Introduction
As a major constituent of the vascular system,
vascular smooth muscle cells (VSMCs) crucially influence the
processes of sustaining vascular structure and blood pressure
(1,2). The ectopic proliferation and migration
of VSMCs have been demonstrated to be implicated in the development
of various cardiovascular diseases, including pulmonary artery
hypertension, vein graft failure and atherosclerosis (3,4). Under
normal circumstances, VSMCs remain in a non-proliferative and
non-migrating states (5). However,
the proliferation and migration of VSMCs can be markedly elevated
in the presence of extracellular factors, including basic
fibroblast growth factor, tumor necrosis factor-α and
platelet-derived growth factor (PDGF)-BB (6,7). Of
these, PDGF-BB, a well-known potent mitogen, contributes
effectively to increase VSMC proliferation and migration (8). Investigating the associated regulatory
mechanisms underlying VSMC proliferation may be useful for the
prevention of cardiovascular diseases. Inhibition of HSF1-FAM3A-ATP
signaling pathway in VSMC can alleviate Ang II (angiotensin
II)-induced constriction and vascular remodeling, hypertension, and
cardiac hypertrophy in mice, thus HSF1 inhibitors may be used to
treat hypertension (9). Dysfunction
of VSMCs is crucial in the pathogenesis of proliferative
cardiovascular diseases, and knockdown of miR-146a could
significantly inhibit the proliferative and migratory properties of
VSMCs in vitro via regulating NF-κB expression (10). VSMCs lose differentiation markers
and gain uncontrolled proliferative activity during the early
stages of atherosclerosis, and in response to vascular injury or
alterations in local environmental cues, differentiated/contractile
VSMCs are capable of switching to a dedifferentiated phenotype
characterized by increased proliferation, migration and
extracellular matrix synthesis together with decreased expression
of contractile markers (11).
MicroRNAs (miRNAs or miRs) are small non-coding RNAs
that regulate genes by binding to the 3′-untranslated regions
(3′-UTRs) of target mRNA (9).
Accumulating research has indicated that miRNAs exert pivotal
effects in the development and occurrence of vascular diseases
(12). For instance, miR-24 was
reported to attenuate vascular remodeling in high-glucose
(HG)-induced VSMCs by inhibiting their proliferation and migration
(13). miR-19 was identified to
serve protective effects against myocardial infarction (14). Additionally, miR-130a was
demonstrated to modulate the proliferation and migration of VSMCs
in various vascular tissues (15).
The results of these previous studies revealed that miRNAs are
crucial regulators of vascular function maintenance. Recently,
miR-125a-5p has been demonstrated to be implicated in the
progression of multiple regulatory processes, including VSMCs
proliferation (16). miR-125b is
involved in vascular calcification in vitro and in
vivo by, at least partly, targeting SP7 (17). miR-125a-5p is highly expressed in
VSMCs and inhibits the PDGF-BB pathway by targeting E26
transformation-specific (ETS)-1 and is, therefore, a potential
regulator of the phenotypic switch of VSMCs (16). It has been reported that miR-7 is
closely correlated with biological processes (18). miR-7 was reported to regulate
autophagy and the ubiquitin-proteasome system in human muscle
cells. Low levels of miR-7 promoted both processes and high levels
of miR-7 repressed them. Furthermore, miR-7 was demonstrated to
inhibit tumor metastasis and reverse epithelial-mesenchymal
transition through protein kinase/extracellular signal-regulated
kinase (ERK)1/2 inactivation by targeting epidermal growth factor
receptor (EGFR) in epithelial ovarian cancer (18). However, the possible roles of
miR-125a-5p and miR-7 in VSMCs growth and migration remain to be
elucidated.
EGFR, a member of ErbB receptor family, is
extensively distributed in human tissues (15). EGFR overexpression is associated
with diverse solid tumors, including ovarian cancer and colorectal
cancer (19). EGFR has been
demonstrated to activate the ERK pathway, which manages cellular
processes, such as cell growth and differentiation (20). More importantly, EGFR has been
indicated to influence the proliferation and migration of VSMCs
(21). However, it is unclear
whether EGFR is affected by miR-125a-5p and miR-7 in VSMCs.
The current study investigated the functional role
of miR-125a-5p or miR-7 on cell growth, migration and invasion in
VSMCs treated by PDGF-BB. The present study hypothesized that
miR-125a-5p and miR-7 were involved in the proliferation and
migration of VSMCs by targeting EGFR. Therefore, the current study
aimed to establish the functional roles of miR-125a-5p and miR-7 on
the proliferation of VSMCs through EGFR.
Materials and methods
Cell culture and treatment
VSMCs, which were derived from the thoracic aortas
of rats, were obtained from American Type Culture Collection. VSMCs
were cultured in DMEM (Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
at 5% CO2 and 37°C. VSMCs were then incubated at 37°C
with PDGF-BB (Sigma-Aldrich; Merck KGaA) at various concentrations
(0, 5, 10, 20 and 40 ng/ml) for 24 h prior to subsequent
experiments.
miR-125a-5p mimics (5′-UCCCUGAGACCCUUUAACCUGUGA-3′),
miR-125a-5p inhibitors (5′-UGCCAGUCUCUAGGUCCCUGAGAC-3′), miR-7
mimics (5′-UGGAAGACUAGUGAUUUUGUUGU-3′), miR-7 inhibitors
(5′-UUGGAUGUUGGCCUAGUUCUGUGU-3′), NC mimics
(5′-UGAACAGUGUUACGUACGAUACC-3′), NC inhibitor
(5′-GGUUCGUACGUACACUGUUCA-3′), pcDNA-EGFR and pcDNA3.1 empty
vectors were commercially synthesized by Shanghai GenePharma Co.,
Ltd. Cell transfections were conducted using
Lipofectamine® 2000 reagent (Beyotime Institute of
Biotechnology), according to the manufacturer's protocol. The
concentration of miR-125a-5p mimics was 50 nM, and the
concentration of miR-125a-5p inhibitor was 100 nM. At 48 h
post-transfection, cells were incubated at 37°C with 20 ng/ml
PDGF-BB for another 24 h prior to subsequent experiments (22,23).
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from VSMCs was collected using
TRIzol® reagent (Thermo Fisher Scientific, Inc.) and
subjected to TaqMan one-step reverse transcription (Applied
Biosystems; Thermo Fisher Scientific, Inc.). RT-qPCR was conducted
on an ABI Prism 7500 (Applied Biosystems; Thermo Fisher Scientific,
Inc.), according to the manufacture's protocol. The relative
expressions of miRNAs were quantified using 2−ΔΔCq
method (24). U6 was used as the
control. The sequences for the primers used are as follows:
miRNA-125a-5p forward, 5′-GCTCCCTGAGACCCT-3′ and reverse,
5′-GAGCAGGCTGGAGAA-3′; miR-7 forward, 5′-CTGTTACTATGGTAGCGACACTG-3′
and reverse, 5′-CACACTGGAGGATTACATTCCC-3′; U6 forward,
5′-CTCGCTTCGGCAGCACA-3′ and reverse, 5′-AACGCTTCACGAATTTGCGT-3′.
The reaction conditions were 94 °C for 3 min, 94°C for 45 sec, 57°C
for 45 sec and 72°C for 45 sec for 30 cycles, and final extension
at 72°C for 10 min.
Cell counting kit-8 (CCK-8) and EdU
assays
Cell growth was analyzed using CCK-8 and EdU assays.
VSMCs (seeding density, 2×103) were maintained in
96-well plates for 24 and 48 h at 37°C in a CO2
incubator. A total of 10 µl CCK-8 (Beyotime Institute of
Biotechnology) was added and VSMCs were incubated at 37°C for 4 h.
Absorbance was analyzed at 450 nm using a microplate reader
(Bio-Rad Laboratories, Inc.).
For the EdU assays, VSMCs (seeding density,
2×103) were grown in 96-well plates at 37°C for 24 h.
Cells were then incubated with 50 µM EdU at 37°C for 2 h and fixed
with 4% formaldehyde at room temperature for 30 min. The nuclei
were counterstained for 15 min at room temperature with 100 ng/ml
DAPI. EdU-positive cells were observed using a fluorescence
microscope (magnification, ×200; Olympus Corporation). EdU-positive
cells were quantified by ImageJ software version 4.3 (National
Institutes of Health).
Wound healing assay
VSMCs (seeding density, 5×104) were grown
in 6-well plates up to 100% confluence and scratched by a sterile
200 µl pipette tip. Cells were then washed with serum-free medium
three times and cultured for 24 h at 37°C. Images were obtained
using an optical microscope (magnification, ×100; BX-51; Olympus
Corporation).
Transwell assay
Transwell assays were used to determine the
migratory and invasive capacity of VSMCs. For the migration assays,
cells (seeding density, 5×104) were suspended in 200 µl
serum-free DMEM and placed into the upper chamber of the Transwell
inserts (8 µm pore size; Corning, Inc.). A total of 600 µl DMEM
supplemented with 20% FBS was added to the lower chamber of the
Transwell. The cells were incubated for 24 h at 37°C.
For the invasion assays, the upper chamber of the
Transwell was pre-coated with Matrigel (BD Biosciences) for 60 min
at 37°C prior to the assay. Cells (seeding density,
5×104) were suspended in 200 µl serum-free DMEM and
placed into the upper chamber of the Transwell inserts (8 µm pore
size; Corning, Inc.). At 24 h post-incubation, non-migrating cells
were removed using a cotton swab. Cells that migrated into the
lower chamber of the Transwell were fixed in 5% glutaraldehyde for
30 min at 4°C and stained with 0.1% crystal violet for 10 min at
room temperature. A total of six nonoverlapping visual fields were
randomly selected for cell counting, and images were captured using
a light microscope (magnification, ×100).
Luciferase reporter assay
Luciferase reporter assay were performed as
previously described (19).
TargetScan (www.targetscan.org/vert_72/) was used to investigate
the putative target genes of miR-125a-5p and miR-7. Luciferase
reporter assays were used to research the association between EGFR
and miR-125a-5p or miR-7 in VSMCs. Wild- and mutant-type EGFR
vectors (EGFR-WT or EGFR-MUT) were synthesized by Shanghai
GenePharma Co., Ltd. and cloned into luciferase genes (Shanghai
GenePharma, Co., Ltd.). VSMCs (5×104) were plated in
24-well plates for 24 h at 37°C, followed by transfection with
EGFR-WT or EGFR-MUT vectors and miR-125a-5p mimics, miR-125a-5p
inhibitors, miR-7 mimics, miR-7 inhibitors or NCs using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). At 48 h post-transfection, luciferase activity
was measured using a Dual Luciferase reporter assay (Promega
Corporation) and normalized to Renilla luciferase.
Western blotting
Total protein was extracted from VSMCs using RIPA
lysis buffer (Bio-Rad Laboratories, Inc.) and protein concentration
was measured with a BCA kit (Beyotime Institute of Biotechnology).
Protein (20 µg) was isolated using 10% SDS-PAGE gels and
transferred to PVDF membranes. The membranes were blocked for 1 h
at room temperature in Tris Buffer Saline Tween supplemented with
0.5% Tween and 5% skimmed milk. Subsequently, the membranes were
treated with primary antibodies overnight at 4°C and then incubated
with horseradish peroxidase (HRP)-conjugated rabbit anti-mouse
Immunoglobulin G (IgG) H&L antibodies (1:2,000; cat. no.
ab6728) or HRP-conjugated goat anti-rabbit IgG H&L antibodies
(1:2,000; cat. no. ab6721) at room temperature for 2 h. The primary
antibodies used were as follows: Anti-p38 mitogen-activated protein
kinase (38 kDa, p38; 1:1,000; cat. no. ab227426),
anti-phosphorylated-p38 (38 kDa, p-p38; 1:1,000; cat. no. ab45381),
anti-matrix metalloproteinase-2 (74 kDa, MMP-2; 1:1,000; cat. no.
ab97779), anti-MMP-9 (78 kDa, 1:1,000; cat. no. ab73734), anti-EGFR
(134 kDa, 1:1,000; cat. no. ab131498) and anti-GAPDH (37 kDa,
1:2,000; cat. no. ab9485) all antibodies from Abcam. Protein bands
were visualized using an ECL kit (Beyotime Institute of
Biotechnology) and quantified using ImageJ software version 4.3
(National Institutes of Health).
Statistical analysis
Data was analyzed using GraphPad Prism software
(version 5.0; GraphPad Software, Inc.) and presented as the mean ±
standard deviation. Each experiment was performed in triplicate.
Student's t-test was used to compare the difference between
two groups and one-way ANOVA analysis followed by Tukey's post-hoc
test was used to compare the differences between multiple groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-125a-5p and miR-7 are
downregulated in PDGF-BB-treated VSMCs
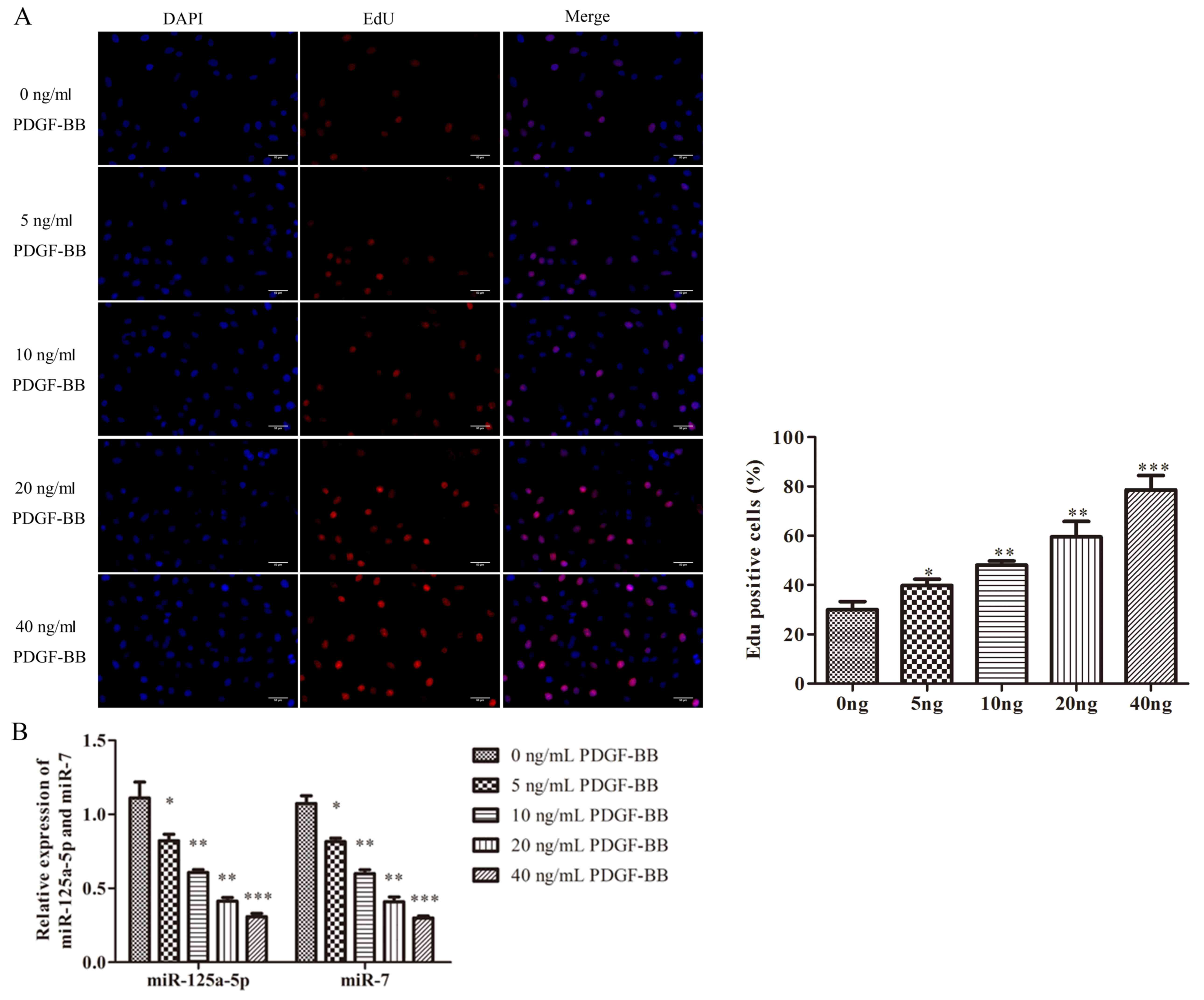
It has been well-documented that PDGF-BB is the most
potent stimuli that triggers the proliferation of VSMCs (7). EdU assays were performed to
investigate the effects of PDGF-BB on VSMCs. The viability of VSMCs
was promoted by PDGF-BB in a concentration-dependent manner,
indicating that PDGF-BB increased the proliferation of VSMCs
successfully (Fig. 1A).
Furthermore, the expression of miR-125a-5p and miR-7 was examined
by RT-qPCR to determine the possible role of miR-125a-5p or miR-7
in PDGF-BB-treated VSMCs. PDGF-BB significantly decreased the
expression of miR-125a-5p and miR-7 in a concentration-dependent
manner, indicating that miR-125a-5p and miR-7 were downregulated in
PDGF-BB-treated VSMCs (Fig. 1B).
The results revealed that dysregulated miR-125a-5p and miR-7 may be
associated with the proliferative state of VSMCs.
miR-125a-5p and miR-7 restrain the
growth of PDGF-BB-treated VSMCs
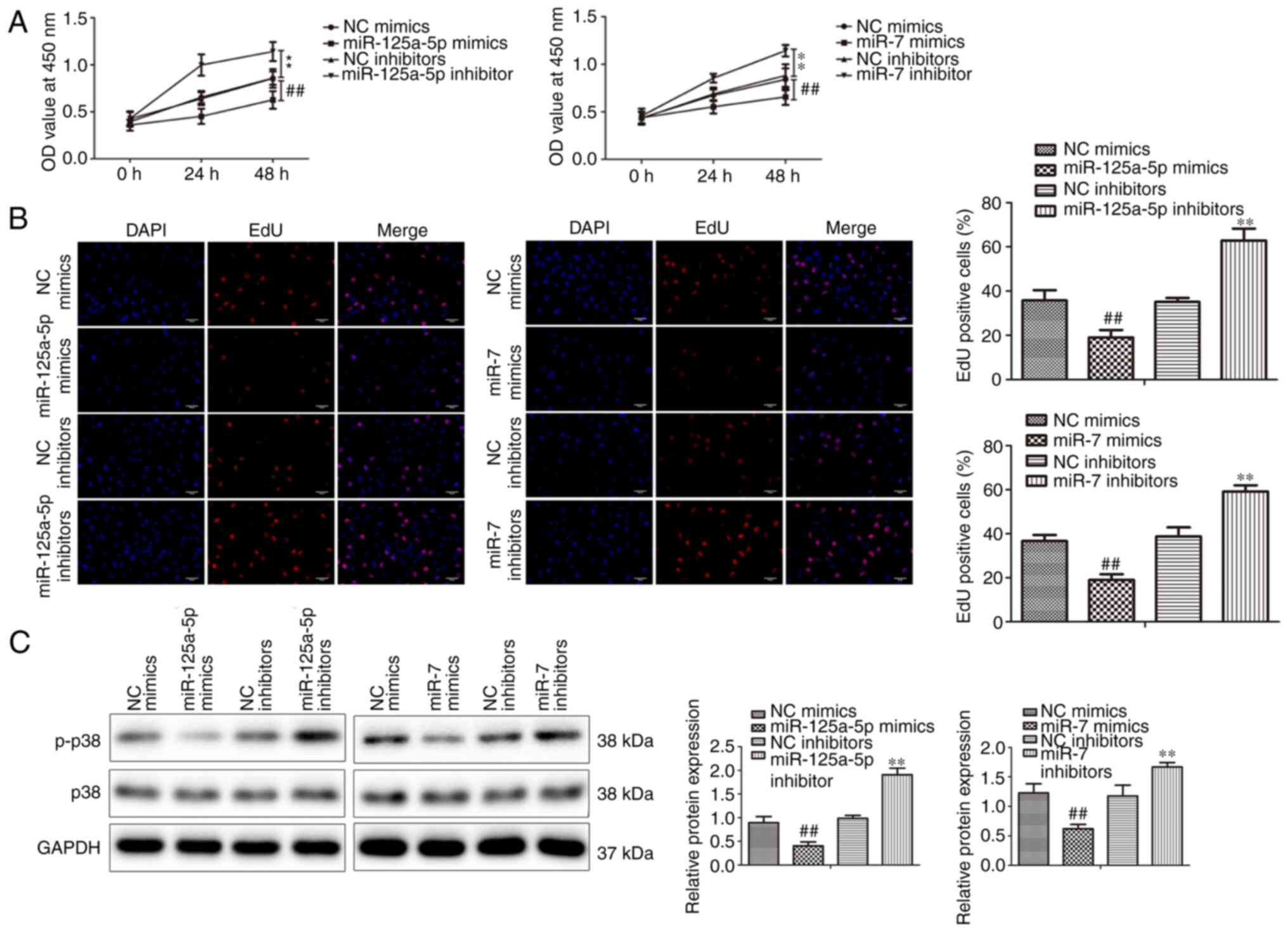
VSMCs were transfected with miR-125a-5p and miR-7
mimics or inhibitors or NCs and incubated with PDGF-BB to
investigate the biological impacts of miR-125a-5p and miR-7 on the
growth of PDGF-BB-treated VSMCs. CCK-8 assays indicated that
miR-125a-5p and miR-7 mimics significantly decreased the
proliferation of VSMCs compared with mimic NC groups in a
time-dependent manner (Fig. 2A).
Additionally, the inhibition of miR-125a-5p and miR-7 significantly
promoted the proliferation of VSMCs compared with inhibitor NC
groups. The results of the EdU assays revealed that the miR-125a-5p
and miR-7 mimics significantly suppressed the proliferation of
VSMCs, while inhibitors demonstrated the opposite effect (Fig. 2B). Furthermore, the protein
expression of cell proliferation marker p38 (21) was assessed via western blotting.
miR-125a-5p and miR-7 mimics significantly inhibited p38
phosphorylation, while inhibitors promoted p38 phosphorylation
(Fig. 2C). p38 expression was not
markedly different between groups. In summary, the results
indicated that miR-125a-5p and miR-7 inhibited the proliferation of
PDGF-BB-treated VSMCs.
miR-125a-5p and miR-7 repress the
migration and invasion of PDGF-BB-treated VSMCs
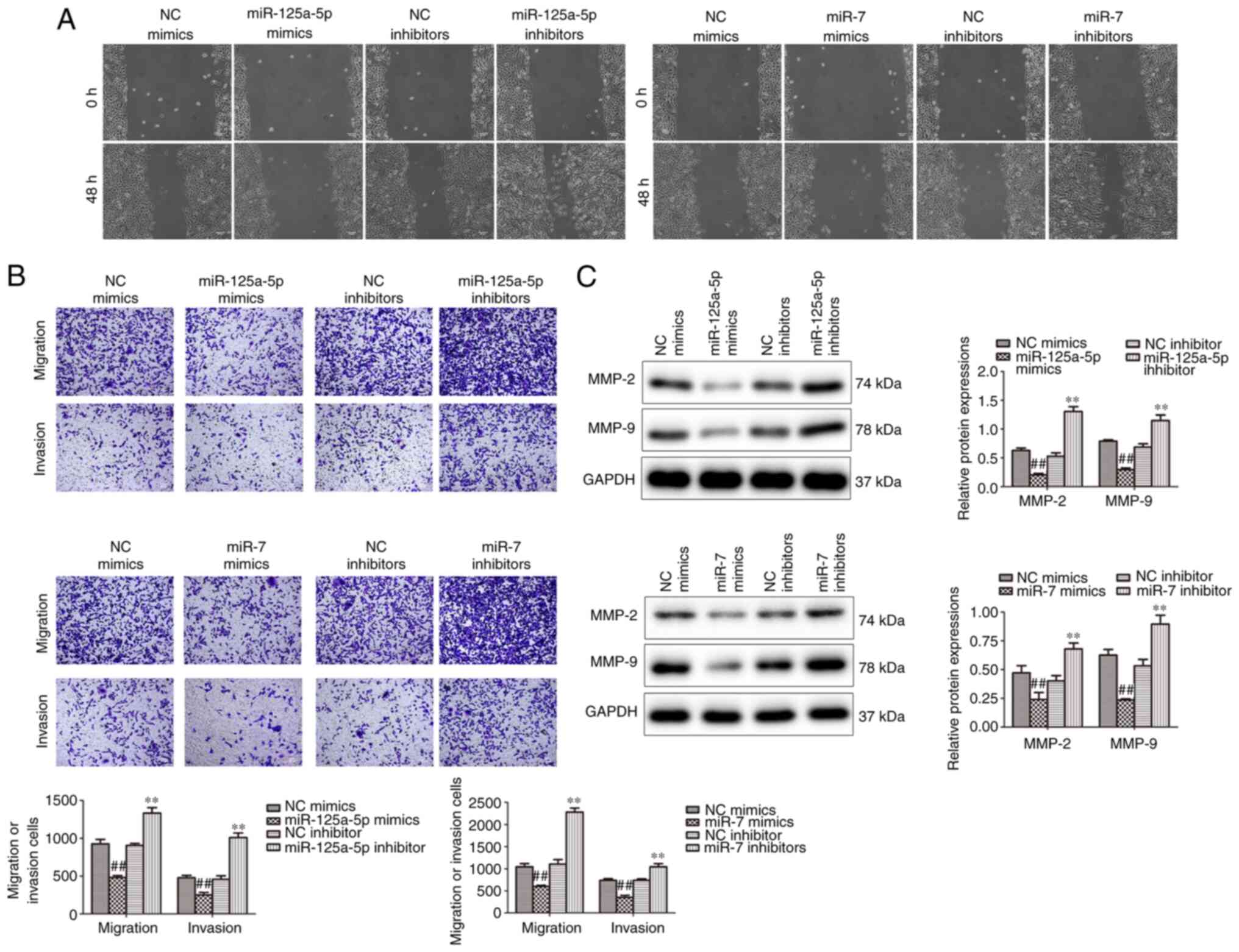
Wound healing and Transwell assays were performed to
analyze the effect of miR-125a-5p and miR-7 on the migration and
invasion of PDGF-BB-treated VSMCs. The results demonstrated that
miR-125a-5p and miR-7 mimics markedly attenuated migration, while
inhibition promoted the migration of PDGF-BB-treated VSMCs
(Fig. 3A). Furthermore, Transwell
assays verified that miR-125a-5p and miR-7 mimics markedly
decreased the migration and invasion in PDGF-BB-treated VSMCs,
while inhibitors promoted migration and invasion (Fig. 3B). Furthermore, the expression of
MMP-2 and MMP-9, which modulate the migration of VSMCs, were
determined by western blotting (25). The results demonstrated that MMP-2
and MMP-9 protein levels were significantly reduced in the
miR-125a-5p and miR-7 mimic groups and significantly increased in
the inhibitor groups compared with their corresponding NCs
(Fig. 3C). The results indicated
that miR-125a-5p and miR-7 decreased the migration and invasion of
VSMCs stimulated by PDGF-BB.
EGFR is a direct target of miR-125a-5p
and miR-7 in VSMCs
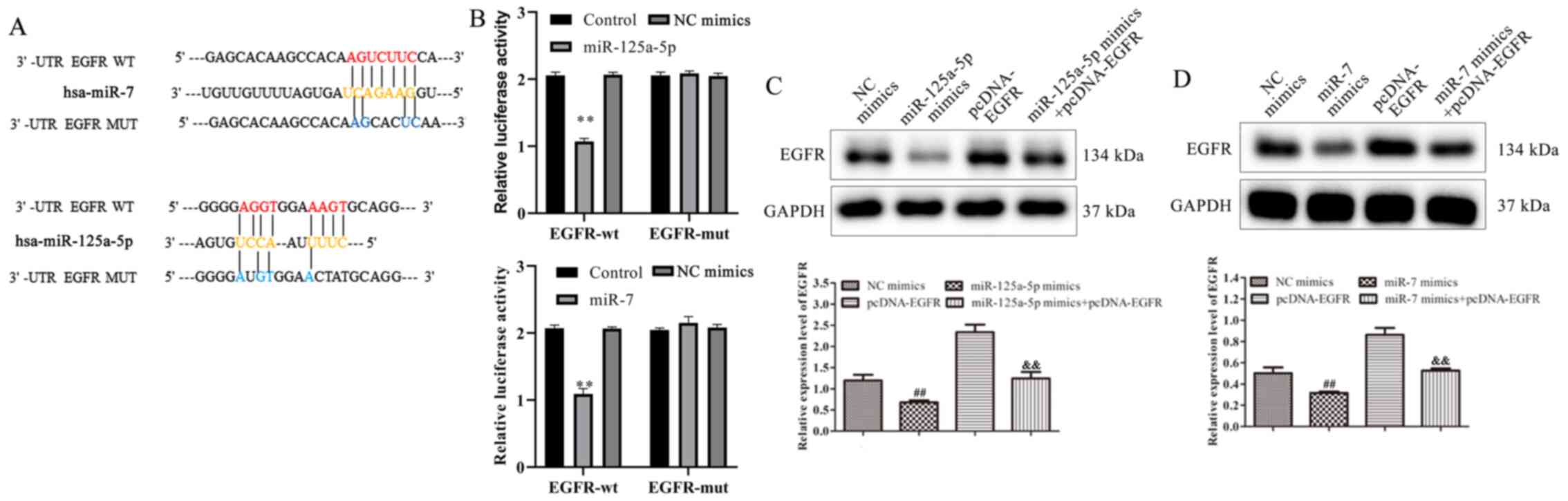
Bioinformatics analysis was used to predict
potential target genes to further investigate the underlying
mechanisms of miR-125a-5p and miR-7 in the proliferation and
migration of VSMCs. According to the results, EGFR was reported to
contain binding sites for miR-125a-5p and miR-7 (Fig. 4A). To confirm the binding sites,
luciferase reporter assays were conducted. Co-transfection with
miR-125a-5p or miR-7 mimics and EGFR-WT decreased the relative
luciferase activity of VSMCs (Fig.
4B). The results of the mutated EGFR target sequences did not
demonstrate significant differences in luciferase activity. To
investigate the association between miR-125a-5p or miR-7 and EGFR,
miR-125a-5p or miR-7 mimics and pcDNA-EGFR were co-transfected into
PDGF-BB-treated VSMCs. The results of western blotting indicated
that EGFR expression was significantly decreased in miR-125a-5p and
miR-7 mimics group compared with NC mimics, while overexpression of
EGFR reversed the miR-125a-5p or miR-7 mimic-mediated EGFR
inhibition (Fig 4C and D). The
results demonstrated that EGFR was identified as a target gene of
miR-125a-5p and miR-7 and that these miRNAs negatively regulated
EGFR expression by binding to its 3′-UTR.
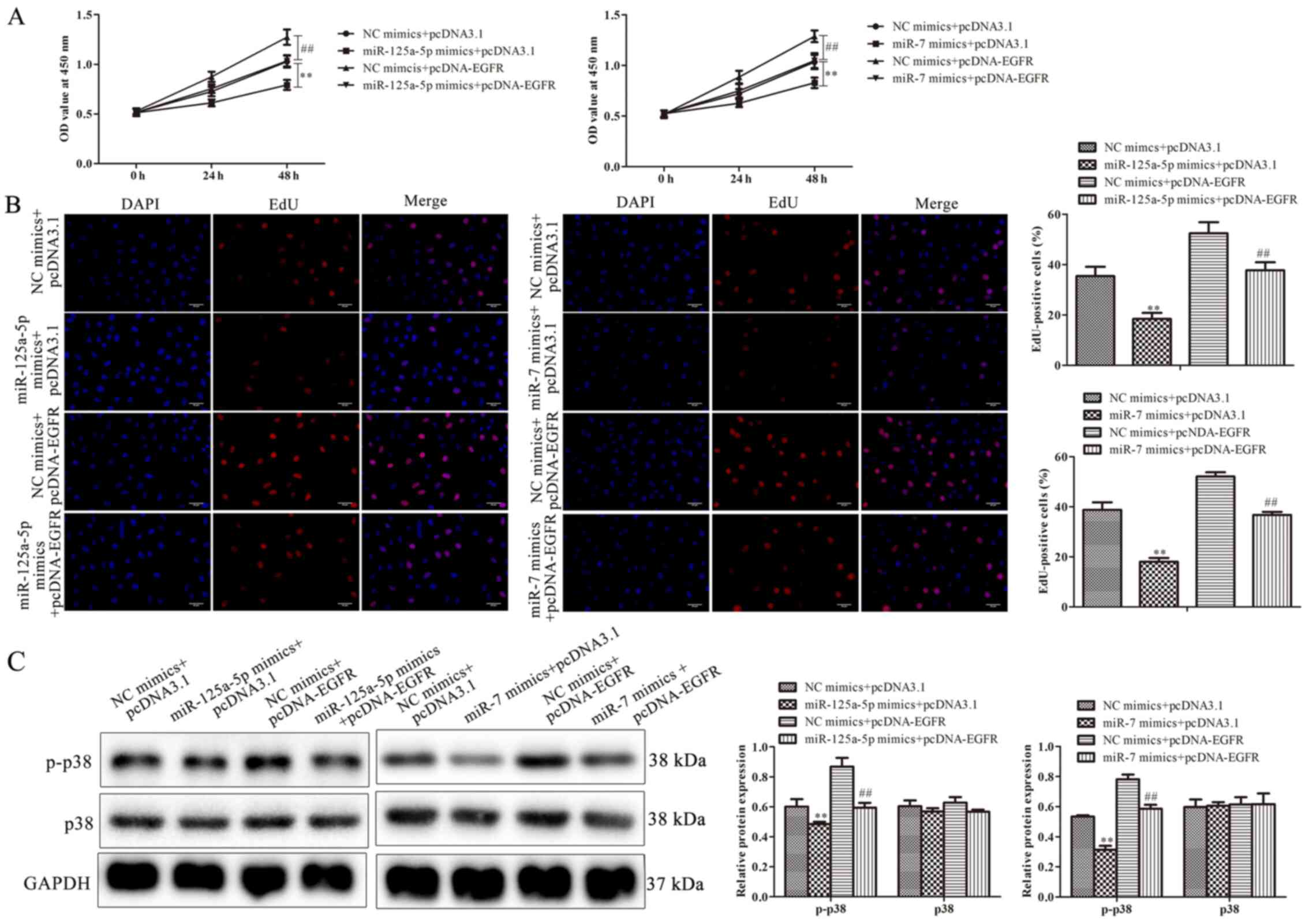
miR-125a-5p and miR-7 inhibits cell
proliferation by targeting EGFR
To further determine whether EGFR was involved in
miR-125a-5p- and miR-7-mediated cell growth of PDGF-BB-treated
VSMCs, VSMCs were co-transfected with pcDNA-EGFR and miR-125a-5p or
miR-7 mimics, followed by PDGF-BB treatment. The results of the
CCK-8 assay revealed that EGFR overexpression significantly
increased the proliferation of PDGF-BB-treated VSMCs compared with
the NC group (Fig. 5A). Similar to
these results, EdU assays also indicated that miR-125a-5p and miR-7
mimics reduced the proliferation of PDGF-BB-treated VSMCs, while
EGFR overexpression increased proliferation (Fig. 5B). Cells co-treated with pcDNA-EGFR
and miR-125a-5p or miR-7 mimics reversed EGFR-mediated
proliferation (Fig. 5B).
miR-125a-5p or miR-7 mimics reduced the level of p-p38, while
pcDNA-EGFR had the opposite effect; however, cells co-treated with
pcDNA-EGFR eliminated the decrease in p-p38 expression induced by
miR-125a-5p or miR-7 mimics (Fig.
5C). The results indicated that EGFR was associated with the
inhibition of PDGF-BB-treated VSMC proliferation mediated by
miR-125a-5p or miR-7.
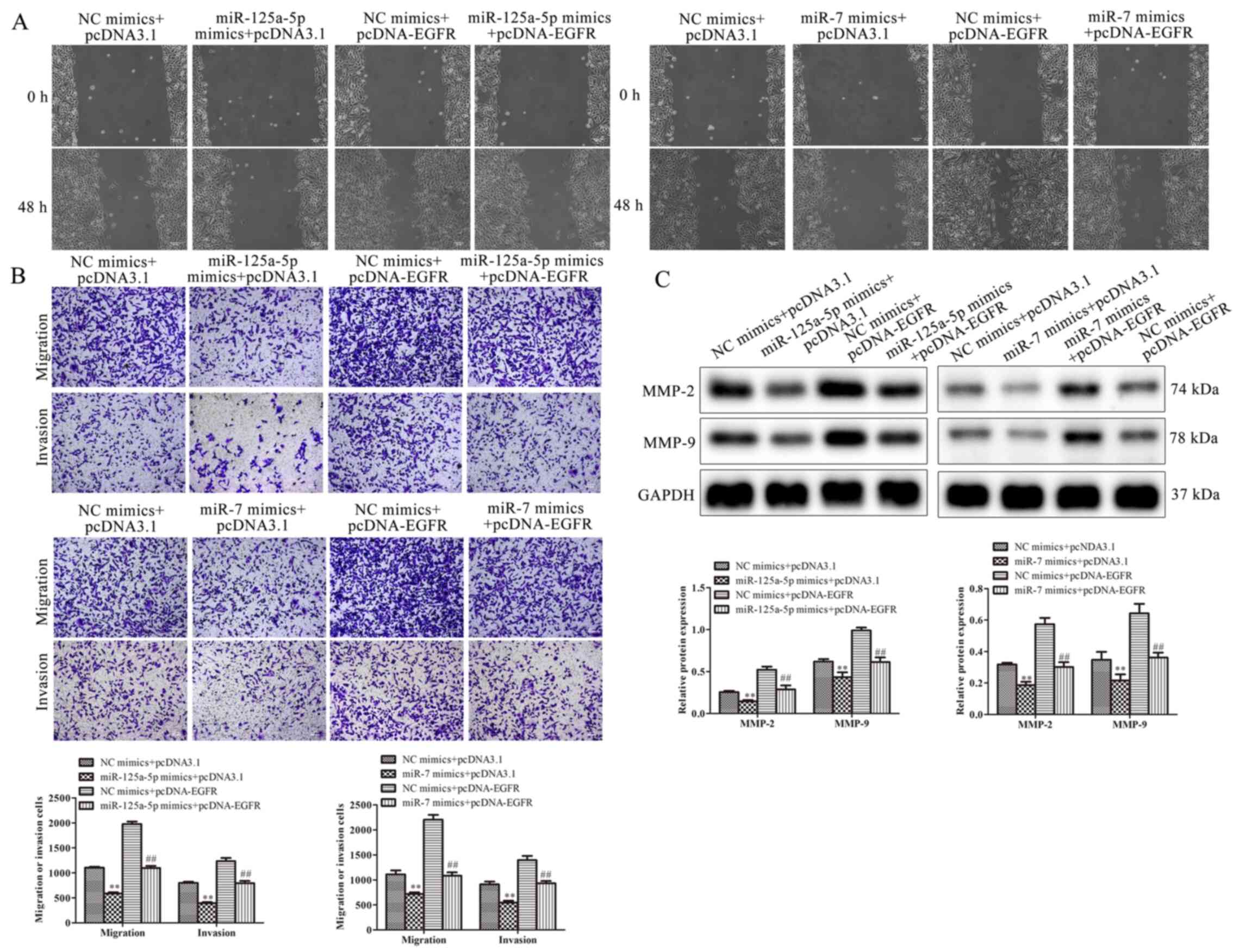
miR-125a-5p and miR-7 suppress
migration and invasion by targeting EGFR
The effect of EGFR and miR-125a-5p or miR-7 on the
migration and invasion of VSMCs was investigated. The results
demonstrated that miR-125a-5p and miR-7 mimics decreased migration,
while EGFR overexpression increased the migration of
PDGF-BB-treated VSMCs. Cells co-treated with EGFR markedly
ameliorated the suppressive function of miR-125a-5p and miR-7
mimics on migration (Fig. 6A).
Furthermore, the results of Transwell assays revealed that
miR-125a-5p and miR-7 mimics decreased migration and invasion,
while EGFR upregulation reversed this result (Fig. 6B). These results were consistent
with those of wound healing. The expression of migratory-associated
proteins, including MMP-2 and MMP-9, were examined using western
blotting. The expression of MMP-2 and MMP-9 was significantly
reduced in VSMCs transfected with miR-125a-5p and miR-7 mimics
compared with the NC group, while cells co-transfected with EGFR
exhibited attenuated MMP-2 and MMP-9 levels (Fig. 6C). In summary, the results
demonstrated that miR-125a-5p and miR-7 repressed the migration and
invasion of PDGF-BB-induced VSMCs by regulating EGFR.
Discussion
The results of the current study revealed that
miR-125a-5p and miR-7 were downregulated in PDGF-BB-treated VSMCs,
while upregulation of miR-125a-5p and miR-7 significantly
suppressed cell growth and migration in vitro. The
transfected miRNAs impeded the growth and migration of
PDGF-BB-treated VSMCs by targeting EGFR. Therefore, miR-125a-5p and
miR-7 may be novel regulators implicated in the proliferation and
migration of VSMCs. However, a limitation of the current study was
that other cells lines or types of cell were not investigated.
Accumulating evidence has indicated that miR-125a-5p
modulates the progression of cancer, the differentiation of
skeletal muscle cells and the inhibition of angiogenesis (26–31).
miR-125a-5p has been established as a tumor suppressor that
prevents the development of human diseases including colorectal and
bladder cancer (32–34). Similarly, miR-7 acts as a tumor
inhibitor in glioblastoma, gastric, liver, breast, head and neck,
non-small cell lung carcinoma, melanoma, cervical, prostate,
colorectal, thyroid, ovarian and schwannoma carcinoma (35). However, the roles of miR-125a-5p and
miR-7 in VSMCs are yet to be elucidated. The current study
demonstrated that miR-125a-5p and miR-7 were associated with the
proliferation and migration of PDGF-BB-treated VSMCs.
Overexpression of miR-125a-5p and miR-7 suppressed the cell growth
and migration of PDGF-BB-treated VSMCs, while downregulation of
miR-125a-5p and miR-7 exerted the opposite effect. Additionally,
the expression of the miRNAs were negatively associated with
expression of p-p38, MMP-2 and MMP-9, which are proliferation and
migration marker genes of VSMCs. It has been demonstrated that
MMP-9 degrades collagen IV, collagen V and gelatin in the
extracellular matrix and promotes tumor invasion and metastasis
(36–39). Li et al (40) examined the effect of MMP-9 on the
angiogenesis of gastric cancer cells using gelatin zymography, the
results of which indicated that MMP-9 was positively associated
with angiogenesis progression in patients with gastric cancer
(40). These results were
consistent with that of the current study. The results verified
that miR-125a-5p and miR-7 regulated the proliferation and
migration of VSMCs.
It is well documented that miRNAs modulate the
biological functions of cells by downregulating mRNA targets
post-transcriptionally (41). Due
to the suppression of miR-125a-5p and miR-7 on the growth and
migration of VSMCs, the current study hypothesized that target
gene(s) should be positively associated with cell proliferation and
migration. To explore the mechanism underlying the inhibition of
the proliferation and migration of VMSCs mediated by miR-125a-5p
and miR-7, the putative target gene for both miRNAs was identified
using bioinformatics tools. EGFR was considered a candidate target
for miR-125a-5p and miR-7 in VMSCs. As a member of ErbB family,
EGFR serves an important role in various pathophysiological
processes, including cell tumorigenesis and transformation
(42,43). The association between EGFR and
miR-125a-5p or miR-7 was further validated by luciferase reporter
assays and western blotting. EGFR had a negative association with
the expression of miR-125a-5p and miR-7 in VSMCs. Previous studies
have demonstrated that miR-125 and miR-7 target numerous genes,
including EGFR (44,45). The results of the current study are
therefore consistent with these. In VSMCs, EGFR is involved with
cell growth, migration, inflammation and vascular remodeling,
consequently triggering vascular diseases, including
atherosclerosis and hypertension (46). In the current study, overexpression
of EGFR accelerated cell growth and migration of PDGF-BB-treated
VSMCs. Furthermore, the results demonstrated that EGFR
overexpression abrogated the miR-125a-5p and miR-7-mediated
inhibition of VSMC proliferation and migration. In summary, the
results verified that miR-125a-5p and miR-7 decreased the migration
and invasion of PDGF-BB-treated VSMCs by targeting EGFR. However, a
limitation of the current study is that the effect of miR-125a-5p
and miR-7 on EGFR luciferase activity and protein level were not
investigated. These should be researched in future studies.
In conclusion, the results demonstrated that the
expression of miR-125a-5p and miR-7 was decreased in
PDGF-BB-stimulated VSMCs. Following this, EGFR was reported to be a
target of miR-125a-5p and miR-7, and overexpression of EGFR
reversed the inhibition of VSMC proliferation, migration and
invasion mediated by miR-125a-5p and miR-7. The results indicated
that miR-125a-5p and miR-7 may serve as novel molecular targets in
the proliferation and migration of VSMCs.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
HZ and XL conceived and designed the study. SL and
YH performed the literature search and acquisition of data. DG and
YW contributed to analysis and interpretation of data. HZ and XL
drafted and revised the manuscript. HZ and XL confirm the
authenticity of all the raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lacolley P, Regnault V, Nicoletti A, Li Z
and Michel JB: The vascular smooth muscle cell in arterial
pathology: A cell that can take on multiple roles. Cardiovasc Res.
95:194–204. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen Q, Zhang H, Liu Y, Adams S, Eilken H,
Stehling M, Corada M, Dejana E, Zhou B and Adams RH: Endothelial
cells are progenitors of cardiac pericytes and vascular smooth
muscle cells. Nat Commun. 7:124222016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Régent A, Ly KH, Lofek S, Clary G, Tamby
M, Tamas N, Federici C, Broussard C, Chafey P, Liaudet-Coopman E,
et al: Proteomic analysis of vascular smooth muscle cells in
physiological condition and in pulmonary arterial hypertension:
Toward contractile versus synthetic phenotypes. Proteomics.
16:2637–2649. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bennett MR, Sinha S and Owens GK: Vascular
smooth muscle cells in atherosclerosis. Circ Res. 118:692–702.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
In This Issue. Proc Natl Acad Sci.
104:7731–7732. 2007.doi: 10.1073/iti1907104. View Article : Google Scholar
|
|
6
|
Kato Y, Yokoyama U, Fujita T, Umemura M,
Kubota T and Ishikawa Y: Epac1 deficiency inhibits basic fibroblast
growth factor-mediated vascular smooth muscle cell migration. J
Physiol Sci. 69:175–184. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim SH, Yun SJ, Kim YH, Ha JM, Jin SY, Lee
HS, Kim SJ, Shin HK, Chung SW and Bae SS: Essential role of
krüppel-like factor 5 during tumor necrosis factor α-induced
phenotypic conversion of vascular smooth muscle cells. Biochem
Biophys Res Commun. 463:1323–1327. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shawky NM and Segar L: Sulforaphane
inhibits platelet-derived growth factor-induced vascular smooth
muscle cell proliferation by targeting mTOR/p70S6kinase signaling
independent of Nrf2 activation. Pharmacol Res. 119:251–264. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiang R, Chen J, Li S, Yan H, Meng Y, Cai
J, Cui Q, Yang Y, Xu M, Geng B and Yang J: VSMC-specific deletion
of FAM3A attenuated ang ii-promoted hypertension and cardiovascular
hypertrophy. Circ Res. 126:1746–1759. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dong S, Xiong W, Yuan J, Li J, Liu J and
Xu X: MiRNA-146a regulates the maturation and differentiation of
vascular smooth muscle cells by targeting NF-κB expression. Mol Med
Rep. 8:407–412. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Frismantiene A, Philippova M, Erne P and
Resink TJ: Smooth muscle cell-driven vascular diseases and
molecular mechanisms of VSMC plasticity. Cell Signal. 52:48–64.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang Y, Zhang L, Wang Y, Ding H, Xue S,
Qi H and Li P: MicroRNAs or long noncoding RNAs in diagnosis and
prognosis of coronary artery disease. Aging Dis. 10:353–366. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cai W, Zhang J and Yang J, Fan Z, Liu X,
Gao W, Zeng P, Xiong M, Ma C and Yang J: MicroRNA-24 attenuates
vascular remodeling in diabetic rats through PI3K/Akt signaling
pathway. Nutr Metab Cardiovasc Dis. 29:621–632. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gao F, Kataoka M, Liu N, Liang T, Huang
ZP, Gu F, Ding J, Liu J, Zhang F, Ma Q, et al: Therapeutic role of
miR-19a/19b in cardiac regeneration and protection from myocardial
infarction. Nat Commun. 10:18022019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu WH, Hu CP, Chen XP, Zhang WF, Li XW,
Xiong XM and Li YJ: MicroRNA-130a mediates proliferation of
vascular smooth muscle cells in hypertension. Am J Hypertens.
24:1087–1093. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gareri C, Iaconetti C, Sorrentino S,
Covello C, De Rosa S and Indolfi C: miR-125a-5p modulates
phenotypic switch of vascular smooth muscle cells by targeting
ETS-1. J Mol Biol. 429:1817–1828. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Goettsch C: miRNA-125b and their targets
in cardiovascular calcification. Proc Physiol Soc. 13:1234–1246.
2013.
|
|
18
|
Fan X, Liu M, Tang H, Leng D, Hu S, Lu R,
Wan W and Yuan S: MicroRNA-7 exerts antiangiogenic effect on
colorectal cancer via ERK signaling. J Surg Res. 240:48–59. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yano S, Kondo K, Yamaguchi M, Richmond G,
Hutchison M, Wakeling A, Averbuch S and Wadsworth P: Distribution
and function of EGFR in human tissue and the effect of EGFR
tyrosine kinase inhibition. Anticancer Res. 23:3639–3650.
2003.PubMed/NCBI
|
|
20
|
Clauditz TS, Gontarewicz A, Lebok P,
Tsourlakis MC, Grob TJ, Münscher A, Sauter G, Bokemeyer C, Knecht R
and Wilczak W: Epidermal growth factor receptor (EGFR) in salivary
gland carcinomas: Potentials as therapeutic target. Oral Oncol.
48:991–996. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee JS, Kim SY, Kwon CH and Kim YK:
EGFR-dependent ERK activation triggers hydrogen peroxide-induced
apoptosis in OK renal epithelial cells. Arch Toxicol. 80:337–346.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schreier B, Döhler M, Rabe S, Schneider B,
Schwerdt G, Ruhs S, Sibilia M, Gotthardt M, Gekle M and Grossmann
C: Consequences of epidermal growth factor receptor (ErbB1) loss
for vascular smooth muscle cells from mice with targeted deletion
of ErbB1. Arterioscler Thromb Vasc Biol. 31:1643–1652. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dong X, Hu H, Fang Z, Cui J and Liu F:
CTRP6 inhibits PDGF-BB-induced vascular smooth muscle cell
proliferation and migration. Biomed Pharmacother. 103:844–850.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-(Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ye G, Huang K, Yu J, Zhao L, Zhu X, Yang
Q, Li W, Jiang Y, Zhuang B, Liu H, et al: MicroRNA-647 Targets
SRF-MYH9 axis to suppress invasion and metastasis of gastric
cancer. Theranostics. 7:3338–3353. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sun L, Lian JX and Meng S: MiR-125a-5p
promotes osteoclastogenesis by targeting TNFRSF1B. Cell Mol Biol
Lett. 24:232019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu H, Ma Y, Liu C, Li P and Yu T: Reduced
miR-125a-5p level in non-small-cell lung cancer is associated with
tumour progression. Open Biol. 8:1801182018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vo DT, Karanam NK, Ding L, Saha D, Yordy
JS, Giri U, Heymach JV and Story MD: miR-125a-5p functions as tumor
suppressor microRNA and is a marker of locoregional recurrence and
poor prognosis in head and neck cancer. Neoplasia. 21:849–862.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li H, An X, Bao L, Li Y, Pan Y, He J, Liu
L, Zhu X, Zhang J, Cheng J and Chu W: MiR-125a-3p-KLF15-BCAA
regulates the skeletal muscle branched-chain amino acid metabolism
in nile tilapia (Oreochromis niloticus) during starvation.
Front Genet. 11:8522020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Che P, Liu J, Shan Z, Wu R, Yao C, Cui J,
Zhu X and Wang J, Burnett MS, Wang S and Wang J: miR-125a-5p
impairs endothelial cell angiogenesis in aging mice via RTEF-1
downregulation. Aging Cell. 13:926–934. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zheng X, Wu Z, Xu K, Qiu Y, Su X, Zhang Z
and Zhou M: Interfering histone deacetylase 4 inhibits the
proliferation of vascular smooth muscle cells via regulating
MEG3/miR-125a-5p/IRF1. Cell Adh Migr. 13:41–49. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang X, Qiu J, Kang H, Wang Y and Qian J:
miR-125a-5p suppresses colorectal cancer progression by targeting
VEGFA. Cancer Manag Res. 10:5839–5853. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y, Zhang D, Lv J, Wang S and Zhang
Q: MiR-125a-5p suppresses bladder cancer progression through
targeting FUT4. Biomed Pharmacother. 108:1039–1047. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu L, Li Y, Yin L, Qi Y, Sun H, Sun P, Xu
M, Tang Z and Peng J: miR-125a-5p ameliorates hepatic glycolipid
metabolism disorder in type 2 diabetes mellitus through targeting
of STAT3. Theranostics. 8:5593–5609. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Horsham JL, Ganda C, Kalinowski FC, Brown
RA, Epis MR and Leedman PJ: MicroRNA-7: A miRNA with expanding
roles in development and disease. Int J Biochem Cell Biol.
69:215–224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ha YM, Nam JO and Kang YJ: Pitavastatin
regulates ang II induced proliferation and migration via IGFBP-5 in
VSMC. Korean J Physiol Pharmacol. 19:499–506. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang BF, Jiang H, Chen J, Guo X, Hu Q and
Yang S: KDM3A inhibition attenuates high concentration
insulin-induced vascular smooth muscle cell injury by suppressing
MAPK/NF-κB pathways. Int J Mol Med. 41:1265–1274. 2018.PubMed/NCBI
|
|
38
|
Park HJ, Kim MK, Kim Y, Bae SS, Kim HJ,
Bae SK and Bae MK: Gastrin-releasing peptide promotes the migration
of vascular smooth muscle cells through upregulation of matrix
metalloproteinase-2 and −9. BMB Rep. 50:628–633. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Qi S, Perrino S, Miao X, Lamarche-Vane N
and Brodt P: The chemokine CCL7 regulates invadopodia maturation
and MMP-9 mediated collagen degradation in liver-metastatic
carcinoma cells. Cancer Lett. 483:98–113. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li TJ, Jiang YM, Hu YF, Huang L, Yu J,
Zhao LY, Deng HJ, Mou TY, Liu H, Yang Y, et al:
Interleukin-17-producing neutrophils link inflammatory stimuli to
disease progression by promoting angiogenesis in gastric cancer.
Clin Cancer Res. 23:1575–1585. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang P, Guan Q, Zhou D, Yu Z, Song Y and
Qiu W: miR-21 inhibitors modulate biological functions of gastric
cancer cells via PTEN/PI3K/mTOR pathway. DNA Cell Biol. 37:38–45.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schreier B, Schwerdt G, Heise C, Bethmann
D, Rabe S, Mildenberger S and Gekle M: Substance-specific
importance of EGFR for vascular smooth muscle cells motility in
primary culture. Biochim Biophys Acta. 1863:1519–1533. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Spano JP, Lagorce C, Atlan D, Milano G,
Domont J, Benamouzig R, Attar A, Benichou J, Martin A, Morere JF,
et al: Impact of EGFR expression on colorectal cancer patient
prognosis and survival. Ann Oncol. 16:102–108. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhou X, Hu Y, Dai L, Wang Y, Zhou J, Wang
W, Di W and Qiu L: MicroRNA-7 inhibits tumor metastasis and
reverses epithelial-mesenchymal transition through AKT/ERK1/2
inactivation by targeting EGFR in epithelial ovarian cancer. PLoS
One. 9:e967182014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lin T, Ren Q, Zuo W, Jia R, Xie L, Lin R,
Zhao H, Chen J, Lei Y, Wang P, et al: Valproic acid exhibits
anti-tumor activity selectively against
EGFR/ErbB2/ErbB3-coexpressing pancreatic cancer via induction of
ErbB family members-targeting microRNAs. J Exp Clin Cancer Res.
38:1502019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Schreier B, Gekle M and Grossmann C: Role
of epidermal growth factor receptor in vascular structure and
function. Curr Opin Nephrol Hypertens. 23:113–121. 2014. View Article : Google Scholar : PubMed/NCBI
|