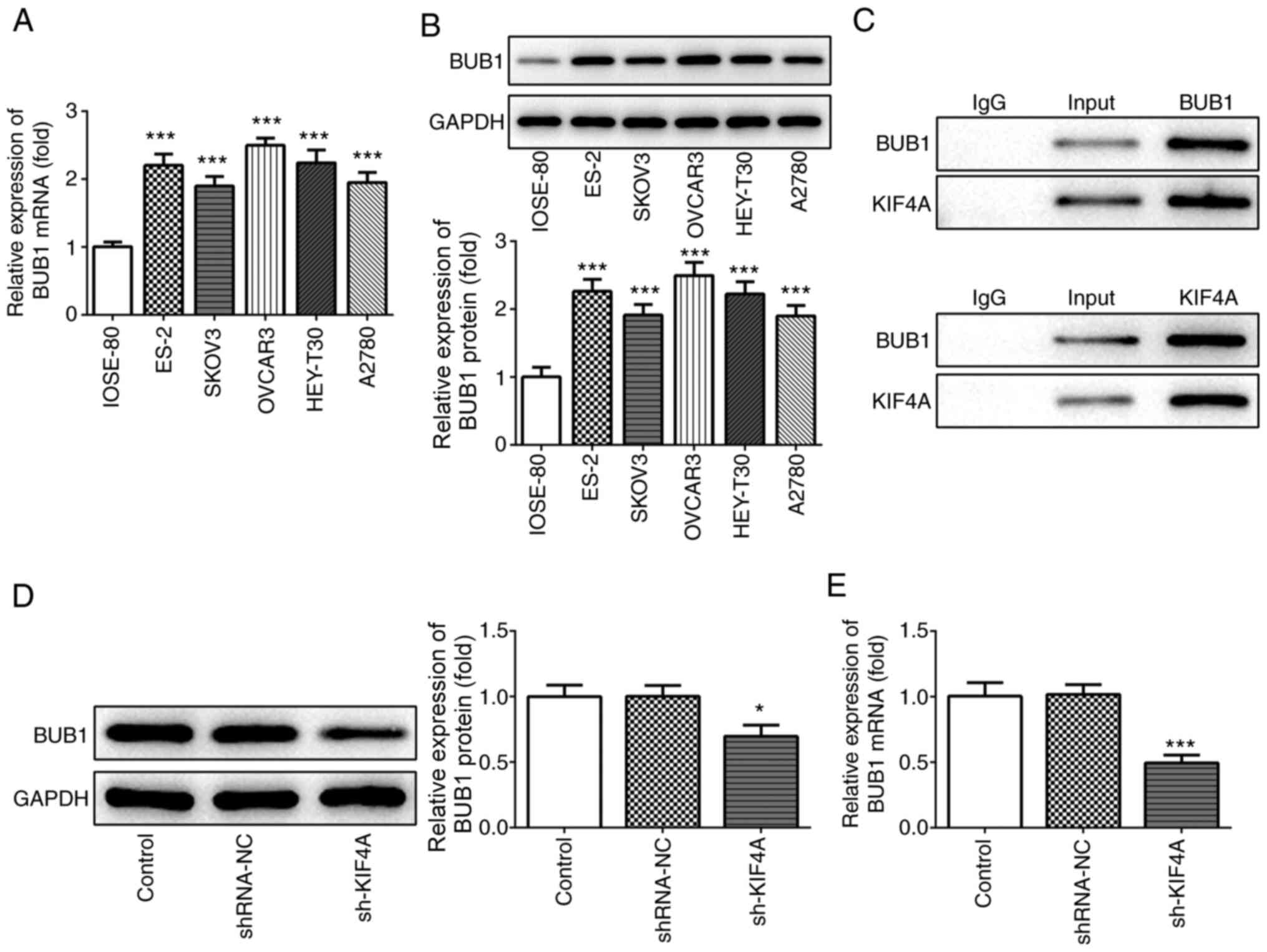
KIF4A expression is upregulated in
ovarian cancer
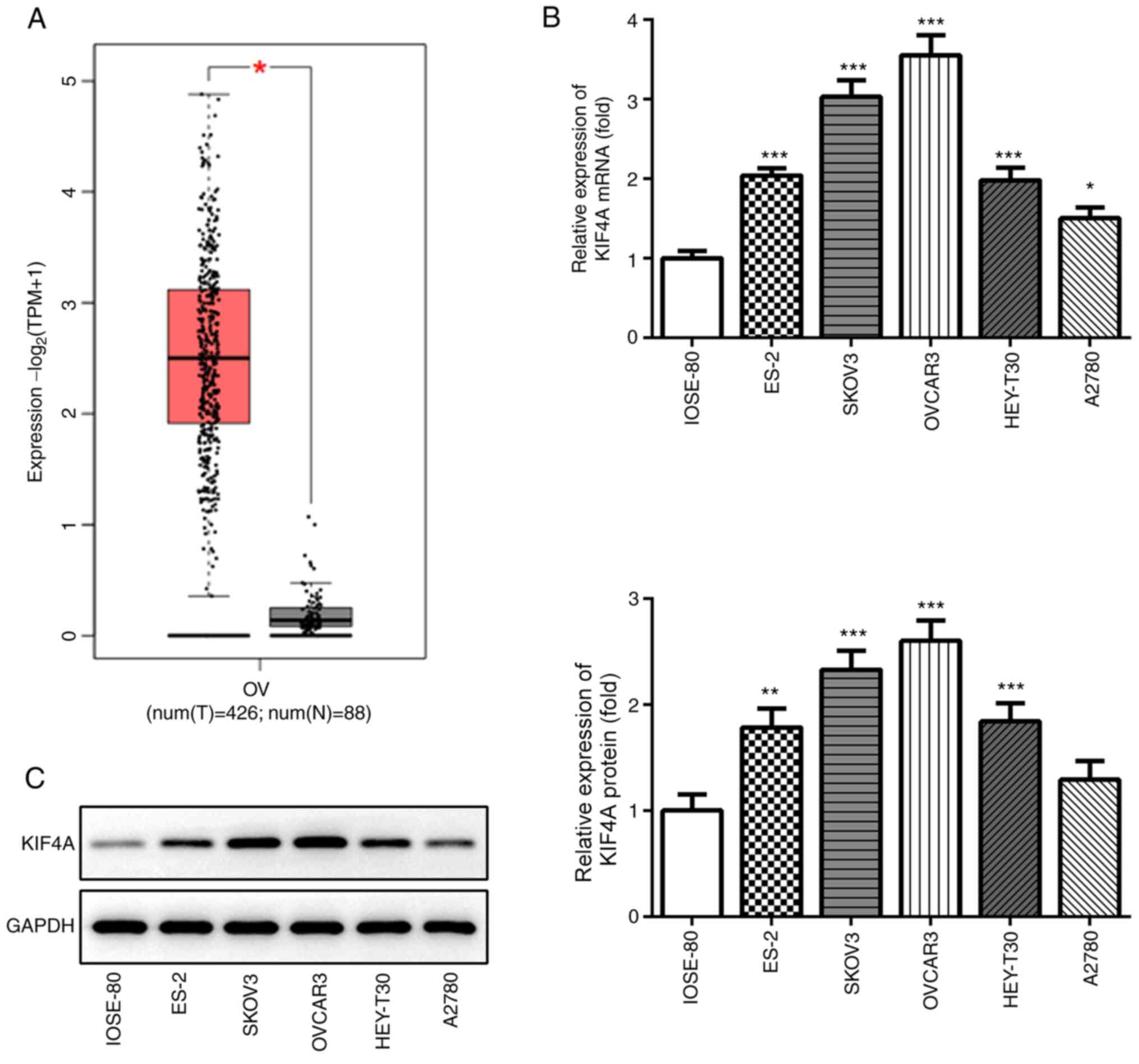
The bioinformatics analysis results using GEPIA
indicated that KIF4A expression levels were significantly
upregulated in ovarian cancer tumor samples compared with adjacent
healthy tissues (Fig. 1A).
Subsequently, KIF4A expression was detected in a normal ovarian
cell line (IOSE-80) and several ovarian cancer cell lines via
RT-qPCR and western blotting. KIF4A mRNA and protein expression
levels were significantly higher in ovarian cancer cell lines
compared with IOSE-80 cells, except KIF4A protein expression in
A2780 cells (Fig. 1B and C). Among
the ovarian cancer cell lines, KIF4A expression levels were highest
in OVCAR3 cells, which were selected for further experiments.
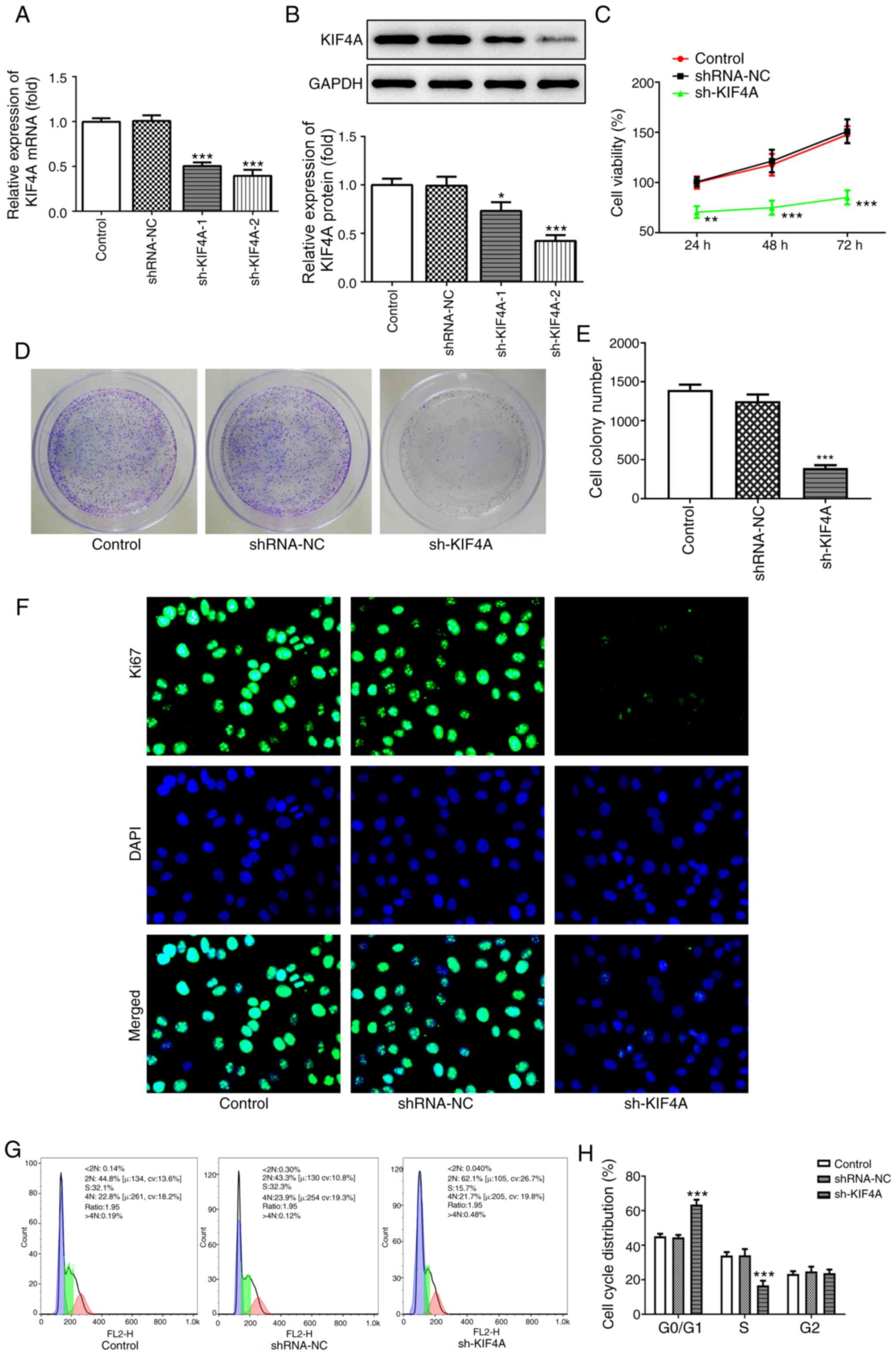
KIF4A knockdown inhibits OVCAR3 cell
proliferation and migration, and promotes OVCAR3 cell
apoptosis
To explore the molecular function of KIF4A in
ovarian cancer, cells were transfected with sh-KIF4A-1 and
sh-KIF4A-2. Compared with the shRNA-NC group, KIF4A mRNA and
protein expression levels were significantly downregulated
following transfection with sh-KIF4A-1 or sh-KIF4A-2 (Fig. 2A and B). Due to higher transfection
efficiency, sh-KIF4A-2 was used for subsequent experiments. A
series of cellular functional experiments were performed. Cell
viability and colony formation were significantly inhibited by
KIF4A knockdown compared with the shRNA-NC group (Fig. 2C-E). Furthermore, with increasing
incubation times, the difference in cell viability between the
shRNA-NC and sh-KIF4A groups increased. The immunofluorescence
assay results revealed that Ki67 expression levels in the sh-KIF4A
group were markedly decreased compared with the shRNA-NC group
(Fig. 2F). The flow cytometry assay
results demonstrated that compared with the shRNA-NC group, KIF4A
knockdown significantly induced cell cycle arrest at the G0/G1
phase, but significantly reduced the number of S phase cells
(Fig. 2G and H). Furthermore, the
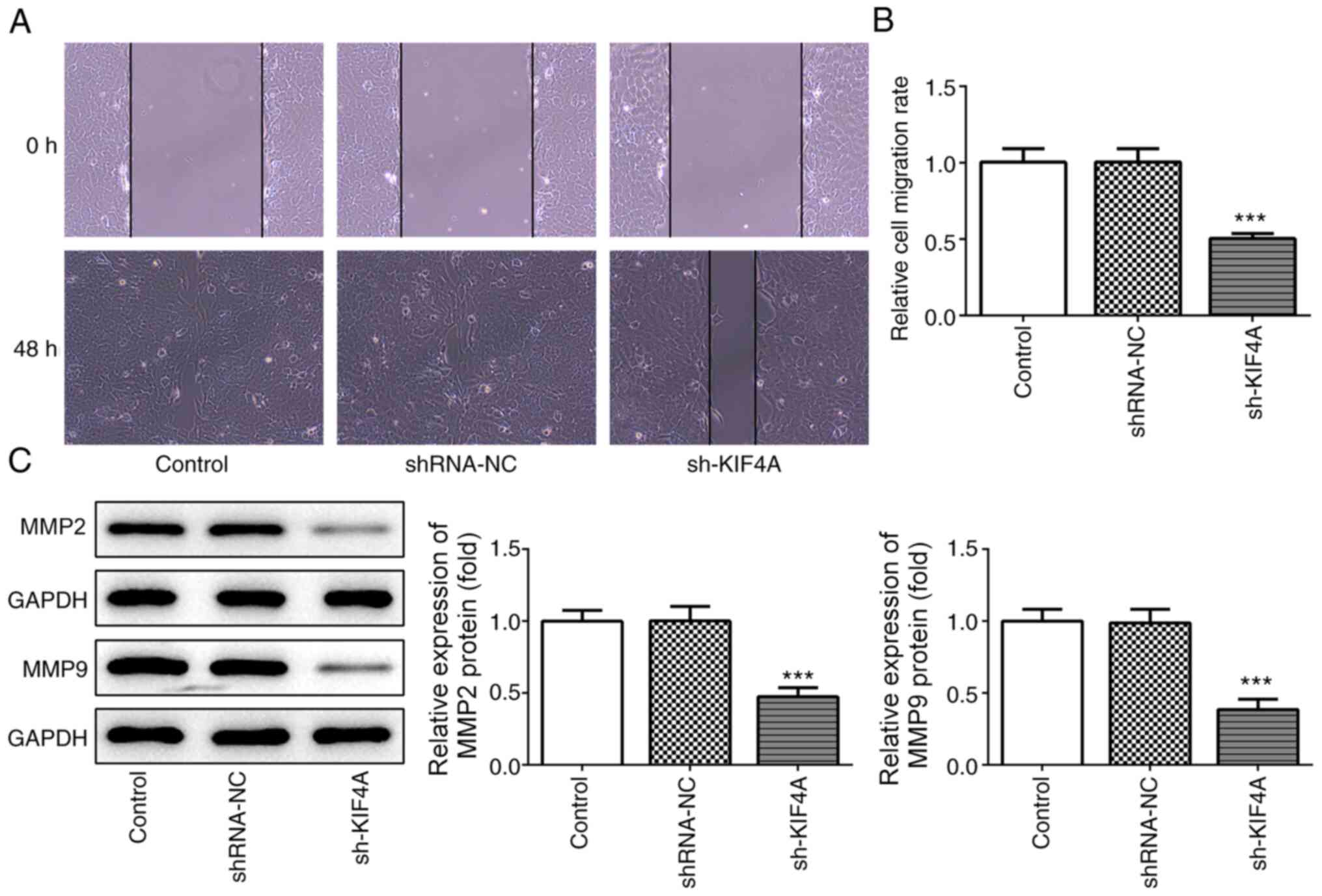
wound healing assay results demonstrated that cell migration was
significantly decreased by KIF4A knockdown compared with the
shRNA-NC group (Fig. 3A and B),
which was consistent with the decreased expression levels of MMP2
and MMP9 in KIF4A-knockdown OVCAR3 cells compared with
shRNA-NC-transfected OVCAR3 cells (Fig.
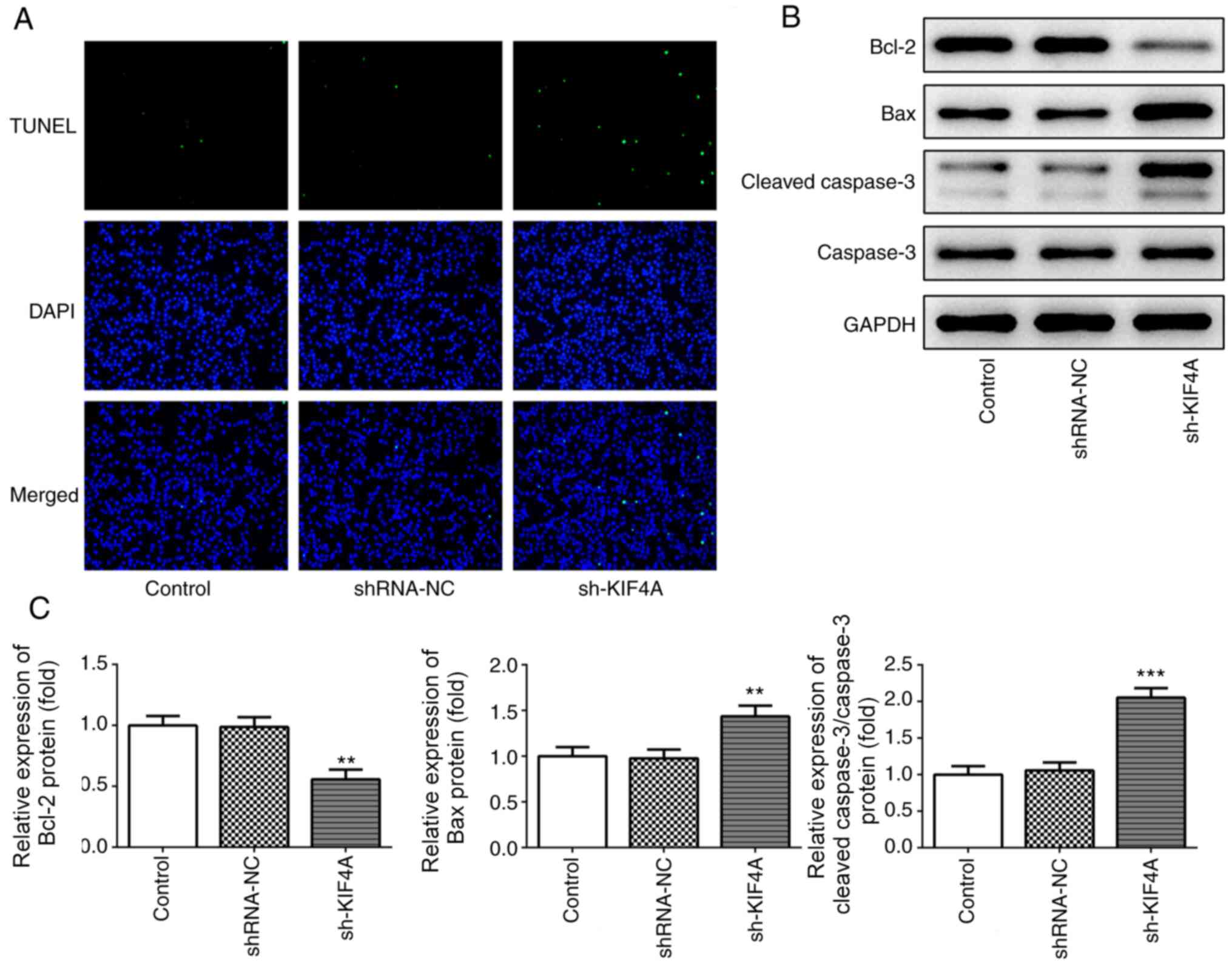
3C). In addition, the TUNEL assay results demonstrated that the
number of fluorescent green dots was notably increased following
transfection with sh-KIF4A in OVCAR3 cells compared with the
shRNA-NC group (Fig. 4A),
indicating that KIF4A knockdown markedly promoted cell apoptosis.
Compared with the shRNA-NC group, KIF4A knockdown significantly
downregulated Bcl-2 expression, an antiapoptotic indicator
(22), whereas Bax and cleaved
caspase3 expression levels, which are proapoptotic indicators
(22), were significantly
upregulated (Fig. 4B and C),
further demonstrating that KIF4A knockdown markedly promoted cell
apoptosis.
BUB1 expression is upregulated in
ovarian cancer cells and regulated by KIF4A
Subsequently, the expression levels of BUB1 were
detected in the normal ovarian cell line (IOSE-80) and several
ovarian cancer cell lines. BUB1 mRNA and protein expression levels
were significantly upregulated in ovarian cancer cell lines
compared with IOSE-80 cells (Fig. 5A
and B). The interaction of KIF4A and BUB1 was verified by
performing the co-immunoprecipitation assay (Fig. 5C). Furthermore, KIF4A knockdown
significantly decreased BUB1 mRNA and protein expression levels in
OVCAR3 cells compared with the shRNA-NC group (Fig. 5D and E), indicating that KIF4A
positively regulated BUB1 expression in ovarian cancer.
BUB1 overexpression reverses KIF4A
knockdown-mediated effects on ovarian cancer cells
To further investigate the regulatory mechanism
between KIF4A and BUB1 in ovarian cancer, OVCAR3 cells were
transfected with pcDNA-BUB1. Following transfection with
pcDNA-BUB1, BUB1 mRNA and protein expression levels were
significantly increased compared with the pcDNA-NC group (Fig. 6A and B). Subsequently, OVCAR3 cells
were co-transfected with sh-KIF4A and pcDNA-NC or pcDNA-BUB1. The
CCK-8 assay results demonstrated that BUBI overexpression
significantly decreased KIF4A knockdown-mediated inhibitory effects
on cell viability at the 48 and 72 h time points (Fig. 6C). Additionally, BUB1 overexpression
weakened KIF4A knockdown-mediated inhibitory effects on colony
formation and Ki67 expression (Fig. 6D
and E), indicating that KIF4A knockdown-induced inhibition of
cell proliferation was reversed by BUB1 overexpression.
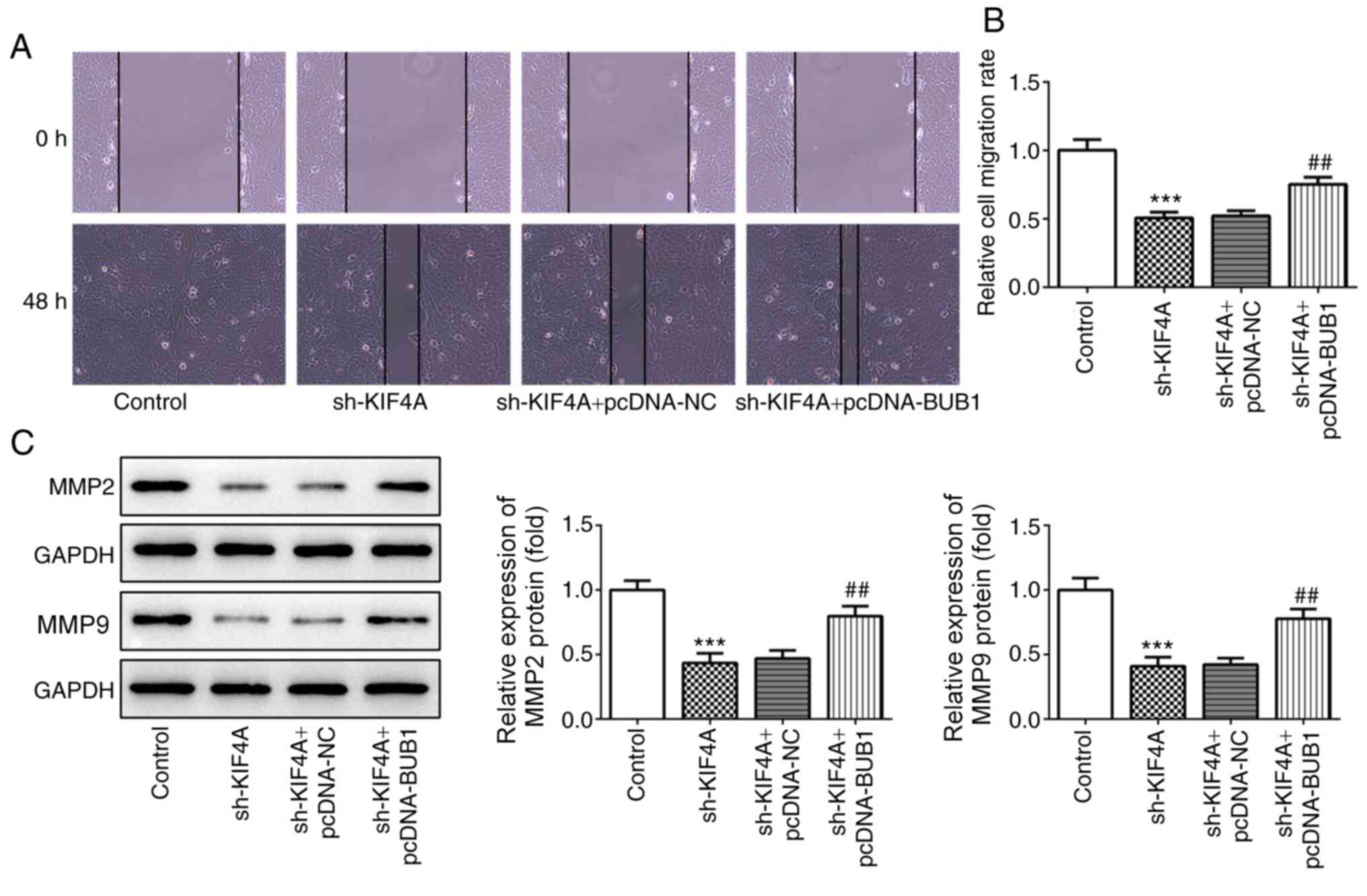
Furthermore, KIF4A knockdown-induced inhibition of cell migration
was also reversed by BUB1 overexpression (Fig. 7A and B), and KIF4A
knockdown-mediated downregulation of MMP2 and MMP9 expression
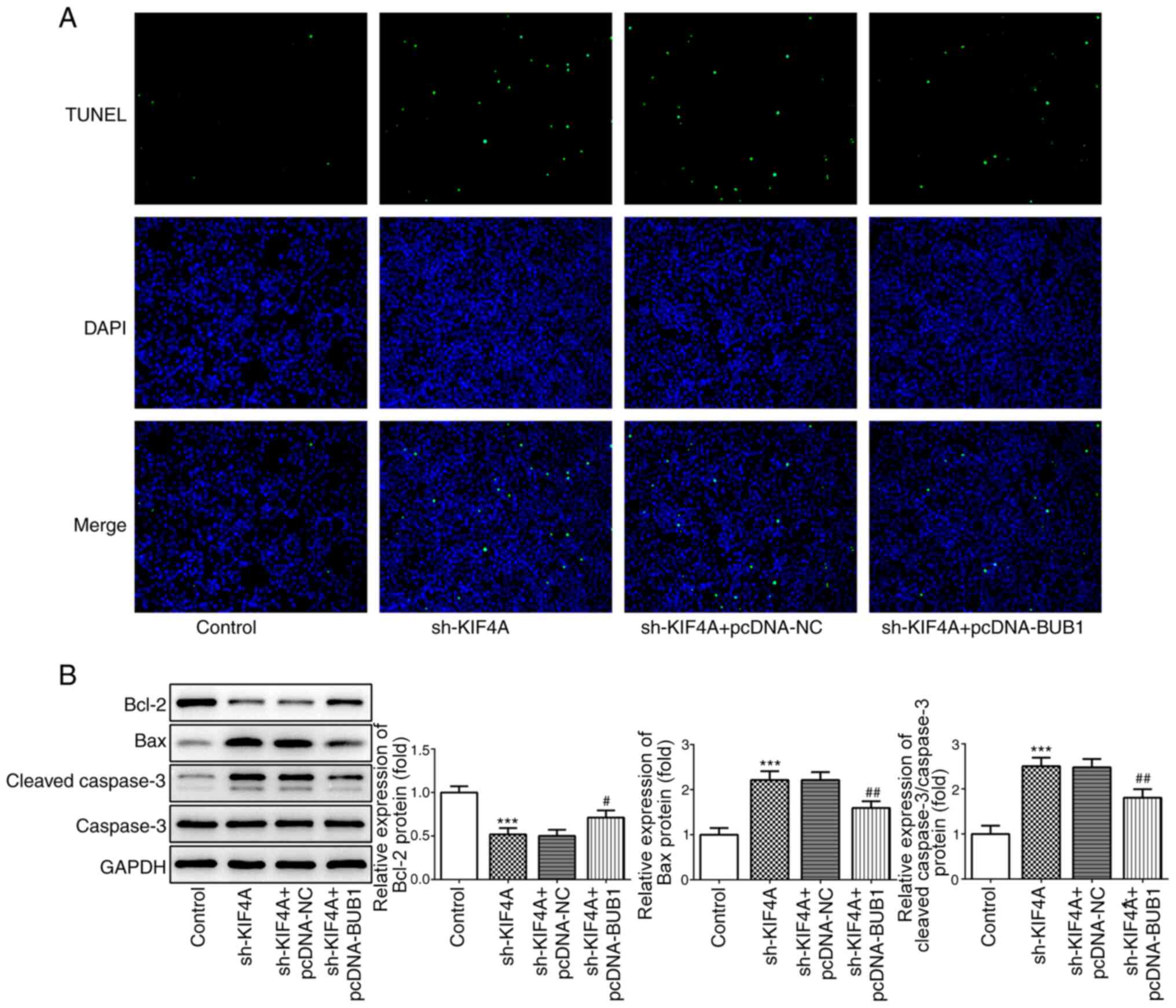
levels was inhibited by BUB1 overexpression (Fig. 7C). The TUNEL assay results
demonstrated that the number of apoptotic cells was reduced
following co-transfection with sh-KIF4A and pcDNA-BUB1 compared
with transfection with sh-KIF4A alone (Fig. 8A). In addition, compared with the
control group, KIF4A knockdown significantly downregulated Bcl-2
expression, but significantly upregulated Bax and cleaved caspase 3
expression levels, which was reversed by BUB1 overexpression
(Fig. 8B), suggesting that BUB1
overexpression promoted cell apoptosis.
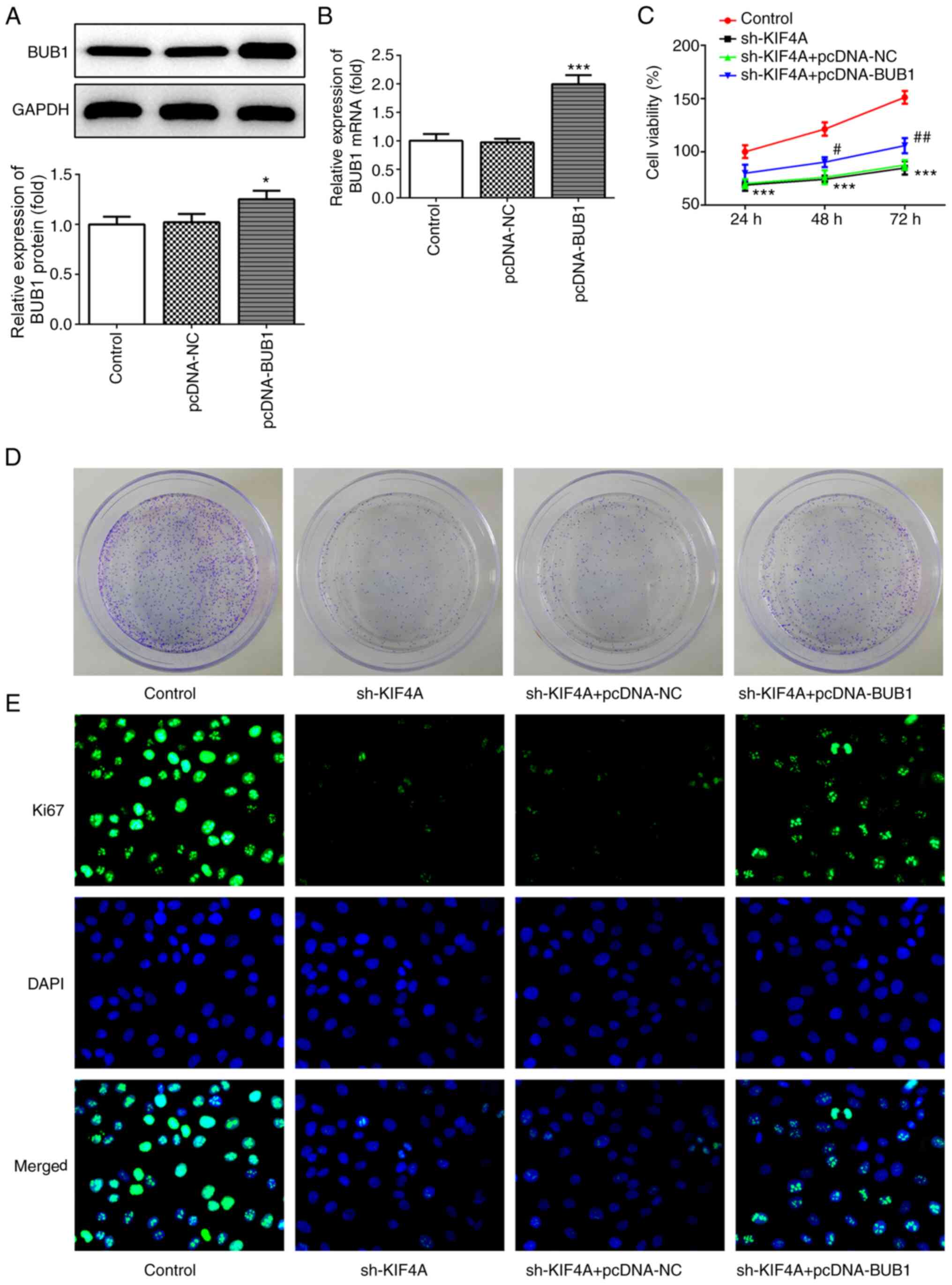 | Figure 6.BUB1 overexpression reverses
KIF4A-mediated inhibitory effects on cell proliferation. OVCAR3
cells were transfected with pcDNA-NC, pcDNA-BUB1, sh-KIF4A,
sh-KIF4A + pcDNA-NC or sh-KIF4A + pcDNA-BUB1. BUB1 (A) protein and
(B) mRNA expression levels were measured via western blotting and
reverse transcription-quantitative PCR, respectively. (C) Cell
viability was assessed by performing Cell Counting Kit-8 assays.
Cell colony formation was (D) determined by performing colony
formation assays. Magnification, ×1. (E) Ki67 expression was
detected via immunofluorescence. Magnification, ×400. *P<0.05
and ***P<0.001 vs. pcDNA-NC or control; #P<0.05
and ##P<0.01 vs. sh-KIF4A + pcDNA-NC. BUB1, budding
uninhibited by benzimidazoles 1; KIF4A, kinesin family member 4A;
NC, negative control; sh, short hairpin RNA. |
Discussion
Ovarian cancer is one of the most common lethal
gynecological malignancies worldwide, accounting for 2.5% of all
malignancies in women (23). Due to
the vague symptoms of ovarian cancer, and the lack of early
screening and detection techniques, 70–75% of patients with ovarian
cancer are first diagnosed at an advanced stage (24). Therefore, identifying and
understanding novel specific targets of ovarian cancer is important
to improve diagnosis and therapeutic strategies. The present study
demonstrated that KIF4A and BUB1 expression levels were
significantly increased in ovarian cancer cell lines compared with
IOSE-80 cells. Moreover, KIF4A interacted with BUB1 and positively
regulated BUB1 expression. By modulating BUB1, KIF4A knockdown
significantly inhibited ovarian cancer cell viability, colony
formation and migration, and markedly induced cell apoptosis
compared with the shRNA-NC group, hindering the development of
ovarian cancer. Therefore, the present study indicated that KIF4A
knockdown exerted anticancer activities in ovarian cancer; thus,
KIF4A knockdown might serve as a potential therapeutic strategy for
ovarian cancer.
KIF4A serves as an oncogene in various types of
cancer, and KIF4A can regulate cellular functions and behaviors to
influence the occurrence and development of malignancies. For
example, Hou et al (10)
demonstrated that KIF4A could enhance cell proliferation by
promoting cell cycle progression in vivo and in
vitro, contributing to the development of colorectal cancer.
Hou et al (13) reported
that KIF4A overexpression enhanced hepatocellular carcinoma cell
proliferation and migration, whereas KIF4A knockdown reduced cell
proliferation and migration, indicating the potential role of KIF4A
in mediating tumor initiation and progression. In the present
study, the results demonstrated that KIF4A expression was
significantly upregulated in ovarian cancer cell lines compared
with IOSE-80 cells. Moreover, compared with the shRNA-NC group,
KIF4A knockdown significantly inhibited cell viability, colony
formation and migration, and markedly promoted cell apoptosis,
revealing the potential value of KIF4A in regulating the
tumorigenesis of ovarian cancer.
BUB1 has been recognized as an oncogene in several
types of cancer. It has been reported that BUB1 downregulation can
inhibit cell proliferation, migration and invasion, and increase
the rate of apoptosis in hepatocellular carcinoma cells (25). Clinical gene analysis identified a
close association between BUB1 expression and poor clinical
outcomes in patients with breast cancer with metastasis (26). Additionally, BUB1 has been reported
to serve as a key proliferation-associated gene in non-small cell
lung cancer (27). The present
study demonstrated that BUB1 mRNA and protein expression levels
were significantly increased in ovarian cancer cell lines compared
with IOSE-80 cells. In addition, the co-immunoprecipitation assay
results further demonstrated the interaction between KIF4A and
BUB1. The subsequent functional experiments revealed that BUB1
overexpression weakened KIF4A knockdown-mediated inhibitory effects
on cell viability, colony formation and migration, and KIF4A
knockdown-induced cell apoptosis, indicating a potential mechanism
underlying the molecular action of KIF4A in ovarian cancer.
In conclusion, to the best of our knowledge, the
present study was the first to suggest that aberrant KIF4A
expression might regulate the progression of ovarian cancer.
Compared with the shRNA-NC group, KIF4A knockdown suppressed
ovarian cancer cell viability, colony formation and migration, and
promoted ovarian cancer cell apoptosis via downregulating BUB1
expression. The results of the present study suggested that
therapeutic strategies targeting the KIF4A/BUB1 axis might be
beneficial for the treatment of ovarian cancer. A key limitation of
the present study was the lack of investigation of the role of
KIF4A in ovarian cancer using other cell lines. Therefore, future
studies should use additional cell lines to verify the results of
the present study.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LY designed the study, performed the experiments,
interpreted the data and critically revised the manuscript. WJ
performed the experiments, analyzed the data and drafted the
manuscript. All authors read and approved the final manuscript. LY
was accountable for the work in ensuring that questions related to
the integrity of any part of the work are appropriately
investigated and resolved. LY and WJ confirm the authenticity of
all the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Webb PM and Jordan SJ: Epidemiology of
epithelial ovarian cancer. Best Pract Res Clin Obstet Gynaecol.
41:3–14. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Stewart C, Ralyea C and Lockwood S:
Ovarian Cancer: An Integrated Review. Semin Oncol Nurs. 35:151–156.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vargas AN: Natural history of ovarian
cancer. Ecancermedicalscience. 8:4652014.PubMed/NCBI
|
|
4
|
Wang L, Yan W, Li X, Liu Z, Tian T, Chen
T, Zou L and Cui Z: S100A10 silencing suppresses proliferation,
migration and invasion of ovarian cancer cells and enhances
sensitivity to carboplatin. J Ovarian Res. 12:1132019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rath O and Kozielski F: Kinesins and
cancer. Nat Rev Cancer. 12:527–539. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ha MJ, Yoon J, Moon E, Lee YM, Kim HJ and
Kim W: Assignment of the kinesin family member 4 genes (KIF4A and
KIF4B) to human chromosome bands Xq13.1 and 5q33.1 by in situ
hybridization. Cytogenet Cell Genet. 88:41–42. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mazumdar M, Sundareshan S and Misteli T:
Human chromokinesin KIF4A functions in chromosome condensation and
segregation. J Cell Biol. 166:613–620. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hu CK, Coughlin M, Field CM and Mitchison
TJ: KIF4 regulates midzone length during cytokinesis. Curr Biol.
21:815–824. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kurasawa Y, Earnshaw WC, Mochizuki Y,
Dohmae N and Todokoro K: Essential roles of KIF4 and its binding
partner PRC1 in organized central spindle midzone formation. EMBO
J. 23:3237–3248. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hou PF, Jiang T, Chen F, Shi PC, Li HQ,
Bai J and Song J: KIF4A facilitates cell proliferation via
induction of p21-mediated cell cycle progression and promotes
metastasis in colorectal cancer. Cell Death Dis. 9:4772018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xue D, Cheng P, Han M, Liu X, Xue L, Ye C,
Wang K and Huang J: An integrated bioinformatical analysis to
evaluate the role of KIF4A as a prognostic biomarker for breast
cancer. OncoTargets Ther. 11:4755–4768. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao H, Chen X, Cai Q, Shang Z and Niu Y:
Increased KIF4A expression is a potential prognostic factor in
prostate cancer. Oncol Lett. 15:7941–7947. 2018.PubMed/NCBI
|
|
13
|
Hou G, Dong C, Dong Z, Liu G, Xu H, Chen
L, Liu L, Wang H and Zhou W: Upregulate KIF4A Enhances
Proliferation, Invasion of Hepatocellular Carcinoma and Indicates
poor prognosis Across Human Cancer Types. Sci Rep. 7:41482017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang D, He Y, Wu B, Deng Y, Wang N, Li M
and Liu Y: Integrated bioinformatics analysis for the screening of
hub genes and therapeutic drugs in ovarian cancer. J Ovarian Res.
13:102020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bolanos-Garcia VM and Blundell TL: BUB1
and BUBR1: Multifaceted kinases of the cell cycle. Trends Biochem
Sci. 36:141–150. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fu X, Chen G, Cai ZD, Wang C, Liu ZZ, Lin
ZY, Wu YD, Liang YX, Han ZD, Liu JC, et al: Overexpression of BUB1B
contributes to progression of prostate cancer and predicts poor
outcome in patients with prostate cancer. OncoTargets Ther.
9:2211–2220. 2016.PubMed/NCBI
|
|
17
|
Stahl D, Braun M, Gentles AJ, Lingohr P,
Walter A, Kristiansen G and Gütgemann I: Low BUB1 expression is an
adverse prognostic marker in gastric adenocarcinoma. Oncotarget.
8:76329–76339. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
von Mering C, Huynen M, Jaeggi D, Schmidt
S, Bork P and Snel B: STRING: A database of predicted functional
associations between proteins. Nucleic Acids Res. 31:258–261. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feng H, Gu ZY, Li Q, Liu QH, Yang XY and
Zhang JJ: Identification of significant genes with poor prognosis
in ovarian cancer via bioinformatical analysis. J Ovarian Res.
12:352019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang X and Chen T: CUL4A regulates
endometrial cancer cell proliferation, invasion and migration by
interacting with CSN6. Mol Med Rep. 23:232021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Y, Yang X, Ge X and Zhang F:
Puerarin attenuates neurological deficits via Bcl-2/Bax/cleaved
caspase-3 and Sirt3/SOD2 apoptotic pathways in subarachnoid
hemorrhage mice. Biomed Pharmacother. 109:726–733. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Holschneider CH and Berek JS: Ovarian
cancer: Epidemiology, biology, and prognostic factors. Semin Surg
Oncol. 19:3–10. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Maringe C, Walters S, Butler J, Coleman
MP, Hacker N, Hanna L, Mosgaard BJ, Nordin A, Rosen B, Engholm G,
et al ICBP Module 1 Working Group, : Stage at diagnosis and ovarian
cancer survival: Evidence from the International Cancer
Benchmarking Partnership. Gynecol Oncol. 127:75–82. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu B, Xu T, Liu H, Min Q, Wang S and Song
Q: MiR-490-5p suppresses cell proliferation and invasion by
targeting BUB1 in hepatocellular carcinoma cells. Pharmacology.
100:269–282. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tang D, Zhao X, Zhang L, Wang Z and Wang
C: Identification of hub genes to regulate breast cancer metastasis
to brain by bioinformatics analyses. J Cell Biochem. 120:9522–9531.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pabla S, Conroy JM, Nesline MK, Glenn ST,
Papanicolau-Sengos A, Burgher B, Hagen J, Giamo V, Andreas J, Lenzo
FL, et al: Proliferative potential and resistance to immune
checkpoint blockade in lung cancer patients. J Immunother Cancer.
7:272019. View Article : Google Scholar : PubMed/NCBI
|