Introduction
Stroke is the second leading cause of mortality
worldwide, with an annual mortality of ~5.5 million, and causes
disability in 50% of survivors (1).
Ischemic stroke accounts for 75–85% of all strokes and is a
pathological state characterized by limited blood flow (2). With regards to hindered cerebral blood
supply, the most common clinical treatment for ischemic stroke is
restoration of perfusion and reoxygenation to the affected area
(3). However, reoxygenation and
blood flow recovery are associated with aggravation of tissue
injury and a severe inflammatory response, which is known as
cerebral ischemia-reperfusion (CIR) injury (4,5). Due
to the complex mechanism of cerebral ischemia, clinical treatment
is difficult. Therefore, it is urgent to find effective treatment
for cerebral ischemia.
Dexmedetomidine (DEX) is a novel type of analgesic
(6). Multiple studies have shown
that DEX exerts its role via anti-inflammatory and anti-apoptotic
pathways, and it is used in the treatment of neuropathic pain and
other diseases (7–9). However, the mechanism of the
protective effect of DEX is still unclear. Certain studies have
shown that DEX serves a neuroprotective role by inhibiting
endoplasmic reticulum stress (10,11).
The present study investigated whether DEX inhibits expression of
microRNA (miRNA/miR)-199a and improves neurocyte injury induced by
CIR injury.
Previously, non-coding RNA has attracted significant
attention (12–15). Non-coding RNAs comprise ribosomal,
transfer, small nuclear and long non-coding RNA, as well as miRNA.
miRNAs are endogenous short single-stranded RNAs (12). Previ-ous studies have investigated
the role of miRNAs in cancer, particularly the potential of miRNAs
as specific biomarkers of cancer (13,14).
In addition to cancer, miRNA serves an important role in regulation
of gene expression in various biological pathophysiological
processes (15). In the central
nervous system, pathological conditions, including stroke,
schizophrenia and neurodegenerative disease, significantly alter
expression of miRNAs in the brain and affect the occurrence of
disease (16,17). miRNAs have been reported to be
associated with IR injury, including miR-455 and miR-199a.
Furthermore, miR-199a has been shown to recognize and target
sirtuin 1, thereby repressing hypoxia preconditioning and longevity
of cardiac myocytes (18–20). Therefore, it was hypothesized that
DEX may serve a role in CIR injury by targeting miR-199a.
In the present study it was hypothesized that the
neuroprotection of DEX may be associated with miR-199a in CIR
injury. Thus, the miR-199a antagomir was used to determine the
mechanism of DEX in a rat model of CIR injury.
Materials and methods
Animals
In total, 72 healthy SPF grade Sprague Dawley male
rats (weight, 180–220 g; age, 6–8 weeks) were purchased from Jinan
Pengyue Experimental Animal Co., Ltd. [licence no. SCXK (Lu)
20190003]. All rats were housed under controlled temperature
(22±2°C) and humidity (55±5%), with free access to food and water,
and were subjected to adaptive feeding for 1 week under a 12-h
light/dark cycle. Animal experiments were performed in accordance
with the Guide for the Care and Use of Laboratory Animals, 8th
edition (21) and were approved by
the Animal Protection and Use Committee of Yantaishan Hospital
(Yantai, China).
CIR model
Before the operation, rats were fasted for 12 h and
allowed to drink water; the percentage of maximum body weight loss
was 0.10%. According to the Zea Longa method (22), the CIR model was established using
middle cerebral artery occlusion. Rats were anesthetized via
intraperitoneal injection of sodium pentobarbital (50 mg/kg) and
fixed in a supine position. The common, internal and external
carotid arteries were carefully separated along the midline of the
neck. Next, a nylon thread was inserted into the internal carotid
artery. Upon feeling slight resistance, the insertion of the nylon
thread was halted and fixed in place. After blocking blood flow for
2 h, the thread was removed, and reperfusion was permitted for 24
h. The wound was sutured layer-by-layer after the operation. During
the operation, the room temperature was maintained at 37.0±0.5°C,
and the rectal temperature, respiratory rate and heart rate of
animals were monitored. After animals woke up, gait and behavior
were observed, and the success of modeling was judged according to
the following scoring standard, as previously described (8,22,23):
Grade 0 (normal), symptoms without neurological impairment; grade 1
(mild), inextensibility of the left forepaw when lifting the rat
tail; grade 2 (moderate), circling to the left side while walking;
grade 3 (severe), walking difficulty and leaning to the left; grade
4 (very severe), inability to walk spontaneously. Rats with a score
of 1–3 were selected for subsequent experiments. The thread in rats
in the sham operation group was removed immediately after inserting
the thread plug without retaining the middle cerebral artery.
Intracerebroventricular injection
Following routine anesthesia using sodium
pentobar-bital (50 mg/kg), the rats were fixed on a stable
stereotactic instrument. After skin preparation and disinfection
using 2.5% iodine, the top skin of the head was cut (depth, 1–2
cm), and the anterior fontanel was exposed. Head fixation of rats
was as follows: The nose was in the middle of the fixator; front
and rear fontanelle were kept at the same level; and the head was
firmly fixed. The diameter of the drill hole was ~0.5 mm, and the
bregma coordinates were as follows: AP, 0.8 mm; L, 1.5 mm; and V,
4.5 mm, based on the rat brain stereotaxic atlas (24). The miR-199a antagomir or negative
con-trol was slowly injected into the lateral ventricle at an
injection rate of 0.5 µl/min using a microinjector. Then, the
needle was slowly removed and the small hole was sealed with
medical bone wax. The incision was sutured following local routine
disinfection.
Animal grouping and treatment
The animals were randomly divided into the following
groups: i) Sham; ii) CIR model; iii) DEX; iv) miR-199a antagomir
(anta-miR); v) anta-miR + negative control (NC); and vi) DEX +
anta-miR (n=12/group). In the Sham group, the thread was removed
immediately after inserting the thread plug, without retaining the
middle cerebral artery and rats were fed normally. In the CIR model
group, CIR injury was induced. In the DEX group, 3 µg/kg/h DEX was
injected into the caudal vein 24 h before the operation and during
reperfusion, as previously described (8). At 0.5 h before reperfusion, animals in
the anta-miR group were injected with 5 µl anta-miR (Shanghai
GenaPharma Co., Ltd.; http://www.genepharma.com/en/) in the breg-ma
coordinates AP, 0.8 mm; L, 1.5 mm; and V, 4.5 mm (based on the rat
brain stereo-taxic atlas). In the NC antagomir group (anta-NC), 0.5
h before perfusion, 5 µl anta-miR NC (5′-UAACCAAUGUGCAGACUACUGU-3′;
Shanghai GenaPharma Co., Ltd.), was injected into the right
ventricle at the aforementioned coordinates. In the DEX + anta-miR
group, DEX was administered twice: 3 µg/kg/h DEX was injected into
the tail vein 24 h before the operation and at the beginning of
reperfusion, separately. In addition, 5 h before reperfusion, 5 µl
anta-miR was injected into the right ventricle of rats (bregma
coordinates: AP, 0.8 mm; L, 1.5 mm; and V, 4.5 mm, based on the Rat
Brain Stereotaxic Atlas). Following 24 h reperfusion, the rats were
anesthetized with 3% pentobarbital sodium (45 mg/kg), followed by
euthanasia via cervical dislocation.
The brain tissue of 6 rats in each group was fixed
in 4% paraformaldehyde at 37°C for 24 h for
2,3,5-triphenyltetrazolium chloride (TTC) and hematoxylin and eosin
(H&E) staining, immunofluorescence and immunohistochemistry.
Brain tissue from another 6 rats in each group was stored in liquid
nitrogen for western blotting and reverse
transcription-quantitative PCR (RT-qPCR) analysis.
Improved mNSS
The neurobehavioral score of the experimental groups
was assessed in a blinded manner. The neurological function of rats
was evaluated before modeling and at 6, 12 and 24 h after
reperfusion. The mNSS behavioral score is primarily used to assess
ability, such as movement, sensation, balance and reflex, of rats
(25). The score ranges from 0 to
18. The scoring criteria is showed in Table I. The higher the score, the more
serious the neurological deficit. A total of 8 rats in each group
were randomly selected for mNSS behavioral tests at 1, 3 and 7 days
after the operation. The maximum score was 18 points, with 1–6
points allocated for mild, 7–12 for moderate and 13–18 for severe
damage.
 | Table I.Modified neurological severity score
(maximum points=18). |
Table I.
Modified neurological severity score
(maximum points=18).
| Test | Points |
|---|
| Raising rat by
tail | 3 |
| Flexion
of forelimb | 1 |
| Flexion
of hindlimb | 1 |
| Head
moves >10° to vertical axis within 30 sec | 1 |
| Placing rat on
floor | 3 |
| Normal
walk | 0 |
|
Inability to walk
straight | 1 |
|
Circling toward the paretic
side | 2 |
| Fall
down to the paretic side | 3 |
| Sensory | 2 |
| Placing
(visual and tactile) | 1 |
|
Proprioceptive (deep
sensation, pushing paw against table edge to stimulate limb
muscles) | 1 |
| Beam balance | 6 |
|
Balances with steady
posture | 0 |
| Grasps
side of beam | 1 |
| Hugs
beam; one limb falls from beam | 2 |
| Hugs
beam; two limbs fall from beam or spins on beam (>60 sec) | 3 |
|
Attempts to balance on beam
but falls off (>40 sec) | 4 |
|
Attempts to balance on beam
but falls off (>20 sec) | 5 |
| Falls
off; no attempt to balance or hang on to beam (20 sec) | 6 |
| Reflex absence and
abnormal movement | 4 |
| Pinna
(head shake when auditory meatus is touched) | 1 |
| Corneal
(eye blink when cornea is lightly touched with cotton) | 1 |
| Startle
(motor response to brief snapping clipboard noise) | 1 |
|
Seizure, myoclonus,
myodystony | 1 |
TTC staining
The volume of cerebral infarction was measured by
TTC staining. Briefly, 5-µm coronal slices were cut and soaked in
2% TTC dye solution (cat. no. D025-1-3; Nanjing Jiancheng
Bioengineering Institute) and incubated at 37°C for 30 min. The
slides were washed with ddH2O and then stained by 2% TTC
at 37°C for 30 min. Finally, slides were fixed with 4%
paraformaldehyde at 37°C for 24 h, imaged with a camera
(DSC-RX100M6; Sony Corporation)and processed with ImageJ software
5.0 (National Institutes of Health).
RT-qPCR
Animal tissue total RNA extraction kit (cat. no.
N066; Nanjing Jiancheng Bioengineering Institute) was used to
extract total RNA from cortical tissue, according to the
manufacturer's protocol, and agarose gel electrophoresis was used
to detect the RNA integrity. A PrimeScript™ RT kit (cat. no. K1622;
Thermo Fisher Scientific, Inc.) was used to reverse transcribe
total RNA into cDNA at 37°C according to the manufacturer's
protocol. RT-qPCR was performed using SYBR Green PCR Master mix on
an ABI 7300 Real Time PCR system (both Applied Biosystems; Thermo
Fisher Scientific, Inc.). Primer 5.0 (Premier Biosoft
International) was used to design RT-qPCR primers as follows:
miR-199a forward (F), 5′-AGAAGGCGATTGATACGAGTCA-3′ and reverse (R),
5′-GGTCTCCCCAGTGTTCAGATA-3′; and U6 F, 5′-GTGCAGGGTCCGAGGT-3′ and
R, 5′-CGCTTCGGCAGCACAT-3′. The thermocycling conditions used for
RT-qPCR were as follows: Denaturation at 95°C for 20 sec; followed
by 40 cycles of annealing at 60°C for 30 sec and extension at 72°C
for 30 sec. The melting curve was drawn and relative expression of
miR-199a was analyzed by the 2−ΔΔCq method (26).
H&E staining
The brain tissue was fixed in 4% paraformaldehyde at
37°C for 24 h, dehydrated and embedded in paraffin. The sections
were cut into 4-µm thick slices (three slices/group). The slices
were placed into an oven at 60°C for 12 h. Next, the slices were
immersed in xylene I and II for 15 min; 100% ethanol I and II for
10 min; 95, 90, 80 and 70% gradient ethanol for 5 min; and
ddH2O for 5 min to remove residual ethanol. The slices
were then stained with 0.3% hematoxylin for 5 min, and then stained
blue with ddH2O for 4 min at 37°C. Next, the sections
were briefly immersed in 1% hydrochloric acid-ethanol, washed and
observed under an optical light microscope (BX51; Olympus
Corporation) at ×400 magnification until differentiation was
moderately completed. The slices were then immersed in 0.5% eosin
for 10 min at 37°C, and the residual eosin was rapidly rinsed off
using ddH2O. Next, the slices were immersed in a 70, 80,
90, 95, 100 I and 100% II ethanol gradient series for dehydration
for 5 min each and washed with xylene. Finally, the slices were
sealed with neutral resin, and observed and photographed under an
optical light microscope (BX51; Olympus Corporation) at ×400
magnification.
Nissl staining
Staining was performed in accordance with the
instructions of the Nissl staining kit (cat. no. BB44357-1; BestBio
Science; http://www.bestbio.com.cn/). A 6-µm
thick paraffin-embedded section was dewaxed using xylene and
hydrated using gradient ethanol, washed with ddH2O,
stained with 0.5% methylene blue for 10 min at 37°C and Nissl
differentiated for 10 min at 37°C. Under an optical light
microscope, Nissl bodies were observed at ×400 magnification. Next,
the slices were treated with 0.5% ammonium molybdate solution for 5
min at 37°C, washed with distilled water to prevent decolorization,
dehydrated with anhydrous ethanol and then sealed with neutral glue
after being made transparent with xylene. The morphology of
cortical neurons was observed under a light microscope (cat. no.
BX51; Olympus Corporation) at ×400 magnification.
Immunofluorescence
After being fixed with 4% paraformaldehyde at 37°C
for 24 h, brain tissue was embedded in paraffin and sectioned at a
thickness of 4 µm. The slices were incubated at 50°C for 1 h and
then washed with PBS. Next, blocking at 37°C was performed by
addition of 10% goat serum (cat. no. 16210064; Gibco; Thermo Fisher
Scientific, Inc.) for 1 h, followed by addition of mouse monoclonal
antibody against rat NeuN (1:1,000; cat. no. ab104224; Abcam) and
rabbit anti-rat microtubule-associated proteins 1A/1B light chain
3B (LC3B) polyclonal antibody (1:100; cat. no. ab222776; Abcam)
overnight at 4°C. Next, fluorescently labeled secondary antibody
(FITC-labeled goat anti mouse IgG; 1:1,000; cat. no. ab678; Abcam)
and Cy3-labeled goat anti-rabbit IgG (1:500; cat. no. ab6939;
Abcam) were incubated with the membrane at room tem-perature for 1
h. DAPI staining (5 µg/ml) was conducted for 5 min at 25°C,
followed by fluorescence quenching agent sealing, fluorescence
microscope observation with the magnification of ×400 and
imaging.
Immunohistochemistry
Paraffin-embedded sections (thickness, 5 µm) of
brain tissue were incubated in an oven at 60°C overnight and
dewaxed in water. The antigen activity was restored by the
high-pressure repair method, and 3% endogenous peroxidase ac-tivity
was inactivated by H2O2 solution, followed by
blocking with 10% goat serum (cat. no. 16210064; Gibco; Thermo
Fisher Scientific, Inc.) at room temperature for 60 min. Next,
rabbit anti-rat Beclin1 (1:500; cat. no. ab62557; Abcam), p62
(1:400; cat. no. ab91526; Abcam) and LC3B (1:2,000; cat. no.
ab51520; Abcam) monoclonal antibodies were added at 4°C for
overnight. The NC group was incubated at 4°C overnight with PBS
instead of primary antibody. After rewarming at room temperature
every oth-er day, horseradish peroxidase (HRP)-conjugated goat
anti-rabbit IgG H&L secondary antibody was added (1:5,000; cat.
no. ab205718; Abcam) at 37°C for 10–15 min. Following 0.05% DAB
staining at room temperature for 5 min, the sections were stained
with hematoxylin at room temperature for 5 min. Following
differentiation, the sections were dehydrated with 70% alcohol, 80%
alcohol, 90% alcohol and 100% alcohol, made transparent and sealed.
Sections were finally imaged under an optical light microscope
(BX51; Olympus Corporation) at ×400 magnification. The pathological
images were collected and analyzed by Motic Medical 6.0 software
[Motic (Xiamen) Electric Group Co., Ltd.]. A total of five
non-overlapping visual fields were observed in each section. The
number of Beclin1, p62 and LC3B-positive cells was counted, and the
average value was calculated. Cells with brown cytoplasm (observed
under light microscopy at ×400 magnification) were regarded as
positive.
Western blotting
The protein expression levels of Beclin1, p62, LC3B
and cleaved caspase-3 in rat cerebral cortex were detected by
western blotting. The cerebral cortex tissue was removed from
liquid nitrogen and placed into a precooling mortar for 15 min.
Protein lysate was added to the mortar to wash the ground tissue.
After mixing, the suspension was placed into an Eppendorf tube. The
suspension was placed on ice for 20 min and centrifuged at 850 × g
for 20 min at 4°C before the supernatant was collected. The total
protein of each sample was detected by a BCA protein quantification
kit. The supernatant was incubated with denatured 4X loading buffer
solution (with DTT; cat. no. P1015; Beijing Solarbio Science &
Technology Co., Ltd.) at a ratio of 3:1 and placed in a water bath
at 100°C for 10 min. Equal quantities of protein samples (30 µg)
were separated by 10% SDS-PAGE and transferred to a PVDF membrane.
The PVDF membrane was blocked with 5% skimmed milk powder at 4°C
for 60 min and incubated with rabbit anti-rat Beclin1 (1:1,000;
cat. no. ab189494; Abcam), p62 (cat. no. ab52875; Abcam), cleaved
caspase-3 (1:500; cat. no. ab49822; Abcam), LC3B (1:200; cat. no.
ab222776; Abcam) and β-actin (1:1,000; cat. no. ab8277; Abcam)
polyclonal antibodies at 4°C overnight. After washing the membrane
with washing solution, goat anti-rabbit IgG H&L (HRP) secondary
antibody (1:5,000; cat. no. ab205718; Abcam) was added and
incubated for 1 h at room temperature in the dark. Next, the
exposure solution was prepared according to the manufacturer's
instructions of the enhanced chemiluminescence kit (Thermo Fisher
Scientific, Inc.), and the color rendering expo-sure was added to
the PVDF film. ImageJ software 5.0 (National Institutes of Health)
was used for quantitative analysis.
Statistical analysis
The data were analyzed with GraphPad Prism 5.0
statistical analysis software (GraphPad Software, Inc.), and three
experimental repeats were performed and the results were expressed
as the mean ± SD. One-way ANOVA was used, followed by Tukey's post
hoc test, for comparisons between groups. P<0.05 was consid-ered
to indicate a statistically significant difference.
Results
DEX improves CIR injury in rats by
inhibiting miR-199a
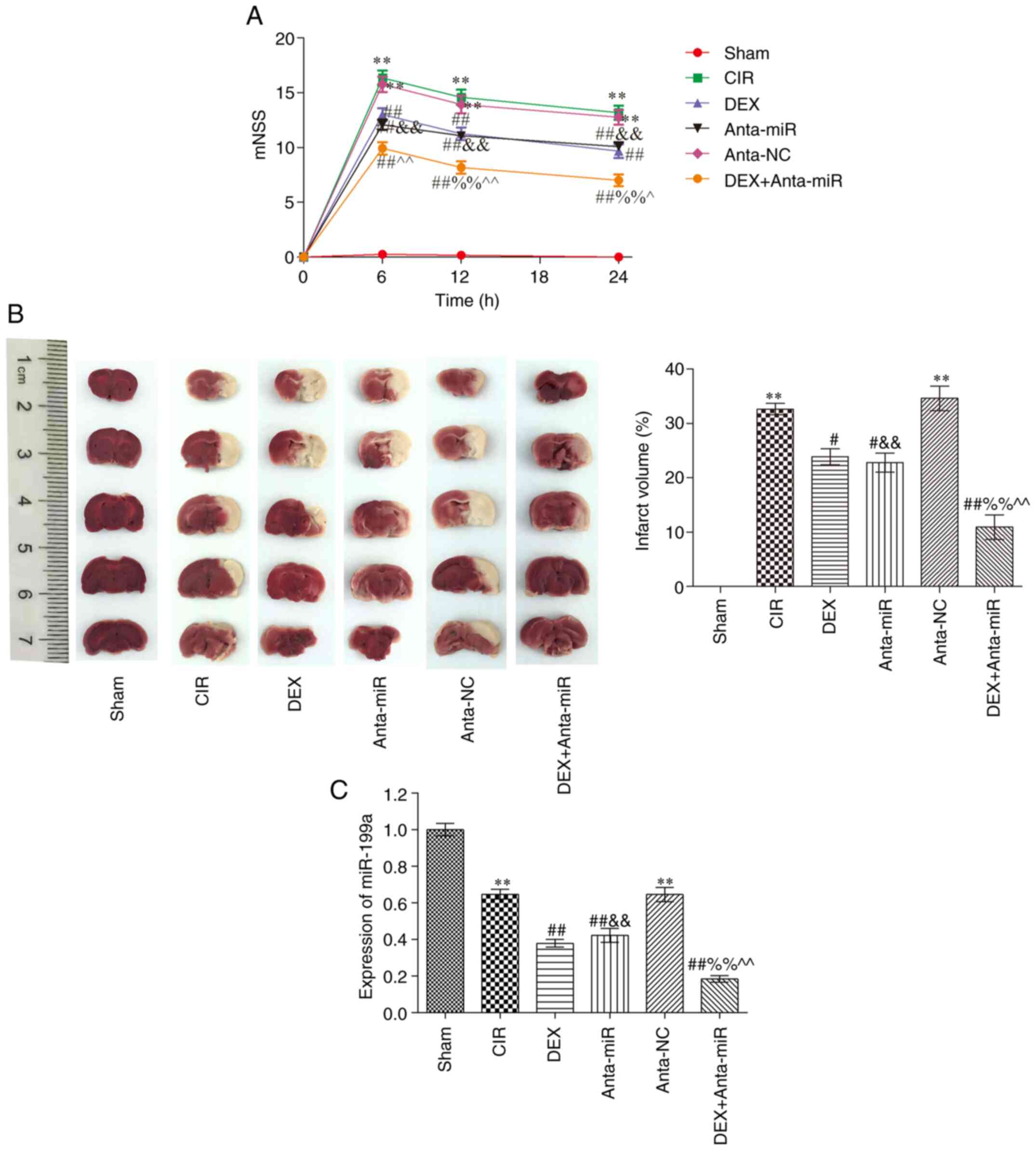
Compared with the Sham group, the mNSS of the CIR
group was significantly higher at 6 h after injury, at which point
neurological deficit was greatest (Fig.
1A). At 12 and 24 h after injury, mNSS decreased and
neurological function recovered. mNSS of the DEX and anta-miR
groups were significantly lower than those of the CIR group across
all three time points, and mNSS significantly decreased with time.
Compared with the rats in the DEX and anta-miR groups, the scores
of the DEX + anta-miR group were significantly decreased but did
not reach the level of the Sham group. Fig. 1B shows slices of one rat brain from
each group. There was no cerebral infarction in the Sham group.
Compared with the Sham group, the area of cerebral infarction in
the CIR and anta-NC groups was significantly increased. Following
DEX and anta-miR treatment, the cerebral infarction area
significantly decreased. Compared with the DEX or anta-miR groups,
the infarct area was further decreased in DEX + anta-miR group.
miR-199a was weakly expressed in the cortex of rats with CIR. DEX
and anta-miR pretreatment decreased the content of miR-199a, and
the content of miR-199a was further decreased in DEX + anta-miR
group (Fig. 1C). These data
revealed that the neuroprotection of DEX was associated with
miR-199a.
Effect of DEX on cerebral cortex
pathology in rats with CIR
The nucleoli of neurons in the cortex of the Sham
group were clear and the neurons exhibited normal morphology
(Fig. 2A). In the CIR and anta-NC
groups, neuron and nucleolus loss and nuclear pyk-nosis were
observed. Following DEX or anta-miR intervention, the loss of
neurons and pyknosis decreased. Following co-treatment with DEX and
anta-miR, pyknosis of the nucleus was weak, and almost no loss of
neurons was observed. Nissl staining revealed that neurons in the
Sham group contained abundant Nissl bodies with normal structure
and clear integrity (Fig. 2B). In
the CIR and anta-NC groups, the normal neuronal structure was
destroyed, and the number of Nissl bodies was decreased. The number
of Nissl bodies in the DEX and anta-miR groups was significantly
higher than in the CIR group, and the number of Nissl bodies
increased further after DEX was combined with anta-miR treatment.
These results suggested that DEX improved the brain pathological
injury by regulating miR-199a.
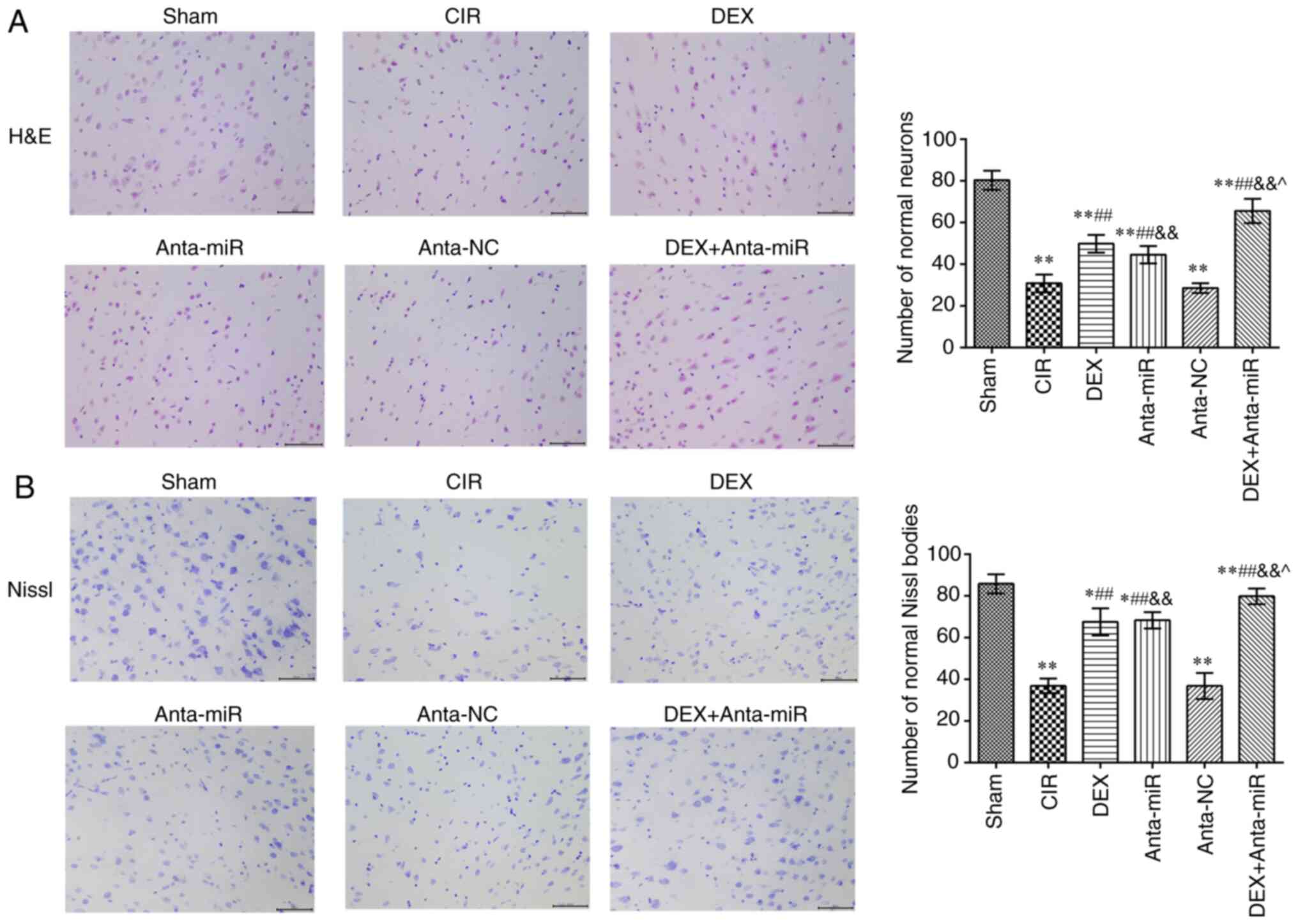 | Figure 2.Effects of DEX on histopathology of
the cerebral cortex in rats with CIR. (A) H&E staining. Scale,
50 µm. (B) Nissl staining. Scale, 50 µm. Magnification, ×400 (n=6).
*P<0.05, **P<0.01 vs. Sham; ##P<0.01 vs. CIR;
&&P<0.01 vs. anta-NC; ^P<0.05
vs. DEX. CIR, cerebral ischemia-reperfusion; DEX, dexmedetomidine;
anta, antagomir; miR, microRNA; NC, negative control; H&E,
hematoxylin and eosin. |
Effect of DEX on expression of LC3B in
the cerebral cortex of rats with CIR
Fig. 3 shows
fluorescence microscopy results of double staining with LC3B (red)
and NeuN (green). The fluorescence intensity of LC3B in the
cerebral cortex of the CIR and anta-NC groups was significantly
higher than that of the Sham group, while the NeuN inten-sity
decreased. The fluorescence intensity of LC3B and NeuN increased
following DEX or anta-miR treatment compared with that of the CIR
group. LC3B was primarily expressed in the cytoplasm, whereas NeuN
was expressed in the nucleus. This indicated that the mechanism of
DEX regulating miR-199a was associated with autophagy.
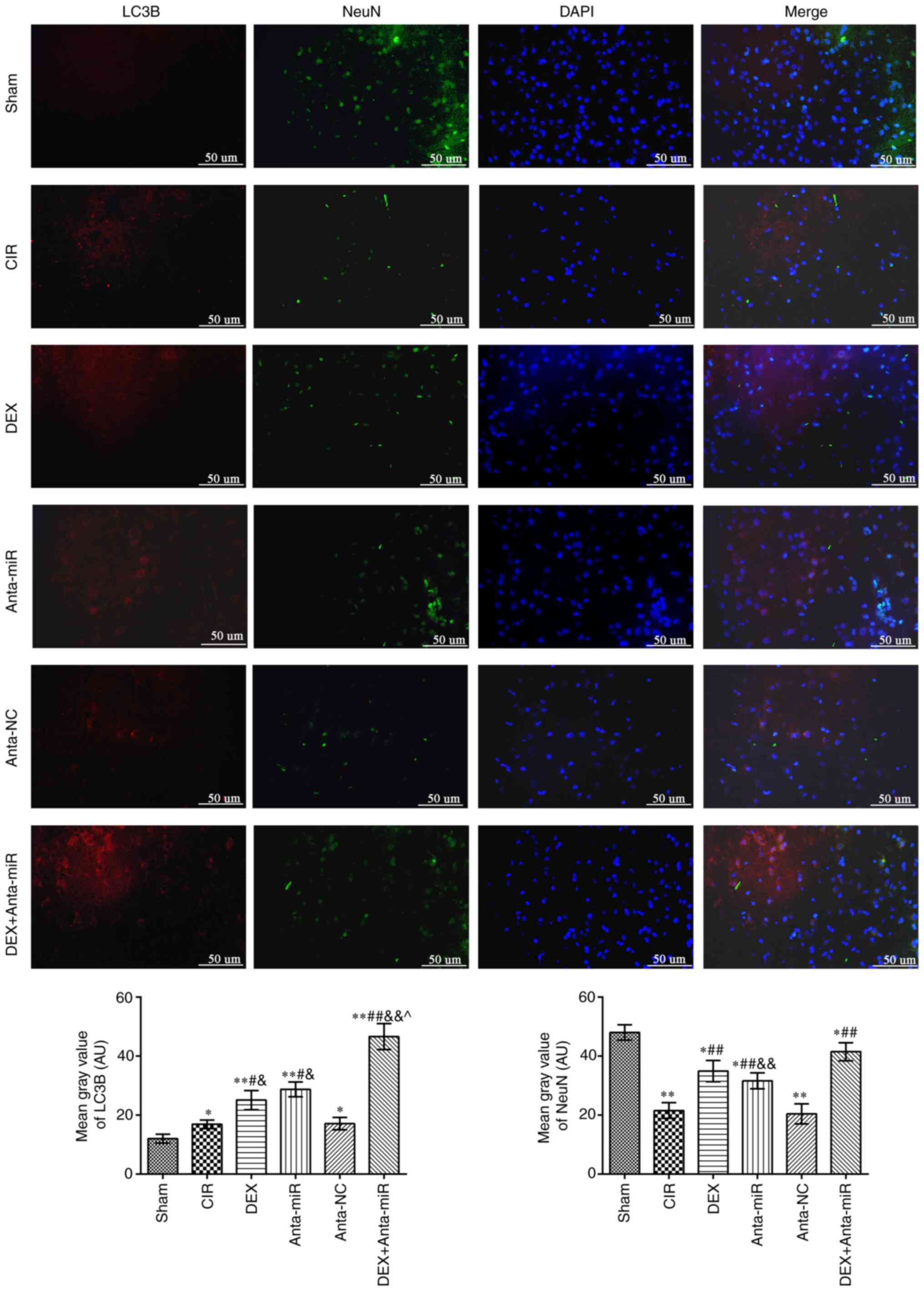 | Figure 3.DEX induces co-localization of NeuN
and LC3B in the cerebral cortex of rats with CIR.
Immunofluorescence staining was used to detect co-expression of
NeuN and LC3B. Scale bar, 50 µm. Magnification, ×400 (n=6).
*P<0.05, **P<0.01 vs. Sham; #P<0.05,
##P<0.01 vs. CIR; &P<0.05,
&&P<0.01 vs. anta-NC; ^P<0.05
vs. DEX. LC3B, microtubule-associated proteins 1A/1B light chain
3B; CIR, cerebral ischemia-reperfusion; DEX, dexmedetomidine; anta,
antagomir; miR, microRNA; NC, negative control. |
Effect of DEX on expression of
autophagy-associated proteins in the cerebral cortex of rats with
CIR
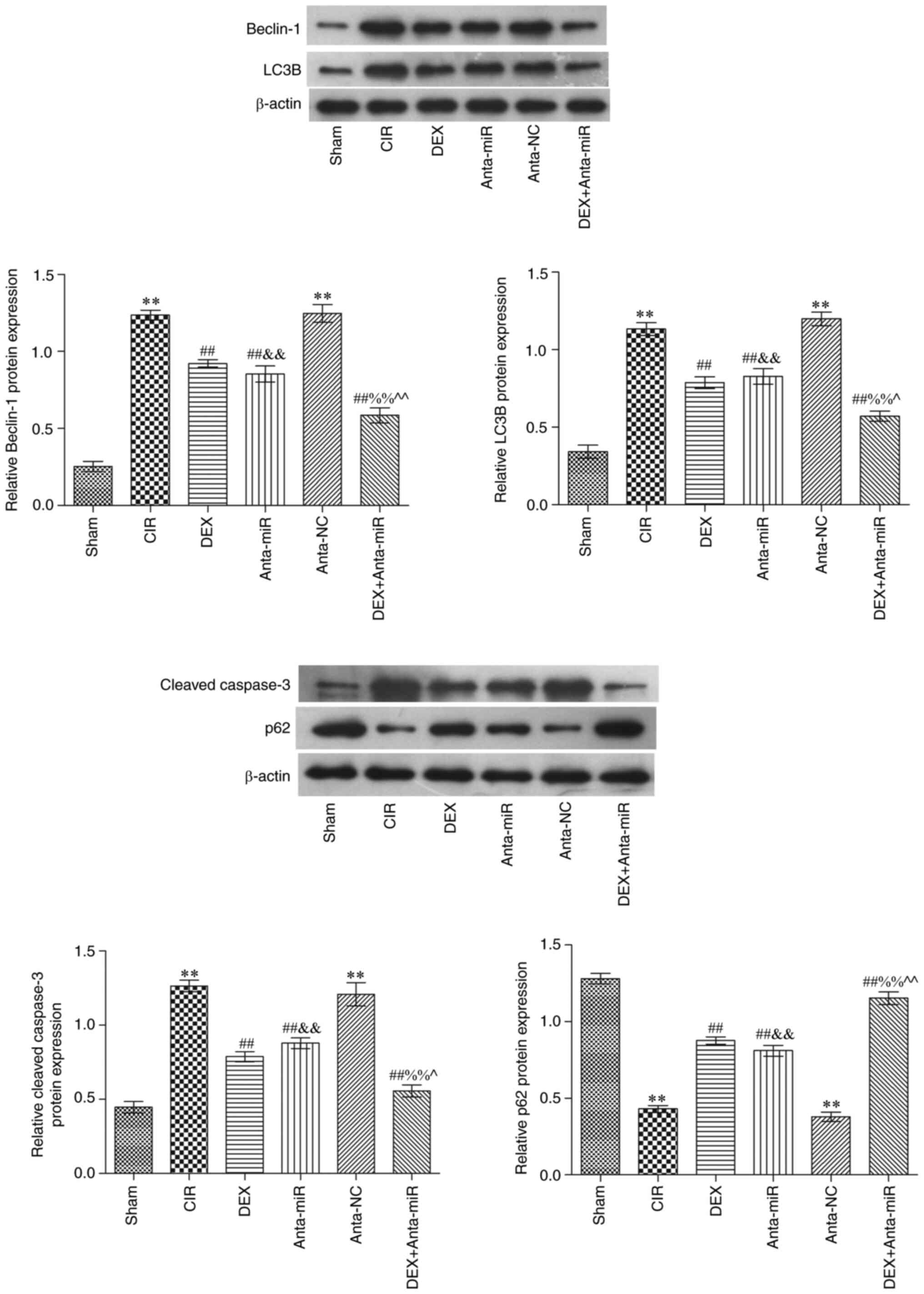
Immunohistochemical analysis of autophagy-associated
proteins in the cerebral cortex revealed that expression of Beclin1
and LC3B in the CIR and anta-NC groups was significantly increased,
while the expression of p62 was significantly decreased compared
with the Sham group (Fig. 4).
Following DEX and anta-miR inter-vention, Beclin1 and LC3B protein
expression levels were significantly lower than those in the CIR
group, while p62 protein expression significantly increased In
addition, the expression levels of Beclin1 and LC3B decreased,
while protein expression of p62 was increased following
co-treatment with DEX and anta-miR. These data further demonstrated
that DEX regulation of miR-199a in CIR was associated with
autophagy.
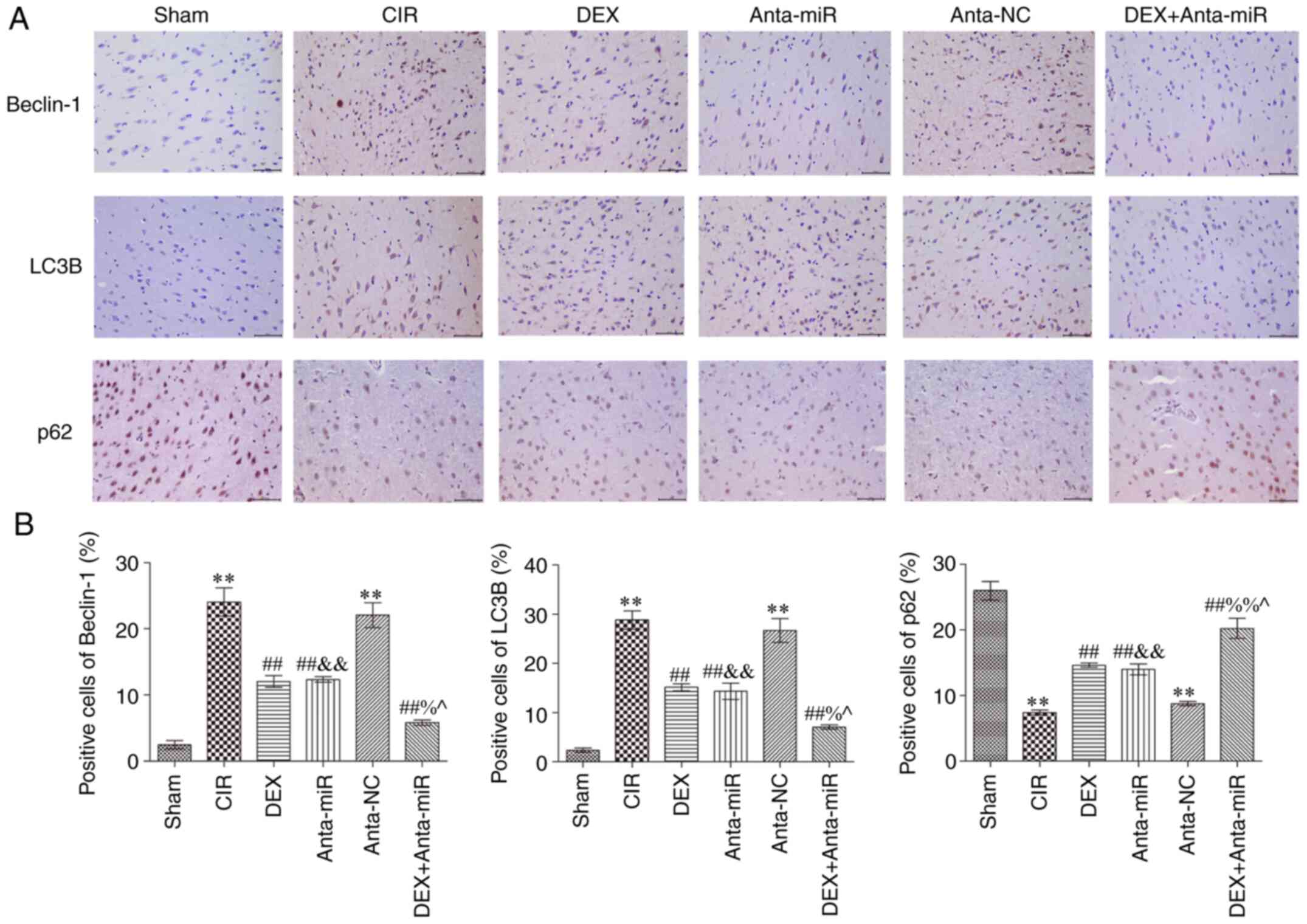 | Figure 4.DEX inhibits expression of Beclin1,
LC3B and p62 in the cerebral cortex of rats with CIR. (A)
Immunohistochemical staining. Scale, 50 µm. (B) Quantitative
analysis of Beclin1, LC3B and p62 levels. Magnification, ×400
(n=6). **P<0.01 vs. Sham; ##P<0.01 vs. CIR;
&&P<0.01 vs. anta-NC; ^P<0.05
vs. DEX; %P<0.05, %%P<0.01 vs.
anta-miR. CIR, cerebral ischemia-reperfusion; LC3B,
microtubule-associated proteins 1A/1B light chain 3B; DEX,
dexmedetomidine; Anta, antagomir; miR, microRNA; NC, negative
control. |
Effects of DEX on expression of
Beclin1, LC3B, p62 and cleaved caspase-3 in the cerebral cortex of
rats with CIR
Western blot analysis showed that Beclin1, LC3B and
cleaved caspase-3 protein expression levels were significantly
increased in the CIR and anta-NC groups, while p62 protein
expression was significantly decreased compared with the Sham group
(Fig. 5). Compared with the CIR
group, protein expression levels of p62 significantly increased
following DEX and anta-miR intervention. In addition, the
expression levels of Beclin1, LC3B and cleaved caspase-3 decreased,
while those of p62 increased following co-treatment with DEX and
anta-miR. These results were consistent with the results of the
immunohistochemistry.
Discussion
Previous studies have shown that CIR causes
oxidative stress, calcium regulation dis-order and autophagy
(27–29). Autophagy is a common type of
programmed cell death, which relies on the catabolism of autophagy
lysosomes to degrade cytoplasmic content and recycle resources
(30). Abnormal autophagy is
associated with tumors, as well as neurodegenerative, metabolic and
immune disease (31). LC3 is a
marker of autophagy and is primarily involved in the formation of
autophagy (32). The precursor
molecule of LC3 is cleaved by autophagy-related 4B cysteine peptide
to form the cytoplasmic form LC3-i. In mammals, there are four LC3
isoforms (LC3A, LC3B, LC3B2 and LC3C), and their expression varies
in different tissue and cell types (33). In previous studies (32–34),
LC3B has been demonstrated to be highly expressed in the brain and
endocrine tissue, but not in liver, colon, myocardium or spleen.
Beclin1, p62 and cleaved caspase-3 are associated with LC3B
(33). The present study detected
co-expression of LC3B and the neuron marker NeuN. It was found that
DEX inhibited autophagy and decreased nerve cell injury.
Previous studies have found that DEX improves the
survival rate of neurons following transient global or focal
cerebral ischemia in rats, and serves an important protective role
in neuronal injury induced by cerebral ischemia by inhibiting
activation of astrocytes (11,35).
The present study found that DEX decreased mNSS and improved
pathological damage of the cerebral cortex. A previous study also
found that DEX protects PC12 cells from oxidative damage via
regulation of miR-199a/hypoxia inducible factor 1α (36). Thus, the present study aimed to
investigate the association between DEX and miR-199a in neuron
injury. It was found that DEX inhibited the expression of miR-199a.
A previous study found that DEX decreases glutamate agonist-induced
neuronal apoptosis, which is associated with increased expression
levels of brain-derived neurotrophic factor in astrocytes (37). To the best of our knowledge,
however, the association between DEX and autophagy in CIR has not
been reported, and the mechanism of DEX in vitro needs
further investigation.
There is limited research on the temporal changes of
miRNAs in cerebral ischemic tolerance (38). It has been reported that 24 h after
ischemic preconditioning, the expression of miRNA in the cortex of
adult mice changes (38,39). miR-199a shows similar temporal
changes in cerebral cortex and striatum, and overexpression of
miR-199a decreases brain microvascular endothelial cell (BMEC)
death, inflammation and angiogenesis (40). These results confirm that miR-199a
has a protective effect on BMECs by inhibiting the
pathophysiological process during and after oxygen-glucose
deprivation and reperfusion. This provides a novel insight for
understanding the molecular mecha-nism of microvascular injury,
blood-brain barrier dysfunction and inflammation following ischemic
stroke, which is important for improving the treatment of ischemic
stroke (40,41). The present study found that DEX and
anta-miR pretreatment decreased levels of miR-199a. Therefore, the
present results showed that DEX protected against CIR injury by
targeting miR-199a. This finding may serve as a guide for the
treatment of patients with stroke. However, the mechanism of DEX
and miR-199a in cerebral ischemia remains unclear. Further in
vitro studies with specific cells or co-cultured cells are
needed to improve understanding of the potential mechanisms and to
identify prom-ising therapeutic targets.
In conclusion, DEX inhibited miR-199a, activated
autophagy and alleviated injury of nerve cells induced by CIR in
rats.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corre-sponding author on reasonable
request.
Authors' contributions
YL conceptualized and designed the study and
software. YZ and YL curated the data, performed the experiments and
wrote the manuscript. HZ and XM visualized the data, performed the
experiments and supervised the study. WZ contributed the study
design, wrote, reviewed and edited the manuscript. YL and HZ
confirmed the authenticity of all the raw data. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal
Protection and Use Committee of Yan-taishan Hospital (Yantai,
China; approval no. 2020-15).
Patient consent for publication
Not application.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Paul S and Candelario-Jalil E: Emerging
neuroprotective strategies for the treatment of ischemic stroke: An
overview of clinical and preclinical studies. Exp Neurol.
335:1135182021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Barteczek P, Li L, Ernst AS, Böhler LI,
Marti HH and Kunze R: Neuronal HIF-1α and HIF-2α deficiency
improves neuronal survival and sensorimotor function in the early
acute phase after ischemic stroke. J Cereb Blood Flow Metab.
37:291–306. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Choi JI, Ha SK, Lim DJ, Kim SD and Kim SH:
S100ß, matrix metalloproteinase-9, D-dimer, and heat shock protein
70 are serologic biomarkers of acute cerebral infarction in a mouse
model of transient MCA occlusion. J Korean Neurosurg Soc.
61:548–558. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Doherty J and Baehrecke EH: Life, death
and autophagy. Nat Cell Biol. 20:1110–1117. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carloni S, Albertini MC, Galluzzi L,
Buonocore G, Proietti F and Balduini W: Melatonin reduces
endoplasmic reticulum stress and preserves sirtuin 1 expression in
neuronal cells of newborn rats after hypoxia-ischemia. J Pineal
Res. 57:192–199. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang W and Zhang J: Dexmedetomidine
preconditioning protects against lung inju-ry induced by
ischemia-reperfusion through inhibition of autophagy. Exp Ther Med.
14:973–980. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhang W, Zhang JQ, Meng FM and Xue FS:
Dexmedetomidine protects against lung ischemia-reperfusion injury
by the PI3K/Akt/HIF-1α signaling pathway. J Anesth. 30:826–833.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang YQ, Tang YF, Yang MK and Huang XZ:
Dexmedetomidine alleviates cerebral ischemia-reperfusion injury in
rats via inhibition of hypoxia-inducible factor-1α. J Cell Biochem.
Nov 19–2018.(Epub ahead of print). doi: 10.1002/jcb.28058.
|
|
9
|
Begum G, Yan HQ, Li L, Singh A, Dixon CE
and Sun D: Docosahexaenoic acid reduces ER stress and abnormal
protein accumulation and improves neuronal function following
traumatic brain injury. J Neurosci. 34:3743–3755. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hetta DF, Kamal EE, Mahran AM, Ahmed DG,
Elawamy A and Abdelraouf AM: Efficacy of local dexmedetomidine
add-on for spermatic cord block anesthesia in patients undergoing
intrascrotal surgeries: Randomized controlled multicenter clinical
trial. J Pain Res. 10:2621–2628. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu C, Fu Q, Mu R, Wang F, Zhou C, Zhang
L, Yu B, Zhang Y, Fang T and Tian F: Dexmedetomidine alleviates
cerebral ischemia-reperfusion injury by inhibiting endoplasmic
reticulum stress dependent apoptosis through the
PERK-CHOP-Caspase-11 pathway. Brain Res. 1701:246–254. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo H, Ingolia NT, Weissman JS and Bartel
DP: Mammalian microRNAs predom-inantly act to decrease target mRNA
levels. Nature. 466:835–840. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guo D, Ma J, Yan L, Li T, Li Z, Han X and
Shui S: Down-regulation of lncrna MALAT1 attenuates neuronal cell
death through suppressing beclin1-dependent autophagy by regulating
mir-30a in cerebral ischemic stroke. Cell Physiol Biochem.
43:182–194. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu P, Zuo X, Deng H, Liu X, Liu L and Ji
A: Roles of long noncoding RNAs in brain development, functional
diversification and neurodegenerative diseases. Brain Res Bull.
97:69–80. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang J, Yuan L, Zhang X, Hamblin MH, Zhu
T, Meng F, Li Y, Chen YE and Yin KJ: Altered long non-coding RNA
transcriptomic profiles in brain microvascular endothelium after
cerebral ischemia. Exp Neurol. 277:162–170. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ebert MS and Sharp PA: Roles for microRNAs
in conferring robustness to biolog-ical processes. Cell.
149:515–524. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu J, Wang F, Yang GH, Wang FL, Ma YN, Du
ZW and Zhang JW: Human microRNA clusters: Genomic organization and
expression profile in leukemia cell lines. Biochem Biophys Res
Commun. 349:59–68. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Calame K: MicroRNA-155 function in B
Cells. Immunity. 27:825–827. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yao S, Tang B, Li G, Fan R and Cao F:
miR-455 inhibits neuronal cell death by targeting TRAF3 in cerebral
ischemic stroke. Neuropsychiatr Dis Treat. 12:3083–3092. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yin KJ, Deng Z, Huang H, Hamblin M, Xie C,
Zhang J and Chen YE: miR-497 regulates neuronal death in mouse
brain after transient focal cerebral ischemia. Neurobiol Dis.
38:17–26. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the Care and Use of Laboratory Animals. 8th
edition. National Academies Press (US); Washington, DC: pp.
963–965. 2011
|
|
22
|
Longa EZ, Weinstein PR, Carlson S and
Cummins R: Reversible middle cerebral artery occlusion without
craniectomy in rats. Stroke. 20:84–91. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sha R, Han X, Zheng C, Peng J, Wang L,
Chen L and Huang X: The effects of electroacupuncture in a rat
model of cerebral ischemia-reperfusion injury following middle
cerebral artery occlusion involves microRNA-223 and the PTEN
signaling pathway. Med Sci Monit. 25:10077–10088. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang D, Li Z, Zhang Y, Wang G, Wei M, Hu
Y, Ma S, Jiang Y, Che N, Wang X, et al: Targeting of
microRNA-199a-5p protects against pilocarpine-induced status
epilepticus and seizure damage via SIRT1-p53 cascade. Epilepsia.
57:706–716. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen J, Sanberg PR, Li Y, Wang L, Lu M,
Willing AE, Sanchez-Ramos J and Chopp M: Intravenous administration
of human umbilical cord blood reduces behavioral deficits after
stroke in rats. Stroke. 32:2682–2688. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hei C, Liu P, Yang X, Niu J and Li PA:
Inhibition of mTOR signaling confers protection against cerebral
ischemic injury in acute hyperglycemic rats. Int J Biol Sci.
13:878–887. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang M, Li J, Peng Q, Liu Y, Liu W, Luo
C, Peng J, Li J, Yung KK and Mo Z: Neuroprotective effects of
bilobalide on cerebral ischemia and reperfusion injury are
associated with inhibition of pro-inflammatory mediator production
and down-regulation of JNK1/2 and p38 MAPK activation. J
Neuroinflammation. 11:1672014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Halestrap AP: Calcium, mitochondria and
reperfusion injury: A pore way to die. Biochem Soc Trans.
34:232–237. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Carloni S, Girelli S, Scopa C, Buonocore
G, Longini M and Balduini W: Activation of autophagy and Akt/CREB
signaling play an equivalent role in the neuroprotective effect of
rapamycin in neonatal hypoxia-ischemia. Autophagy. 6:366–377. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chu CT: Mechanisms of selective autophagy
and mitophagy: Implications for neurodegenerative diseases.
Neurobiol Dis. 122:23–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Descloux C, Ginet V, Clarke PG, Puyal J
and Truttmann AC: Neuronal death after perinatal cerebral
hypoxia-ischemia: Focus on autophagy-mediated cell death. Int J Dev
Neurosci. 45:75–85. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Qian X, Li X, Cai Q, Zhang C, Yu Q, Jiang
Y, Lee JH, Hawke D, Wang Y, Xia Y, et al: Phosphoglycerate kinase 1
phosphorylates beclin1 to induce autophagy. Mol Cell.
65:917–931.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang L, Chen C, Zhang X, Li X, Chen Z,
Yang C, Liang X, Zhu G and Xu Z: Neuroprotective Effect of Curcumin
against cerebral ischemia-reperfusion via mediating autophagy and
inflammation. J Mol Neurosci. 64:129–139. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Aryan HE, Box KW, Ibrahim D, Desiraju U
and Ames CP: Safety and efficacy of dexmedetomidine in
neurosurgical patients. Brain Inj. 20:791–798. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wu L, Xi Y and Kong Q: Dexmedetomidine
protects PC12 cells from oxidative damage through regulation of
miR-199a/HIF-1α. Artif Cells Nanomed Biotechnol. 48:506–514. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yin D, Zhou S, Xu X, Gao W, Li F, Ma Y,
Sun D, Wu Y, Guo Q, Liu H, et al: Dexmedetomidine attenuated early
brain injury in rats with subarachnoid haemorrhage by suppressing
the inflammatory response: The TLR4/NF-κB pathway and the NLRP3
inflammasome may be involved in the mechanism. Brain Res.
1698:1–10. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang P, Liang J, Li Y and Li J, Yang X,
Zhang X, Han S, Li S and Li J: Down-regulation of miRNA-30a
alleviates cerebral ischemic injury through enhancing beclin
1-mediated autophagy. Neurochem Res. 39:1279–1291. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Rane S, He M, Sayed D, Vashistha H,
Malhotra A, Sadoshima J, Vatner DE, Vat-ner SF and Abdellatif M:
Downregulation of miR-199a derepresses hypoxia-inducible
factor-1alpha and Sirtuin 1 and recapitulates hypoxia
preconditioning in cardiac myocytes. Circ Res. 104:879–886. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zuo Y, Wang Y, Hu H and Cui W:
Atorvastatin protects myocardium against ischemia-reperfusion
injury through inhibiting miR-199a-5p. Cell Physiol Biochem.
39:1021–1030. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu WH, Yao XY, Yu HJ, Huang JW and Cui LY:
Downregulation of miR-199a may play a role in 3-nitropropionic acid
induced ischemic tolerance in rat brain. Brain Res. 1429:116–123.
2012. View Article : Google Scholar : PubMed/NCBI
|