Introduction
Over the past four decades, the prevalence of
obesity has tripled worldwide (1).
As the number of individuals that are overweight and obese
increases, the consumption of low-calorie sweeteners has also
gradually increased each year (2).
Although low-calorie synthetic sweeteners, such as aspartame and
saccharine, may have a beneficial effect on weight loss, several
studies have reported adverse effects (3). Therefore, nature-derived sugar
substitutes with low calorific contents, such as D-psicose, have
been developed.
D-psicose, also known as D-allulose, is produced in
small quantities in various fruits. It is an epimer of fructose
that can be enzymatically produced using D-psicose 3-epimerase
(4). In addition to its
anti-obesity effects, D-psicose has also been found to inhibit
cancer cell proliferation by inducing apoptosis and cell cycle
arrest (5–7). Since the purpose of using low-calorie
sweeteners is to help lose weight, it is likely that individuals
will also be regularly exercising while consuming D-psicose.
However, the effect of the chronic consumption of D-psicose on
exercising muscle tissue remains unclear. A previous in vivo
study reported that the oral administration of D-psicose was well
absorbed and distributed throughout the body; the highest
accumulation was found in the liver and kidneys, followed by the
lungs, spleen and skeletal muscle tissue (8). Skeletal muscles comprise >40% of
body organ systems. The muscles in those who exercise regularly are
highly metabolically active and use the majority of the simple
sugars available. Thus, the use of D-psicose by an obese individual
who regularly exercises may lead to muscle cells being highly
exposed to this natural sugar. Therefore, the effect of D-psicose
on muscle cells under exercise-induced oxidative stress requires
further investigation.
During exercise, the rate of muscle contractile
activity exponentially increases, which leads to overactive
mitochondria in muscle cells and the production of reactive oxygen
species (ROS), such as superoxide free radicals (9). Certain types of free radicals have
been detected in muscle tissue and increased free-radical activity
has been shown to lead to extensive muscle damage. The produced
reactive species can positively or negatively modulate muscle cells
(10). Previous studies that have
used hydrogen peroxide (H2O2) treatment for
C2C12 myogenic cells to mimic exercise-induced alterations in
skeletal muscles have revealed that oxidative stress could damage
proliferating myoblasts (11,12).
Apoptosis is an evolutionarily conserved process
that serves an important role in the musculoskeletal system during
development, homeostasis and disease pathology (13). H2O2 and its
reactive by-products act as potential mediators of apoptosis
induced by diverse stimuli (14).
Numerous signaling pathways have been found to be regulated by
exercise-induced ROS generation, leading to muscle cell remodeling
or death by apoptosis (10,11,13).
Among these, MAPK was demonstrated to have a crucial role in
exercise physiology. Apoptosis is a type of programmed cell death
that occurs in response to unnecessary cell proliferation and
facilitates the elimination of injured cells. The ratio of
endogenous pro- to anti-apoptotic proteins is the major determining
factor of cell fate. The overactivation of proapoptotic proteins in
cells is known to inhibit cell cycle progression and cell
proliferation (15). Notably,
several previous studies have reported that D-psicose upregulated
the expression levels of CDKs, leading to cell cycle arrest and
apoptosis in several cell types (5–7).
The present study aimed to determine the effects of
D-psicose on C2C12 myogenic cells. Low dose
H2O2 was used to mimic the generation of ROS
during exercise and to determine how D-psicose affected skeletal
muscle cells under oxidative stress-induced conditions, such as
exercise. The results revealed that D-psicose, in the presence of
physiological concentrations of H2O2, induced
the generation of ROS, triggered cell cycle arrest and initiated
apoptosis in C2C12 myogenic cells. To the best of our knowledge,
this was the first study to demonstrate the toxic effects of
D-psicose on muscle cells treated with H2O2,
which were suggested to occur via the MAPK signaling pathway.
Materials and methods
Chemicals
D-psicose, H2O2,
N-acetylcysteine (NAC), JNK inhibitor VIII and SB203580 were all
purchased from Sigma-Aldrich (Merck KGaA).
Cell culture and treatment
C2C12 cells were obtained from the Human Sciences
Research Resources Bank (Japan Human Sciences Foundation, Tokyo,
Japan). Cells were cultured in low glucose DMEM (Thermo Fisher
Scientific, Inc.), supplemented with 10% heat-inactivated FBS
(Gibco; Thermo Fisher Scientific, Inc.), and maintained at 37°C in
a humidified atmosphere with 5% CO2. For each
experiment, cells were seeded in a 6-well plate and pretreated with
1, 2 or 5 mM D-psicose for 3 h and then with 100 µM
H2O2 for 2 h, after which the medium was
changed, followed by incubation for 19 h.
MTT assay
An MTT assay was used to evaluate cell viability.
Briefly, cells were seeded into 96-well plates at a density of
3×103 cells/well and incubated overnight. Following the
incubation, 20 µl sterile MTT dye (5 mg/ml) was added to each well
and incubated for another 3 h at 37°C. After removal of the medium,
100 µl DMSO was added to each well and incubated for a further 10
min. The absorbance was measured at a wavelength of 570 nm using a
microplate reader (16). Eight
replicate wells were used for each concentration.
Flow cytometric analysis of
apoptosis
Cells were stained with Annexin V-FITC and propidium
iodide (PI; cat. no. BMS500FI-300; Thermo Fisher Scientific, Inc.)
as previously described (17).
Apoptotic cells were subsequently analyzed using flow cytometry (BD
FACSCanto II flow cytometer and FACSDiva ver. 6.1; BD Biosciences)
according to the manufacturer's protocol.
Colony formation assay
Cells (1×103/well) were seeded into
6-well plates and cultured in the indicated media for 10–15 days.
Subsequently, the media was removed, cells were washed twice in
PBS, fixed with 4% paraformaldehyde for 1 h at room temperature and
stained with crystal violet for 40 min at room temperature. Plates
were then thoroughly washed with water and air-dried.
Cell cycle distribution analysis
Cells were harvested after 24 h of treatment and
centrifuged at 200 × g for 5 min at room temperature. Cell pellets
were suspended in 100 µl PBS prior to being fixed with 75% (v/v)
cold ethanol for 2 h at room temperature and then stained with a PI
solution containing DNase-free RNase A for 30 min at room
temperature in the dark. Analysis was performed using a flow
cytometer according to the manufacturer's protocol.
Measurement of intracellular ROS
levels
Intracellular ROS levels were detected using
dihydroethidium (DHE; Molecular Probes; Thermo Fisher Scientific,
Inc.) using flow cytometry. DHE produces blue fluorescence in the
cytosol until it is oxidized by superoxide to 2-hydroxyethidium,
which then intercalates within cellular DNA and stains the nucleus
a bright fluorescent red. Following incubation with 4 µM DHE for 15
min at 37°C, the cells were washed twice with PBS and the
intracellular ROS levels (%) were analyzed using flow
cytometry.
Measurement of the mitochondrial
membrane potential (MMP)
Following treatment, the cells were collected and
treated with 10 nM tetramethylrhodamine methyl ester perchlorate
(TMRM; Molecular Probes; Thermo Fisher Scientific, Inc.) in 1 ml
FBS (1%) diluted in PBS for 15 min at 37°C. TMRM is a cationic
fluorophore widely used for staining cellular mitochondria and the
mitochondrial matrix. The percentage of cells with potential MMP
loss was analyzed by flow cytometry gated for red TMRM
fluorescence, using an excitation wavelength of 488 nm and an
emission wavelength of 575 nm (18).
Western blotting
Cells were harvested, washed with PBS and total
protein was extracted using RIPA lysis buffer [150 mM NaCl, 1%
Triton X-100 (v/v), 1% sodium deoxycholate, 0.1% SDS, 1 µg/ml each
of aprotinin, pepstatin and leupeptin, 1 mM EGTA, 50 mM Tris–HCI;
pH 7.5] for 20 min on ice. Following a brief sonication, lysates
were centrifuged at 13,000 × g for 10 min at 4°C, the supernatant
was collected and the protein contents in the supernatant were
measured using a Coomassie (Bradford) Protein Assay kit (Thermo
Fisher Scientific, Inc.). An equal amount of protein (10 µg) was
loaded per lane and separated via SDS-PAGE. The separated proteins
were subsequently transferred onto PVDF membranes and blocked with
5% skimmed milk in TBS-Tween 20 (150 mM NaCl, 50 mM Tris, 0.1%
Tween-20; pH 7.5) for 1 h at room temperature. The membrane was
then incubated overnight at 4°C with the following primary
antibodies: Anti-MCL1 apoptosis regulator BCL2 family member
(Mcl-1; cat. no. CST68542; 1:1,000), anti-cleaved caspase-3 (cat.
no. CST9661; 1:1,000), anti-JNK (cat. no. CST9258; 1:1,000),
anti-phosphorylated (p)-JNK (cat. no. CST4668; 1:1,000),
anti-ERK1/2 (cat. no. CST4695; 1:1,000), anti-p-ERK1/2 (cat. no.
CST4370; 1:2,000), anti-p38 (cat. no. CST8690; 1:1,000), anti-p-p38
(cat. no. CST4511; 1:1,000), anti-p-checkpoint kinase 1 (Chk1; cat.
no. CST2348; 1:1,000), anti-p-DNA polymerase δ 1, catalytic subunit
(CDC2; cat. no. CST4539; 1:1,000), anti-sirtuin 3 (SIRT3; cat. no.
CST5490; 1:1,000) (all Cell Signaling Technology, Inc.), anti-Bax
(cat. no. ab32503; 1:5,000), anti-cleaved-poly(ADP-ribose)
polymerase 1 (PARP1; cat. no. ab32064; 1:1,000; both Abcam),
anti-cell division cycle 25 C (CDC25C; cat. no. sc327; 1:1,000),
anti-superoxide dismutase 2 (SOD2; cat. no. sc30080; 1:1,000),
anti-Bcl-2 (cat. no. sc7382; 1:400) and anti-β-actin (cat. no.
sc8432; 1:1,000; all Santa Cruz Biotechnology, Inc.). Following the
primary antibody incubation, the membranes were incubated for 2 h
at room temperature with the following secondary antibodies:
Anti-rabbit IgG, HRP-linked Antibody (cat. no. CST7074; 1:3,000)
and anti-mouse IgG, HRP-linked Antibody (cat. no. CST7076; 1:2,000;
both Cell Signaling Technology, Inc.). Protein bands were
visualized using ECL (Amersham; Cytiva) according to the
manufacturer's protocol (19).
Densitometric analysis was performed using ImageJ software 1.50i
(National Institutes of Health).
Statistical analysis
All experiments are representative of three
independent biological replicates. Data are presented as the mean ±
SD. Multiple comparisons were performed using a one-way or two-way
ANOVA with a Tukey's post hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
D-psicose treatment exerts no
cytotoxic effects on resting muscle cells
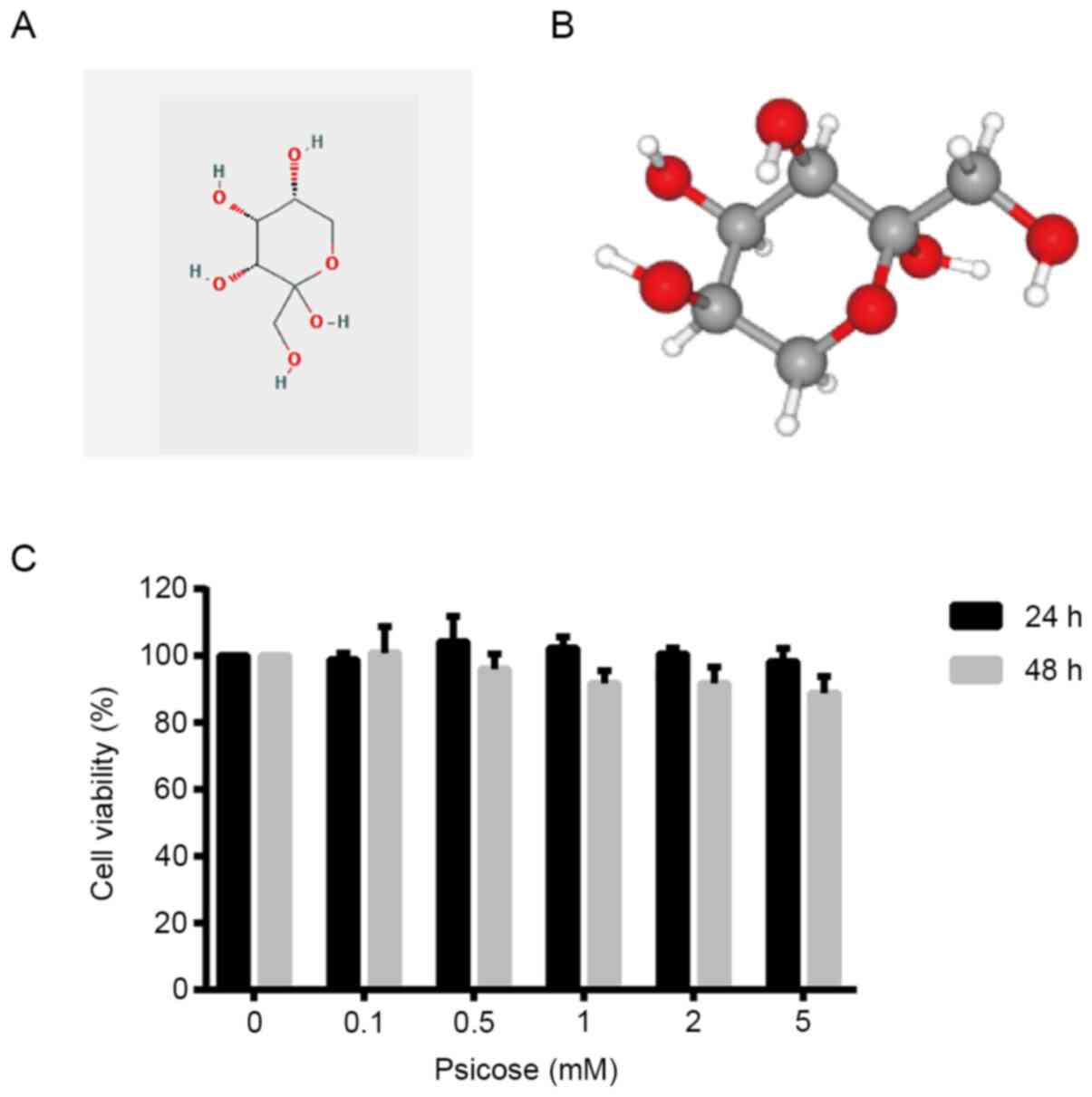
D-psicose is a ketohexose monosaccharide with the
following molecular formula:
C6H12O6. It is a C3 epimer of
fructose that was first derived from the antibiotic psicofuranine,
which contains one ketone group that acts as a reducing agent
(4). The three-dimensional
structure produces a white, odorless powder that is soluble in
water (Fig. 1A and B). The present
study first investigated the effect of D-psicose on C2C12 myogenic
cells in resting conditions (without oxidative stress). The results
demonstrated that increasing concentrations of D-psicose (0–5 mM)
exerted no cytotoxic effects on resting C2C12 cells at 24 or 48 h
post-treatment (Fig. 1C), which
suggested the potential safe application of D-psicose as a sugar
replacement in daily food products.
D-psicose exacerbates
H2O2-induced cell damage in C2C12 cells
To mimic exercise-induced oxidative stress in muscle
cells, C2C12 cells were treated with H2O2.
The results revealed that as the concentration of
H2O2 increased, the viability of C2C12 cells
decreased; ~10% of the cells had decreased viability following 100
µM H2O2 treatment and only 60% of cells
remained viable following 500 µM H2O2
treatment (Fig. 2A). Thus, 100 µM
H2O2 was selected as the optimal
concentration to act as an exercise mimetic in further
experiments.
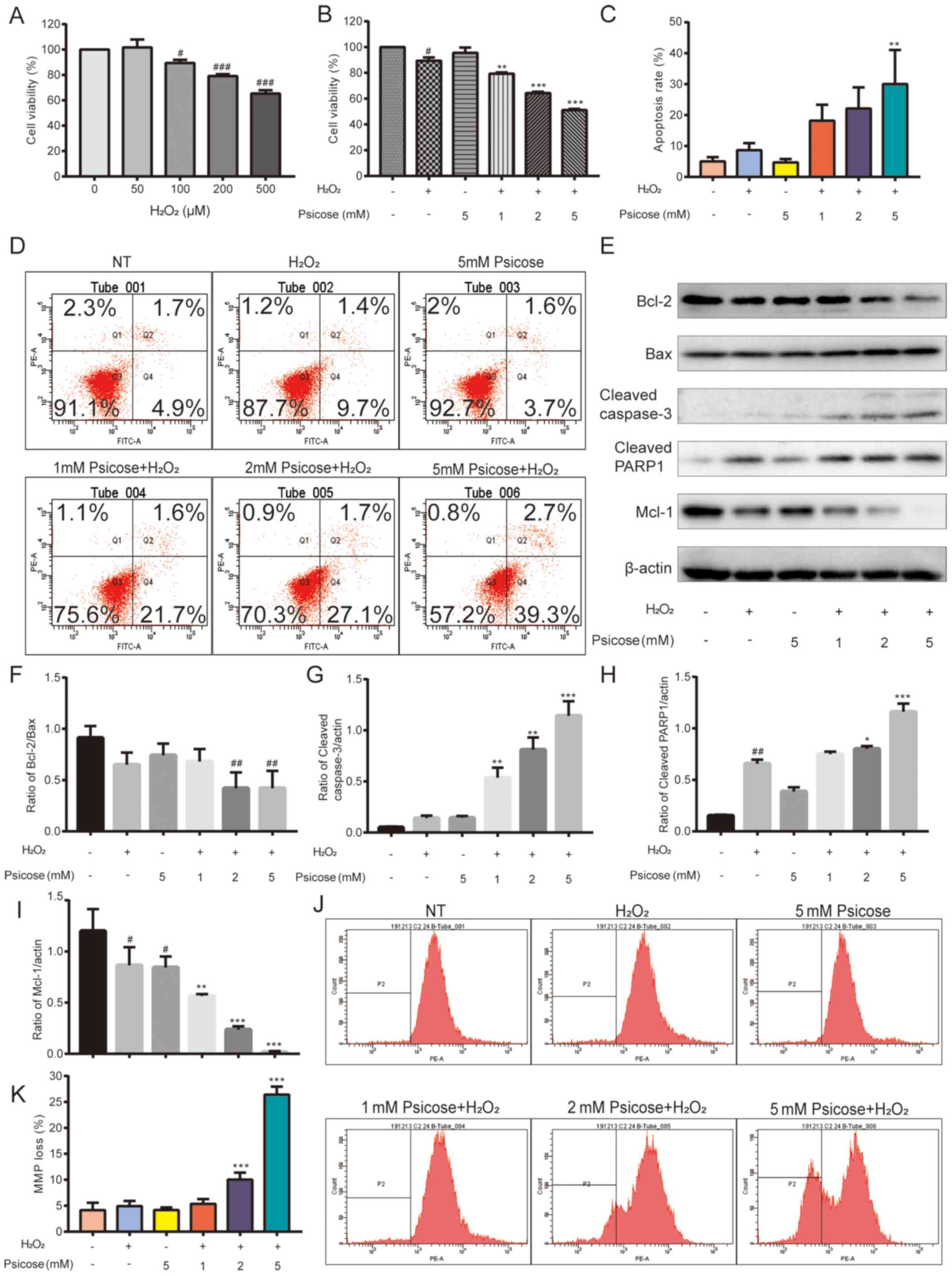 | Figure 2.D-psicose enhances
H2O2-induced C2C12 cell damage. (A) C2C12
cells were pretreated with 0–500 µM H2O2 for
2 h, after which the medium was changed. Following incubation for
24 h, a MTT assay was used to analyze cell viability. The
concentration of 100 µM H2O2 was determined
to be a moderate inducer of oxidative stress. C2C12 cells were
pretreated with 1, 2 or 5 mM D-psicose for 3 h and then with 100 µM
H2O2 for 2 h, after which the medium was
changed. Following incubation for 19 h, (B) an MTT assay, (C)
Annexin V/propidium iodide staining, (D) flow cytometry, (J and K)
tetramethylrhodamine methyl ester perchlorate staining and (E-I)
western blotting were used to analyze cell viability, cell
apoptosis, MMP loss and the expression levels of apoptosis-related
proteins. Data are presented as the mean ± SEM of independent
experiments. #P<0.05, ##P<0.01 and
###P<0.001 vs. NT; *P<0.05, **P<0.01 and
***P<0.001 vs. H2O2 (n=3).
H2O2, hydrogen peroxide; MMP, mitochondrial
membrane potential; Mcl-1, MCL1 apoptosis regulator BCL2 family
member; PARP-1, anti-cleaved-poly(ADP-ribose) polymerase 1; NT, No
treatment. |
C2C12 cells were pre-incubated with D-psicose for 3
h and then treated with H2O2 for 2 h, after
which the cell medium was replaced with fresh cell medium, and
cells were incubated for a further 19 h. Cell viability analysis
revealed a dose-dependent decrease in cell viability in cells
treated with D-psicose in the presence of
H2O2. Notably, viability was lost in >50%
of cells following 5 mM D-psicose treatment (Fig. 2B).
Subsequently, whether H2O2
induced apoptosis in D-psicose pretreated cells was determined
(Fig. 2C and D). To investigate the
effect of D-psicose on the apoptotic pathway, expression levels of
proteins associated with apoptosis were analyzed using western
blotting. D-psicose was found to potentiate
H2O2-induced apoptosis by downregulating the
expression levels of the anti-apoptotic proteins, Bcl-2 and Mcl-1,
and upregulating the expression levels of the proapoptotic protein
Bax (Fig. 2E-I). A concomitant
increase in the cleavage of caspase-3 and PARP1 was also observed,
suggesting that D-psicose initiated caspase-mediated apoptosis.
When cells are under immense oxidative stress, mitochondria lose
their membrane integrity and exercising muscles rely on
mitochondria for the majority of their energy production. Thus, the
effects of D-psicose on mitochondrial function were investigated.
The treatment with H2O2 or D-psicose alone
did not affect the mitochondria of muscle cells; however, in the
presence of D-psicose pretreatment, H2O2
treatment significant decreased the MMP in C2C12 cells (Fig. 2J and K).
D-psicose exacerbates
H2O2-induced apoptosis in C2C12 cells through
the MAPK signaling pathway
Exercising is an intermittent form of cellular
stress that was discovered to activate proteins of the MAPK
signaling pathways in the skeletal muscle of rats (20). In addition, the
H2O2-induced upregulation of ERK, p38 and JNK
expression was reported to regulate cell death in C2C12 cells
(21). To investigate the role of
MAPKs in the damaging effect of D-psicose in exercising skeletal
muscle cells, the protein expression levels of ERK, p38 and JNK
were analyzed in H2O2-stimulated C2C12 cells.
The expression levels of p-JNK and p-p38 were significantly
upregulated following the pretreatment with increasing
concentrations of D-psicose in H2O2-treated
C2C12 cells; however, the expression levels of p-ERK were
downregulated (Fig. 3A-D). These
data suggested that JNK and p38 may have a role in the regulation
of D-psicose-induced apoptosis in muscle cells under oxidative
stress conditions.
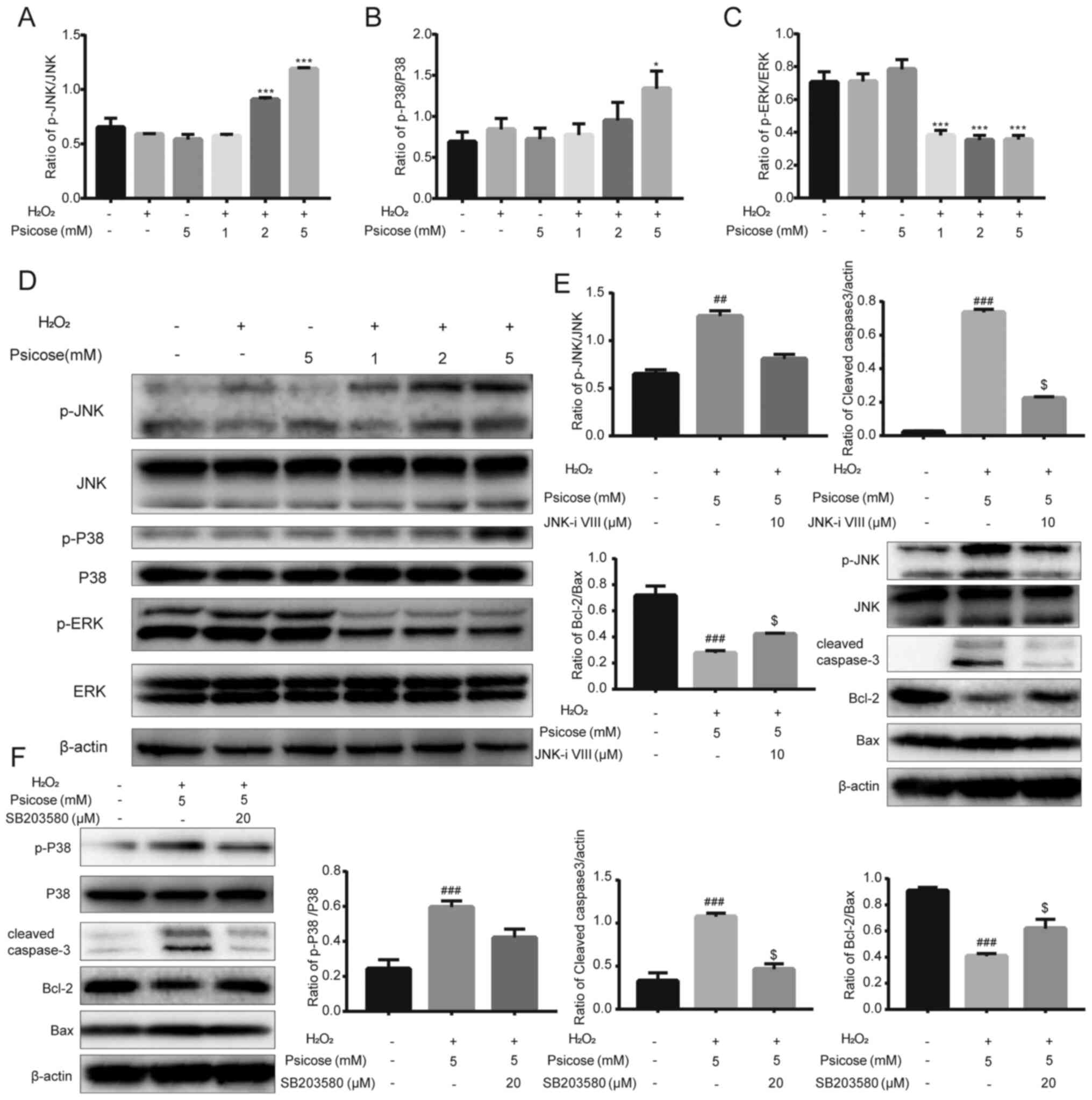 | Figure 3.D-psicose enhances
H2O2-induced apoptosis of C2C12 cells through
the MAPK signaling pathway. C2C12 cells were pretreated with 1, 2
or 5 mM D-psicose for 3 h and then 100 µM
H2O2 for 2 h, after which the medium was
changed. Following incubation for 3 h, protein expression levels of
(A) p-JNK and JNK, (B) p-P38 and P38, and (C) p-ERK and ERK were
analyzed using western blotting. (D) Representative western
blotting image of p-JNK, JNK, p-P38, P38, p-ERK and ERK. (E) C2C12
cells were pretreated with 5 mM D-psicose + 100 µM
H2O2 or 5 mM D-psicose + 10 µM JNK inhibitor
VIII + 100 µM H2O2 for 1 h. The expression
levels of JNK, p-JNK, cleaved caspase-3, Bcl-2 and Bax were
analyzed using western blotting. (F) C2C12 cells were pretreated
with 5 mM D-psicose + 100 µM H2O2 or 5 mM
D-psicose + 20 µM SB203580 + 100 µM H2O2 for
1 h. The expression levels of P38, p-P38, cleaved caspase-3, Bcl-2
and Bax were analyzed using western blotting. β-actin was used as
the loading control. Data are presented as the mean ± SEM of
independent experiments. ##P<0.01 and
###P<0.001 vs. NT; *P<0.05 and ***P<0.001 vs.
H2O2; $P<0.05 vs. D-psicose +
H2O2 (n=3). H2O2,
hydrogen peroxide; p-, phosphorylated. |
To validate these findings, muscle cells were
exposed to JNK and p-38 inhibitors (JNK inhibitor VIII and
SB203580, respectively). Following the pre-incubation of the C2C12
cell culture with JNK inhibitor VIII or SB203580, followed by
treatment with D-psicose and H2O2, the
expression levels of p-JNK and p38 were downregulated.
Concurrently, both of the inhibitors also reversed
D-psicose-induced apoptotic effects by upregulating Bcl-2
expression levels and inhibiting the cleavage of caspase-3
(Fig. 3E and F).
D-psicose pretreatment enhances
H2O2-induced oxidative stress via ROS
generation
The generation of ROS and activation of
apoptosis-related proteins serves a key role in triggering
apoptosis. Previously, it was reported that
H2O2 induced high levels of oxidative stress
and apoptosis in proliferating skeletal myoblasts (22). Hence, the present study sought to
determine whether the same was true for the effect of
D-psicose-induced apoptosis in C2C12 cells. The results revealed
that pretreatment with increasing concentrations of D-psicose
increased the production of ROS in
H2O2-treated C2C12 cells, while this effect
was significantly reversed upon exposure to NAC (Fig. 4A and B). In addition, MAPK signaling
molecules were observed to be activated by D-psicose, while the
effects of H2O2 treatment were markedly
blocked by NAC. Furthermore, the phosphorylation levels of JNK and
p38 were reduced, while the phosphorylation levels of ERK were
increased. Concurrently, the levels of apoptosis were inhibited,
which was evident by the upregulated expression levels of Bcl-2,
and downregulated expression levels of Bax and cleaved caspase-3
(Fig. 4C). As downregulated
expression levels of SIRT3 were previously reported to promote the
generation of ROS and inhibit the SIRT3/SOD2-mediated deactivation
process of superoxide radicals (23), the present study also investigated
the effect of D-psicose and H2O2 treatment on
the expression levels of SIRT3 and SOD2 in C2C12 cells. The results
revealed that the pretreatment with D-psicose significantly
downregulated the expression levels of SIRT3 and SOD2 in
H2O2-treated C2C12 cells in a dose-dependent
manner (Fig. 4D).
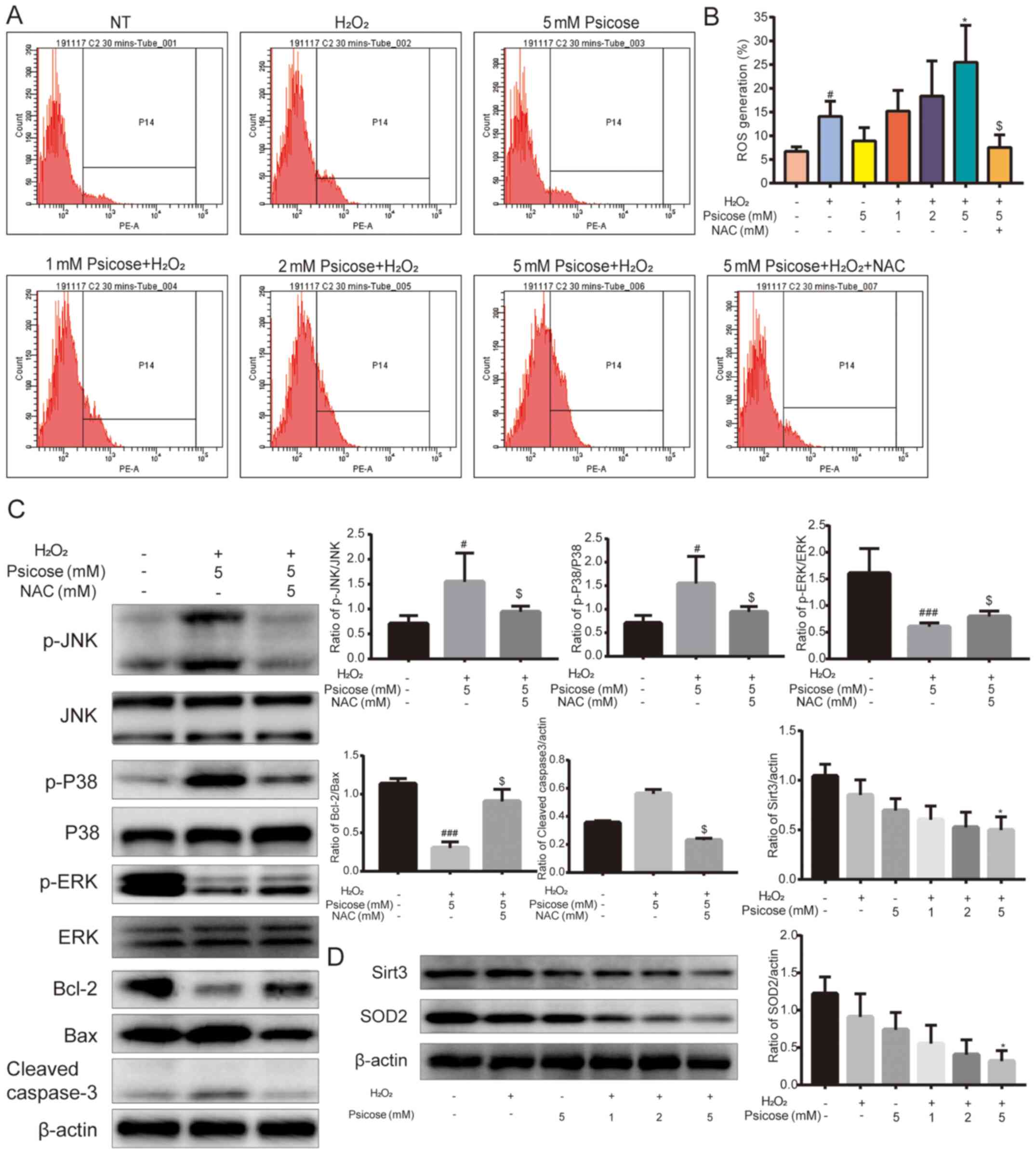 | Figure 4.Co-treatment of D-psicose and
H2O2 increases ROS generation in C2C12 cells.
C2C12 cells were pretreated with 1, 2 or 5 mM D-psicose for 3 h and
then 100 µM H2O2 for 2 h, after which the
medium was changed. Following incubation for 30 min, ROS generation
was analyzed using dihydroethidium staining and flow cytometry (A)
Representative flow cytometry plots and (B) quantification of ROS
generation. (C) C2C12 cells were pretreated with 5 mM D-psicose +
100 µM H2O2 or 5 mM D-psicose + 5 mM NAC +
100 µM H2O2 for 1 h. The expression levels of
p-p38, p-JNK, p-ERK, cleaved caspase-3, Bcl-2 and Bax were analyzed
using western blotting. (D) C2C12 cells were pretreated with 1, 2
or 5 mM D-psicose for 3 h and with 100 µM
H2O2 for 2 h, after which the medium was
changed. Following incubation for 3 h, the expression levels of
SIRT3 and SOD2 were analyzed using western blotting. Data are
presented as the mean ± SEM of independent experiments.
#P<0.05 and ###P<0.001 vs. NT;
*P<0.05 vs. H2O2; $P<0.05
vs. D-psicose + H2O2 (n=3).
H2O2, hydrogen peroxide; ROS, reactive oxygen
species; p-, phosphorylated; SIRT3, sirtuin 3; SOD2, superoxide
dismutase 3; NAC, N-acetylcysteine; NT, No treatment. |
D-psicose and
H2O2 induce cell cycle arrest and inhibit the
proliferation of C2C12 cells
The effect of D-psicose on muscle cell proliferation
under conditions of oxidative stress were subsequently
investigated. The combined treatment with D-psicose and
H2O2 increased the percentage of cells in the
sub-G1 phase of the cell cycle compared with cells in
other phases of the cell cycle (Fig. 5A
and B). This finding suggested the potential inhibitory effect
of D-psicose on cell viability and proliferation. Furthermore, a
colony formation assay was performed for 5 days post-treatment to
determine the long-term effects of D-psicose and
H2O2 treatment on muscle cell proliferation.
Both the number and size of colonies were decreased in the
treatment groups, while few colonies formed from
H2O2-stimulated C2C12 cells pretreated with 5
mM D-psicose (Fig. 5C). In
addition, the expression levels of several proteins associated with
the cell cycle were analyzed using western blotting. The results
revealed that, in C2C12 cells, D-psicose treatment in the presence
of H2O2 downregulated the expression levels
of CDC25C and the phosphorylation of CDC2 (Fig. 5D-G). These findings indicated that
D-psicose may potentiate cell cycle arrest and decrease the
long-term survival of muscle cells under oxidative stress
conditions.
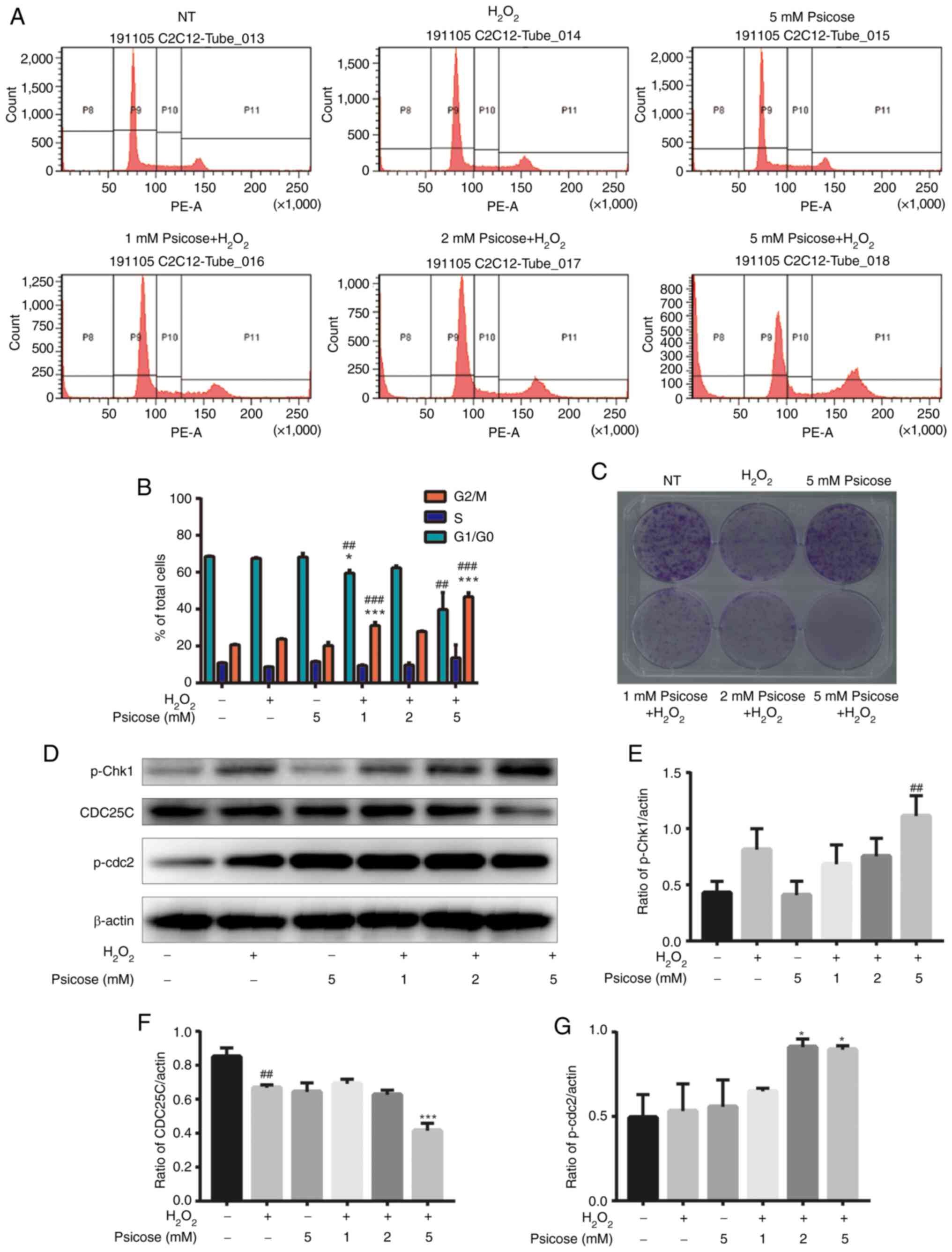 | Figure 5.Co-treatment with D-psicose and
H2O2 induces G2/M phase cell cycle
arrest in C2C12 myoblasts. C2C12 cells were pretreated with 1, 2 or
5 mM D-psicose for 3 h and then with 100 µM
H2O2 for 2 h, after which the medium was
changed. Following incubation for 24 h, the SubG1 cell
population was analyzed using propidium iodide staining and flow
cytometry. (A) Representative flow cytometry plots and (B)
quantification of cells in each state of the cell cycle. (C) Colony
formation assays were performed using cells seeded at a density of
1×103 cells/well into 6-well plates for 24 h, treated as
described in part (A) and incubated for 5 days. (D) C2C12 cells
were pretreated with 1, 2 or 5 mM D-psicose for 3 h and then with
100 μM H2O2 for 2 h, after which the medium was changed. Following
incubation for 3 h, the expression levels of p-Chk1, CDC25C and
p-CDC2 were analyzed using western blotting. The relative
expression levels of (E) p-Chk1, (F) CDC25C and (G) p-CDC2 were
quantified relative to the value of β-actin. Data are presented as
the mean ± SEM of independent experiments. ##P<0.01
and ###P<0.001 vs. NT; *P<0.05 and ***P<0.001
vs. H2O2 (n=3). H2O2,
hydrogen peroxide; p-, phosphorylated; Chk1, checkpoint kinase 1;
CDC25C, cell division cycle 25 C; CDC2, DNA polymerase δ 1,
catalytic subunit; NT, No treatment. |
Discussion
The present study aimed to determine the effect of
D-psicose on ROS generation and H2O2-induced
apoptosis in C2C12 myogenic cells. H2O2 is a
widely used method to establish exercise-induced oxidative stress
in cells (11,12,22).
The present results revealed that the treatment of C2C12 cells with
D-psicose under oxidative stress conditions elevated the levels of
ROS, decreased the MMP and apoptosis, and induced cell cycle
arrest. Moreover, the lethal effects of D-psicose on exercising
muscle cells were found to be mediated via the MAPK family
molecules, p-JNK and p-p38 (Fig.
6). Notably, the observed effects were reversed following
treatment with the ROS inhibitor, NAC, which is consistent with the
findings of a previous study (24).
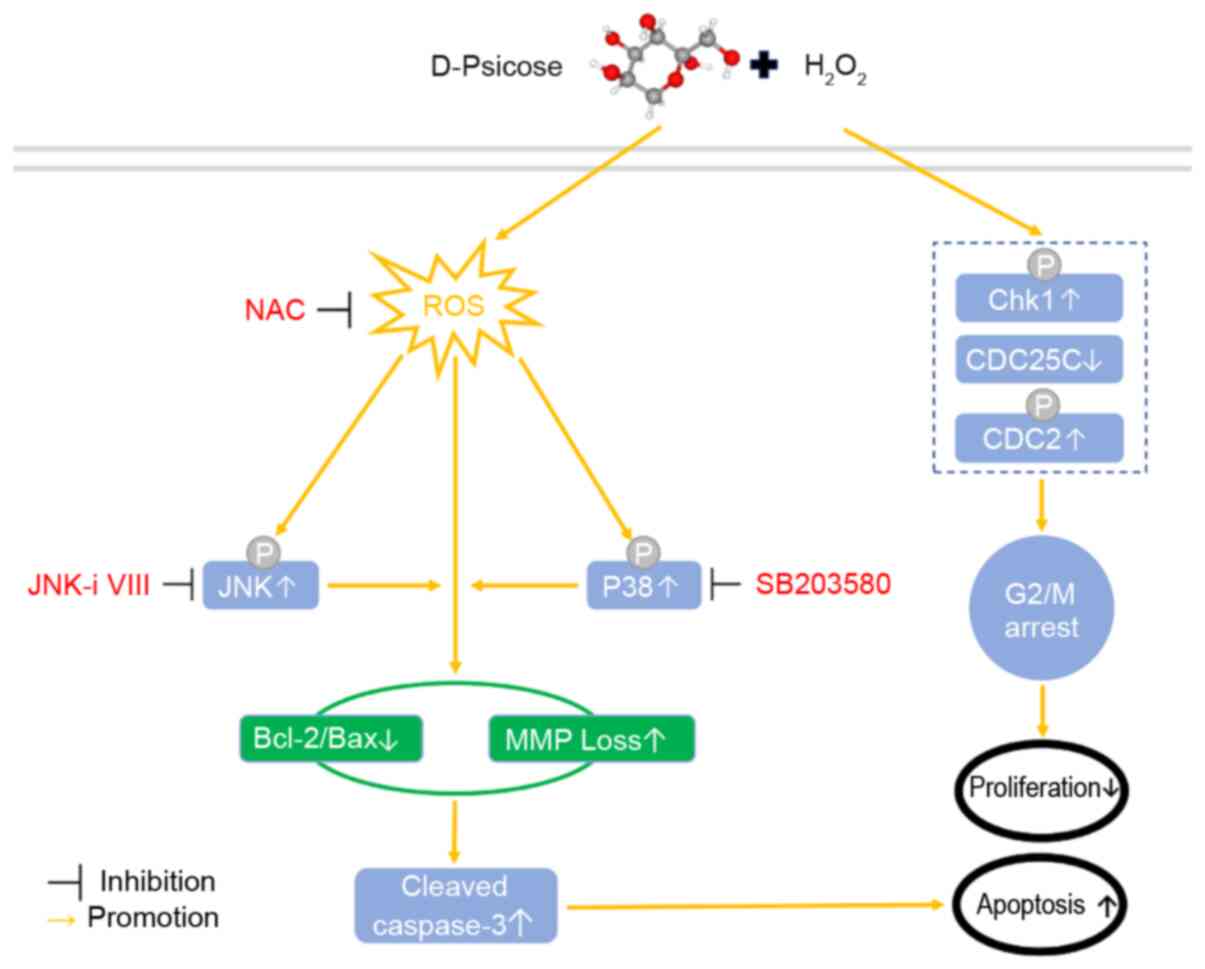 | Figure 6.Proposed molecular mechanism through
which D-psicose and H2O2 may induce apoptosis
and G2/M cell cycle arrest. H2O2,
hydrogen peroxide; CDC25C, cell division cycle 25 C; CDC2, DNA
polymerase δ 1, catalytic subunit; Chk1, checkpoint kinase 1;
H2O2, hydrogen peroxide; MMP, membrane
mitochondrial potential; ROS, reactive oxygen species; NAC,
N-acetylcysteine; p-, phosphorylated. |
Oxidative stress is the production of excessive
reactive oxidants that exceeds the cellular antioxidant capability.
In 1992, it was reported that contracting rodent skeletal muscle
released superoxide radicals into the interstitial spaces (25). Bailey et al (26) also demonstrated that exercise
resulted in a greater production and accumulation of intramuscular
free radicals in human skeletal muscle cells. ROS produced in
contracting skeletal muscle facilitates the strength of the muscle
contraction; however, if ROS concentrations exceed a certain level,
it reduces force generation and causes fatigue (27). The findings of the present study
revealed that low doses of H2O2 induced
oxidative stress in muscle cells, which may be due to
exercise-induced ROS generation. However, the concomitant exposure
to D-psicose further increased the production of ROS, leading to
cell death. On the other hand, pretreatment with NAC reversed the
increased the production of ROS, returning the levels of ROS to
almost homeostatic levels, thereby providing a protective effect.
These results were consistent with previous studies, which showed
that antioxidant infusion in humans during voluntary exercise
substantially enhanced performance during prolonged exercise
(28,29).
The balance between antioxidants and pro-oxidants is
crucial for the survival of aerobic organisms. A previous
experiment in rats demonstrated that exercise promoted the
expression and activity of SOD2 in skeletal muscle mitochondria,
which subsequently maintained cellular oxidant-antioxidant
homeostasis (30). Therefore, as
the exercise-induced upregulation of SOD2 in skeletal muscle served
a key role in the prevention of superoxide accumulation, it could
be hypothesized that the loss of SOD2 may have devastating effects
on muscle cell survival. SIRT3 was discovered to regulate SOD2 via
deacetylation and activation of FOXO3a, and SOD2 can catalyze the
neutralization process involving the conversion of superoxide into
water and hydrogen peroxide (23,31).
The results of the present study revealed that the treatment with
D-psicose alone, in addition to in the presence of
H2O2, exerted inhibitory effects on the
expression levels of SIRT3 and SOD2.
D-psicose was found to suppress post-prandial serum
glucose levels and reduce the accumulation of body fat, which
indicates its beneficial effects on obesity-related metabolic
disturbances (8). In the past, this
type of sugar supplement was used in tabletop packets in cafés and
restaurants. Currently, these sugars are added to everyday food
products to promote the sales of food items as healthy
alternatives. In both children and adults, the consumption of
low-calorie sweeteners has significantly increased, primarily due
to an increased awareness of their non-nutritive value and
potential effects in weight loss (32). Hence, future studies should aim to
enhance the existing knowledge on the metabolic and health effects
of low-calorie sweeteners. A long-term study on D-psicose toxicity
in rats revealed that the final body weight and weight gain in
D-psicose-fed animals were significantly decreased compared with
the control group (33). On the one
hand, these findings may be considered as beneficial for weight
loss; however, D-psicose may be reducing the body weight by
decreasing the number of muscle cells. Yagi and Matsuo (33) investigated the long-term (18 months)
toxicity of D-psicose in rats and found a significant increase in
organ weight of the liver, kidneys, brain, lungs and pancreas,
which suggested that the accumulation of fat, and that muscle
proteins may be broken down to promote toxic effects. The present
data suggested that the treatment with D-psicose was safe for
muscle cells; however, under stressful conditions, D-psicose could
exert deleterious effects on the survival of the muscle cells.
Previous studies have shown that D-psicose increased
G2/M phase cell cycle arrest and decreased the
population of liver cells in the S phase (5). Cell cycle progression is tightly
regulated by a complex network of positive and negative cell cycle
regulatory molecules, such as cyclins and CDKs. CDC25C is involved
in the DNA damage checkpoints by activating CDK complexes that
drive the cell cycle and is a key mediator of cell cycle
progression (34). The current
findings demonstrated that the treatment with D-psicose in
H2O2-stimulated muscle cells led to the
accumulation of cells in the sub-G1 phase and the
downregulation of the expression levels of the cell
cycle-regulating protein, CDC25C.
In conclusion, the findings of the present study
suggested that under physiological oxidative stress conditions,
D-psicose may exert negative effects on muscle cells by promoting
the production of ROS and triggering cell cycle arrest and
apoptosis. The results further indicated that the negative effects
of D-psicose on exercising muscle cells may be mediated by MAPK and
may be partly due to slow cell cycle progression and the failure to
repair DNA damage, which ultimately leads to apoptosis.
Acknowledgements
Not applicable.
Funding
This work was supported by the National Natural
Science Foundation of China (grant no. 81330043) and Beijing
Municipal Health Commission (grant nos. BMHC-2019-9 and
BMHC-2018-4).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ZW and LS designed the study. ZW and YL performed
the experiments and collected the data. ZC and CW confirm the
authenticity of all the raw data. ZW interpreted the data with help
from LS, YL, JM, YW, QF, YZ, HI, ZC and CW. ZW, LS and YL analyzed
the results. ZW and JM wrote the manuscript with help from ZC and
CW. ZC and CW were responsible for funding acquisition. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chu DT, Minh Nguyet NT, Dinh TC, Thai Lien
NV, Nguyen KH, Nhu Ngoc VT, Tao Y, Son LH, Le DH, Nga VB, et al: An
update on physical health and economic consequences of overweight
and obesity. Diabetes Metab Syndr. 12:1095–1100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sylvetsky AC and Rother KI: Trends in the
consumption of low-calorie sweeteners. Physiol Behav. 164:446–450.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Swithers SE: Artificial sweeteners produce
the counterintuitive effect of inducing metabolic derangements.
Trends Endocrinol Metab. 24:431–441. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
He W, Mu W, Jiang B, Yan X and Zhang T:
Construction of a food grade recombinant bacillus subtilis based on
replicative plasmids with an auxotrophic marker for
biotransformation of d-Fructose to d-Allulose. J Agric Food Chem.
64:3243–3250. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sui L, Dong Y, Watanabe Y, Yamaguchi F,
Hatano N, Tsukamoto I, Izumori K and Tokuda M: The inhibitory
effect and possible mechanisms of D-allose on cancer cell
proliferation. Int J Oncol. 27:907–912. 2005.PubMed/NCBI
|
|
6
|
Sui L, Dong Y, Watanabe Y, Yamaguchi F,
Hatano N, Izumori K and Tokuda M: Growth inhibitory effect of
D-allose on human ovarian carcinoma cells in vitro. Anticancer Res.
25:2639–2644. 2005.PubMed/NCBI
|
|
7
|
Mitani T, Hoshikawa H, Mori T, Hosokawa T,
Tsukamoto I, Yamaguchi F, Kamitori K, Tokuda M and Mori N: Growth
inhibition of head and neck carcinomas by D-allose. Head Neck.
31:1049–1055. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Tsukamoto I, Hossain A, Yamaguchi F,
Hirata Y, Dong Y, Kamitori K, Sui L, Nonaka M, Ueno M, Nishimoto K,
et al: Intestinal absorption, organ distribution, and urinary
excretion of the rare sugar D-psicose. Drug Des Devel Ther.
8:1955–1964. 2014.PubMed/NCBI
|
|
9
|
Qaisar R, Bhaskaran S and Van Remmen H:
Muscle fiber type diversification during exercise and regeneration.
Free Radic Biol Med. 98:56–67. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Derbre F, Ferrando B, Gomez-Cabrera MC,
Sanchis-Gomar F, Martinez-Bello VE, Olaso-Gonzalez G, Diaz A,
Gratas-Delamarche A, Cerda M and Viña J: Inhibition of xanthine
oxidase by allopurinol prevents skeletal muscle atrophy: role of
p38 MAPKinase and E3 ubiquitin ligases. PLoS One. 7:e466682012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fan X, Hussien R and Brooks GA:
H2O2-induced mitochondrial fragmentation in C2C12 myocytes. Free
Radic Biol Med. 49:1646–1654. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Siu PM, Wang Y and Alway SE: Apoptotic
signaling induced by H2O2-mediated oxidative stress in
differentiated C2C12 myotubes. Life Sci. 84:468–481. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sandri M and Carraro U: Apoptosis of
skeletal muscles during development and disease. Int J Biochem Cell
Biol. 31:1373–1390. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jackson MJ, Edwards RH and Symons MC:
Electron spin resonance studies of intact mammalian skeletal
muscle. Biochim Biophys Acta. 847:185–190. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Goldar S, Khaniani MS, Derakhshan SM and
Baradaran B: Molecular mechanisms of apoptosis and roles in cancer
development and treatment. Asian Pac J Cancer Prev. 16:2129–2144.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zakki SA, Muhammad JS, Li JL, Sun L, Li
ML, Feng QW, Li YL, Cui ZG and Inadera H: Melatonin triggers the
anticancer potential of phenylarsine oxide via induction of
apoptosis through ROS generation and JNK activation. Metallomics.
12:396–407. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hertveldt K, Philippe J, Thierens H,
Cornelissen M, Vral A and De Ridder L: Flow cytometry as a
quantitative and sensitive method to evaluate low dose radiation
induced apoptosis in vitro in human peripheral blood lymphocytes.
Int J Radiat Biol. 71:429–433. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu J, Liu T, Rios Z, Mei Q, Lin X and Cao
S: Heat shock proteins and cancer. Trends Pharmacol Sci.
38:226–256. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li M, Kondo T, Zhao QL, Li FJ, Tanabe K,
Arai Y, Zhou ZC and Kasuya M: Apoptosis induced by cadmium in human
lymphoma U937 cells through Ca2+-calpain and
caspase-mitochondria-dependent pathways. J Biol Chem.
275:39702–39709. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Goodyear LJ, Chang PY, Sherwood DJ,
Dufresne SD and Moller DE: Effects of exercise and insulin on
mitogen-activated protein kinase signaling pathways in rat skeletal
muscle. Am J Physiol. 271:E403–E408. 1996.PubMed/NCBI
|
|
21
|
Ronda AC, Vasconsuelo A and Boland R:
Extracellular-regulated kinase and p38 mitogen-activated protein
kinases are involved in the antiapoptotic action of
17beta-estradiol in skeletal muscle cells. J Endocrinol.
206:235–246. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Caporossi D, Ciafre SA, Pittaluga M,
Savini I and Farace MG: Cellular responses to H(2)O(2) and
bleomycin-induced oxidative stress in L6C5 rat myoblasts. Free
Radic Biol Med. 35:1355–1364. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lu J, Cheng K, Zhang B, Xu H, Cao Y, Guo
F, Feng X and Xia Q: Novel mechanisms for superoxide-scavenging
activity of human manganese superoxide dismutase determined by the
K68 key acetylation site. Free Radic Biol Med. 85:114–126. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Coffey VG, Zhong Z, Shield A, Canny BJ,
Chibalin AV, Zierath JR and Hawley JA: Early signaling responses to
divergent exercise stimuli in skeletal muscle from well-trained
humans. FASEB J. 20:190–192. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Reid MB, Shoji T, Moody MR and Entman ML:
Reactive oxygen in skeletal muscle. II. Extracellular release of
free radicals. J Appl Physiol (1985). 73:1805–1809. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bailey DM, Lawrenson L, McEneny J, Young
IS, James PE, Jackson SK, Henry RR, Mathieu-Costello O, McCord JM
and Richardson RS: Electron paramagnetic spectroscopic evidence of
exercise-induced free radical accumulation in human skeletal
muscle. Free Radic Res. 41:182–190. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nemes R, Koltai E, Taylor AW, Suzuki K,
Gyori F and Radak Z: Reactive oxygen and nitrogen species regulate
key metabolic, anabolic, and catabolic pathways in skeletal muscle.
Antioxidants (Basel). 7:852018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Medved I, Brown MJ, Bjorksten AR and
McKenna MJ: Effects of intravenous N-acetylcysteine infusion on
time to fatigue and potassium regulation during prolonged cycling
exercise. J Appl Physiol (1985). 96:211–217. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
McKenna MJ, Medved I, Goodman CA, Brown
MJ, Bjorksten AR, Murphy KT, Petersen AC, Sostaric S and Gong X:
N-acetylcysteine attenuates the decline in muscle Na+,K+-pump
activity and delays fatigue during prolonged exercise in humans. J
Physiol. 576:279–288. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hollander J, Fiebig R, Gore M, Ookawara T,
Ohno H and Ji LL: Superoxide dismutase gene expression is activated
by a single bout of exercise in rat skeletal muscle. Pflugers Arch.
442:426–434. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim YS, Gupta Vallur P, Phaeton R,
Mythreye K and Hempel N: Insights into the Dichotomous Regulation
of SOD2 in Cancer. Antioxidants (Basel). 6:862017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Magnuson BA, Carakostas MC, Moore NH,
Poulos SP and Renwick AG: Biological fate of low-calorie
sweeteners. Nutr Rev. 74:670–689. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yagi K and Matsuo T: The study on
long-term toxicity of d-psicose in rats. J Clin Biochem Nutr.
45:271–277. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cho YC, Park JE, Park BC, Kim JH, Jeong
DG, Park SG and Cho S: Cell cycle-dependent Cdc25C phosphatase
determines cell survival by regulating apoptosis signal-regulating
kinase 1. Cell Death Differ. 22:1605–1617. 2015. View Article : Google Scholar : PubMed/NCBI
|