Introduction
Ischemic heart disease is a major disease with the
highest mortality and morbidity rate (1.655%) in the world, an
estimated 31% of all deaths worldwide are due to cardiovascular
diseases (1–3). At present, the most effective
treatment for myocardial infarction is primary percutaneous
coronary intervention, but there is no effective way to prevent
myocardial ischemia-reperfusion (I/R) injury (4). Furthermore, continuous perfusion after
ischemia can cause myocardial I/R injury, leading to reperfusion
arrhythmia, myocardial shock and other pathological changes
(5). Therefore, alleviating
myocardial I/R injury is considered particularly important for the
prevention and treatment of ischemic myocardial injury.
Long non-coding (lnc)RNAs are >200 nucleotides in
length and have no significant protein-coding potential. lncRNAs
are associated with different cellular biological processes, such
as cell apoptosis, invasion and proliferation (6). Furthermore, it has been found that
lncRNAs are involved in the pathogenesis of cardiovascular disease
(7). Upregulation of lncRNA H19
imprinted maternally expressed transcript reduced myocardial
infarction-induced myocardial injury through the regulation of
KDM3A expression (7). Inhibition of
lncRNA KCNQ1OT1 protected against oxygen glucose
deprivation/reperfusion (OGD/R)-induced myocardial cell injury via
the p38/MAPK/NF-κB signaling pathway (8). A recent study demonstrated that
lncRNAs can act as competing endogenous (ce)RNAs to specifically
bind to microRNAs (miRNAs) and reduce the regulatory effect of
miRNAs on targeted mRNAs (9). For
example, lncRNA downregulation of lncRNA MALAT1 inhibited
OGD/R-induced myocardial cell injury by acting as a miRNA (miR)-20b
sponge and regulating autophagy (10). Inhibition of lncRNA GAS5 can
alleviate myocardial reperfusion injury by acting as a ceRNA for
miR-335 and regulating Rho-associated coiled-coil containing
protein kinase 1 expression (11).
It has been reported that lncRNA SNHG8 acts as a potential
biomarker and may participate in ischemia in myocardial cells
(12). Results from the present
study further identified the association between SNHG8 and
miR-335 in protection from hypoxia-ischemia-reoxygenation
(HI/R)-induced myocardial cell injury.
Heart failure, myocardial infarction and I/R injury
are the main factors in the pathogenesis of heart disease that may
result in cell death (13).
Cardiomyocytes are lost as a result of apoptosis and necrosis in
heart disease (14). Inhibition of
lncRNA HOTAIR could increase the apoptotic rate of hypoxia-induced
cardiomyocytes (15). LncRNA Gpr19
could protect OGD/R-induced myocardial cell injury by reducing cell
apoptosis and activation of the miR-324-5p/mitochondrial fission
regulator 1 axis (16). Bcl-2 and
Bax proteins, members of the Bcl-2 family, and caspase-3, their
downstream target, are considered to be the main regulators of cell
apoptosis; they are expressed in the cytoplasm in the form of
zymogens and subsequently activate cell apoptosis (17). Knockdown of lncRNA ZFAS1 expression
could decrease I/R-induced myocardial cell apoptosis by regulating
the changes in Bcl-2, Bax and cleaved caspase-3 expression
(18). Overexpression of lncRNA
SNHG8 was reported to inhibit chronic cerebral
ischemia-induced neuron apoptosis (19). Therefore, the present study
hypothesized that SNGH8 may protect against HI/R-induced myocardial
injury by regulating apoptosis. Furthermore, it has been reported
that the RAS signaling pathways serve a role in the development of
a number of diseases, such as cancer and cardiac diseases (20,21).
RAS p21 protein activator 1 (RASA1; also known as p120RasGAP) was
the first RasGAP protein to be identified; it primarily acts by
negatively regulating RAS signaling (22,23).
RASA1 has been shown to serve a role in the cardiac myocyte growth
induced by hypertrophic stimuli (24).
Therefore, the aim of the present study was to
investigate the protective effects of lncRNA SNHG8 in
HI/R-induced myocardial damage. SNHG8 may serve an important
role by regulating miR-335 and RASA1 expression, suggesting that
SNHG8 could be a novel therapeutic strategy for treatment of
myocardial I/R injury.
Materials and methods
Cell culture and HI/R conditions
The embryonic rat cardiomyocyte-derived H9C2 cell
line was purchased from the American Type Culture Collection and
grown in Gibco Dulbecco's modified Eagle's medium (DMEM; Thermo
Fisher Scientific, Inc.), which contained Gibco 10% fetal bovine
serum (FBS; Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin (MilliporeSigma) in a humidified incubator
under standard conditions (5% CO2). After a 24-h
incubation at 37°C, cells were washed three times with
phosphate-buffered saline (PBS) and the growth media was replaced
with glucose- and FBS-free DMEM. Cells were then cultured in a
Thermo 3131 hypoxic incubator (<0.1% O2, 5%
CO2 and 95% N2; Thermo Fisher Scientific,
Inc.) at 37°C for 24 h (Hypoxia-ischemic; HI). Following treatment,
the cells then were reoxygenated in an incubator in a 5%
CO2 atmosphere for 2 h (Hypoxia-ischemic-reoxygenate;
HI/R). H9C2 cells were used to HI/R conditions to induce a
myocardial I/R injury model in vitro.
Transfection
H9C2 cells were seeded into 24-well plates at a
density of 1×105 cells/well. The cells were cultured
overnight before transfection at 37°C. Next, transfection with
SNHG8 siRNA (100 nM), RASA1 siRNA (100 nM), negative control
(NC; scrambled sequence) siRNA (Santa Cruz Biotechnology, Inc.),
RASA1 plasmid, NC (empty plasmid) plasmid (OriGene Technologies,
Inc.), miR-335 mimics, NC-mimics inhibitor or NC-inhibitor
(Guangzhou RiboBio Co., Ltd.) into H9C2 cells was performed using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
transfection medium was replaced with complete medium 6 h after
transfection, and the cells were incubated for the indicated times.
All treatments were started 24 h after transfection. The siRNA
sequences are presented in Table I
and Fig. S1 demonstrates the
transfection efficacy as determined by reverse
transcription-quantitative PCR (RT-qPCR).
 | Table I.siRNAs, miRNA, miRNA inhibitors and
NC sequences used for transfections. |
Table I.
siRNAs, miRNA, miRNA inhibitors and
NC sequences used for transfections.
| Gene | Sequence
(5′→3′) |
|---|
| NC siRNA |
CATAGCGGTGTAGTAAAGCATAATA |
| SNHG8 siRNA |
ATTACGATGGATGATGGAAACATA |
| RASA1 siRNA |
TAGGATATTACAGTCACGT |
| NC-mimic |
UUCUCCGAACGUGUCACGUTT |
| miR-335 mimics | UCAAGAGCAA
UAACGAAAAAUGU |
| NC inhibitor |
GTGTAACACGTCTATACGCCCA |
| miR-335
inhibitor |
ACAUUUUUCGUUAUUGCUCUUGA |
Cell viability
Cell viability was examined using a Cell Counting
Kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Inc.). The
H9C2 cells transfected with SNHG8 siRNA, miR-335 mimics,
inhibitor, RASA1 siRNA or NC were seeded into 96-well plates at a
density of 3×103 cells/well incubation at 37°C for 24 h.
Then, 10 µl CCK-8 solution was added to the cells (100 µl/well) and
incubated at 37°C for 3 h. An MRX II microplate reader (Dynex
Technologies, Inc.) was used to detect the absorbance at 450
nm.
Western blot analysis
The cells were lysed by the cell lysis buffer (Cell
Signaling Technology, Inc.) and samples centrifuged at 300 × g for
5 min at 4°C after the lysis treatment. The supernatant was
collected and the protein concentrations measured with a BCA
Protein Assay kit (Sigma-Aldrich; Merck KGaA). The proteins (20
µg/lane) were separated by 10% SDS-PAGE and then transferred to
polyvinylidene difluoride membranes (EMD Millipore). The membranes
were blocked with Tris-buffered saline (TBS) and 0.1% Tween 20
(TBS-T) containing 5% bovine serum albumin (BSA; Sigma-Aldrich;
Merck KGaA) at room temperature and then incubated with anti-Bax
(cat. no. ab182734), anti-Bcl-2 (cat. no. ab194583) or anti-cleaved
caspase-3 (cat. no. ab2302; all from Abcam; 1:1,000 in TBST
containing 5% BSA) primary antibodies overnight at 4°C. The
membranes were washed with TBST three times and then incubated with
the appropriate horseradish peroxidase-labeled secondary antibody
(1:2,000, Cell Signaling Technology, Inc.; cat. no. 7076) for 2 h
at room temperature. Finally, the ECL Plus detection system (EMD
Millipore) was used to examine the protein bands and the expression
levels of the target proteins were semi-quantified by detecting the
optical density value of each band. GAPDH (Cell Signaling
Technology, Inc.; cat. no. 5174; 1:1,000) served as an internal
control.
EdU assay
The H9C2 cells were seeded in 96-well plates at a
density of 3×103 cells/well in culture media. The medium
was replaced with serum-free medium to synchronize the cells. After
24 h, the serum-free medium was replaced with growth media for 48
h. Cell proliferation was assessed using a Click-iT EdU imaging kit
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions.
RT-qPCR analysis
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to extract total RNA from the
H9C2 cells according to the manufacturer's instructions. Total RNA
was then reverse transcribed to cDNA using a PrimeScript RT Reagent
kit (Takara Biotechnology, Co., Ltd.; cat. no. RR047A). qPCR was
then performed using a SYBR Green RT-PCR Kit (Takara Biotechnology,
Co., Ltd.). The PCR conditions were as follows: 40 cycles of 95°C
for 30 sec, 60°C for 34 sec and 72°C for 30 sec. U6 (Takara
Biotechnology, Co., Ltd.; cat. no. 638315) and β-actin served as
the internal controls. The U6 was used for miRNA, and mRNA and
lncRNA was normalization by β-actin. The relative expression levels
of lncRNA SNHG8, miR-335 and RASA1 expression were assessed
using the comparative 2−ΔΔCq method (25) and primer sequences are presented in
Table II and Table SI.
| Table II.Primer sequences used for reverse
transcription-quantitative PCR. |
Table II.
Primer sequences used for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| SNHG8 | F:
GACACAAGGTGGCTATGGTGCTG |
|
| R:
CATGGTGGTCGTCGCGCTAAC |
| miR-335 | F:
GCGGTCAAGAGCAATAACGAA |
|
| R:
GTGCAGGGTCCGAGGTATTC |
| RASA1 | F:
CTGGAGATTATTCCCTGTATTTTCG |
|
| R:
TGTTCTTTCCGATAGTGGTCTATGA |
| β-actin | F:
CCTCTATGCCAACACAGTGC |
|
| R:
CATCGTACTCCTGCTTGCTG |
Dual-luciferase reporter assays
The cDNA fragments of SNHG8 carrying the
wild-type (WT) or mutated (MuT) binding sites of miR-335 and the
3′-untranslated region (UTR) of the amplified RASA1 fragment
containing the predicated target sites for miR-335 were synthesized
by Shanghai GenePharma Co., Ltd. H9C2 cells (1×105
cells/well) were seeded into 24-well plates and co-transfected with
50 ng recombinant luciferase vectors, 10 ng pGL3 vectors and 50 nM
miR-335 mimics, miR-335 inhibitor or NC-mimic, NC-inhibitor using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.).
After transfection for 48 h, the cells were lysed and luciferase
activities were evaluated using the Dual-Luciferase Reporter Assay
System (Promega Corporation) and a luminometer (Glomax20/20;
Promega Corporation).
Lactate dehydrogenase (LDH) release
analysis
The expression of LDH was determined by Cytotoxicity
LDH Assay Kit-WST (Roche Diagnostics). Briefly, 50 µl cell
suspension (2.5×104 cells) was added to each well of a
96-well plate after transfection with or without SNHG8
siRNA, RASA1 siRNA or miR-335 inhibitor and incubated at 37°C for 1
h. Lysis buffer (10 µl) was added to each well and incubated at
37°C for 30 min. Then, 100 µl working solution was added to each
well. The plate was protected from light and incubated at room
temperature for 30 min. Finally, 50 µl stop solution was added to
each well and the absorbance was measured at 490 nm using a
microplate reader.
Detection of cell apoptosis
The number of apoptotic cells was determined by flow
cytometry using the Annexin V-FITC Apoptosis Detection Kit (Dojindo
Molecular Technologies, Inc.). Different treatment groups of H9C2
cells were treated with a trypsin-EDTA (0.25%) solution and
centrifuged at 300 × g for 3 min at 4°C. Annexin V Binding
Solution, diluted 10-fold, was added to make a final cell
concentration of 1×106 cells/ml. The cell suspension
(100 µl) was transferred to a new tube, 5 µl Annexin V-FITC
conjugate was added and then 5 µl PI solution was added to the cell
suspension. The samples were protected from light and incubated for
15 min at room temperature. Finally, 10-fold diluted Annexin V
binding solution (400 µl) was added and the percentage of early +
late apoptotic cells was quantified by flow cytometry with a
FACSCalibur system equipped with the CellQuest software (version
5.1; BD Biosciences).
Bioinformatics analysis
Bioinformatics analysis was performed using StarBase
v 3.0 (http://starbase.sysu.edu.cn/index.php) and miRTarBase
(http://mirtarbase.cuhk.edu.cn/php/index.php) to
determine the association between miRNA and lncRNA (26,27).
Statistical analysis
Data are expressed as the mean ± SD, and statistical
analysis was performed using GraphPad Prism 8 software (GraphPad
Software, Inc.). Statistical comparisons were conducted using
unpaired Student's t-test or one-way analysis of variance with
Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Protective effect of SNHG8 on HI/R
myocardial injury
To determine the protective effect of lncRNA on HI/R
myocardial injury, the expression levels of lncRNA were detected in
the control group and HI/R myocardial cells. The results revealed
that lncRNA SNHG8 was the most markedly upregulated lncRNA
(Fig. 1A). It was also found that
treatment with SNHG8 siRNA significantly increased
HI/R-induced cell viability and proliferation, as analyzed by the
CCK-8 assay and EdU analysis. LDH analysis demonstrated that
treatment with SNHG8 siRNA reduced HI/R-induced cell damage
(Fig. 1B-D). Subsequently,
apoptotic rates and the change of apoptotic-related protein
expressions following SNHG8 siRNA treatment in the HI/R
myocardial cells was examined. As presented in Fig. 1E and F, HI/R-induction increased
apoptosis and the protein expression levels of Bax and cleaved
caspase-3 and reduced the expression of Bcl-2 protein, whereas HI/R
treated cells transfected with SNHG8 siRNA exhibited
decreased cell apoptosis and Bcl-2 and cleaved caspase-3 expression
and increased the expression of Bcl-2 protein. These data indicated
that inhibition of SNHG8 may protect against HI/R-induced
myocardial cell injury.
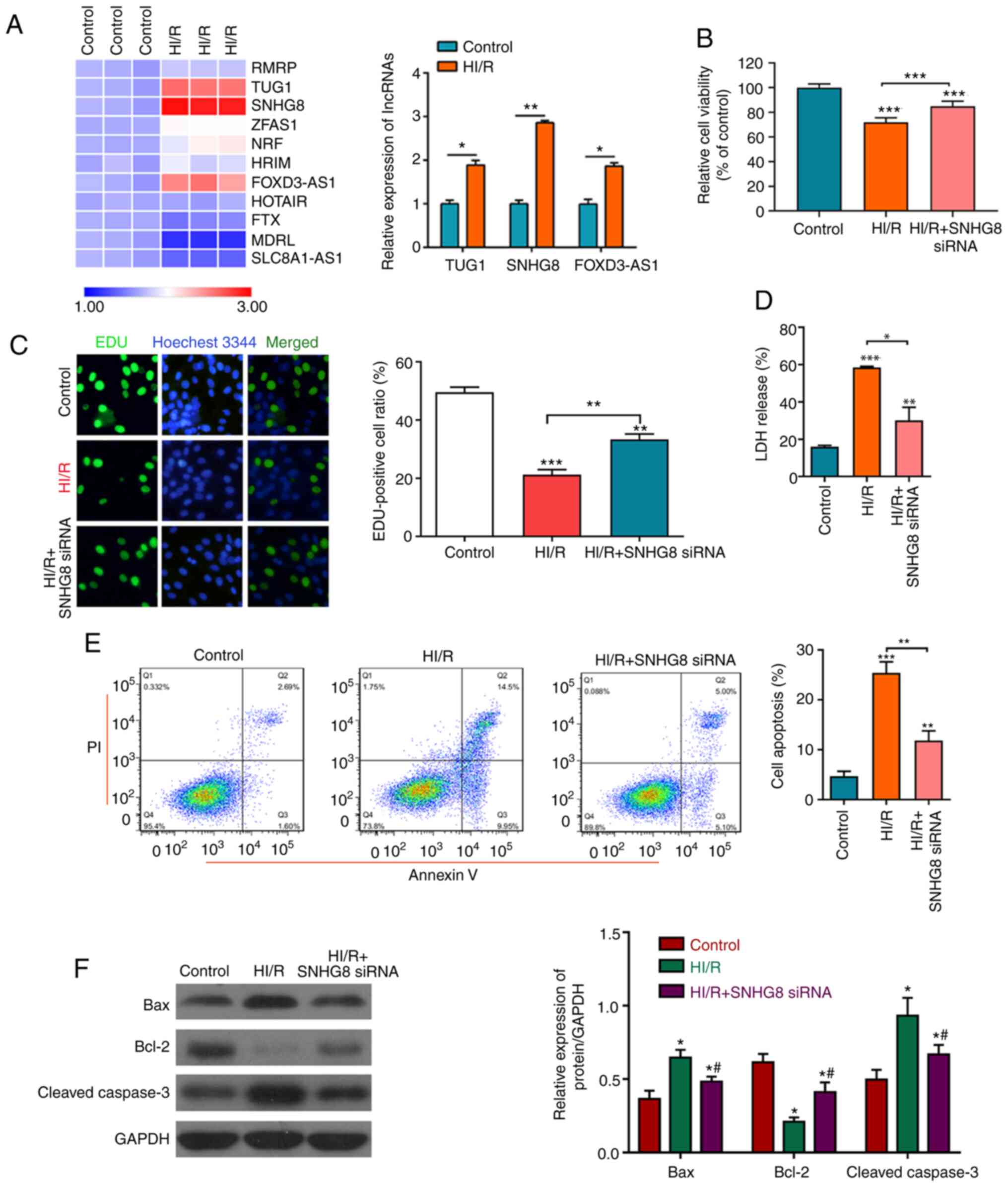 | Figure 1.Protective effect of SNHG8 on
HI/R myocardial injury. (A) Reverse transcription-quantitative PCR
was used to determine the expression levels of various lncRNAs
before or after HI/R injury in H9C2 myocardial cells. *P<0.05,
**P<0.01. (B) Cell Counting Kit-8 assay was used to examine cell
viability under different conditions. ***P<0.001 vs. Control.
(C) Cell proliferation was determined by EdU analysis. **P<0.01,
***P<0.001 vs. Control. (D) LDH assay was used to detect LDH
release. **P<0.01, ***P<0.001 vs. Control. *P<0.05. (E)
Flow cytometric analysis of cell apoptosis in different groups.
**P<0.01, ***P<0.001 vs. Control. (F) The expression levels
of Bax, Bcl-2 and cleaved caspase-3 were determined by western
blotting. *P<0.05 vs. Control. #P<0.05 vs. HI/R.
FOXD3-AS1, forkhead box D3 antisense RNA1; HI/R,
hypoxia-ischemia-reoxygenation; LDH, lactate dehydrogenase; siRNA,
small interfering RNA; SNHG8, small nucleolar RNA host gene
8; TUG1, taurine upregulated 1. |
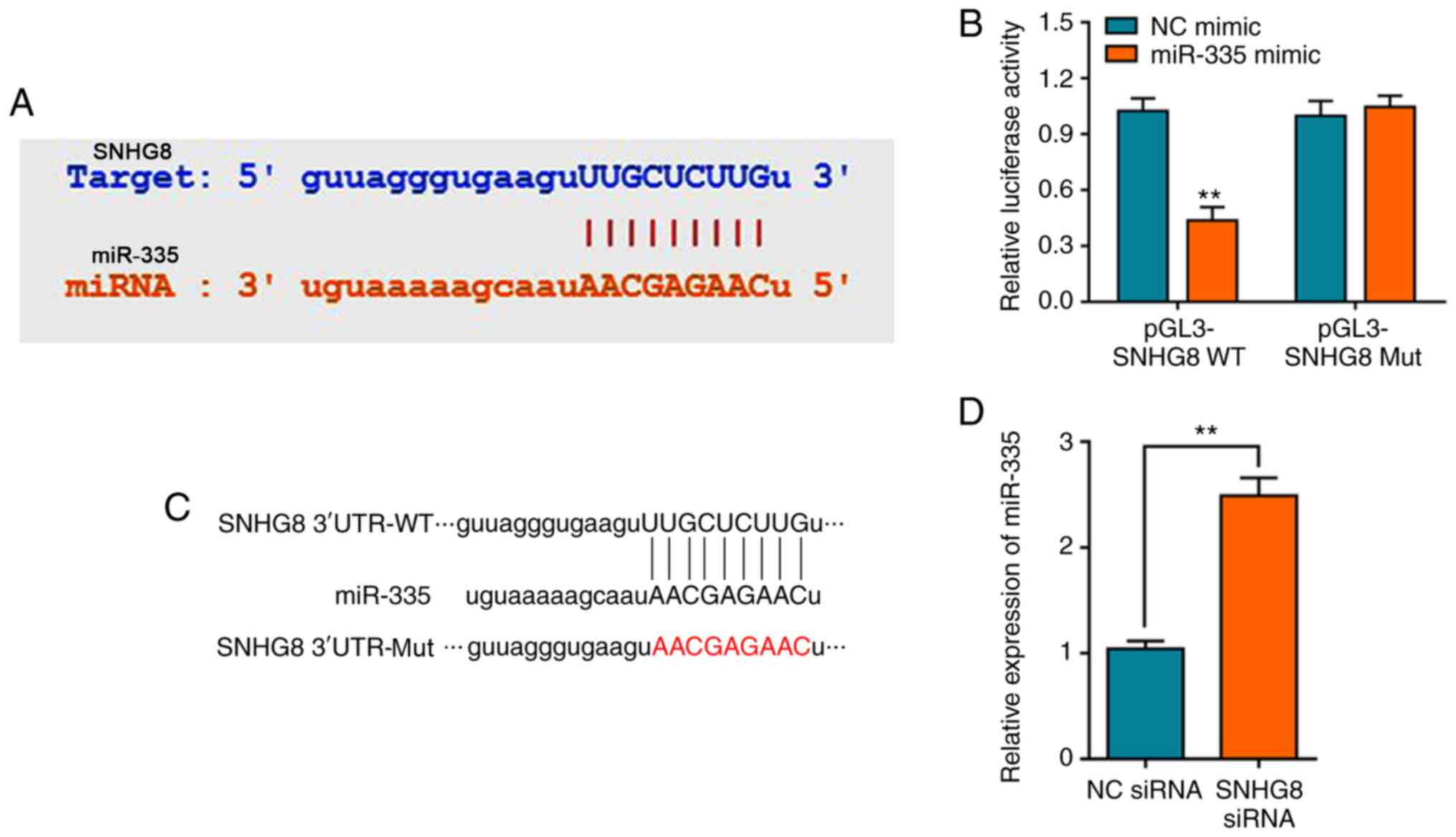
SNHG8 regulates miR-335
expression
It has previously been reported that lncRNA mediates
HI/R injury through the regulation of miRNAs, such as lncRNA MALAT1
which can regulate miRNA-20b after HI/R injury (9,28).
Bioinformatics analysis was performed to screen potential miRNAs
that have complementary base pairing with SNHG8. We firstly
determined the expression of miRNA after under HI/R condition,
showing that miR-335 was (Fig.
S2A). StarBase analysis identified miR-335 as a potential
target of SNHG8 (Fig. 2A).
Consistent with our predication, the dual-luciferase reporter assay
verified that miR-335 mimics could decrease the luciferase activity
of pGL3-SNHG8-wild-type (WT), whereas no significant
difference on pGL3-SNHG8-mutant (Mut) was observed (Fig. 2B). The schematic diagram of the
relationship between SNHG8 and miR-335 was shown in Fig. 2C. Furthermore, silencing of
SNHG8 resulted in upregulation of miR-335 expression
(Fig. 2D).
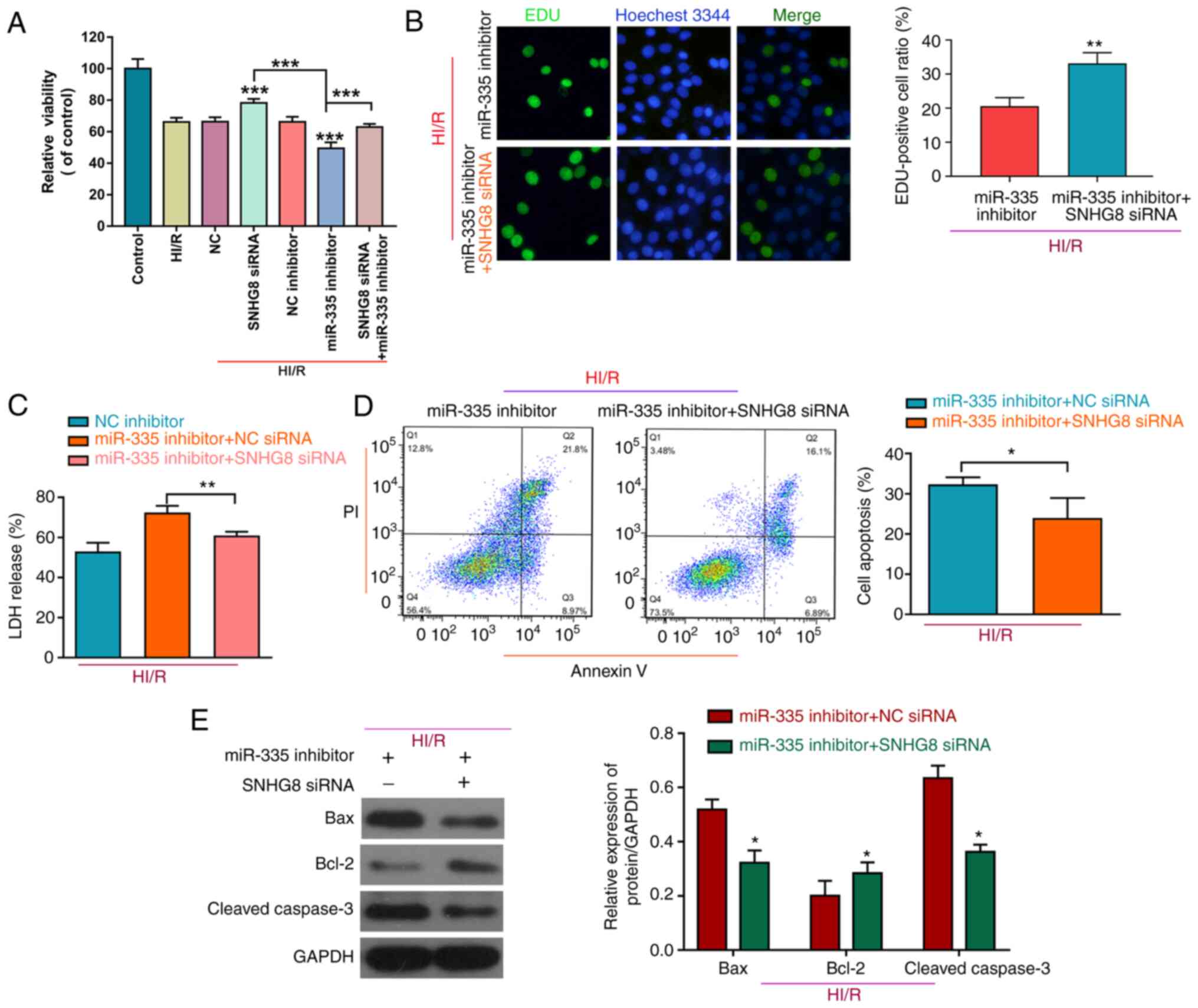
SNHG8 serves a protective role in HI/R
myocardial injury by regulating miR-335
To further determine the role of miR-335 on HI/R
myocardial injury, H9C2 cells were co-transfected with a miR-335
inhibitor and SNHG8 siRNA. As shown in Fig. 3A, treatment with the miR-335
inhibitor reduced cell viability compared with HI/R, whereas
inhibition of SNHG8 increased cell viability after
co-transfected with the miR-335 inhibitor on HI/R-induced H9C2
myocardial cell injury. Furthermore, SNHG8 siRNA combined
with the miR-335 inhibitor was able to enhance cell proliferation
and reduce LDH release compared with the miR-335 inhibitor alone
group (Fig. 3B and C,
respectively). SNHG8 siRNA reduced the miR-335
inhibitor-induced increase in the number of apoptotic cells under
the HI/R condition in H9C2 cells (Fig.
3D). Western blot analysis of apoptosis-related proteins
demonstrated that, Bax was decreased and Bcl-2 was increased
following co-transfection with SNHG8 siRNA and miR-335
inhibitor (Fig. 3E).
miR-335 negatively regulates the
expression of RASA1
Based on the aforementioned results, TargetScan and
miRTarBase were used to investigate the binding site of miR-335
(29,30). The overlapping analysis of the 196
differentially expressed mRNAs identified by TargetScan and the
predicted single mRNA identified by miRTarBase indicated that only
RASA1 interacted with miR-335 (Fig.
S2B). The results indicated that miR-335 targets the 3′-UTR of
the RASA1 mRNA (Fig. 4A). The
schematic diagram of the relationship between miR-335 and RASA1 is
shown in Fig. 4B. A dual-luciferase
reporter assay was used to further verify this prediction, and the
results indicated that miR-335 mimics decreased the luciferase
activity of pGL3-RASA1-WT, whereas no effect on pGL3-RASA1-Mut was
observed (Fig. 4C). In addition, it
was demonstrated that overexpression of miR-335 significantly
downregulated the expression level of RASA1 mRNA, whereas
transfection with the miR-335 inhibitor had the opposite effect on
the expression of RASA1 (Fig. 4D).
The expression of RASA1 following transfection of HI/R-induced H9C2
cells with SNHG8 siRNA was also confirmed. The results
demonstrated that SNHG8 siRNA treatment significantly
downregulated the expression of RASA1 (Fig. 4E).
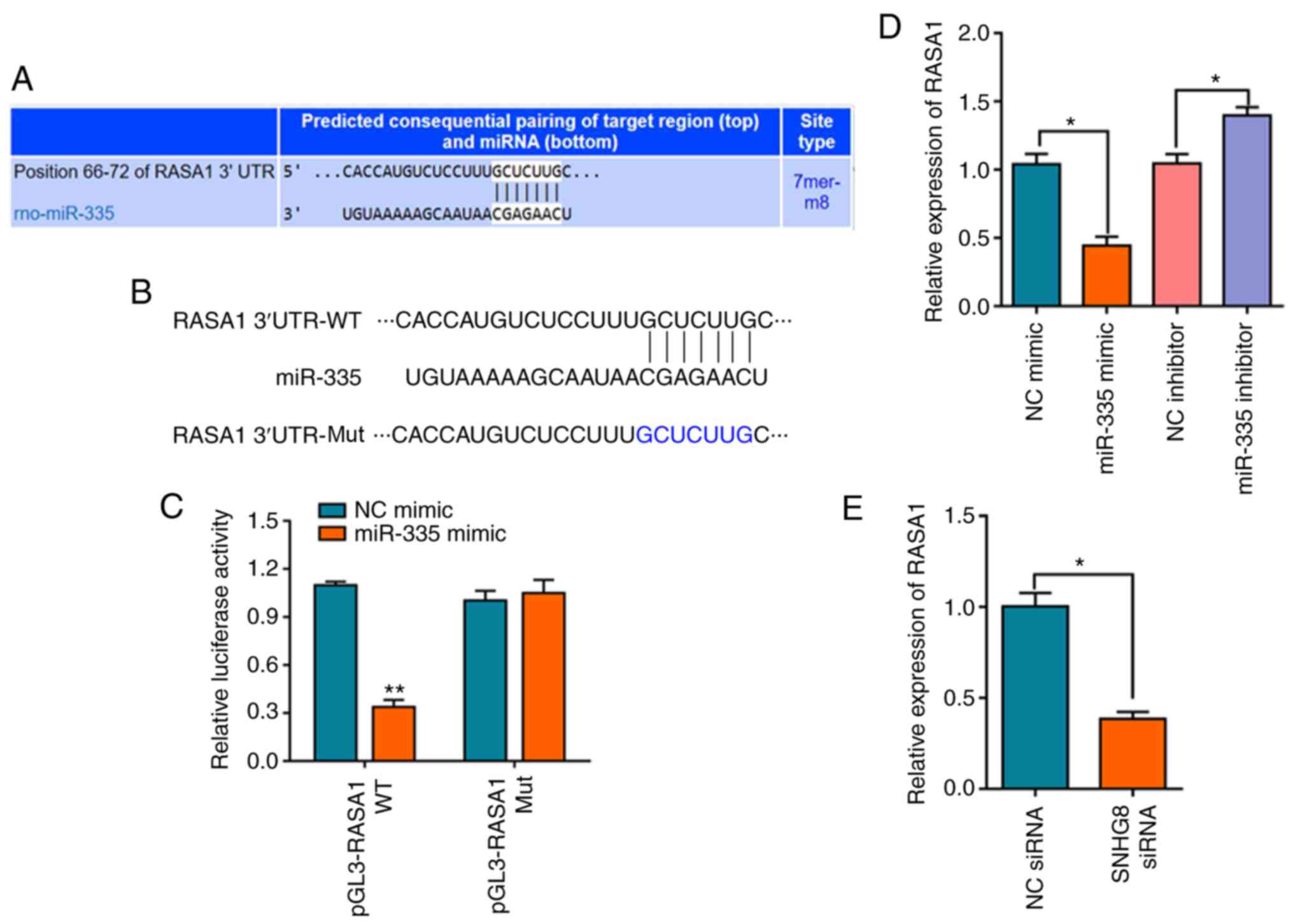 | Figure 4.miR-335 negatively regulates the
expression of RASA1. (A) TargetScan analysis was used to predict
matches between miR-335 and the RASA1 3′-UTR. (B) Schematic diagram
of the relationship between miR-335 and RASA1. (C) Relative
luciferase activities of WT and Mut RASA1 reporter plasmid in H9C2
cells after co-transfection with miR-335 mimics. **P<0.01 vs. NC
mimic. (D) The expression level of RASA1 mRNA was determined by
reverse transcription-quantitative PCR after transfection with the
miR-335 mimic, inhibitor or NC. *P<0.05. (E) SNHG8 siRNA
transfection downregulated RASA1 expression. *P<0.05. miR,
microRNA; Mut, mutant; NC, negative control; RASA1, RAS p21 protein
activator 1; rno, Rattus norvegicus; siRNA, small
interfering RNA; SNHG8, small nucleolar RNA host gene 8;
UTR, untranslated region; WT, wild-type. |
SNHG8 protects against HI/R myocardial
injury through regulation of miR-335 and RASA1
To investigate whether miR-335 and RASA1 mediated
the protective effect of SNHG8 on HI/R-induced H9C2 cell
injury, a series of experiments was performed. Knockdown of RASA1
protected HI/R-induced H9C2 cells by increasing cell viability and
cell proliferation, and decreasing LDH release, whereas compared
with RASA1 siRNA + HI/R group, there was no significant difference
in cell viability, proliferation and LDH following treatment with
SNHG8 siRNA and RASA1 siRNA (Fig. 5A-C). Moreover, following RASA1
silencing, SNHG8 did not further reduce apoptosis, again
demonstrating that SNHG8 may act through RASA1 (Fig. 5D). Western blot analysis showed
that, compared with the RASA1 siRNA group, there was no significant
difference in Bax and Bcl-2 expression after combined treatment
with RASA1 siRNA and SNHG8 siRNA (Fig. 5E).
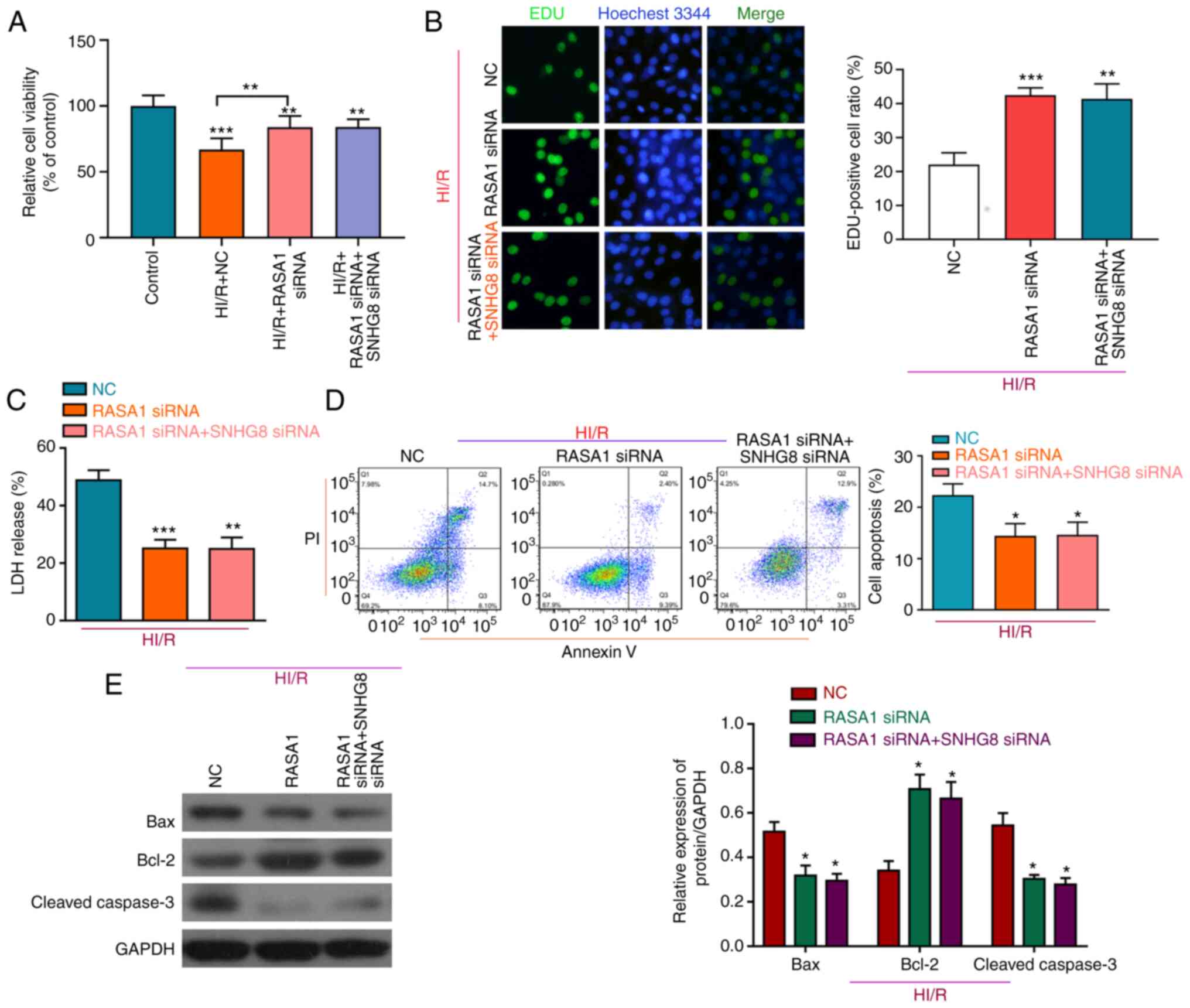 | Figure 5.RASA1 mediates the protective effect
of SNHG8 on HI/R myocardial injury. (A) Cell Counting Kit-8
viability assay and (B) EdU analysis of cell proliferation after
transfection with RASA1 siRNA alone or after co-transfection with
SNHG8 siRNA. **P<0.01, ***P<0.001 vs. Control. (C) LDH
expression was determined following transfection with RASA1 siRNA
alone or co-transfection with SNHG8 siRNA. **P<0.01,
***P<0.001 vs. NC. (D) The number of apoptotic cells was
determined by flow cytometry. *P<0.05 vs. NC. (E) Western blot
analysis of Bax, Bcl-2 and cleaved caspase-3 protein expression
levels in H9C2 cells following transfection RASA1 siRNA alone or
co-transfection of RASA1 siRNA and SNHG8 siRNA. *P<0.05
vs. NC. HI/R, hypoxia-ischemia-reoxygenation; LDH, lactate
dehydrogenase; miR, microRNA; NC, negative control; RASA1, RAS p21
protein activator 1; siRNA, small interfering RNA; SNHG8,
small nucleolar RNA host gene 8. |
The protective effect of SNHG8 combined with
RASA1 overexpression plasmid on HI/R induced H9C2 cells was
analyzed further. The data demonstrated that RASA1 overexpression
reduced cell viability, whereas SNHG8 siRNA combined with
RASA1 overexpression increased viability (Fig. S2C). These results demonstrated that
lncRNA SNHG8 may regulate the expression of RASA1 and that
RASA1 may mediate the effects SNHG8 on the damage to cells.
These data indicated that SNHG8 may protect HI/R-induced
H9C2 cell injury through the regulation of miR-335 and RASA1
expression.
Discussion
Myocardial I/R injury is one of the major causes of
mortality worldwide (31).
Myocardial ischemia leads to hypoxia and cell apoptosis, further
aggravating myocardial tissue damage (32). Therefore, investigating strategies
to reduce myocardial injury during hypoxia are essential for
treatment of heart failure and angina pectoris.
Studies have indicated that lncRNAs are related to
cerebral ischemia and that these lncRNAs could regulate the
development of ischemic cerebrovascular disease (33,34).
They also act as ceRNAs, regulating specific RNA transcription by
competitively binding with miRNAs. This lncRNA-miRNA crosstalk
might be particularly important for the control and treatment of
myocardial I/R injury and recovery after ischemia, and lncRNA-miRNA
crosstalk have also emerged as new regulators in myocardial injury
(31). Upregulation of lncRNA HULC
could reduce HI/R-induced H9C2 cell apoptosis and inflammation
through the miR-377-5p-NLRP3/caspase-1/IL-1β axis (35). Silencing of lncRNA PVT1 could reduce
HI/R injury-induced cell apoptosis and autophagy via the
miR-186/Beclin-1 signaling pathway (36). lncRNA SNHG8, located on
4qq26, belongs to the lncRNA family and is 1,062 nt in length
(37). A recent study indicated
that lncRNA SNHG8 was significantly upregulated in AMI and
that it may be correlated with regulation of myocardial cell
necrosis and apoptosis (12). The
present study found that, after HI/R induction, SNHG8
exhibited the highest expression among the lncRNAs examined, and it
was associated with ischemic myocardial cells, which was consistent
with studies by Zhuo et al (12) and Zhang and Bian (38). Inhibition of SNHG8 could
protect against HI/R-induced myocardial injury by increasing cell
viability and proliferation, and by decreasing cell apoptosis.
In the present study, a bioinformatics approach was
used to predict that the SNHG8 transcript contained a
miR-335 binding region. It has previously been reported that
miR-335 serves an important role in myocardial I/R injury (39), which is consistent with the results
of the present study. The present findings also demonstrated that
miR-335 inhibitor treatment could increase cell apoptosis and
reduce cell viability or proliferation in HI/R-induced H9C2 cells,
with co-transfection with SNHG8 siRNA, the effect of the
miR-335 inhibitor on increasing apoptosis was also reversed.
miRNAs serve an important role in cardiovascular
disease by interacting with downstream mRNAs (40,41).
For example, upregulation of miR-149 protected against myocardial
I/R damage by inhibiting forkhead box O3 expression (42). miR-7b overexpression reduced
HI/R-induced H9C2 apoptosis via the hypoxia inducible factor-1α/p38
signaling pathway (43). The
present study demonstrated that miR-335 negatively regulates the
expression of RASA1. Combined with the aforementioned results, it
was confirmed that knockdown of SNHG8 could decrease RASA1
expression. Moreover, the present study confirmed that RASA1
mediated the protective effect of SNHG8 in HI/R-induced
myocardial cell injury. Therefore, it was demonstrated that
SNHG8 may control HI/R-induced myocardial cell injury by
regulating miR-335 and RASA1.
The main factors leading to reperfusion injury are
oxidative stress, inflammation and apoptosis (44). Myocardial apoptosis is also related
to ventricular dysfunction subsequent to cardiac surgery (45). Therefore, inhibition of myocardial
cell apoptosis after ischemia might be considered an effective
treatment to improve diastolic function after ischemia. The Bcl-2
family, including Bcl-2 and Bax, are related to the regulation of
cell apoptosis (46). The present
study revealed that SNHG8 could regulate apoptotic-related
proteins Bax, Bcl-2 and cleaved caspase-3 by regulating miR-335 and
RASA1. However, the present study also had certain limitations. The
protective effect of SNHG8 was verified under HI/R-induced
H9C2 cells in vitro; however, the experiment could be
extended to demonstrate the effect in vivo. Additionally,
further identification and confirmation of the precise mechanisms
underlying RASA1 are required, such as whether RASA1 regulates
Ras/MEK1/2/ERK1/2 and PI3K/Akt signaling.
In conclusion, the present study demonstrated that
transfection with SNHG8 siRNA protected against HI/R-induced
injury in H9C2 cells by mediating the regulation of miR-335 and
RASA1, thus indicating that SNHG8 may be an effective target
for treatment of myocardial I/R injury.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The current study was funded by The Traditional
Chinese Medicine Research Project of Jiangxi Province (grant no.
2019A151).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
DR and LH conceived the idea. YL and PZ performed
the experiments. FW, XZ and DY analyzed the data. DR and YL wrote
the manuscript. All authors have read and approved the final
manuscript. DR and LH confirm the authenticity of all the raw
data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Severino P, D'Amato A, Pucci M, Infusino
F, Adamo F, Birtolo LI, Netti L, Montefusco G, Chimenti C, Lavalle
C, et al: Ischemic heart disease pathophysiology paradigms
overview: From plaque activation to microvascular dysfunction. Int
J Mol Sci. 21:81182020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reed GW, Rossi JE and Cannon CP: Acute
myocardial infarction. Lancet. 389:197–210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Devereaux PJ and Szczeklik W: Myocardial
injury after non-cardiac surgery: Diagnosis and management. Eur
Heart J. 41:3083–3091. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jokinen E: Coronary artery disease in
patients with congenital heart defects. J Intern Med. 288:383–389.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Khan MA, Hashim MJ, Mustafa H, Baniyas MY,
Al Suwaidi SKBM, AlKatheeri R, Alblooshi FMK, Almatrooshi MEAH,
Alzaabi MEH, Al Darmaki RS and Lootah SNAH: Global epidemiology of
ischemic heart disease: Results from the global burden of disease
study. Cureus. 12:e93492020.PubMed/NCBI
|
|
6
|
Li M, Duan L, Li Y and Liu B: Long
noncoding RNA/circular noncoding RNA-miRNA-mRNA axes in
cardiovascular diseases. Life Sci. 233:1164402019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang Y: The novel regulatory role of
lncRNA-miRNA-mRNA axis in cardiovascular diseases. J Cell Mol Med.
22:5768–5775. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li X, Dai Y, Yan S, Shi Y, Han B, Li J,
Cha L and Mu J: Down-regulation of lncRNA KCNQ1OT1 protects against
myocardial ischemia/reperfusion injury following acute myocardial
infarction. Biochem Biophys Res Commun. 491:1026–1033. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sui C, Dong Z, Yang C, Zhang M, Dai B,
Geng L, Lu J, Yang J and Xu M: LncRNA FOXD2-AS1 as a competitive
endogenous RNA against miR-150-5p reverses resistance to sorafenib
in hepatocellular carcinoma. J Cell Mol Med. 23:6024–6033. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang S, Yao T, Deng F, Yu W, Song Y, Chen
J and Ruan Z: LncRNA MALAT1 promotes oxygen-glucose deprivation and
reoxygenation induced cardiomyocytes injury through sponging
miR-20b to Enhance beclin1-mediated autophagy. Cardiovasc Drugs
Ther. 33:675–686. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu N, Zhang X, Bao Y, Yu H, Jia D and Ma
C: Down-regulation of GAS5 ameliorates myocardial
ischaemia/reperfusion injury via the miR-335/ROCK1/AKT/GSK-3β axis.
J Cell Mol Med. 23:8420–8431. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhuo LA, Wen YT, Wang Y, Liang ZF, Wu G,
Nong MD and Miao L: LncRNA SNHG8 is identified as a key regulator
of acute myocardial infarction by RNA-seq analysis. Lipids Health
Dis. 18:2012019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shekhar A, Heeger P, Reutelingsperger C,
Arbustini E, Narula N, Hofstra L, Bax JJ and Narula J: Targeted
imaging for cell death in cardiovascular disorders. JACC Cardiovasc
Imaging. 11:476–493. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu T, Ding W, Ao X, Chu X, Wan Q, Wang Y,
Xiao D, Yu W, Li M, Yu F and Wang J: ARC regulates programmed
necrosis and myocardial ischemia/reperfusion injury through the
inhibition of mPTP opening. Redox Biol. 20:414–426. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao L, Liu Y, Guo S, Yao R, Wu L, Xiao L,
Wang Z, Liu Y and Zhang Y: Circulating long noncoding RNA HOTAIR is
an essential mediator of acute myocardial infarction. Cell Physiol
Biochem. 44:1497–1508. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang L, Guo B, Liu S, Miao C and Li Y:
Inhibition of the LncRNA Gpr19 attenuates ischemia-reperfusion
injury after acute myocardial infarction by inhibiting apoptosis
and oxidative stress via the miR-324-5p/Mtfr1 axis. IUBMB Life.
72:373–383. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Siddiqui WA, Ahad A and Ahsan H: The
mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update.
Arch Toxicol. 89:289–317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang P, Yang D, Yu L and Shi Y:
Downregulation of lncRNA ZFAS1 protects H9c2 cardiomyocytes from
ischemia/reperfusion-induced apoptosis via the miR5903p/NFκB
signaling pathway. Mol Med Rep. 22:2300–2306. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu J, An P, Xue Y, Che D, Liu X, Zheng J,
Liu Y, Yang C, Li Z and Yu B: Mechanism of
Snhg8/miR-384/Hoxa13/FAM3A axis regulating neuronal apoptosis in
ischemic mice model. Cell Death Dis. 10:4412019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Soleimani A, Rahmani F, Saeedi N,
Ghaffarian R, Khazaei M, Ferns GA, Avan A and Hassanian SM: The
potential role of regulatory microRNAs of RAS/MAPK signaling
pathway in the pathogenesis of colorectal cancer. J Cell Biochem.
120:19245–19253. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu X, Xu Y, Deng Y and Li H: MicroRNA-223
regulates cardiac fibrosis after myocardial infarction by targeting
RASA1. Cell Physiol Biochem. 46:1439–1454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tao H, Yang JJ, Chen ZW, Xu SS, Zhou X,
Zhan HY and Shi KH: DNMT3A silencing RASSF1A promotes cardiac
fibrosis through upregulation of ERK1/2. Toxicology. 323:42–50.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Trial J, Entman ML and Cieslik KA:
Mesenchymal stem cell-derived inflammatory fibroblasts mediate
interstitial fibrosis in the aging heart. J Mol Cell Cardiol.
91:28–34. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Diao X, Shen E, Wang X and Hu B:
Differentially expressed microRNAs and their target genes in the
hearts of streptozotocin-induced diabetic mice. Mol Med Rep.
4:633–640. 2011.PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang HY, Lin YC, Li J, Huang KY, Shrestha
S, Hong HC, Tang Y, Chen YG, Jin CN, Yu Y, et al: MiRTarBase 2020:
Updates to the experimentally validated microRNA-target interaction
database. Nucleic Acids Res. 48(D1): D148–D154. 2020.PubMed/NCBI
|
|
28
|
Yang L, Lu Y, Ming J, Pan Y, Yu R, Wu Y
and Wang T: SNHG16 accelerates the proliferation of primary
cardiomyocytes by targeting miRNA-770-5p. Exp Ther Med.
20:3221–3227. 2020.PubMed/NCBI
|
|
29
|
Han J, LaVigne CA, Jones BT, Zhang H,
Gillett F and Mendell JT: A ubiquitin ligase mediates
target-directed microRNA decay independently of tailing and
trimming. Science. 370:eabc95462020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shi CY, Kingston ER, Kleaveland B, Lin DH,
Stubna MW and Bartel DP: The ZSWIM8 ubiquitin ligase mediates
target-directed microRNA degradation. Science. 370:eabc93592020.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gu S, Tan J, Li Q, Liu S, Ma J, Zheng Y,
Liu J, Bi W, Sha P, Li X, et al: Downregulation of LAPTM4B
contributes to the impairment of the autophagic flux via unopposed
activation of mTORC1 signaling during myocardial
ischemia/reperfusion injury. Circ Res. 127:e148–e165. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liang S, Ren K, Li B, Li F, Liang Z, Hu J,
Xu B and Zhang A: LncRNA SNHG1 alleviates
hypoxia-reoxygenation-induced vascular endothelial cell injury as a
competing endogenous RNA through the HIF-1α/VEGF signal pathway.
Mol Cell Biochem. 465:1–11. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ghafouri-Fard S, Shoorei H and Taheri M:
Non-coding RNAs participate in the ischemia-reperfusion injury.
Biomed Pharmacother. 129:1104192020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bao MH, Szeto V, Yang BB, Zhu SZ, Sun HS
and Feng ZP: Long non-coding RNAs in ischemic stroke. Cell Death
Dis. 9:2812018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liang H, Li F, Li H, Wang R and Du M:
Overexpression of lncRNA HULC attenuates myocardial
ischemia/reperfusion injury in rat models and apoptosis of
hypoxia/reoxygenation cardiomyocytes via targeting miR-377-5p
through NLRP3/Caspase1/IL1β signaling pathway inhibition. Immunol
Invest. Jul 17–2020.(Epub ahead of print). View Article : Google Scholar
|
|
36
|
Ouyang M, Lu J, Ding Q, Qin T, Peng C and
Guo Q: Knockdown of long non-coding RNA PVT1 protects human AC16
cardiomyocytes from hypoxia/reoxygenation-induced apoptosis and
autophagy by regulating miR-186/Beclin-1 axis. Gene.
754:1447752020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang P, Li S, Chen Z, Lu Y and Zhang H:
LncRNA SNHG8 promotes proliferation and invasion of gastric cancer
cells by targeting the miR-491/PDGFRA axis. Hum Cell. 33:123–130.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang Y and Bian Y: Long non-coding RNA
SNHG8 plays a key role in myocardial infarction through affecting
hypoxia-induced cardiomyocyte injury. Med Sci Monit.
26:e9240162020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wu N, Zhang X, Du S, Chen D and Che R:
Upregulation of miR-335 ameliorates myocardial ischemia reperfusion
injury via targeting hypoxia inducible factor 1-alpha subunit
inhibitor. Am J Transl Res. 10:4082–4094. 2018.PubMed/NCBI
|
|
40
|
Zhang Y, Fang J and Ma H: Inhibition of
miR-182-5p protects cardiomyocytes from hypoxia-induced apoptosis
by targeting CIAPIN1. Biochem Cell Biol. 96:646–654. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xiao X, Lu Z, Lin V, May A, Shaw DH, Wang
Z, Che B, Tran K, Du H and Shaw P: MicroRNA miR-24-3p reduces
apoptosis and regulates Keap1-Nrf2 pathway in mouse cardiomyocytes
responding to ischemia/reperfusion injury. Oxid Med Cell Longev.
2018:70421052018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lin J, Lin H, Ma C, Dong F, Hu Y and Li H:
MiR-149 aggravates pyroptosis in myocardial ischemia-reperfusion
damage via silencing FoxO3. Med Sci Monit. 25:8733–8743. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sheng Z, Lu W, Zuo Z, Wang D, Zuo P, Yao Y
and Ma G: MicroRNA-7b attenuates ischemia/reperfusion-induced H9C2
cardiomyocyte apoptosis via the hypoxia inducible factor-1/p-p38
pathway. J Cell Biochem. 120:9947–9955. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu X, Zhang L, Qin H, Han X, Zhang Z,
Zhang Z, Qin SY and Niu J: Inhibition of TRAF3 expression
alleviates cardiac ischemia reperfusion (IR) injury: A mechanism
involving in apoptosis, inflammation and oxidative stress. Biochem
Biophys Res Commun. 506:298–305. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pereira RM, Mekary RA, da Cruz Rodrigues
KC, Anaruma CP, Ropelle ER, da Silva ASR, Cintra DE, Pauli JR and
de Moura LP: Protective molecular mechanisms of clusterin against
apoptosis in cardiomyocytes. Heart Fail Rev. 23:123–129. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Banjara S, D Sa J, Hinds MG and Kvansakul
M: The structural basis of Bcl-2 mediated cell death regulation in
hydra. Biochem J. 477:3287–3297. 2020. View Article : Google Scholar : PubMed/NCBI
|