Introduction
Glioma is the most prevalent type of primary tumor
of the central nervous system and accounts for >75% of malignant
brain tumors (1,2). Histologically, glial-derived tumors,
or gliomas, share characteristics of normal glial cells, glial or
neural precursors and stem cells, and are generally named based on
these similarities (3–5). Recent advances in molecular
understanding have provided new insights and novel molecular
subclassification of gliomas. These molecular subtypes include
subependymal giant cell astrocytoma, pilocytic astrocytoma,
epithelioid glioblastoma, oligodendroglioma and mesenchymal
glioblastoma (2,3). Early detection of gliomas may be
difficult due to their deep location in the brain and lack of
obvious clinical symptoms at the early stages (4,6).
Despite the modest survival benefit of standard therapies,
including radiation, chemotherapy and surgery, for patients with
glioma, locoregional failure in the form of persistent or recurrent
disease still occurs in the majority of patients (7–11).
Therefore, innovative biomarkers and more specific targeted
treatments for eradicating glioma are required.
Non-coding RNAs (ncRNAs) are a class of functional
RNAs that do not encode proteins, including microRNAs
(miRNAs/miRs), long non-coding RNAs (lncRNAs) and circular RNAs
(circRNAs) (12–14). Previous studies have reported that
the majority of miRNAs and lncRNAs have regulatory functions,
including regulation of the evolution of stem cells, cardiomyocytes
and epithelial cells, and regulation of the translation,
stabilization and degradation of mRNA (15–19).
With the development of several state-of-the-art technologies, such
as the next generation of high-throughput sequencing techniques,
gene silencing and gene editing, emerging evidence has demonstrated
that, not only miRNAs and lncRNAs, but also circRNAs, have
regulatory functions, and are associated with a variety of diseases
(20–24). circRNAs can interact with protein
complexes, RNA molecules and DNA molecules to exert their effects,
and regulate a wide variety of physiological and pathological
processes (25–27).
circRNAs are a type of ncRNA characterized by a
circular configuration that consists of either prototypical
spliceosome-mediated or lariat-type splicing, which provides a
strong stability and resistance to RNases (28,29).
Numerous studies have shown that several circRNAs are abnormally
expressed in human glioma and may be associated with the
carcinogenesis of glioma. For example, circ-PRKCI (30), circ-CPA4 (31), circ-CDC45 (32), circ-SCAF11 (33), circ-POSTN (34) and circ-PCMTD1 (35) have been found to act as tumor
suppressors or promoters in glioma progression by regulating the
proliferation, invasion, metastasis, angiogenesis and apoptosis of
glioma cells. The aim of the present review was to describe the
potential functional implications of circRNAs and to summarize the
current knowledge on circRNA research in human glioma, including
the expression, regulatory pathways and treatment potential of
circRNAs, as well as the current challenges of circRNA studies in
human glioma.
Brief overview of circRNAs
General characteristics of
circRNAs
circRNAs were first identified in plants, where it
was revealed that they encode subviral agents by electron
microscopy in 1976 (36). At the
beginning of the 21st century, researchers reported the existence
of circRNAs in a wide range of eukaryotic cells and tissue samples
from humans and other species (37). With the development of circRNA
identification and selection techniques, it was reported that
circRNAs consist of diverse combinations of sequences and domains,
including intronic, exonic, intergenic and antisense chain regions
(38). Subsequently, the detected
types and quantity of circRNAs have increased. According to the
components of the parental genes, circRNAs can be grouped into
three main subtypes: Circular intronic RNAs (ciRNAs), exonic
circRNAs (ecircRNAs) and exon-intron circRNAs (EIciRNAs) (39–42).
circRNAs are resistant to degradation by
ribonucleases due to the lack of a free 5′ or 3′ end, which ensures
their stability compared with their linear counterparts (43). circRNAs are extensively present in
body fluids, and are covalently closed single-stranded RNA
molecules that are generated via multiple cyclization modes,
including lariat-driven cyclization, RNA-binding protein
(RBP)-driven cyclization, base-pairing-driven cyclization and ciRNA
biosynthesis (44,45). The cyclization modes and specific
types of circRNAs are shown in Fig.
1. Selective circularization combined with alternative splicing
can generate numerous circRNAs from one gene (20).
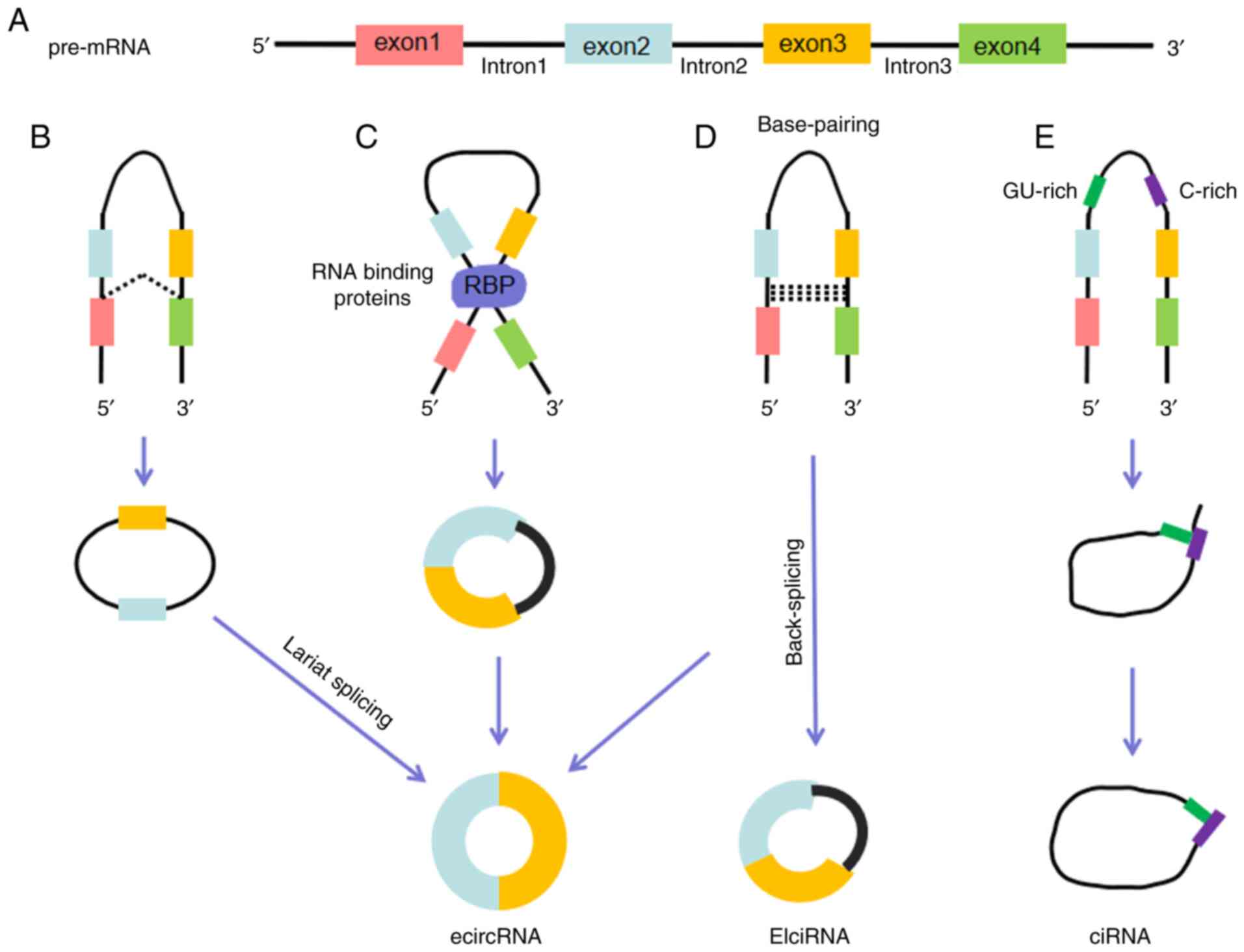 | Figure 1.Biogenesis of circRNAs. (A) Pre-mRNA
contains introns and exons, and can generate a variety of circRNAs
through different splicing mechanisms. (B) Lariat-driven
cyclization. When a pre-mRNA is spliced, the 3′ hydroxyl of the
upstream exon interacts with the 5′ phosphate of the downstream
exon to form a covalent linkage, producing a lariat that contains
exons and introns. The 2′ hydroxyl of the 5′ intron reacts with the
5′ phosphate of the 3′ intron, followed by an interaction between
the 3′ hydroxyl of the 3′ exon and the 5′ phosphate of the 5′ exon,
through which an ecircRNA is formed. (C) RBP-driven cyclization.
RBPs can promote the interaction of the downstream intron and
upstream intron, causing the formation of an ecircRNA. (D)
Base-pairing-driven cyclization. The downstream introns and
upstream introns are paired based on inverse-repeating or
complementary sequences. The introns are removed or retained to
form ecircRNA or EIciRNA. (E) Biosynthesis of ciRNA. Formation of
ciRNAs mainly depends on a 7-nt GU-rich element and an 11-nt C-rich
element to escape debranching and exonucleolytic degradation.
circRNA, circular RNA; RBP, RNA-binding protein; ecircRNA, exonic
circRNA; EIciRNA, exon-intron circRNA; ciRNA, circular intronic
RNA. |
Research on circRNAs has confirmed numerous features
of circRNAs, including diversity, specificity and conservation
(46). In comparison with linear
RNAs, the majority of circRNAs are hyper-conserved at the
nucleotide-level among different species (47). In 2021, Varela-Martinez et al
(48) identified a large number of
sheep circRNAs, of which 1,606 were homologous to human circRNAs.
Based on RNA sequencing from differentiated cell lines and brain
tissues, it was demonstrated that numerous circRNAs are tissue- and
stage-specifically expressed (49,50).
Due to their unusual stability, highly conserved sequence and
expression specificity, circRNAs may serve as candidates for
clinical biomarker research.
Biological functions of circRNAs
A growing body of studies have revealed that
circRNAs can be produced in any region of the genome and can be
cycled by multiple cyclization modes (51–53).
Those features create multiformity in the sequence and length of
circRNAs, eventually resulting in the multiplicity of circRNA
functions. The biological functions of circRNAs in a wide variety
of human neoplasms include acting as miRNA sponges, interacting
with proteins, regulating gene splicing or transcription,
translating proteins, or peptide and epigenetic regulation. The
most typical role of circRNAs is to serve as miRNA sponges
(54–57). Mechanistically, miRNAs can block the
translation of target mRNA via complementary pairing with the
target mRNA 3′-untranslated region (UTR), which impacts the
stability of the target mRNA and modulates the expression in the
cell nucleus via combining with gene promoters (52,53).
However, circRNAs can suppress the expression of miRNAs via
adsorption and, simultaneously, influence the molecular level of
downstream target genes (54). Li
et al (55) reported that
circ_PSD3 may serve as a miR-637 sponge to modulate the expression
of downstream target genes in papillary thyroid carcinoma. In
addition, other circRNAs, such as circRNA_0005529 and circ-TGFBR2,
have been reported to interact with miR-527 and miR-107 to regulate
human gastric cancer cell proliferation and metastasis, and
nasopharyngeal carcinoma cell viability, respectively (58,59).
Notably, the same circRNA may even have the opposite function in
different tumor cells, through targeting different miRNAs. For
example, circ-FOXO3 has been reported to target miR-23a to inhibit
esophageal squamous cell cancer proliferation, but may also target
miR-29a-3p to promote prostate cancer progression (60,61).
Notably, numerous circRNAs that do not possess miRNA-binding sites
still have a wide range of biological functions, indicating that
circRNAs have their own additional regulatory functions. Previous
studies have revealed that circRNAs may interact with RBPs or other
proteins to form circRNA-protein complexes, which may impact the
function and mechanisms of action of binding proteins (62,63).
For example, circ-FOKX2 may integrate with Y-box binding protein 1
and heterogeneous nuclear ribonucleoprotein K to facilitate the
upregulation of the oncogenes NUF2 and pyridoxal kinase, ultimately
accelerating pancreatic ductal adenocarcinoma progression (64). This form of action of circRNAs has
also been reflected in the significant downregulation of
circ-DCUN1D4 in lung adenocarcinoma (65). Furthermore, hsa_circ_0008367 has
been shown to interact with the RBP ALKBH5, thus regulating
ferroptosis in hepatocellular carcinoma cells (66).
Beyond the aforementioned two functions, circRNAs
can also modulate gene transcription and alternative splicing
(67,68). Conn et al (69) demonstrated that a circRNA derived
from exon 6 of the SEPALLATA3 (SEP3) gene increased the abundance
of the homology exon-skipped alternative splicing variant and
combined with its homology DNA locus, thereby affecting the
splicing efficiency of SEP3 mRNA. Furthermore, previous studies
identified that EIciRNAs, such as circ-PAIP2 and circ-EIF3J, may
constitute complexes with U1 small nuclear ribonucleoprotein
(snRNP) via specific RNA-RNA interactions to shape EIciRNA-U1snRNP
complexes, which can interact with the polymerase II transcription
complex to promote transcription of the parental gene (70–72).
According to recent studies, numerous circRNAs have been identified
in the cytoplasm, indicating that they may be translated into
proteins (73,74). Further studies revealed that
cap-independent translation of circRNAs can occur via internal
ribosome entry site 119 or following the incorporation of the
m6A RNA modification in the 5′-UTR. Wu et al
(75) demonstrated that circ-SMO
encoded a fresh protein, SMO-193aa, which may drive the
tumorigenicity of glioma cells. In addition, Zheng et al
(76) revealed that circ-PPP1R12A
can carry an open reading frame, which coded a protein known as
circ-PPP1R12A-73aa. Mechanistically, circ-PPP1R12A-73aa facilitated
the progression of colon cancer via stimulating the Hippo-YAP
signal pathway.
circRNAs and glioma
Numerous studies have demonstrated that several
oncogenic and anti-oncogenic circRNAs are abnormally expressed in
human glioma compared with their expression levels in normal
tissues or tumor-adjacent tissues. For example, the expression
levels of circ-FBXW7 and circ-AKT3 were lower in glioma cell lines
and cancer tissues from patients with glioma (77,78).
By contrast, circ-CPA4 (31),
circ-CDC45 (32) and circ-POSTN
(34) have been revealed to be
highly expressed in glioma cell lines and cancer tissues from
patients with glioma. Mechanistically, these abnormally expressed
circRNAs may modulate the migration, apoptosis, proliferation,
invasion and angiogenesis of glioma, mainly through acting as a
sponge targeting miRNAs. The main examples have been summarized in
Table I. A meaningful role may be
played by circRNAs in the carcinogenesis of glioma. The following
subsections address the effect of these differentially expressed
circRNAs on human glioma.
 | Table I.Upregulated or downregulated
circRNAs, and their functions and mechanisms in glioma. |
Table I.
Upregulated or downregulated
circRNAs, and their functions and mechanisms in glioma.
| Authors, year | circRNA | Sample | Expression
change | Function | Possible
mechanism | (Refs.) |
|---|
| Yang et al,
2018 | circ-FBXW7 | Glioma tissues and
glioma cells | Down | Tumorigenicity
(−) |
circ-FBXW7/FBXW7-185aa |
(77) |
| Xia et al,
2019 | circ-AKT3 | Glioma tissues and
glioma cells | Down | Tumorigenicity
(−) |
circ-AKT3/AKT3-174aa |
(78) |
| Wu et al,
2021 | circ-SMO | Glioma tissues and
glioma cells | Up | Tumorigenicity
(+) |
circ-SMO/SMO-193aa |
(75) |
| Shi et al,
2019 | circ-0014359 | Glioma tissues and
glioma cells | Up | Proliferation
(+) |
circ-0014359/miR-153/p-AKTser473 |
(85) |
| Chen et al,
2020 | circ_PTN | Glioma cells | Up | Proliferation
(+) |
circ_PTN/miR-122/SOX6 |
(86) |
| Peng et al,
2019 | circCPA4 | Glioma tissues and
glioma cells | Up | Proliferation (+)
Prognosis |
circCPA4/let-7/CPA4 |
(31) |
| He et al,
2020 | circ-MAPK4 | Glioma tissues and
glioma cells | Up | Apoptosis (−) |
circ-MAPK4/miR-125a-3p/p-p38/MAPK |
(92) |
| Cao et al,
2019 |
hsa_circ_0037251 | Glioma cells | Up | Apoptosis (−) |
hsa_circ_0037251/miR-1229-3p/ mTOR |
(93) |
| Chi et al,
2019 | circRNA-104075 | Glioma cells | Up | Apoptosis (−) |
circRNA-104075/Bcl-9 |
(94) |
| He et al,
2019 | circ_002136 | Glioma cells | Up | Angiogenesis
(+) |
circ_002136/miR-138-5p/SOX13/SPON2 |
(99) |
| He et al,
2019 | circ-DICER1 | Glioma cells | Up | Angiogenesis
(+) |
circ-DICER1/miR-103a-3p
(miR-382-5p)/ZIC4 | (100) |
| Liu et a,
2020 | circ-ATXN1 | Glioma cells | Up | Angiogenesis
(+) |
circ-ATXN1/miR-526b-3p/MMP2(VEGFA) | (101) |
| Zuo et al,
2019 | circ-SMAD7 | Glioma tissues and
glioma cells | Up | Migration (+)
Invasion (+) |
circ-SMAD7/PCNA | (105) |
| Yang et al,
2019 | circ-POSTN | Glioma tissues and
glioma cells | Up | Migration (+)
Invasion (+) |
Circ-POSTN/miR-1205 |
(34) |
| Yi et al,
2019 |
hsa_circ_0034642 | Glioma tissues and
glioma cells | Up | Migration (+)
Invasion (+) |
hsa_circ_0034642/miR-1205/BATF3 | (106) |
| Su et al,
2019 | circDENND2A | Glioma tissues and
glioma cells | Up | Migration (+)
Invasion (+) |
circDENND2A/miR-625-5p | (107) |
| Qu et al,
2019 | circ_0079593 | Glioma tissues and
glioma cells | Up | Prognosis | circ_0079593 /
miR-182 (miR-433) | (111) |
| Liu et al,
2020 | circCDC45 | Glioma tissues and
glioma cells | Up | Prognosis | circCDC45/miR-516b
(miR-527) |
(32) |
| Hua et al,
2019 | circCEP128 | Glioma cells | Up | Drug
resistance |
circCEP128/miR-145-5p/ABCG2 | (115) |
| Zhao et al,
2019 | circATP8B4 | Glioma cells | Up |
Radiosensitivity |
circATP8B4/miR-766 | (116) |
| Ding et al,
2019 | circNFIX | Glioma tissues and
glioma cells | Up | Targeted
therapy |
circNFIX/miR-378e/RPN2 | (117) |
circRNAs regulate the tumorigenicity
of glioma cells
Numerous genetic studies on glioma have determined
that gene mutations can promote the tumorigenicity of glioma cells
(2,78,79).
In general, glioma-related genes can be separated into two classes:
Oncogenes and tumor suppressor genes. Activating tumor-promoting
genes or deactivating tumor suppressor genes may lead to the onset
and development of glioma (80,81).
Furthermore, previous research on human glioma specimens has
revealed that circRNAs can alter the expression of certain genes or
proteins by translating proteins or sponging miRNAs to facilitate
the tumorigenicity of glioma cells (82). For example, it was demonstrated that
circ-FBXW7, a circRNA derived from the FBXW7 gene, encoded the
protein FBXW7-185aa, which may suppress the tumorigenicity of
glioma cells. Further research revealed that FBXW7-185aa reduced
the half-life of c-Myc by antagonizing ubiquitin specific peptidase
28-induced c-Myc stability. Additionally, circ-FBXW7 and
FBXW7-185aa levels were reduced in glioblastoma clinical samples,
and the expression of circ-FBXW7 was positively associated with the
overall survival of patients with glioblastoma (77). Furthermore, Xia et al
(78) demonstrated that circ-AKT3
was expressed at low levels in glioblastoma tissues in contrast to
the expression in normal brain tissues. Furthermore, this previous
study reported that circ-AKT3 encodes a 174 amino acid (aa)
non-classical protein named AKT3-174aa. Overexpression of
AKT3-174aa suppressed the tumorigenicity of glioblastoma cells.
Notably, Wu et al (75)
revealed that circ-SMO, a circRNA derived from exons 3–6 of the SMO
gene, encoded protein SMO-193aa, which is known to be essential for
Hedgehog signaling activation in glioblastoma. Furthermore, in
vivo and in vitro experiments verified that SMO-193aa
may directly interact with SMO and enhance SMO cholesterol
modification, thus promoting the tumorigenicity of glioma.
From a genetic point of view, circRNAs are vital
modulators of tumor formation. During the development of glioma,
circRNAs are pivotal players in the glioma-associated gene changes
that cause malignant initiation and progression, as mentioned in
the present review. Therefore, the role of circRNAs in glioma is
considered a promising field for further investigation.
circRNAs activate and sustain
proliferative signaling
Normal brain tissues regulate the generation and
release of proliferative signals that instruct entrance into and
development via the cell growth-and-division cycle, thereby
assuring the maintenance of normal brain tissue architecture and
characteristics (83). Glioma cells
have been suggested to obtain the ability to induce and sustain
positively acting growth-stimulatory signals in various ways
(84). For example, Shi et
al (85) reported that
circ-0014359 exhibited high expression levels in glioma clinical
tissue samples and cells. In addition, it was found that
overexpression of circ-0014359 was associated with downregulation
of miR-153; circ-0014359 acted as a sponge for miR-153 to
facilitate glioma cell proliferation and was suggested to be
involved in the PI3K axis. Further investigation revealed that high
expression of miR-153 decreased the levels of phosphorylated
(p)-AKTser473, whereas inhibiting miR-153 could enhance the
expression of p-AKTser473. Chen et al (86) demonstrated that circ_ PTN exhibited
high expression levels in glioma, and functioned as an initiator of
cell proliferation via its sponging effects on the miR-122 family.
Additionally, SRY-box transcription factor (SOX)6 was predicted as
a target of miR-122, and high expression levels of miR-122 reduced
the expression levels of SOX6. Downregulation of miR-122 reversed
the tumor suppressor role of circ_PTN knockdown, whereas increased
expression of SOX6 affected the apoptosis and inhibition of cell
proliferation induced by miR-122 overexpression. In contrast to
normal brain tissues, Peng et al (31) characterized circ-CPA4 from 28,265
differently expressed circRNAs in glioma tissues. The authors
investigated the functional roles of circ-CPA4 and its downstream
gene. By prediction and validation, the authors verified that
circ-CPA4 interacted with let-7 and served as a sponge for let-7;
circ-CPA4 may absorb let-7 to modulate the levels of CPA4, thus
promoting the proliferation and progression of glioma cells
(31).
Dysregulation of circRNAs may contribute to the
aberrant functioning of proliferation signaling in glioma. As miRNA
sponges, circRNAs may suppress the expression levels of miRNAs via
adsorption and concomitantly affect the expression of downstream
genes. Thus, circRNAs may provide novel insights into controlling
the malignant proliferation of glioma cells.
circRNAs function in cell
apoptosis
Apoptosis is finely modulated at the gene level,
thus leading to the highly efficient elimination of injured cells
(87). The balance between anti-
and pro-apoptotic factors is essential to decide whether a cell
enters apoptosis. Apoptosis is mainly modulated via the caspase
family of proteases and the B-cell lymphoma 2 (Bcl-2) family of
intracellular proteins. Previous studies have demonstrated that
circRNAs are pivotal modulators of a number of cell processes in
glioma, including apoptosis (88,89).
circRNAs can alter the extrinsic and intrinsic apoptotic pathways
in several ways, resulting in a decrease in apoptosis or
development of apoptosis resistance; these involve the injured
death receptor pathway, alteration of the balance between
anti-apoptotic and pro-apoptotic proteins of the Bcl-2 family,
reduced caspase function and impaired p53 function (90,91).
He et al (92) demonstrated
that circ-MAPK4, a circRNA that has been reported to act as a
cancer-promoting gene in glioma, was upregulated and associated
with the clinicopathological stage of glioma (P<0.05). In
addition, this previous study revealed that knockdown of circ-MAPK4
resulted in increased levels of the cleaved form of apoptotic
factors [caspase-9, caspase-3 and poly(ADP-ribose) polymerase],
indicating the anti-apoptotic effect of circ-MAPK4 in glioma
development. In addition, experimental validation suggested that
circ-MAPK4 targeted miR-125a-3p, which activated the p38/MAPK
signaling pathway to modulate cell apoptosis. Another study by Cao
et al (93) revealed that
hsa_circ_0037251 modulated the levels of mechanistic target of
rapamycin through sponging miR-1229-3p and thus regulated cellular
apoptosis. Notably, hsa_circ_0037251 was revealed to be
overexpressed in glioma cells, and silencing of hsa_circ_0037251
facilitated cellular apoptosis and induced cell cycle arrest in the
G1 phase. It is becoming clear that destabilizing the
overall pro- or anti-apoptotic balance of the Bcl-2 family can
induce dysregulated apoptosis in the affected cells. For example,
Chi et al (94) reported
that matrine was able to induce apoptosis via downregulating
circRNA-104075 in glioma cells. circRNA-104075 was highly expressed
in glioma cells and functioned as a suppressor of cell apoptosis by
disrupting the equilibrium of anti-apoptotic/pro-apoptotic factors.
Furthermore, this previous study demonstrated that increased
expression of circRNA-104075 increased p62 and Bcl-2 expression,
but reduced beclin-1, cleaved caspase-3, cleaved caspase-9 and Bax
levels, as well as the light chain (LC)3-II/LC3-I ratio in
matrine-treated glioma cells.
circRNAs can alter the balance of anti- and
pro-apoptotic factors via multiple mechanisms, thus leading to
apoptotic resistance of glioma cells (89–94).
In addition, these findings may provide more evidence on the
association between cell apoptosis and circRNAs to better develop
novel therapeutic strategies for glioma.
circRNAs induce angiogenesis
Similar to normal tissues, tumor growth requires
oxygen and nutrients, along with an ability to excrete carbon
dioxide and metabolic waste products (95). Cancer cells excrete numerous
pro-angiogenic regulators, which result in the creation of abnormal
tumor-associated neovasculature. Tumor-associated neovasculature,
which is characterized by disordered, immature and penetrable blood
vessels, helps to maintain and expand tumor growth (96). Evidence has indicated that
angiogenesis is induced early during the multi-step progression of
glioma in humans and experimental animal models (97). Glioma cells excrete vascular
endothelial growth factor (VEGF) and other pro-angiogenic
regulatory factors to facilitate the development of vascular
endothelial cells. Several of these angiogenic members are
signaling proteins that couple with stimulating or suppressive cell
surface receptors exhibited by vascular endothelial cells. In
addition, glial vascular endothelial cells may excrete a
considerable quantity of factors that accelerate cancer development
(98). Notably, research has shown
that circRNAs can regulate the interactions of these secreted
factors to induce angiogenesis. He et al (99) indicated that circ_002136, spondin 2
(SPON2) and SOX13 were upregulated, whereas miR-138-5p was
downregulated in glioma-exposed endothelial cells. circ_002136
served as a molecular sponge for miR-138-5p, which negatively
regulated SOX13 and modulated glioma angiogenesis. By contrast,
SOX13 improved the level of SPON2 and enhanced the angiogenic
ability of glioma-exposed endothelial cells. Furthermore, silencing
circ_002136 markedly suppressed the activity, metastasis and tubule
formation of glioma-exposed endothelial cells. In another study, it
was demonstrated that the level of circ-DICER1 was increased in
glioma-exposed endothelial cells, and knockdown of circ-DICER1
inhibited the cell activity, metastasis and tubule-forming ability
of glioma-exposed endothelial cells. Furthermore, circ-DICER1
served as a sponge for miR-382-5p or miR-103a-3p, and impaired
their negative modulatory effects on zic family member 4 (ZIC4) in
glioma-exposed endothelial cells. Further research verified that
ZIC4 upregulated the level of its downstream gene heat shock
protein (Hsp)90β, and Hsp90 subsequently promoted the cell
activity, metastasis and tubule-forming ability of glioma-exposed
endothelial cells via stimulating the PI3K/AKT signaling pathway
(100). Additionally, Liu et
al (101) demonstrated that
the levels of circ-ATXN1were increased, whereas miR-526b-3p
expression was decreased in glioma-associated endothelial cells.
circ-ATXN1 promoted angiogenesis of glioma-associated endothelial
cells by interacting with miR-526b-3p. A dual luciferase experiment
elucidated that MMP2 and VEGFA were targets of miR-526b-3p.
Furthermore, silencing circ-ATXN1 inhibited the viability and
angiogenesis of glioma-associated endothelial cells.
Taken together, these findings indicated that
circRNAs are important regulators of angiogenesis in glioma.
circRNAs may regulate the interactions of pro-angiogenic factors
and glioma growth factors to induce angiogenesis. Angiogenesis is
the basis of hematogenous metastasis in glioma and new blood
vessels have been characterized as essential channels for glioma to
transport nutrients (97,98). These circRNA studies may be useful
in designing approaches to improving the survival of patients with
glioma.
circRNAs regulate invasion and
metastasis
Metastasis is generally a late event in cancer and
has limited treatment options. Invasion and migration are
associated with numerous phenotypic and genotypic changes of cells
that are indirectly and directly modulated via diverse extrinsic
and intrinsic factors (102). The
multiple-step procedures of invasion and migration have been
described as a series of discontinuous steps, usually referred to
as the invasion-metastasis cascade. High-grade malignant tumor
tissues are hyperactive and may progress through the steps of the
invasion-metastasis cascade within a short period of time (103). Glioma is an invasive disease that
tends to propagate locally and migrate to other parts of the brain
(104). It is increasingly
apparent that dysregulation of circRNAs may contribute to the
aggressive growth and migration of glioma. For example, circ-SMAD7
promoted the invasion and migration of glioma via upregulating
proliferating cell nuclear antigen (PCNA), whereas knockdown of
circ-SMAD7 suppressed the metastasis and invasion of glioma cells.
Consistently, the mRNA and protein expression levels of PCNA were
suppressed after circ-SMAD7 was downregulated in glioma cells. In
addition, the levels of circ-SMAD7 were increased in glioma tissues
compared with those in normal brain tissues. Furthermore, the
expression of circ-SMAD7 was associated with World Health
Organization tumor stage and Karnofsky Performance Scale score
(105). Another study by Yang
et al (34) revealed that
the levels of circ-POSTN were elevated in glioma and associated
with World Health Organization tumor grades, tumor dimension and
poor prognosis in patients with glioma. circ-POSTN has been shown
to act as a sponge of miR-1205 to accelerate glioma cell invasion
and metastasis in vivo. Additionally, knockdown of
circ-POSTN in glioma cells reduced cell metastasis and invasion
potential (34). Moreover, Yi et
al (106) demonstrated that
hsa_circ_0034642 affected the migration and invasion of glioma via
binding miR-1205 to regulate the basic leucine zipper transcription
factor ATF-like 3 level. Notably, hsa_circ_0034642 exhibited higher
levels of expression in glioma tissue and cells in comparison with
in normal brain tissue. Furthermore, high hsa_circ_0034642
expression was associated with clinical severity and a worse
prognosis. Su et al (107)
characterized a fresh hypoxia-related circRNA, DENN domain
containing 2A (DENND2A), which enhanced the invasion and metastasis
of glioma cells by directly binding to miR-625-5p. In addition,
glioma tissue with highly expressed hypoxia-inducible factor 1α
exhibited a higher level of circ-DENND2A and a lower level of
miR-625-5p.
Based on the aforementioned findings, circRNAs may
affect glioma invasion and metastasis through dynamic and complex
regulatory mechanisms. Invasion and metastasis of glioma cells are
leading factors resulting in patient death. Therefore, further
studies on the effect of circRNAs in glioma cell invasion and
metastasis are expected to help with the development of treatment
strategies for glioma.
circRNAs as diagnostic and prognostic
biomarkers in glioma
In recent years, despite substantial advances in
treatment strategy, the rate of early diagnosis of glioma remains
low and it is difficult to treat patients with late-stage glioma,
thus leading to the relative low survival of this disease (108). Additionally, compared with other
types of cancer, the overall prognosis of patients with glioma is
worse and the survival period is shorter (109). Thus, the identification of
valuable biomarkers for early-stage diagnosis and prognosis is
crucial for improving the survival of patients with glioma. Recent
studies have demonstrated that circRNAs are strongly associated
with the clinicopathological characteristics of glioma, and they
may serve as diagnostic and prognostic biomarkers (32,92,110).
Previous studies indicated that the closed-ring structure
stabilizes circRNAs in plasma and tissues, and protects them from
the enzymatic activity of exonucleases (28,50);
therefore, the unique structure of circRNAs may be a potential
advantage for their use as biomarkers for the diagnosis and
prognosis of glioma. Qu et al (111) revealed that circ_0079593 could be
considered an independent prognostic factor for patients with
glioma based on multivariate analysis. Furthermore, overexpression
of circ_0079593 in glioma was found to be associated with tumor
size, World Health Organization tumor grade and patient survival
rate. Mechanistically, circ_0079593 was revealed to facilitate
glioma development via binding to miR-433 and miR-182. Another
study by Liu et al (32)
indicated that circ-CDC45 expression was increased in glioma
compared with that in paired non-cancerous specimens.
Overexpression of circ-CDC45 was closely associated with larger
tumor size, higher World Health Organization tumor stage and poor
prognosis. Functionally, circ-CDC45 interacted with and modulated
the levels of miR-527 and miR-516b to facilitate cell proliferation
and invasion. Furthermore, Peng et al (31) demonstrated that the levels of
circ-CPA4 were high in glioma compared with those in paired
non-cancerous specimens. Simultaneously, a high level of circ-CPA4
was positively correlated with a worse outcome of glioma.
In conclusion, due to their diverse range of
functions, high stability and presence in easily obtainable body
fluids, circRNAs are considered valuable biomarkers in glioma.
Furthermore, the study of emerging circRNAs will improve our
understanding of the molecular pathways involved, including
glioma-associated gene expression, thereby improving the management
of patients with glioma.
Therapeutic potential of circRNAs in
glioma
Over the last decade, treatment for glioma has
focused on maximum safe surgical resection alongside radiotherapy
and chemotherapy (3). Radiotherapy
and chemotherapy are typical and aggressive treatment approaches
for numerous patients with late-stage and unresectable glioma;
however, the inherent and acquired resistance of glioma cells is a
major limitation to the efficacy of radiotherapy and chemotherapy
(112,113). Recently, given the enhanced
understanding of the etiopathogenesis of glioma, numerous signaling
pathways and molecules are considered potentially suitable for
targeted treatment (114). Recent
studies revealed that certain circRNAs can not only modulate the
resistance of glioma cells, but also block vital glioma-associated
signaling pathways and molecules, thereby modulating the expression
of the corresponding gene. For example, Hua et al (115) demonstrated that the levels of the
circ-CEP128 were higher, whereas miR-145-5p levels were lower in
temozolomide-resistant glioma cells compared with those in parental
cells. Silencing circ-CEP128 suppressed cellular proliferation,
reduced the level of ATP-binding cassette super-family G member 2
(ABCG2) and improved the cytotoxic function of temozolomide in
glioma cells. Furthermore, a mechanistic study demonstrated that
circ-CEP128 may serve as a miR-145-5p sponge to regulate the levels
of the downstream target ABCG2. Additionally, Zhao et al
(116) indicated that the levels
of circ-ATP8B4 were significantly higher in radiation-resistant
glioma cells, and indicated that circ-ATP8B4 may serve as a
molecular sponge to adsorb miR-766 and to facilitate cell
radioresistance. Furthermore, silencing or overexpression of
circRNAs not only elucidates the mechanism underlying the
initiation and development of glioma, but also provides useful
information for the design of targeted treatments to modulate
crucial glioma-associated signaling pathways, molecules and genes.
For example, Ding et al (117) indicated that the levels of
circ-NFIX were increased in glioma cells and tissues compared with
those in matched control samples, and high levels of circ-NFIX
markedly accelerated cell cycle progression, migration and
invasion, and suppressed apoptosis. By contrast, silencing
circ-NFIX led to cell cycle arrest, inhibition of cell migration
and invasion, and enhanced apoptosis in glioma cells.
Mechanistically, circ-NFIX positively regulated ribophorin II via
binding to miR-378e in glioma cells.
These results indicated that circRNAs may provide
novel insights into individualized therapeutic methods for the
treatment of glioma. An appropriate combination of novel targeted
and immunotherapeutic approaches that are circRNA-driven is
considered an attractive strategy for the treatment of human
glioma. However, compared with coding RNAs, the present knowledge
on circRNAs remains inadequate, and circRNAs cannot yet be used in
the clinical setting. Thus, further studies on the treatment of
glioma are needed.
Summary of the regulatory pathway of
circRNAs in glioma biological progression
In glioma, a considerable number of circRNAs are
dysregulated in neoplastic tissue, which may suppress or promote
tumor progression. Notably, the understanding of circRNAs is
rapidly improving, as described in the present review. In addition,
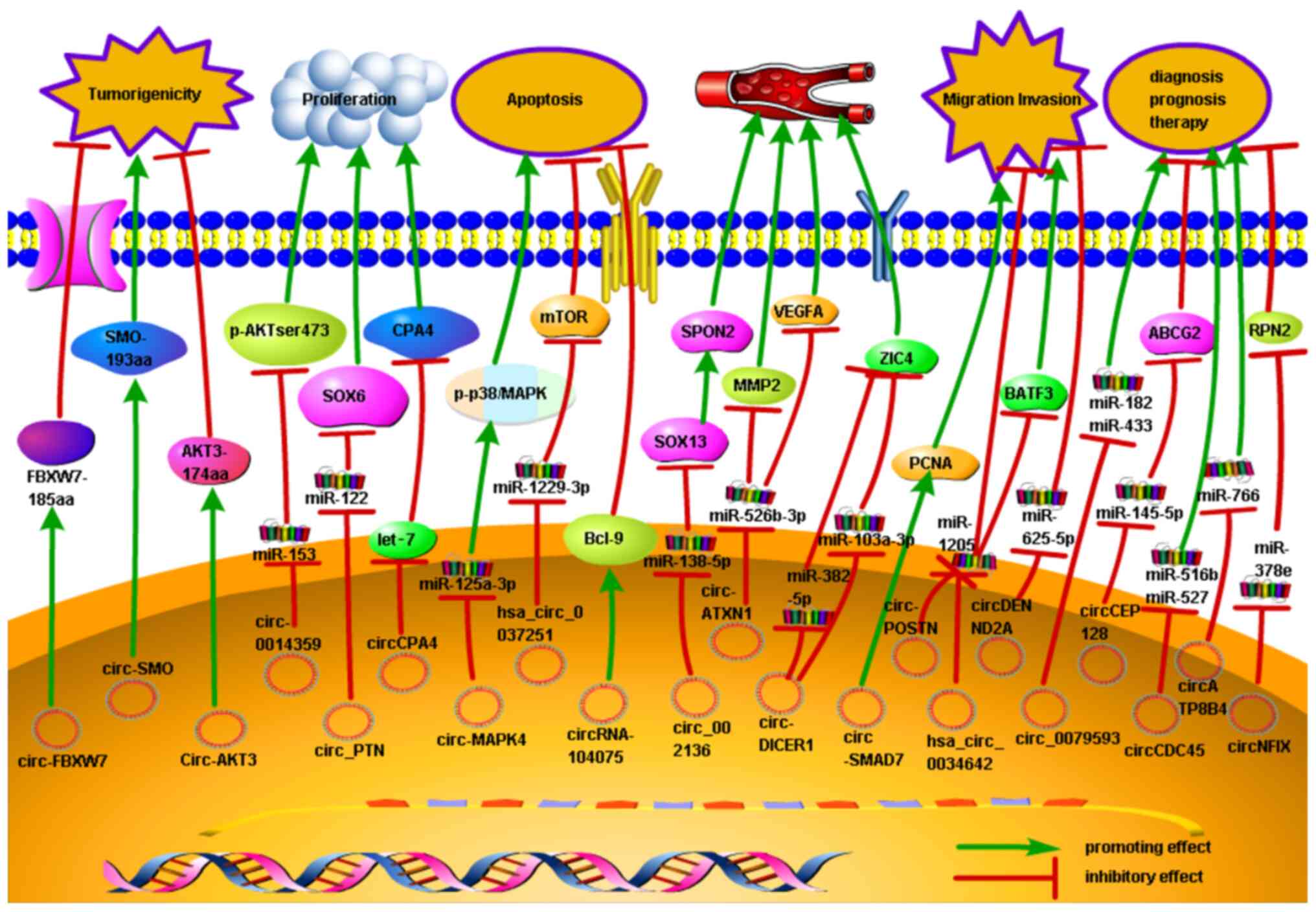
numerous studies (Fig. 2) have
indicated the role of circRNAs in several biological functions in
human glioma, including cellular proliferation, migration,
invasion, apoptosis and angiogenesis. For example, circ-0014359,
circ_PTN, circ-MAPK4 and hsa_circ_0037251 bind to miR-153, miR-122,
miR-125a-3p and miR-1229-3p, respectively, to modulate the
apoptosis and proliferation of glioma cells (85,86,92,93).
Furthermore, other circRNAs, such as circ_002136, hsa_circ_0034642
and circ-DENND2A, regulate the angiogenesis, invasion and migration
of glioma cells by acting as sponges targeting miR-138-5p, miR-1205
and miR-625-5p, respectively (99,106,107).
Online resources
Recently, to facilitate the study of the different
aspects of circRNAs, a number of circRNA research tools have been
developed and improved such as circBase, CircInteractome,
circAtlas, MiOncoCirc, CircR2Disease, TCSD and CSCD (118–124). These databases are currently used
for circRNA identification, localization and characterization
analysis, and as tools for investigating the interaction of
circRNAs with targets. Online resources are essential tools for
investigators to obtain data, and the number of them has rapidly
increased. Some of the online databases used for the study of
circRNAs are shown in Table
II.
| Table II.Databases used for circRNA
research. |
Table II.
Databases used for circRNA
research.
| Authors, year | Database | URL | Annotation | (Refs.) |
|---|
| Glazar et
al, 2014 | circBase | http://www.circbase.org/ | A circRNA database
that contains circRNA information from human, mouse, C.
elegans and Latimeria organisms | (118) |
| Panda et al,
2018 |
CircInteractome | http://circinteractome.nia.nih.gov/ | Predicts binding
sites of proteins to circRNAs. Predicts potential binding sites for
miRNA-circRNA interactions | (119) |
| Ji et al,
2019 | circAtlas | http://circatlas.biols.ac.cn | Includes circRNA
sequence information for humans and other animals | (120) |
| Vo et al,
2019 | MiOncoCirc | https://nguyenjoshvo.github.io/ | Description of
targets that can be used for cancer diagnosis or treatment | (121) |
| Fan et al,
2018 | CircR2Disease | http://bioinfo.snnu.edu.cn/CircR2Disease/ | This tool delivers
a broad database for circRNA dysregulation in different
diseases | (122) |
| Xia et al,
2017 | TSCD | http://gb.whu.edu.cn/TSCD | TSCD distinguishes
the characteristics of human and mouse tissue-specific
circRNAs | (123) |
| Xia et al,
2018 | CSCD | http://gb.whu.edu.cn/CSCD | This database
recognized 272,152 cancer-specific circRNAs from 228 total RNA
samples Predicts cellular localization of circRNAs, RNA-binding
proteins and variable splicing of related genes | (124) |
Conclusion and perspectives
Glioma is a malignancy of the human brain, which
poses a major threat to the health and survival of those affected.
The initiation and development of glioma include various phases and
regulators; however, its pathogenesis requires further
investigation. Currently, identifying novel molecules is crucial
for the ongoing progress in the research of targeted treatments for
glioma. Over the past few years, a large body of evidence has
indicated the effect of circRNAs on the development of glioma.
Databases have played a key role in bioinformatics analyses on
circRNAs and have contributed to considerable advances in this
field. The function of circRNAs includes a wide range of
physiopathological processes. As described in the present review,
circRNAs participate in glioma cell proliferation, invasion,
metastasis and cell cycle progression, and are crucial for the
diagnosis, prognosis and treatment of glioma. In addition, there
are several recommendations for future circRNA research. Firstly,
the present study on circRNAs in glioma mainly summarized studies
that used glioma tissues and cells, whereas other specimens (such
as cerebrospinal fluid and blood) are more easily obtained.
Secondly, the basic mechanisms through which circRNAs participate
in the advance of glioma await further investigation. Finally,
since the final objective of circRNA research is to safely utilize
circRNAs for clinical therapy of glioma, extensive clinical
research is required.
In conclusion, due to the advances in technology for
the assessment and screening of novel circRNAs, and the
improvements in databases, circRNAs may be applied in the future in
the diagnostic, therapeutic and prognostic management of glioma,
thereby leading to great advances in glioma therapy.
Acknowledgements
Not applicable.
Funding
The present study was supported by The Hebei
Province Key research and development projects of China (grant no.
19277737D).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WS performed the literature search and drafted the
manuscript. HZ conceived the idea for the article, and assisted
with editing and drafting the text. XH and LH assisted with editing
and drafting the text. XX participated in the critical review and
substantially revised it. Data authentication is not applicable.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ABCG2
|
ATP-binding cassette super-family G
member 2
|
|
Bcl-2
|
B-cell lymphoma 2
|
|
circRNAs
|
circular RNAs
|
|
ciRNAs
|
circular intronic RNAs
|
|
ecircRNAs
|
exonic circular RNAs
|
|
EIciRNAs
|
exon-intron circular RNAs
|
|
lncRNAs
|
long non-coding RNAs
|
|
miRNAs
|
microRNAs
|
|
ncRNAs
|
non-coding RNAs
|
References
|
1
|
Ng S, Zouaoui S, Bessaoud F, Rigau V, Roux
A, Darlix A, Bauchet F, Mathieu-Daudé H, Trétarre B,
Figarella-Branger D, et al: An epidemiology report for primary
central nervous system tumors in adolescents and young adults: A
nationwide population-based study in France, 2008–2013. Neuro
Oncol. 22:851–863. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sminia P, van den Berg J, van Kootwijk A,
Hageman E, Slotman BJ and Verbakel W: Experimental and clinical
studies on radiation and curcumin in human glioma. J Cancer Res
Clin Oncol. 147:403–409. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maccallini C, Gallorini M, Cataldi A and
Amoroso R: Targeting iNOS As a valuable strategy for the therapy of
glioma. ChemMedChem. 15:339–344. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Daisy Precilla S, Kuduvalli SS and
Thirugnanasambandhar Sivasubramanian A: Disentangling the
therapeutic tactics in GBM: From bench to bedside and beyond. Cell
Biol Int. 45:18–53. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Catalano M, D'Alessandro G, Trettel F and
Limatola C: Role of infiltrating microglia/macrophages in glioma.
Adv Exp Med Biol. 1202:281–298. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang L, Liu F, Weygant N, Zhang J, Hu P,
Qin Z, Yang J, Cheng Q, Fan F, Zeng Y, et al: A novel integrated
system using patient-derived glioma cerebral organoids and
xenografts for disease modeling and drug screening. Cancer Lett.
500:87–97. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ballo MT, Urman N, Lavy-Shahaf G, Grewal
J, Bomzon Z and Toms S: Correlation of tumor treating fields
dosimetry to survival outcomes in newly diagnosed glioblastoma: A
large-scale numerical simulation-based analysis of data from the
phase 3 EF-14 randomized trial. Int J Radiat Oncol Biol Phys.
104:1106–1113. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cloughesy TF, Mochizuki AY, Orpilla JR,
Hugo W, Lee AH, Davidson TB, Wang AC, Ellingson BM, Rytlewski JA,
Sanders CM, et al: Neoadjuvant anti-PD-1 immunotherapy promotes a
survival benefit with intratumoral and systemic immune responses in
recurrent glioblastoma. Nat Med. 25:477–486. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Roura AJ, Gielniewski B, Pilanc P,
Szadkowska P, Maleszewska M, Krol SK, Czepko R, Kaspera W, Wojtas B
and Kaminska B: Identification of the immune gene expression
signature associated with recurrence of high-grade gliomas. J Mol
Med (Berl). 99:241–255. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Eslahi M, Dana PM, Asemi Z, Hallajzadeh J,
Mansournia MA and Yousefi B: The effects of chitosan-based
materials on glioma: Recent advances in its applications for
diagnosis and treatment. Int J Biol Macromol. 168:124–129. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Pedretti S, Masini L, Turco E, Triggiani
L, Krengli M, Meduri B, Pirtoli L, Borghetti P, Pegurri L, Riva N,
et al: Hypofractionated radiation therapy versus chemotherapy with
temozolomide in patients affected by RPA class V and VI
glioblastoma: A randomized phase II trial. J Neurooncol.
143:447–455. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Saw PE, Xu X, Chen J and Song EW:
Non-coding RNAs: The new central dogma of cancer biology. Sci China
Life Sci. 64:22–50. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ryu J, Ahn Y, Kook H and Kim YK: The roles
of non-coding RNAs in vascular calcification and opportunities as
therapeutic targets. Pharmacol Ther. 218:1076752021. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu K, Yu M and Chen Y: Non-coding RNAs
regulating androgen receptor signaling pathways in prostate cancer.
Clin Chim Acta. 513:57–63. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Uddin MN and Wang X: The landscape of long
non-coding RNAs in tumor stroma. Life Sci. 264:1187252021.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fort V, Khelifi G and Hussein SMI: Long
non-coding RNAs and transposable elements: A functional
relationship. Biochim Biophys Acta Mol Cell Res. 1868:1188372021.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kalani M, Hodjati H, Ghoddusi Johari H and
Doroudchi M: Memory T cells of patients with abdominal aortic
aneurysm differentially expressed micro RNAs 21, 92a, 146a, 155,
326 and 663 in response to helicobacter pylori and lactobacillus
acidophilus. Mol Immunol. 130:77–84. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bahreini F, Rayzan E and Rezaei N:
microRNA-related single-nucleotide polymorphisms and breast cancer.
J Cell Physiol. 236:1593–1605. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jiang S: Perspectives on MicroRNA study in
oncogenesis: Where are we? Neoplasia. 23:99–101. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cao YZ, Sun JY, Chen YX, Wen CC and Wei L:
The roles of circRNAs in cancers: Perspectives from molecular
functions. Gene. 767:1451822021. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu Z, Huang Q, Zhang Q, Wu H and Zhong Z:
circRNAs open a new era in the study of cardiovascular disease
(review). Int J Mol Med. 47:49–64. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zaiou M: circRNAs signature as potential
diagnostic and prognostic biomarker for diabetes mellitus and
related cardiovascular complications. Cells. 9:6592020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li L, Sun D, Li X, Yang B and Zhang W:
Identification of key circRNAs in non-small cell lung cancer. Am J
Med Sci. 361:98–105. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Nguyen TM, Zhang XO, Wang L, Phan
T, Clohessy JG and Pandolfi PP: Optimized RNA-targeting
CRISPR/Cas13d technology outperforms shRNA in identifying
functional circRNAs. Genome Biol. 22:412021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Koch L: CRISPR-Cas13 targets circRNAs. Nat
Rev Genet. 22:682021. View Article : Google Scholar
|
|
26
|
Li Y, Feng W, Kong M, Liu R, Wu A, Shen L,
Tang Z and Wang F: Exosomal circRNAs: A new star in cancer. Life
Sci. 269:1190392021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Luo Q, Li X, Fu B, Zhang L, Fang L, Qing
C, Guo Y, Huang Z and Li J: Expression profile and diagnostic value
of circRNAs in peripheral blood from patients with systemic lupus
erythematosus. Mol Med Rep. 23:12021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang X, Ye T, Liu H, Lv P, Duan C, Wu X,
Jiang K, Lu H, Xia D, Peng E, et al: Expression profiles,
biological functions and clinical significance of circRNAs in
bladder cancer. Mol Cancer. 20:42021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu T, Wang M, Jiang L, Ma L, Wan L, Chen
Q, Wei C and Wang Z: circRNAs in anticancer drug resistance: Recent
advances and future potential. Mol Cancer. 19:1272020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang X, Yang H, Zhao L, Li G and Duan Y:
Circular RNA PRKCI promotes glioma cell progression by inhibiting
microRNA-545. Cell Death Dis. 10:6162019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peng H, Qin C, Zhang C, Su J, Xiao Q, Xiao
Y, Xiao K and Liu Q: circCPA4 acts as a prognostic factor and
regulates the proliferation and metastasis of glioma. J Cell Mol
Med. 23:6658–6665. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu J, Hou K, Ji H, Mi S, Yu G, Hu S and
Wang J: Overexpression of circular RNA circ-CDC45 facilitates
glioma cell progression by sponging miR-516b and miR-527 and
predicts an adverse prognosis. J Cell Biochem. 121:690–697. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Meng Q, Li S, Liu Y, Zhang S, Jin J, Zhang
Y, Guo C, Liu B and Sun Y: Circular RNA circSCAF11 accelerates the
glioma tumorigenesis through the miR-421/SP1/VEGFA axis. Mol Ther
Nucleic Acids. 17:669–677. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang Y, Zhang Y, Chen B, Ding L, Mu Z and
Li Y: Elevation of circular RNA circ-POSTN facilitates cell growth
and invasion by sponging miR-1205 in glioma. J Cell Biochem.
120:16567–16574. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zheng SQ, Qi Y, Wu J, Zhou FL, Yu H, Li L,
Yu B, Chen XF and Zhang W: CircPCMTD1 acts as the sponge of
miR-224-5p to promote glioma progression. Front Oncol. 9:3982019.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sanger HL, Klotz G, Riesner D, Gross HJ
and Kleinschmidt AK: Viroids are single-stranded covalently closed
circular RNA molecules existing as highly base-paired rod-like
structures. Proc Natl Acad Sci USA. 73:3852–3856. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yin S, Tian X, Zhang J, Sun P and Li G:
PCirc: Random forest-based plant circRNA identification software.
BMC Bioinformatics. 22:102021. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Goodall GJ and Wickramasinghe VO: RNA in
cancer. Nat Rev Cancer. 21:22–36. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang D, Ni N, Wang Y, Tang Z, Gao H, Ju
Y, Sun N, He X, Gu P and Fan X: circRNA-vgll3 promotes osteogenic
differentiation of adipose-derived mesenchymal stem cells via
modulating miRNA-dependent integrin α5 expression. Cell Death
Differ. 28:283–302. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang J, Liu Y and Shi G: The
circRNA-miRNA-mRNA regulatory network in systemic lupus
erythematosus. Clin Rheumatol. 40:331–339. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen J, Yang J, Fei X, Wang X and Wang K:
circRNA ciRS-7: A novel oncogene in multiple cancers. Int J Biol
Sci. 17:379–389. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cao Z, Zhang Y, Wei S, Zhang X, Guo Y and
Han B: Comprehensive circRNA expression profile and function
network in osteoblast-like cells under simulated microgravity.
Gene. 764:1451062021. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu T, Li Y, Liang X, Liu X and Tang M:
Identification of potential circRNA-miRNA-mRNA regulatory networks
in response to graphene quantum dots in microglia by microarray
analysis. Ecotoxicol Environ Saf. 208:1116722021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wang Z and Lei X: Identifying the sequence
specificities of circRNA-binding proteins based on a capsule
network architecture. BMC Bioinformatics. 22:192021. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhou B, Yang H, Yang C, Bao YL, Yang SM,
Liu J and Xiao YF: Translation of noncoding RNAs and cancer. Cancer
Lett. 497:89–99. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fu LY, Wang SW, Hu MY, Jiang ZL, Shen LL,
Zhou YP, Guo JM and Hu YR: Circular RNAs in liver diseases:
Mechanisms and therapeutic targets. Life Sci. 264:1187072021.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wei G, Zhu J, Hu HB and Liu JQ: Circular
RNAs: Promising biomarkers for cancer diagnosis and prognosis.
Gene. 771:1453652021. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Varela-Martinez E, Corsi GI, Anthon C,
Gorodkin J and Jugo BM: Novel circRNA discovery in sheep shows
evidence of high backsplice junction conservation. Sci Rep.
11:4272021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li Z, Li X, Xu D, Chen X, Li S, Zhang L,
Chan MTV and Wu WKK: An update on the roles of circular RNAs in
osteosarcoma. Cell Prolif. 54:e129362021. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Beilerli A, Gareev I, Beylerli O, Yang G,
Pavlov V, Aliev G and Ahmad A: Circular RNAs as biomarkers and
therapeutic targets in cancer. Semin Cancer Biol. Jan 9–2021.(Epub
ahead of print). doi: 10.1016/j.semcancer.2020.12.026. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang X, Cheng Z, Xu J, Feng M, Zhang H,
Zhang L and Qian L: Circular RNA Arhgap12 modulates
doxorubicin-induced cardiotoxicity by sponging miR-135a-5p. Life
Sci. 265:1187882021. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Khanipouyani F, Akrami H and Fattahi MR:
Circular RNAs as important players in human gastric cancer. Clin
Transl Oncol. 23:10–21. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Luo Y and Huang C: circSFMBT2 facilitates
vascular smooth muscle cell proliferation by targeting
miR-331-3p/HDAC5. Life Sci. 264:1186912021. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Razavi ZS, Tajiknia V, Majidi S, Ghandali
M, Mirzaei HR, Rahimian N, Hamblin MR and Mirzaei H: Gynecologic
cancers and non-coding RNAs: Epigenetic regulators with emerging
roles. Crit Rev Oncol Hematol. 157:1031922021. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li Z, Huang X, Liu A, Xu J, Lai J, Guan H
and Ma J: circ_PSD3 promotes the progression of papillary thyroid
carcinoma via the miR-637/HEMGN axis. Life Sci. 264:1186222021.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang XY and Mao L: Circular RNA
circ_0000442 acts as a sponge of miR-148b-3p to suppress breast
cancer via PTEN/PI3K/Akt signaling pathway. Gene. 766:1451132021.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Sun Q, Qi X, Zhang W and Li X: Knockdown
of circRNA_0007534 suppresses the tumorigenesis of cervical cancer
via miR-206/GREM1 axis. Cancer Cell Int. 21:542021. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang X, Yang H, Jia Y, Xu Z, Zhang L, Sun
M and Fu J: circRNA_0005529 facilitates growth and metastasis of
gastric cancer via regulating miR-527/Sp1 axis. BMC Mol Cell Biol.
22:62021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li W, Lu H, Wang H, Ning X, Liu Q, Zhang
H, Liu Z, Wang J, Zhao W, Gu Y, et al: Circular RNA TGFBR2 acts as
a ceRNA to suppress nasopharyngeal carcinoma progression by
sponging miR-107. Cancer Lett. 499:301–313. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Xing Y, Zha WJ, Li XM, Li H, Gao F, Ye T,
Du WQ and Liu YC: Circular RNA circ-Foxo3 inhibits esophageal
squamous cell cancer progression via the miR-23a/PTEN axis. J Cell
Biochem. 121:2595–2605. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kong Z, Wan X, Lu Y, Zhang Y, Huang Y, Xu
Y, Liu Y, Zhao P, Xiang X, Li L and Li Y: Circular RNA circFOXO3
promotes prostate cancer progression through sponging miR-29a-3p. J
Cell Mol Med. 24:799–813. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Zang J, Lu D and Xu A: The interaction of
circRNAs and RNA binding proteins: An important part of circRNA
maintenance and function. J Neurosci Res. 98:87–97. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Okholm TLH, Sathe S, Park SS, Kamstrup AB,
Rasmussen AM, Shankar A, Chua ZM, Fristrup N, Nielsen MM, Vang S,
et al: Transcriptome-wide profiles of circular RNA and RNA-binding
protein interactions reveal effects on circular RNA biogenesis and
cancer pathway expression. Genome Med. 12:1122020. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wong CH, Lou UK, Li Y, Chan SL, Tong JH,
To KF and Chen Y: circFOXK2 promotes growth and metastasis of
pancreatic ductal adenocarcinoma by complexing with RNA-binding
proteins and sponging miR-942. Cancer Res. 80:2138–2149. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Liang Y, Wang H, Chen B, Mao Q, Xia W,
Zhang T, Song X, Zhang Z, Xu L, Dong G and Jiang F: circDCUN1D4
suppresses tumor metastasis and glycolysis in lung adenocarcinoma
by stabilizing TXNIP expression. Mol Ther Nucleic Acids.
23:355–368. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liu Z, Wang Q, Wang X, Xu Z, Wei X and Li
J: Circular RNA cIARS regulates ferroptosis in HCC cells through
interacting with RNA binding protein ALKBH5. Cell Death Discov.
6:722020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Liu Y, Liu X, Lin C, Jia X, Zhu H, Song J
and Zhang Y: Noncoding RNAs regulate alternative splicing in
cancer. J Exp Clin Cancer Res. 40:112021. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Stagsted LVW, O'Leary ET, Ebbesen KK and
Hansen TB: The RNA-binding protein SFPQ preserves long-intron
splicing and regulates circRNA biogenesis in mammals. Elife.
10:e630882021. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Conn VM, Hugouvieux V, Nayak A, Conos SA,
Capovilla G, Cildir G, Jourdain A, Tergaonkar V, Schmid M, Zubieta
C and Conn SJ: A circRNA from SEPALLATA3 regulates splicing of its
cognate mRNA through R-loop formation. Nat Plants. 3:170532017.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Hu Q and Zhou T: EIciRNA-mediated gene
expression: Tunability and bimodality. FEBS Lett. 592:3460–3471.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kong S, Tao M, Shen X and Ju S:
Translatable circRNAs and lncRNAs: Driving mechanisms and functions
of their translation products. Cancer Lett. 483:59–65. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Lei M, Zheng G, Ning Q, Zheng J and Dong
D: Translation and functional roles of circular RNAs in human
cancer. Mol Cancer. 19:302020. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Wu X, Xiao S, Zhang M, Yang L, Zhong J, Li
B, Li F, Xia X, Li X, Zhou H, et al: A novel protein encoded by
circular SMO RNA is essential for Hedgehog signaling activation and
glioblastoma tumorigenicity. Genome Biol. 22:332021. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Zheng X, Chen L, Zhou Y, Wang Q, Zheng Z,
Xu B, Wu C, Zhou Q, Hu W, Wu C and Jiang J: A novel protein encoded
by a circular RNA circPPP1R12A promotes tumor pathogenesis and
metastasis of colon cancer via Hippo-YAP signaling. Mol Cancer.
18:472019. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Yang Y, Gao X, Zhang M, Yan S, Sun C, Xiao
F, Huang N, Yang X, Zhao K, Zhou H, et al: Novel Role of FBXW7
circular RNA in repressing glioma tumorigenesis. J Natl Cancer
Inst. 110:304–315. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Xia X, Li X, Li F, Wu X, Zhang M, Zhou H,
Huang N, Yang X, Xiao F, Liu D, et al: A novel tumor suppressor
protein encoded by circular AKT3 RNA inhibits glioblastoma
tumorigenicity by competing with active phosphoinositide-dependent
Kinase-1. Mol Cancer. 18:1312019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
El-Hashash AHK: Histone H3K27M mutation in
brain tumors. Adv Exp Med Biol. 1283:43–52. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
He Z, Yang C, He Y, Gong B, Yin C, Feng J,
Chen L, Tang J and Chen Y: CAMTA1, a novel antitumor gene,
regulates proliferation and the cell cycle in glioma by inhibiting
AKT phosphorylation. Cell Signal. 79:1098822021. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Osama M, Mostafa MN and Alvi MA: Astrocyte
elevated gene-1 as a novel therapeutic target in malignant gliomas
and its interactions with oncogenes and tumor suppressor genes.
Brain Res. 1747:1470342020. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhang Y, Liu Q and Liao Q: CircHIPK3: A
promising cancer-related circular RNA. Am J Transl Res.
12:6694–6704. 2020.PubMed/NCBI
|
|
83
|
Smyth LCD, Rustenhoven J, Scotter EL,
Schweder P, Faull RLM, Park TIH and Dragunow M: Markers for human
brain pericytes and smooth muscle cells. J Chem Neuroanat.
92:48–60. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Jiang Y, Zhou J, Luo P, Gao H, Ma Y, Chen
YS, Li L, Zou D, Zhang Y and Jing Z: Prosaposin promotes the
proliferation and tumorigenesis of glioma through toll-like
receptor 4 (TLR4)-mediated NF-κB signaling pathway. EBioMedicine.
37:78–90. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Shi F, Shi Z, Zhao Y and Tian J: circRNA
hsa-circ-0014359 promotes glioma progression by regulating
miR-153/PI3K signaling. Biochem Biophys Res Commun. 510:614–620.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Chen C, Deng L, Nie DK, Jia F, Fu LS, Wan
ZQ and Lan Q: Circular RNA Pleiotrophin promotes carcinogenesis in
glioma via regulation of microRNA-122/SRY-box transcription factor
6 axis. Eur J Cancer Prev. 29:165–173. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ye Z, Zhuo Q, Hu Q, Xu X, Liu M, Zhang Z,
Xu W, Liu W, Fan G, Qin Y, et al: FBW7-NRA41-SCD1 axis
synchronously regulates apoptosis and ferroptosis in pancreatic
cancer cells. Redox Biol. 38:1018072021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Mohamad Anuar NN, Nor Hisam NS, Liew SL
and Ugusman A: Clinical review: Navitoclax as a pro-apoptotic and
anti-fibrotic agent. Front Pharmacol. 11:5641082020. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Hartman ML and Czyz M: BCL-w: Apoptotic
and non-apoptotic role in health and disease. Cell Death Dis.
11:2602020. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yan F, Fan B, Wang J, Wei W, Tang Q, Lu L,
Luo Z, Pu J and Yang SS: circ_0008305-mediated miR-660/BAG5 axis
contributes to hepatocellular carcinoma tumorigenesis. Cancer Med.
10:833–842. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Lou J, Hao Y, Lin K, Lyu Y, Chen M, Wang
H, Zou D, Jiang X, Wang R, Jin D, et al: Circular RNA CDR1as
disrupts the p53/MDM2 complex to inhibit gliomagenesis. Mol Cancer.
19:1382020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
He J, Huang Z, He M, Liao J, Zhang Q, Wang
S, Xie L, Ouyang L, Koeffler HP, Yin D and Liu A: Circular RNA
MAPK4 (circ-MAPK4) inhibits cell apoptosis via MAPK signaling
pathway by sponging miR-125a-3p in gliomas. Mol Cancer. 19:172020.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Cao Q, Shi Y, Wang X, Yang J, Mi Y, Zhai G
and Zhang M: Circular METRN RNA hsa_circ_0037251 promotes glioma
progression by sponging miR-1229-3p and regulating mTOR expression.
Sci Rep. 9:197912019. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Chi G, Xu D, Zhang B and Yang F: Matrine
induces apoptosis and autophagy of glioma cell line U251 by
regulation of circRNA-104075/BCL-9. Chem Biol Interact.
308:198–205. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Zheng X, Liu J, Li X, Tian R, Shang K,
Dong X and Cao B: Angiogenesis is promoted by exosomal DPP4 derived
from 5-fluorouracil-resistant colon cancer cells. Cancer Lett.
497:190–201. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Yetkin-Arik B, Kastelein AW, Klaassen I,
Jansen CHJR, Latul YP, Vittori M, Biri A, Kahraman K, Griffioen AW,
Amant F, et al: Angiogenesis in gynecological cancers and the
options for anti-angiogenesis therapy. Biochim Biophys Acta Rev
Cancer. 1875:1884462021. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Ahir BK, Engelhard HH and Lakka SS: Tumor
development and angiogenesis in adult brain tumor: Glioblastoma.
Mol Neurobiol. 57:2461–2478. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Carlson JC, Cantu-Gutierrez M, Lozzi B,
Huang-Hobbs E, Turner WD, Tepe B, Zhang Y, Herman AM, Rao G,
Creighton CJ, et al: Identification of diverse tumor endothelial
cell populations in malignant glioma. Neuro Oncol. 23:932–944.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
He Z, Ruan X, Liu X, Zheng J, Liu Y, Liu
L, Ma J, Shao L, Wang D, Shen S, et al:
FUS/circ_002136/miR-138-5p/SOX13 feedback loop regulates
angiogenesis in glioma. J Exp Clin Cancer Res. 38:652019.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
He Q, Zhao L, Liu X, Zheng J, Liu Y, Liu
L, Ma J, Cai H, Li Z and Xue Y: MOV10 binding circ-DICER1 regulates
the angiogenesis of glioma via miR-103a-3p/miR-382-5p mediated ZIC4
expression change. J Exp Clin Cancer Res. 38:92019. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Liu X, Shen S, Zhu L, Su R, Zheng J, Ruan
X, Shao L, Wang D, Yang C and Liu Y: SRSF10 inhibits biogenesis of
circ-ATXN1 to regulate glioma angiogenesis via miR-526b-3p/MMP2
pathway. J Exp Clin Cancer Res. 39:1212020. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Novikov NM, Zolotaryova SY, Gautreau AM
and Denisov EV: Mutational drivers of cancer cell migration and
invasion. Br J Cancer. 124:102–114. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Yu-Ju Wu C, Chen CH, Lin CY, Feng LY, Lin
YC, Wei KC, Huang CY, Fang JY and Chen PY: CCL5 of
glioma-associated microglia/macrophages regulates glioma migration
and invasion via calcium-dependent matrix metalloproteinase 2.
Neuro Oncol. 22:253–266. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Li X and Meng Y: Analyses of
metastasis-associated genes in IDH wild-type glioma. BMC Cancer.
20:11142020. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Zuo CY, Qian W, Huang CJ and Lu J:
Circular RNA circ-SMAD7 promoted glioma cell proliferation and
metastasis by upregulating PCNA. Eur Rev Med Pharmacol Sci.
23:10035–10041. 2019.PubMed/NCBI
|
|
106
|
Yi C, Li H, Li D, Qin X, Wang J, Liu Y,
Liu Z and Zhang J: Upregulation of circular RNA circ_0034642
indicates unfavorable prognosis in glioma and facilitates cell
proliferation and invasion via the miR-1205/BATF3 axis. J Cell
Biochem. 120:13737–13744. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Su H, Zou D, Sun Y and Dai Y:
Hypoxia-associated circDENND2A promotes glioma aggressiveness by
sponging miR-625-5p. Cell Mol Biol Lett. 24:242019. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Patil V and Mahalingam K: A four-protein
expression prognostic signature predicts clinical outcome of
lower-grade glioma. Gene. 679:57–64. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Choi J, Kim SH, Ahn SS, Choi HJ, Yoon HI,
Cho JH, Roh TH, Kang SG, Chang JH and Suh CO: Extent of resection
and molecular pathologic subtype are potent prognostic factors of
adult WHO grade II glioma. Sci Rep. 10:20862020. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Zheng K, Xie H, Wu W, Wen X, Zeng Z and
Shi Y: circRNA PIP5K1A promotes the progression of glioma through
upregulation of the TCF12/PI3K/AKT pathway by sponging miR-515-5p.
Cancer Cell Int. 21:272021. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Qu Y, Zhu J, Liu J and Qi L: Circular RNA
circ_0079593 indicates a poor prognosis and facilitates cell growth
and invasion by sponging miR-182 and miR-433 in glioma. J Cell
Biochem. 120:18005–18013. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Ghiaseddin AP, Shin D, Melnick K and Tran
DD: Tumor treating fields in the management of patients with
malignant gliomas. Curr Treat Options Oncol. 21:762020. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Oldrini B, Vaquero-Siguero N, Mu Q, Kroon
P, Zhang Y, Galán-Ganga M, Bao Z, Wang Z, Liu H, Sa JK, et al: MGMT
genomic rearrangements contribute to chemotherapy resistance in
gliomas. Nat Commun. 11:38832020. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Oprita A, Baloi SC, Staicu GA, Alexandru
O, Tache DE, Danoiu S, Micu ES and Sevastre AS: Updated insights on
EGFR signaling pathways in glioma. Int J Mol Sci. 22:5872021.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Hua L, Huang L, Zhang X, Feng H and Shen
B: Knockdown of circular RNA CEP128 suppresses proliferation and
improves cytotoxic efficacy of temozolomide in glioma cells by
regulating miR-145-5p. Neuroreport. 30:1231–1238. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zhao M, Xu J, Zhong S, Liu Y, Xiao H, Geng
L and Liu H: Expression profiles and potential functions of
circular RNAs in extracellular vesicles isolated from
radioresistant glioma cells. Oncol Rep. 41:1893–1900.
2019.PubMed/NCBI
|
|
117
|
Ding C, Wu Z, You H, Ge H, Zheng S, Lin Y,
Wu X, Lin Z and Kang D: CircNFIX promotes progression of glioma
through regulating miR-378e/RPN2 axis. J Exp Clin Cancer Res.
38:5062019. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Glazar P, Papavasileiou P and Rajewsky N:
circBase: A database for circular RNAs. RNA. 20:1666–1670. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Panda AC, Dudekula DB, Abdelmohsen K and
Gorospe M: Analysis of circular RNAs using the web tool
circinteractome. Methods Mol Biol. 1724:43–56. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Ji P, Wu W, Chen S, Zheng Y, Zhou L, Zhang
J, Cheng H, Yan J, Zhang S, Yang P and Zhao F: Expanded expression
landscape and prioritization of circular RNAs in mammals. Cell Rep.
26:3444–3460.e5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Vo JN, Cieslik M, Zhang Y, Shukla S, Xiao
L, Zhang Y, Wu YM, Dhanasekaran SM, Engelke CG, Cao X, et al: The
landscape of circular RNA in cancer. Cell. 176:869–881.e13. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Fan C, Lei X, Fang Z, Jiang Q and Wu FX:
CircR2Disease: A manually curated database for experimentally
supported circular RNAs associated with various diseases. Database
(Oxford). 2018:bay0442018. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Xia S, Feng J, Lei L, Hu J, Xia L, Wang J,
Xiang Y, Liu L, Zhong S, Han L and He C: Comprehensive
characterization of tissue-specific circular RNAs in the human and
mouse genomes. Brief Bioinform. 18:984–992. 2017.PubMed/NCBI
|
|
124
|
Xia S, Feng J, Chen K, Ma Y, Gong J, Cai
F, Jin Y, Gao Y, Xia L, Chang H, et al: CSCD: A database for
cancer-specific circular RNAs. Nucleic Acids Res. 46(D1):
D925–D929. 2018. View Article : Google Scholar : PubMed/NCBI
|