Introduction
Operations, traumas, burns and scalds can cause skin
trauma, and the rapid and effective repair of skin wounds is key to
ensuring the function of the skin barrier. The process of skin
wound healing is a complex regenerative reaction that involves four
stages: Hemostasis, inflammatory response, cell proliferation and
tissue reconstruction (1). During
this dynamic process, excessive collagen deposition and fibroblast
proliferation and activation lead to the formation of pathological
scars, including hypertrophic scars and keloids (2). Pathological scars cause not only pain
and itching but also aesthetic issues and seriously affect the
patients' mental state and quality of life. Therefore, studying the
mechanism of pathological scar formation and hyperplasia, as well
as its prevention and treatment measures is one of the hot topics
in the field of medical plastic surgery and has important clinical
significance (3,4).
In recent years, stem cell therapy has become an
effective treatment and research hotspot of skin wound healing.
Previous studies have shown that adipose-derived mesenchymal stem
cells (ADSCs), one of the most widely used adult stem cells, have a
beneficial ability to promote skin wound healing and reduce scar
formation (5–7). Moreover, it has been reported that
paracrine cytokines, exosomes and other acellular bioactive
derivatives are the main factors via which ADSCs exert their
biological functions (8). Exosomes
are a bilayer phospholipid membrane structure secreted by cells
with a diameter of 40–150 nm and a density range of 1.09–1.18 g/ml,
and contain proteins, lipids and RNA [mRNA and microRNA
(miRNA/miR)] vesicle-like substances (9–11).
Exosomes, as important components in the paracrine pathway of stem
cells, have high application prospects in promoting the repair and
regeneration of skin wounds (12).
Compared with the direct use of stem cells for tissue repair,
exosomes have improved safety and easier storage and
transportation, are fast and efficient, have no ethical
restrictions and have a wide range of sources, amongst other
advantages (13,14). Previous research has shown that
exosomes can participate in various processes of skin tissue repair
and regeneration, as well as promote skin healing and skin tissue
regeneration by enhancing the proliferation and migration of skin
cells (15,16) and angiogenesis (17,18),
in addition to regulating the immune response (19). Thus, these mechanisms provide a new
method to achieve cell-free therapy.
miRNAs are a type of non-coding RNA with a length of
21–23 nucleotides that can affect multiple gene networks
simultaneously to coordinate biological responses (20). miRNAs exist in tissues throughout
the body and can inhibit the expression of target genes by binding
to the 3′ untranslated region (3′UTR), coding region or 5′UTR of
target mRNA, thereby blocking mRNA translation or degrading mRNA
(21). Subsequently, miRNAs can
participate in cell proliferation and various growth and
development processes, such as differentiation, migration,
metabolism and apoptosis (22).
Studies have reported that miRNAs have a significant
effect in regenerative medicine and can regulate the growth of a
variety of tissues in various processes, including skin healing
(23), bone regeneration (24), liver regeneration (25), kidney regeneration (26), and myocardial regeneration (27). miRNAs can obviously regulate the
proliferation of fibroblasts and the synthesis of extracellular
matrix via several molecular mechanisms. Additionally, they are key
regulators of skin morphogenesis and wound healing. For example,
miR-29a has been reported to inhibit fibroblast proliferation,
migration and collagen deposition after skin thermal injury, and
can promote the repair of denatured dermis (28,29).
Furthermore, mesenchymal stem cell (MSC) therapy can improve skin
wound healing in diabetic mice by correcting the miR-29a imbalance
(30). A recent study revealed that
the expression level of miR-29a was significantly downregulated in
human skin keloids, and long non-coding RNA H19 promoted the
proliferation and metastasis of fibroblasts by modifying downstream
miR-29a and collagen type I α 1 chain (31), suggesting that the downregulation of
miR-29a is closely associated with the formation of keloids induced
by excessive proliferation of fibroblasts.
The TGF-β/Smad signaling pathway is considered an
important regulator during skin wound healing and pathological scar
formation (32). Studies have shown
that miR-29b (another member of the miR-29a family) can target
TGF-β1 in fibroblasts to regulate the activation of the
TGF-β1/Smad3 signaling pathway, and thus, promote skin wound
healing and reduce excessive scar formation (6,33,34).
miR-29a has also been reported to attenuate Angiotensin II-induced
left ventricular remodeling by inhibiting TGF-β/Smad signaling
pathway (35). However, whether
miR-29a reduces excessive scar formation via TGF-β/Smad signaling
pathway remains unknown.
Previous studies have reported that miR-29a was
stably expressed in the exosomes of human ADSCs (hADSCs-exo)
(36), but whether ADSCs-exo can
inhibit scar formation by delivering exogenous miR-29a to the wound
site is yet to be determined. The present study aimed to evaluate
the inhibitory effect of hADSCs-exo enriched with miR-29a on scar
formation for wound healing purposes.
Materials and methods
Cell culture and transfection
All cells were purchased from BeNa Culture
Collection. hADSCs (cat. no. BNCC340147), human skin fibroblasts
(HSFs; cat. no. BNCC353686) and human hypertrophic scar fibroblasts
(HSFBs; cat. no. BNCC342248) were cultured in DMEM (HyClone;
Cytiva) containing 10% FBS (Gibco; Thermo Fisher Scientific, Inc.)
and 1% (v/v) penicillin/streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) and were then incubated at 37°C in a humidified
atmosphere containing 5% CO2. All cell experiments were
approved by the Ethics Committee of The First Affiliated Hospital
of Kunming Medical University (approval no. kmmu2021188) due to the
use of non-immortalized human cells, and carried out in accordance
with the cell experiment management regulations of the
hospital.
For the transfection experiment, miR-29a mimics
(5′-UAGCACCAUCUGAAAUCGGUUA-3′), miR-29a inhibitor
(5′-UAACCGAUUUCAGAUGGUGCUA-3′) and corresponding negative controls
(NC; mimics-NC: 5′-UUCUCCGAACGUGUCACGU-3′; inhibitor-NC:
5′-CAGUACUUUUGUGUAGUACAA-3′) were constructed by Guangzhou RiboBio
Co., Ltd. hADSCs were seeded into 6-well plates at
1.5×105 cells/well for 24 h. Then, 100 nM miR-29a mimics
or miR-29a inhibitor was transfected into hADSCs with
Lipofectamine® 2000 transfection reagent (Invitrogen;
Thermo Fisher Scientific, Inc.), following the manufacturer's
instructions. Transfection was performed for 6 h at 37°C with 5%
CO2. At 4 h post-transfection, the transfection
efficiency was detected via reverse transcription-quantitative
(RT-q)PCR (Fig. S1).
Exosome extraction and
identification
Subconfluent hADSCs in the log phase, which were
obtained by filtering the supernatants through 0.22 µm pore filters
(MilliporeSigma) and ultracentrifuging the serum at 100,000 × g
(4°C for 70 min), were supplemented with 10% exosome-free FBS.
After 24 h of incubation at 37°C, conditioned medium was collected
for exosome extraction. Exosomes derived from hADSCs (with or
without miR-29a mimics and inhibitor transfection) were extracted
and purified using a Cell Culture Media Exosome Purification Mini
kit (Norgen Biotek Corp.), according to the manufacturer's
instructions. To observe the ultrastructure of exosomes, 10 µl
exosome samples were added to the copper mesh for precipitation for
1 min, and then the floating solution was removed by filter paper.
Subsequently, 10 µl uranyl acetate was added to the copper mesh to
precipitate for 1 min, and then the floating solution was removed
using filter paper. After drying at room temperature for several
minutes, the image was detected by transmission electron microscopy
(Libra 120; Zeiss AG) at 100 kV. The size distribution and
concentration of exosomes were determined via nanoflow cytometry
(N30 NanoAnalyzer; NanoFCM, Inc.). Antibodies against exosomes
susceptibility (TSG)101 (1:1,000; cat. no. ab125011; Abcam), CD63
(1:1,000; cat. no. ab134045; Abcam) and CD81 (1:1,000; cat. no.
ab109201; Abcam), which are representative markers of exosomes, and
the cellular protein actin (1:5,000; cat. no. ab6276; Abcam) were
used to identify the collected exosomes via western blotting or
nanoflow cytometry.
Effect of hADSCs-exo on the
proliferation of HSFBs
HSFBs were precultured in 6-well plates
(1×104 cells/well) in DMEM containing 10% FBS and 1%
(v/v) penicillin/streptomycin for 4 h. Then, 20 µg miR-29a-modified
hADSCs-exo (mimics-exo or inhibitor-exo) was added to the culture
medium of HSFBs. An equivalent volume of exosome diluent PBS was
added as the control group. After the exosomes were added, the
cells were incubated at 37°C with 5% CO2 and imaged at
0, 24 and 48 h using an inverted-phase contrast light microscope
(Olympus Corporation; magnification, ×10) to observe the
proliferation of the cells.
In addition, whether HSFBs can directly take up in
hADSCs-exo was detected via immunofluorescence labeling. Exosomes
were labeled with the red fluorescent linker PKH67 (Sigma-Aldrich;
Merck KGaA), as previously reported (37). HSFBs were treated with PKH67-labeled
exosomes for 24 h at 37°C with 5% CO2. DAPI dye (cat.
no. ab104139; Abcam) was used for nuclear staining for 10 min at
room temperature. After washing by distilled water to remove the
uninternalized exosomes and excess DAPI dye, the fluorescent images
were visualized using a confocal microscope (Olympus Corporation;
magnification, ×40).
Scratch wound assay
As previously described, an in vitro scratch
wound assay was performed to evaluate cell migration (38). Briefly, HSFBs were cultured in
6-well culture plates (1×104 cells/well) along with DMEM
containing 10% FBS until 100% confluency, and then a scratch was
created in the middle of the culture dish on each well using a
200-µl pipette tip (Pipet Tip Finder, LLC). The wound was 0.45–0.50
mm in width per well. Next, the cultures were switched to
serum-free medium, and 20 µg miR-29a-modified hADSCs-exo
(mimics-exo or inhibitor-exo) was added to the culture medium for
24 and 48 h. Digital images of each wound were acquired under an
inverted-phase contrast microscope (Olympus Corporation;
magnification, ×10) at 0, 24 and 48 h after scratching. Wound
closure (cell migration) was investigated by measuring the wound
area using the commercial software ImageJ (version 1.52a; National
Institutes of Health). The results are presented as the percentage
of the initial wound area using the following formula: Scratch
healing rate (%) = (Area24 h (o Area48
h)-Area0 h)/Area0 h × 100%. Images of
each wound were acquired in three random views, and the mean cell
migration rate of each sample ± SD was presented as the final
result. The assay was repeated in three pooled cell samples.
Dual-luciferase reporter assay
The binding sites between miR-29a and TGF-β2 were
predicted with StarBase (version 2.0; http://starbase.sysu.edu.cn/agoClipRNA.php?source=mRNA).
Luciferase vectors containing the 3′UTR of human TGF-β2 with
miR-29a binding sites and mutant miR-29a binding sites were
purchased from Shanghai GenePharma Co., Ltd. Then, 50 ng vectors
were co-transfected with 20 nM miR-29a mimics or mimics-NC into
1×104 HSFB cells using Lipofectamine® 2000
transfection reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
for 6 h in the cell incubator with 5% CO2 at 37°C. The
pRL-CMV vector containing the CMV enhancer and early promoter
elements of Renilla luciferase (Promega Corporation) was
used as an internal control. After 48 h, the luciferase reporter
activity was determined using a Dual-Luciferase®
Reporter Assay system (Promega Corporation). Firefly luciferase
activity was normalized to Renilla luciferase activity.
TGF-β2/Smad3 agonist and TGF-β
signaling inhibitor treatment
To verify whether miR-29a modified hADSCs-exo
altered the fibrosis of HSFBs via TGF-β2/Smad3 signaling, the
TGF-β2/Smad3 agonist SRI-011381 hydrochloride (10 µM; cat. no.
HY-100347A; MedChemExpress) was used to co-treat HSFBs with
mimics-exo, and the TGF-β signaling inhibitor pirfenidone (AMR69; 3
mM; cat. no. HY-B0673; MedChemExpress) was used to co-treat HSFBs
with inhibitor-exo. Then the cells were incubated at 37°C with 5%
CO2 for 24 h prior to analysis via western blotting.
Animals and experimental protocol
To investigate the effect of miR-29a-modified
hADSCs-exo on skin wound healing and scar formation, a scald skin
model was established in mice according to a previous report
(33). A total of 36 adult male
Kunming mice (age, 6–8 weeks; weight, 22±2 g) from the same
generation were purchased from Hunan SJA Laboratory Animal Co.,
Ltd. (certificate no. 43004700043639). The mice were randomly
divided into four groups: Control group (n=12), thermal injury
group (thermal; n=12), exosomes from miR-29a overexpressed hADSCs
treated group (mimics-exo; n=6) and mimics-exo co-treated with the
TGF-β agonist SRI-011381 hydrochloride group (mimics-exo +
SRI-011381; n=6). Mice were housed individually with a 12-h
light/dark cycle at 22°C with 50% humidity and received ad
libitum food and water. All experiments were approved by the
Animal Ethics Committee of the First Affiliated Hospital of Kunming
Medical University (approval no. kmmu2021188).
Before thermal injury, the mice were anesthetized
with an intraperitoneal injection of 1% sodium pentobarbital (40
mg/kg) and shaved to expose the dorsal skin. After disinfecting the
back skin with 75% alcohol, a plastic tube with a diameter of 2 cm
was placed on the exposed back skin, one end of which was tightly
connected with the skin. Boiling water at 100°C was injected from
the other end, and the tube and boiling water were removed after
continuous contact with the skin for 10 sec. After thermal injury,
the mice were housed individually, the mimics-exo and mimics-exo +
SRI-011381 groups were multidirectionally, subcutaneously injected
with 200 µg exosomes from miR-29a-overexpressing hADSCs, and the
mimics-exo + SRI-011381 group was intraperitoneally injected with
SRI-011381 (RayBiotech, Inc.) at a dosage of 30 mg/kg at the same
time. Meanwhile, the control and thermal injury groups were
subcutaneously injected with PBS at the same volume. All mice were
treated once every 3 days, and 15 days after thermal injury, all
mice were euthanized by an intraperitoneal injection of excessive
1% sodium pentobarbital (120 mg/kg), and scalded skin tissues were
collected for further tests.
RT-qPCR analysis
RT-qPCR analysis was performed on an ABI 7500
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.). All reactions were run in triplicate. The thermocycling
conditions used for qPCR were as follows: Initial denaturation at
95°C for 10 sec; followed by 40 cycles of 95°C for 10 sec and 60°C
for 30 sec. The 2−ΔΔCq method was used to calculate
relative expression levels (39),
and U6 was used to normalize the miRNA expression. Total RNA was
extracted from cells or tissues using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), and miRNA RT-qPCR was
carried out using a TaqMan™ MicroRNA Reverse Transcription kit
(Applied Biosystems; Thermo Fisher Scientific, Inc.) and TaqMan
Universal PCR Master mix (Applied Biosystems; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions. The
primer sequences used were as follows: miR-29a forward,
5′-CTAGCACCATCTGAAATCGGTTA-3′ and reverse,
5′-TGATTGGCTAAAGTCTACCAC-3′; and U6 forward,
5′-CTCGCTTCGGCAGCACATATACT-3′ and reverse,
5′-ACGCTTCACGAATTTGCGTGTC-3′.
Western blotting
The cells or tissues in each group were collected
and washed with PBS and RIPA protein lysis buffer (Invitrogen;
Thermo Fisher Scientific, Inc.) to extract the total proteins. A
BCA protein kit (Pierce; Thermo Fisher Scientific, Inc.) was used
to detect the protein concentration and purity. In total, 40 µg
protein was separated by 10% SDS-PAGE and then transferred to PVDF
membranes via the electric transfer method. The membranes were
blocked with a solution containing 5% skimmed milk powder for 1 h
at room temperature, after which diluted primary antibody was added
and the membranes were incubated at 4°C overnight. The membranes
were then washed three times with membrane washing buffer, and goat
anti-rabbit HRP-labeled secondary antibody (1:2,000; cat. no.
ab205718; Abcam) or goat anti-mouse HRP-labeled secondary antibody
(1:2,000; cat. no. ab205719; Abcam) was added. The blots were
incubated for 1 h at room temperature in the dark and then washed
three times with membrane washing buffer. Finally, ECL solution
(Bio-Rad Laboratories, Inc.) was added for color development, and
images were collected in a gel imaging system. The grayscale value
analysis was performed using ImageJ software (version 1.52a;
National Institutes of Health), and GAPDH was used as the loading
control. The primary antibodies used were as follows: CD63
(1:1,000; cat. no. ab134045; Abcam), CD81 (1:1,000; cat. no.
ab109201; Abcam), TSG101 (1:1,000; cat. no. ab125011; Abcam), actin
(1:5,000; cat. no. ab6276; Abcam), TGF-β2 (1 µg/ml; cat. no.
ab113670; Abcam), phosphorylated (p)-Smad3 (1:2,000; cat. no.
ab52903; Abcam), α-smooth muscle actin (α-SMA; 1:10,000; cat. no.
ab124964; Abcam), collagen I (Col-I; 1:1,000; cat. no. ab34710;
Abcam), Col-III (1:1,000; cat. no. ab7778; Abcam) and GAPDH
(1:1,000; cat. no. ab8245; Abcam).
Histological examinations
Scar tissue samples were fixed in 4%
paraformaldehyde for 24 h at 4°C and then dehydrated by washing
with a series of ethanol solutions (50% ethanol for 2 h; 70%
ethanol for 2 h; 80% ethanol for 2 h; 95% ethanol I for 2 h; 95%
ethanol II for 1.5 h; 100% ethanol I for 1 h; 100% ethanol II for
30 min) at room temperature. Subsequently, samples were embedded in
paraffin wax and cut into 5-µm sections for routine H&E and
Masson staining. H&E staining was performed by staining with
hematoxylin dye (10 min; room temperature) and eosin dye (1 min;
room temperature) to observe the morphological changes of scar
tissue. All slices were imaged using an inverted-phase contrast
light microscope (Olympus Corporation; magnification, ×4 and ×20).
Scar tissue fibrosis was detected by Masson staining using
hematoxylin dye (10 min; room temperature), masson dye (10 min;
room temperature) and 1% light green aqueous solution (5 min; room
temperature). ImageJ software (version 1.52a; National Institutes
of Health) was used to measure the epidermal thickness and
Masson-positive area.
Statistical analysis
Data are presented as the mean ± SEM. The results
from the experiments were analyzed using SPSS 20.0 (IBM Corp.). The
comparison of multiple groups and the comparison between the two
groups were performed by one-way ANOVA or two-way ANOVA followed by
Bonferroni post hoc test. A mixed two-way ANOVA followed by
Bonferroni post hoc test was also used where appliable. GraphPad
Prism 7 (GraphPad Software, Inc.) was used to plot the experimental
data. P<0.05 was considered to indicate a statistically
significant difference, and all experiments were repeated three
times.
Results
miR-29a expression in mouse scar
tissues and HSFBs
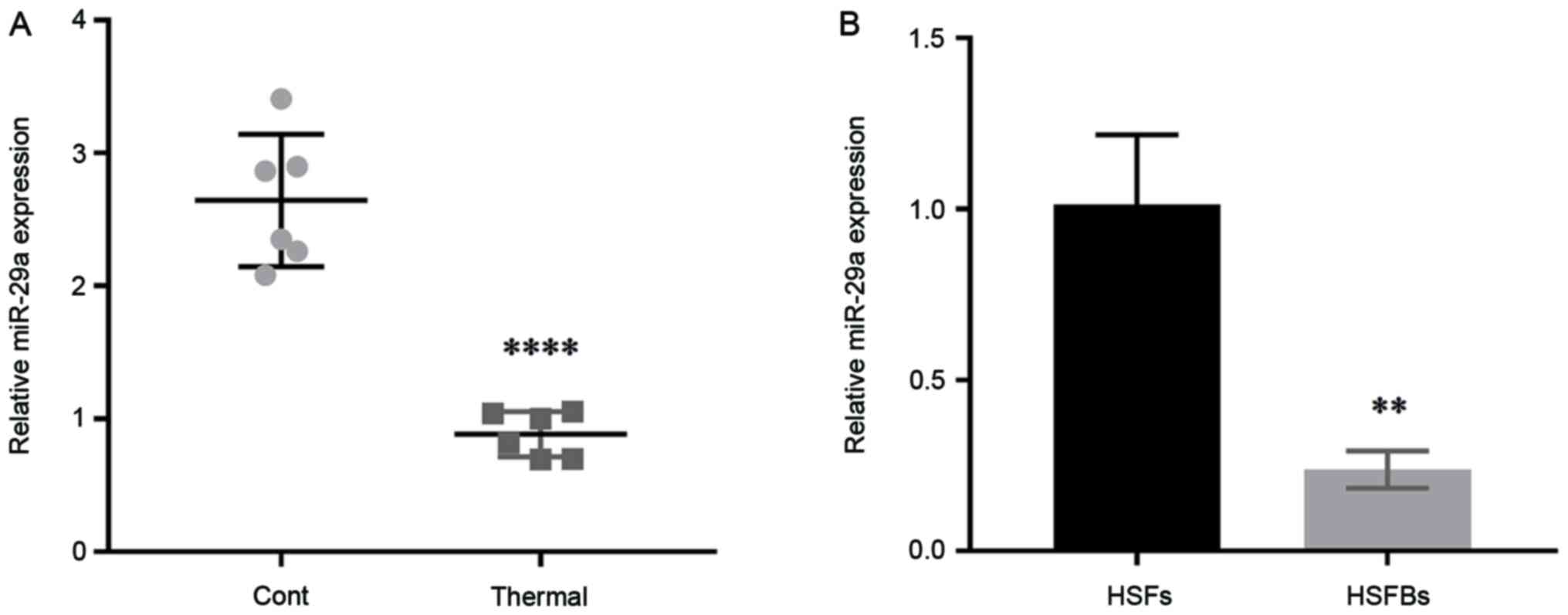
To determine the effect of miR-29a on scar
formation, the expression level of miR-29a was detected in scar
tissues from thermally injured mice and HSFBs, with normal mouse
skin tissues and HSFs used as controls. The results demonstrated
that the expression level of miR-29a in mouse scar tissues was
significantly downregulated compared with that in normal skin
tissues (Fig. 1A), and it was also
lower in HSFBs (Fig. 1B). These
results indicated that miR-29a may act as an inhibitor in the
formation of hypertrophic scars.
Extraction of hADSCs-exo and its
effect on miR-29a expression in HSFBs
First, exosomes were extracted and purified from
hADSCs, then TEM was used to identify the morphology of exosomes
extracted from hADSC cultures (Fig.
2A). The average particle size of exosomes was 81.14 nm and
96.62% of the total particles were 30–150 nm in diameter, and the
average concentration of exosomes was 1.43E + 10/ml (Fig. 2B).
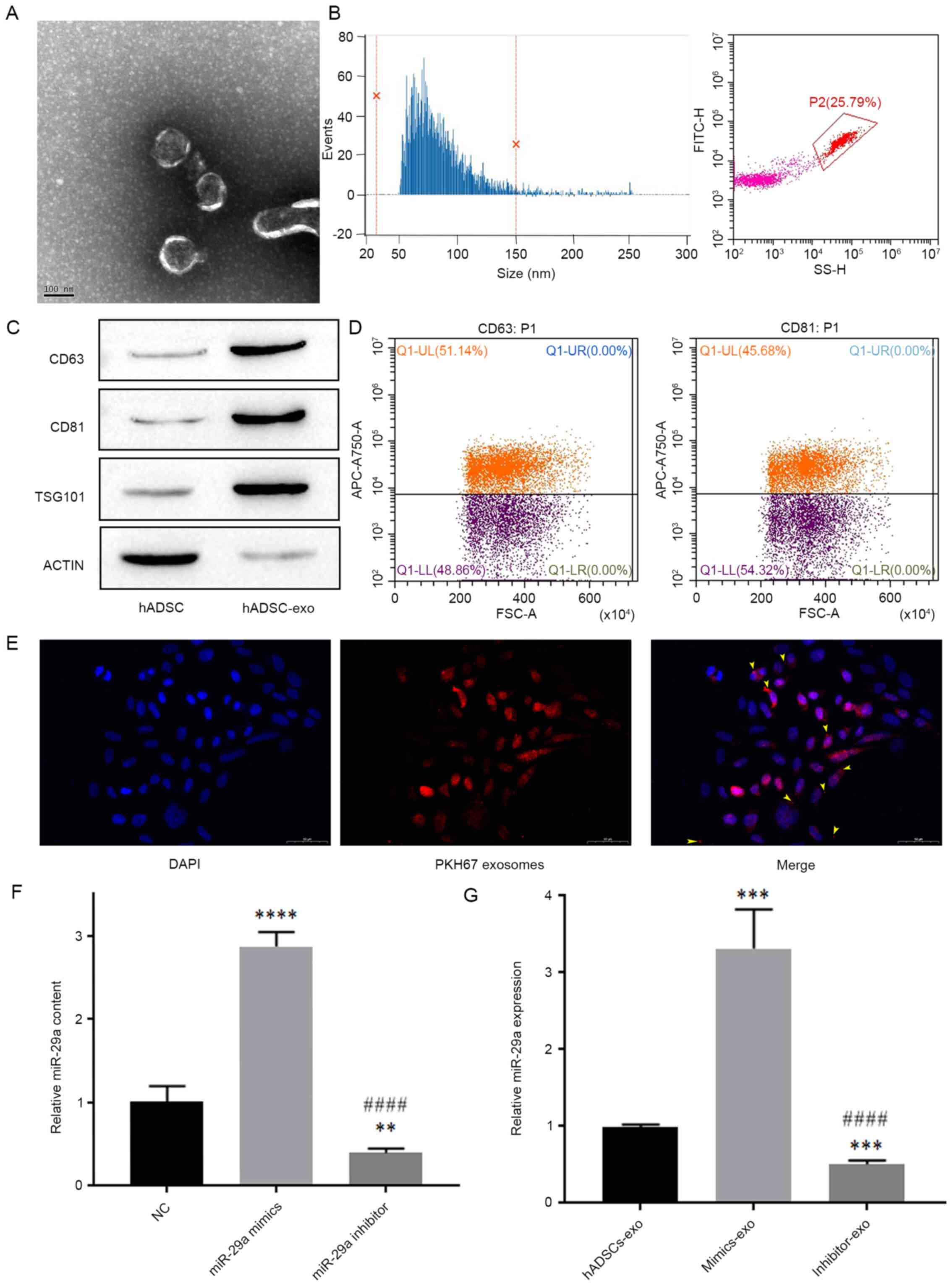 | Figure 2.Exosomes extracted from hADSCs and
their effect on the expression level of miR-29a in HSFBs. (A)
Transmission electron microscopy analysis of exosome
ultrastructure. Scale bar, 100 nm. (B) Nanoflow cytometry analysis
of particle size distribution and particle number. (C) Western
blotting detection of exosome biomarkers. (D) Flow cytometric
analysis of the expression levels of CD63 and CD81 in exosomes. (E)
Representative images of immunofluorescence labeling of exosomes in
HSFBs. Scale bar, 50 µm. (F) Content of miR-29a in different groups
of hADSCs-exo, as analyzed by one-way ANOVA followed by Bonferroni
post hoc test. **P<0.01, ****P<0.0001 vs. NC group;
####P<0.0001 vs. miR-29a mimics group. (G) The
influence of miR-29a-modified hADSCs-exo on the expression level of
miR-29a in HSFBs, as analyzed by one-way ANOVA followed by
Bonferroni post hoc test. ***P<0.001 vs. hADSCs-exo group;
####P<0.0001 vs. mimics-exo group. NC, negative
control; mimics-exo, exosomes from miR-29a overexpressed hADSCs
treated group; inhibitor-exo, exosomes from miR-29a knockdown
hADSCs treated group; miR, microRNA; hADSCs, human adipose-derived
mesenchymal stem cells; TSG101, tumor susceptibility 101. |
Next, the biomarkers of exosomes, including CD63,
CD81 and TSG101, and the cellular protein actin were detected via
western blotting. The results demonstrated that the expression
levels of CD63, CD81 and TSG101 were low in hADSCs, but were high
in hADSCs-exo, while the expression level of the cellular protein
actin was low in hADSCs-exo (Fig.
2C). Additionally, nanoflow cytometry revealed that the
positive rates of the exosome membrane proteins CD63 and CD81 were
both >30% (Fig. 2D), which
indicated that the exosomes were successfully extracted from hADSC
cultures.
Exosomes derived from miR-29a-overexpressing and
miR-29a-inhibited hADSCs were added to the culture medium of HSFBs,
and were labeled with an anti-PKH67 antibody, and DAPI was used to
label the nuclei of HSFBs to determine whether HSFBs can directly
take up hADSCs-exo. The results indicated that HSFBs can directly
take up hADSCs-exo (Fig. 2E). Then,
the expression levels of miR-29a in different groups of hADSCs-exo
and HSFBs were detected via RT-qPCR. First, it was found that
miR-29a mimics and inhibitors could effectively regulate the
content of miR-29a in hADSCs-exo after hADSCs were transfected with
miR-29a mimics or inhibitor (Fig.
2F). In addition, compared with the untreated
hADSCs-exo-treated group, the mimics-exo-treated group showed
higher expression of miR-29a, while the inhibitor-exo-treated group
showed lower expression of miR-29a (Fig. 2G). These results suggested that
miR-29a-modified hADSCs-exo can regulate the expression of miR-29a
in HSFBs.
Effect of miR-29a-modified hADSCs-exo
on the proliferation and migration of HSFBs
To evaluate the effect of miR-29a-modified
hADSCs-exo on HSFBs, HSFBs were divided into different groups and
treated with mimics-exo or inhibitor-exo. PBS was added as a NC,
and then the proliferation of HSFBs was imaged and observed. As
shown in Fig. 3A, HSFBs treated
with mimics-exo showed a lower proliferative ability, while the
proliferative rates of HSFBs in the NC and inhibitor-exo-treated
groups were higher. Furthermore, a cell scratch healing assay was
performed to determine the influence of miR-29a-modified hADSCs-exo
on HSFB migration. Compared with the NC group,
miR-29a-overexpressing hADSCs-exo significantly inhibited the
migration of HSFBs in the mimics-exo-treated group, while the
migration of HSFBs in the inhibitor-exo-treated group was enhanced
compared with that in the mimics-exo-treated group (Fig. 3B). These results indicated that
hADSC-exo-derived miR-29a can inhibit the proliferation and
migration of HSFBs.
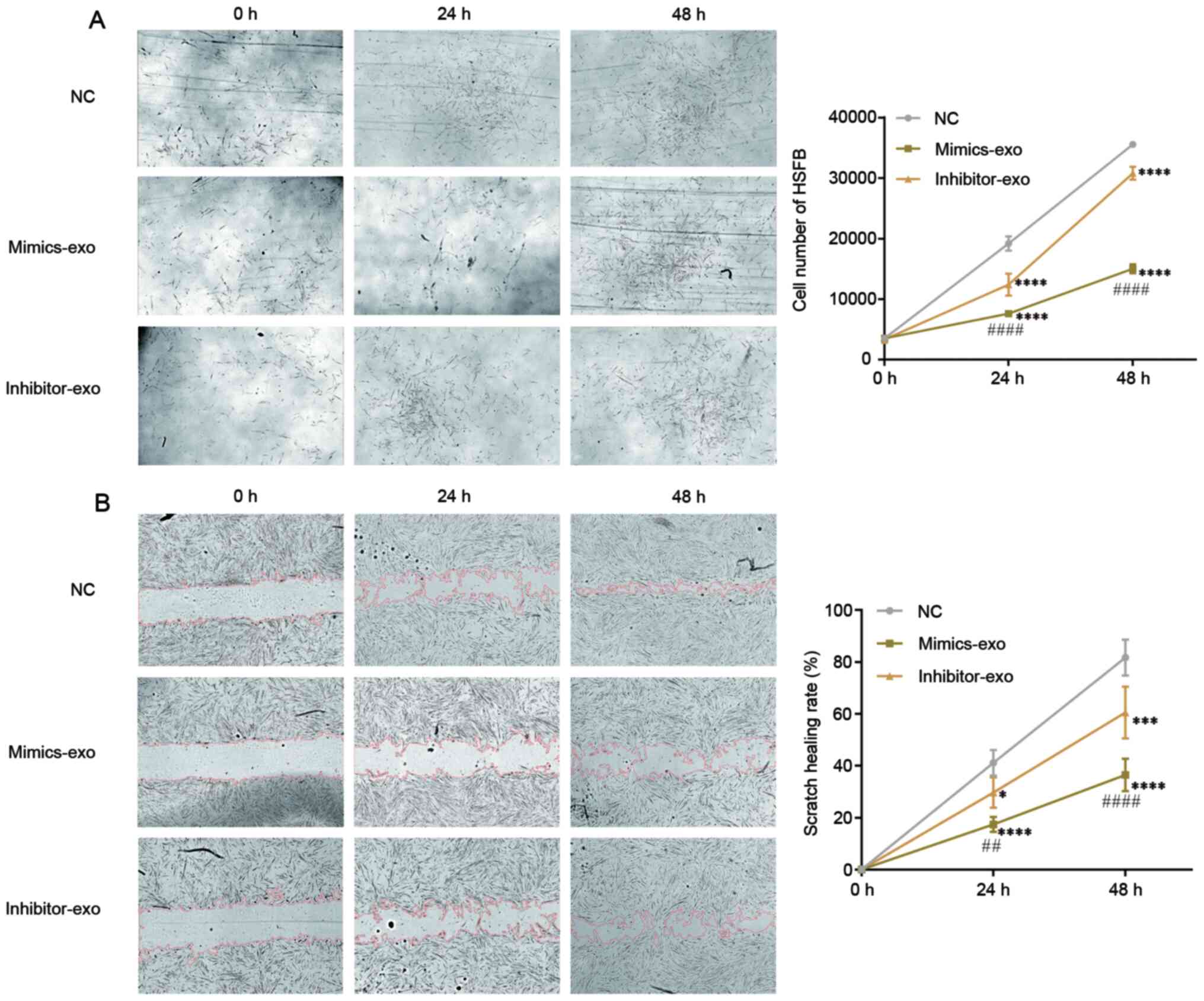 | Figure 3.Effect of miR-29a-modified hADSCs-exo
on the proliferation and migration of HSFBs. (A) Proliferation of
HSFBs treated with different hADSCs-exo. (B) Effect of
miR-29a-modified hADSCs-exo on the scratch healing of HSFBs
(magnification, ×10). Two-way ANOVA followed by Bonferroni post hoc
test was used to compare the differences among different groups.
*P<0.05, ***P<0.001, ****P<0.0001 vs. NC at the same time
point; ##P<0.01, ####P<0.0001 vs.
inhibitor-exo group at the same time point. NC, negative control;
mimics-exo, exosomes from miR-29a overexpressed hADSCs treated
group; inhibitor-exo, exosomes from miR-29a knockdown hADSCs
treated group; miR, microRNA; hADSCs, human adipose-derived
mesenchymal stem cells; HSFBs, human hypertrophic scar
fibroblasts. |
miR-29a-modified hADSCs-exo regulates
the TGF-β2/Smad3 signaling pathway in HSFBs
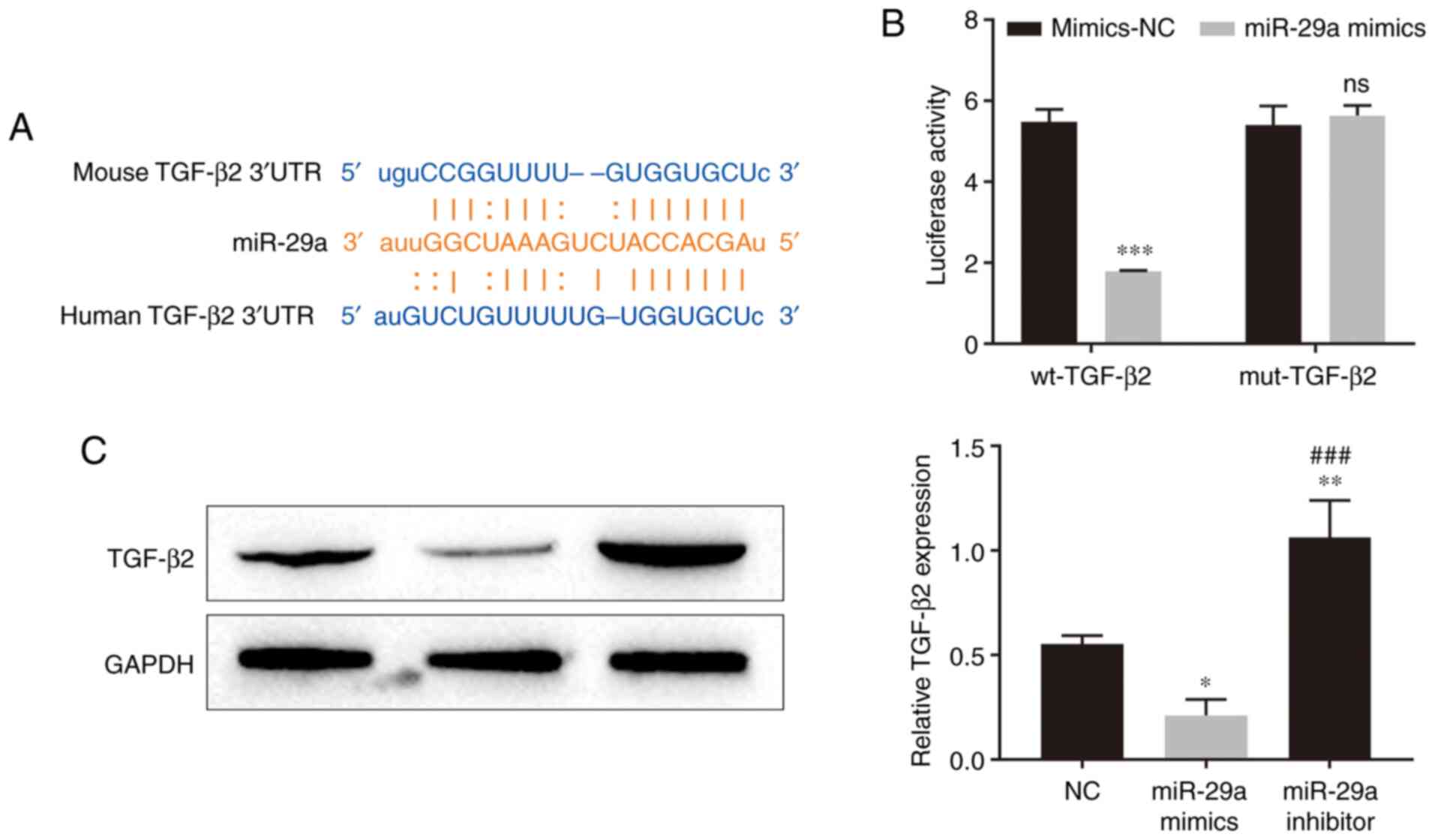
To verify whether hADSC-exo-derived miR-29a inhibits
HSFB proliferation and migration via the TGF-β/Smad signaling
pathway, the targets of miR-29a were scanned in the StarBase online
database, and TGF-β2 was predicted to be one of the targets of
miR-29a. There was a binding site between miR-29a and the 3′UTR of
TGF-β2 mRNA (Fig. 4A). Further
results from the dual-luciferase reporter assay revealed that
miR-29a mimics inhibited the luciferase activity of the wild-type
TGF-β2 3′UTR (Fig. 4B), which
indicated that miR-29a can directly bind to the TGF-β2 3′UTR. The
western blotting results demonstrated that miR-29a mimic
transfection significantly inhibited the expression level of TGF-β2
in HSFBs, while miR-29a inhibitor transfection upregulated TGF-β2
expression in HSFBs (Fig. 4C).
These results suggested that miR-29a can inhibit the expression of
TGF-β2 by binding to the 3′UTR of TGF-β2 mRNA.
To test whether miR-29a-modified hADSCs-exo inhibit
collagen deposition and the fibrosis of HSFBs via the TGF-β2/Smad3
signaling pathway, the TGF-β2/Smad3 agonist SRI-011381
hydrochloride (10 µM) and the TGF-β signaling inhibitor pirfenidone
(AMR69; 3 mM) were used to co-treat HSFBs with mimics-exo and
inhibitor-exo, respectively. Western blot analysis was performed to
evaluate the expression levels of TGF-β2, p-Smad3, Col-I, Col-III
and the fibrotic gene α-SMA in different groups of HSFBs. The data
demonstrated that mimics-exo significantly inhibited the expression
levels of TGF-β2 and p-Smad3 (Fig.
5A-C), as well as downregulated the expression levels of Col-I,
Col-III and α-SMA (Fig. 5A and
D-F), while SRI-011381 reversed the inhibition of
miR-29a-overexpressing hADSCs-exo on the activation of TGF-β2/Smad3
signaling and the fibrosis of HSFBs (Fig. 5). Moreover, AMR69 and inhibitor-exo
cotreatment enhanced the expression level of p-Smad3 (Fig. 5A-C) and upregulated the expression
levels of Col-I and Col-III and compared with the
mimics-exo-treated group (Fig. 5A and
D-F), whereas there was no significant difference in the
expression of TGF-β2 and α-SMA between the AMR69/inhibitor-exo
cotreatment and mimics-exo-treated groups. Thus, inhibition of the
TGF-β2/Smad3 signaling pathway may be a potential mechanism via
which miR-29a-modified hADSCs-exo reduce excessive scar formation
during skin wound healing.
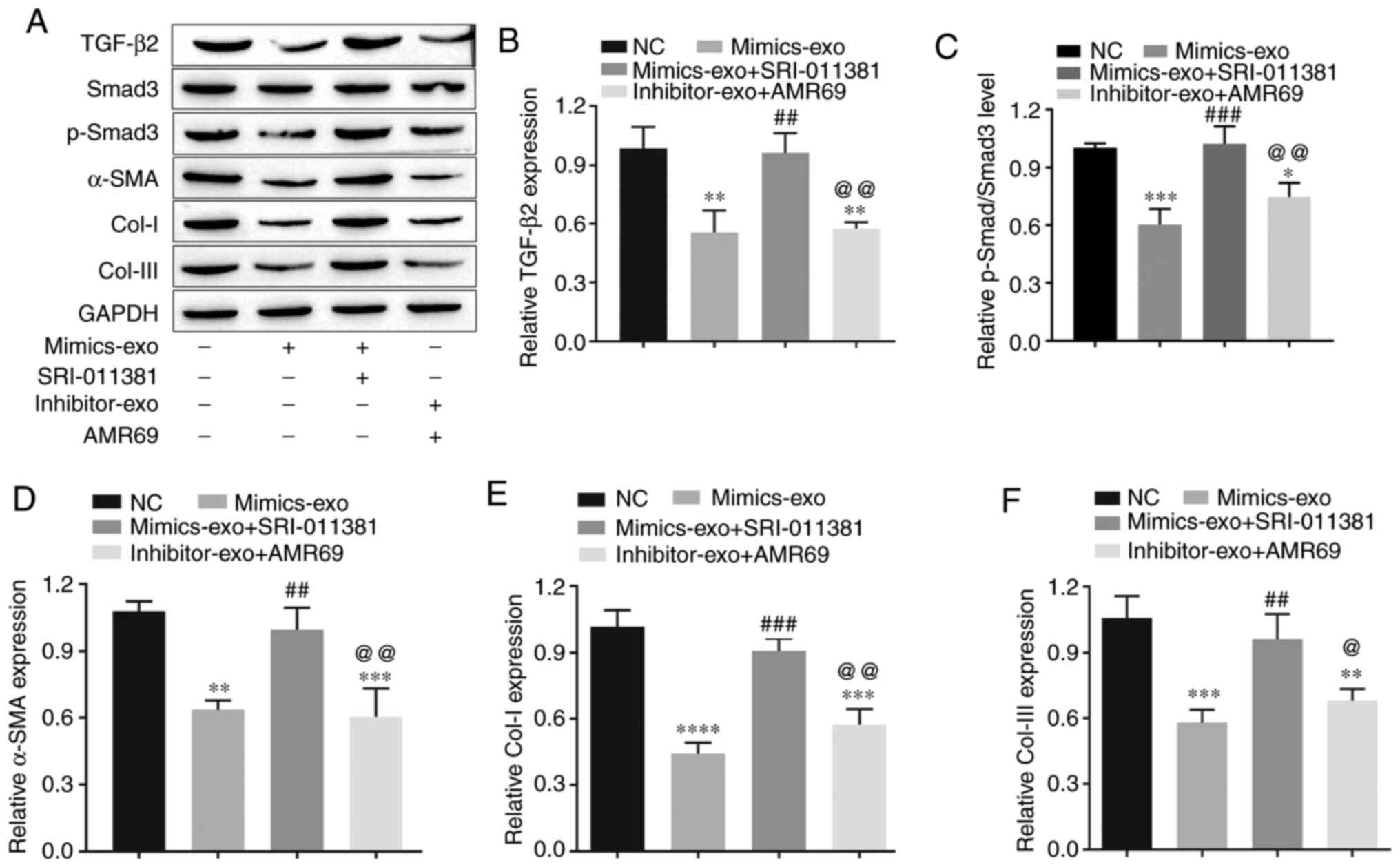 | Figure 5.Effects of miR-29a-modified
hADSCs-exo on the TGF-β2/Smad3 signaling pathway and fibrosis of
human hypertrophic scar fibroblasts. (A) Western blotting results.
(B) Relative expression level of TGF-β2. (C) Relative expression
level of p-Smad3. (D) Relative expression level of the fibrosis
gene α-SMA. (E) Relative expression level of Col-I. (F) Relative
expression level of Col-III. One-way ANOVA followed by Bonferroni
post hoc test was used to compare the differences among different
groups. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001 vs.
NC group; ##P<0.01, ###P<0.001 vs.
mimics-exo group; @P<0.05, @@P<0.01 vs.
mimics-exo + SRI-011381 group. Col-I, collagen I; Col-III, collagen
III; miR, microRNA; NC, negative control; α-SMA, α-smooth muscle
actin; p-, phosphorylated; mimics-exo, exosomes from miR-29a
overexpressed hADSCs treated group; inhibitor-exo, exosomes from
miR-29a knockdown hADSCs treated group; hADSCs, human
adipose-derived mesenchymal stem cells. |
Effects of miR-29a-overexpressing
hADSCs-exo on scar formation in scalded mice
To evaluate the inhibitory effect of
miR-29a-overexpressing hADSCs-exo on scar formation, a scalded skin
mouse model was established, and miR-29a mimics-exo was
subcutaneously injected around the wound. From the gross
observation of scar formation and wound healing after thermal
injury, miR-29a-overexpressing hADSCs-exo showed an obvious effect
on promoting the healing of thermal injury, and SRI-011381 reversed
the effect of miR-29a-overexpressing hADSCs-exo (Fig. 6A and B). As shown in Fig. 6C, the expression level of miR-29a
was significantly downregulated in the thermally injured group.
Subcutaneous injection of miR-29a-overexpressing hADSCs-exo led to
an upregulation of miR-29a, and the agonist of TGF-β, SRI-011381,
had no obvious effect on the expression level of miR-29a in the
skin tissues.
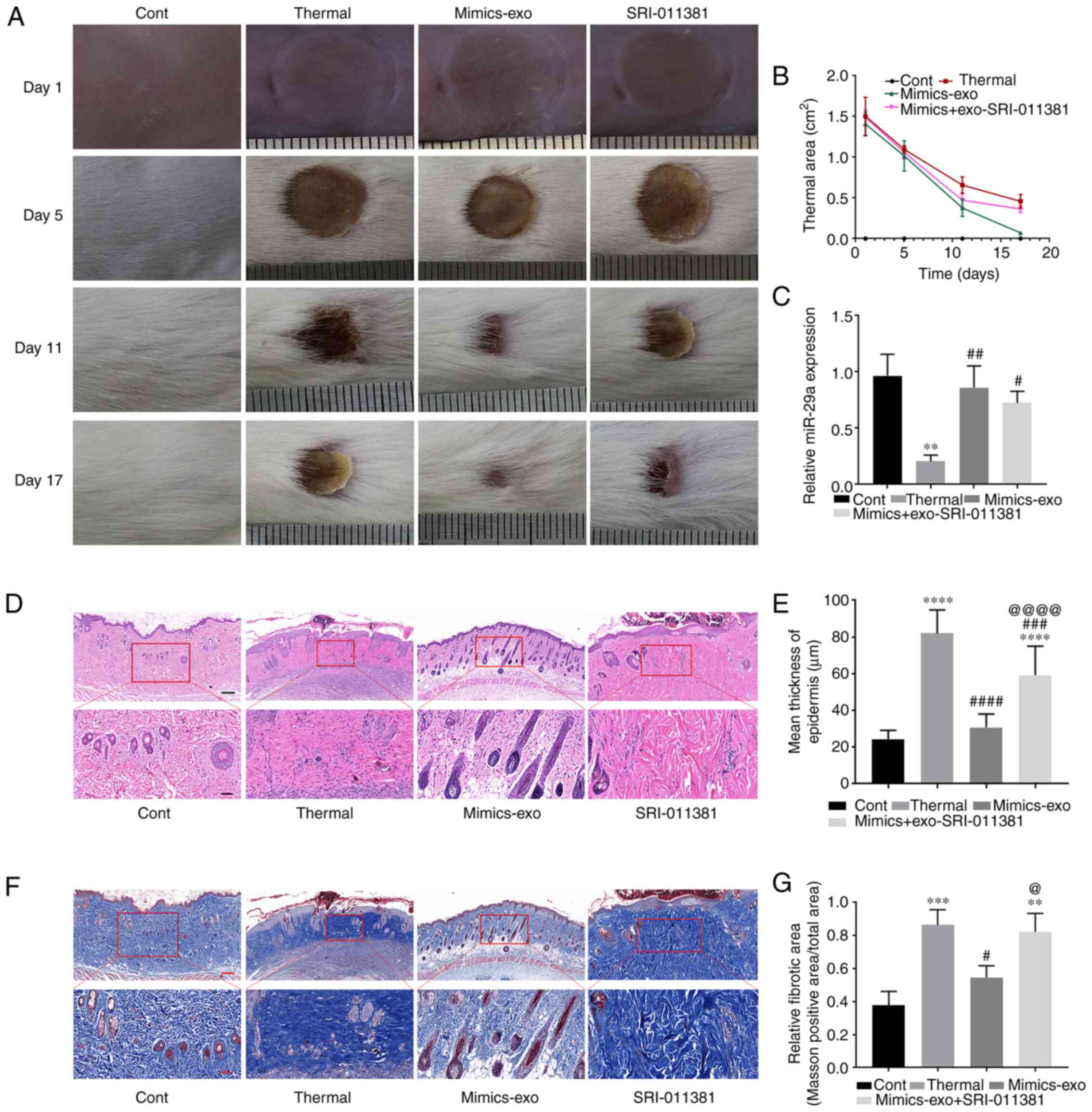 | Figure 6.Effect of miR-29a-overexpressing
hADSCs-exo on scar formation in thermally injured mice. (A)
Representative images of thermal injury of mice on days 1, 5, 11
and 17 after injury. (B) Thermal areas in each group. (C) miR-29a
expression in thermally injured tissues of different groups. (D)
Representative images of H&E staining of scar tissues. (E)
Epidermal thicknesses in different groups. (F) Representative
images of Masson staining of scar tissues. (G) Relative fibrotic
areas in different groups. Bars in the upper panel of D and F
indicate 200 µm, while the bar indicates 50 µm in the lower panel.
One-way ANOVA followed by Bonferroni post hoc test was used to
compare the differences among different groups. **P<0.01,
***P<0.001, ****P<0.0001 vs. Cont group;
#P<0.05, ##P<0.01,
###P<0.001, ####P<0.0001 vs. thermal
group; @P<0.05, @@@@P<0.0001 vs.
mimics-exo group. Cont, control; mimics-exo, exosomes from miR-29a
overexpressed hADSCs treated group; inhibitor-exo, exosomes from
miR-29a knockdown hADSCs treated group; hADSCs, human
adipose-derived mesenchymal stem cells; miR, microRNA. |
Next, histopathological morphological changes in
scalded tissue were determined via H&E staining. Thick dermis
and disordered granulation tissue, obvious infiltration of
inflammatory cells, dense collagen fibers and irregular arrangement
of collagen bundles were observed in thermally injured tissues.
Compared with the thermal injury group, miR-29a-overexpressing
hADSCs-exo markedly reduced the aforementioned morphological
changes, while SRI-011381 reversed the inhibitory effect of
mimics-exo on scar formation (Fig.
6D). Furthermore, Fig. 6E shows
the epidermal thickness measurement results in each group,
suggesting that miR-29a-overexpressing hADSCs-exo could inhibit
epidermal overgrowth during skin wound repair.
Masson staining was performed to observe the
fibrosis of the tissues (Fig. 6F and
G). The dermis of thermally injured tissue was mainly composed
of irregularly dense and disordered blue-stained collagen fibers,
while the mimics-exo treatment group showed more regular loose
collagen fibers. Moreover, the deposition of collagen fibers in the
SRI-011381 treatment group was similar to that in the thermal
injury group.
Effect of miR-29a-overexpressing
hADSCs-exo on the TGF-β2/Smad3 signaling pathway in scar
tissues
To further verify the molecular mechanism by which
hADSC-exo-derived miR-29a inhibits scar formation via the
TGF-β2/Smad3 signaling pathway, the protein expression levels of
TGF-β2, p-Smad3, the fibrosis gene α-SMA and two major
representative collagens, Col-I and Col-III, were measured in
different groups of scar tissues. As shown in Fig. 7A-C, compared with the control group,
the TGF-β2/Smad3 signaling pathway was markedly activated in
thermally injured tissues, and mimics-exo treatment inhibited the
activation of TGF-β2/Smad3 signaling, while SRI-011381 reversed the
inhibition of mimics-exo on this pathway. Furthermore, the
expression levels of α-SMA, Col-I and Col-III were significantly
inhibited by miR-29a-overexpressing hADSCs-exo compared with the
thermal injury group, but the agonist of TGF-β2/Smad3, SRI-011381,
reduced this inhibition (Fig. 7A and
D-F). These results indicated that miR-29a-overexpressing
hADSCs-exo inhibited excessive scar formation by inhibiting the
activation of the TGF-β2/Smad3 signaling pathway.
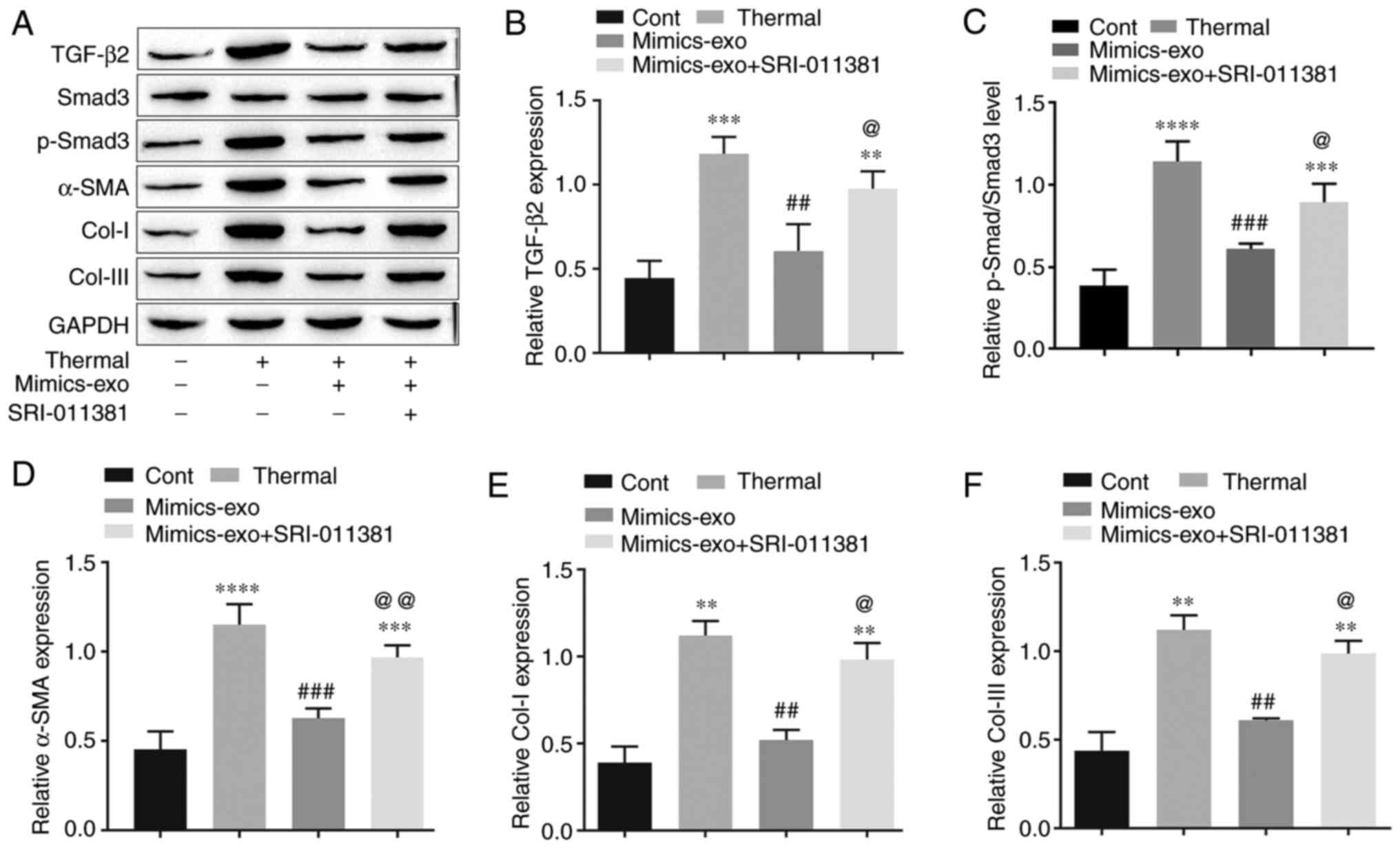 | Figure 7.Effect of microRNA-29a-modified
hADSCs-exo on the TGF-β2/Smad3 signaling pathway in scar tissues.
(A) Western blotting results. (B) Relative expression level of
TGF-β2. (C) Relative expression level of p-Smad3. (D) Relative
expression level of the fibrosis gene α-SMA. (E) Relative
expression level of Col-I. (F) Relative expression level of
Col-III. One-way ANOVA followed by Bonferroni post hoc test was
used to compare the differences among different groups.
**P<0.01, ***P<0.001, ****P<0.0001 vs. Cont group;
##P<0.01, ###P<0.001 vs. thermal group;
@P<0.05, @@P<0.01 vs. mimics-exo group.
Col-I, collagen I; Col-III, collagen III; Cont, control; α-SMA,
α-smooth muscle actin; p-, phosphorylated; mimics-exo, exosomes
from miR-29a overexpressed hADSCs treated group; hADSCs,
humanadipose-derived mesenchymal stem cells. |
Discussion
During wound healing, scar formation and hyperplasia
often occur, not only affecting aesthetics but also causing pain,
itching and other symptoms. These scars and hyperplasia may also
have negative impacts on the patient's physical, psychological and
social functions, and have become an urgent problem to be solved in
the field of plastic surgery (4).
Hypertrophic scars or keloids are more common types of pathological
scars in clinical practice, and their incidence is increasing, with
~11 million patients with keloids in high-income countries in 2000
(40). The excessive proliferation
and fibrosis of fibroblasts caused by the activation of the
TGF-β/Smad signaling pathway contribute primarily to the formation
of pathological scars (2,32). The present study demonstrated that
miR-29a-modified hADSCs-exo inhibited the proliferation and
migration of HSFBs and reduced excessive scar formation via the
targeted inhibition of the TGF-β2/Smad3 signaling pathway.
Several studies have reported that only a small part
of the tissue repair function of stem cells involves the
proliferation and differentiation of stem cells in the damaged
area, while most of them function via paracrine signaling (41). Exosomes reflect not only the
physiological state of the source cells, but also the
differentiation direction of secreted cells. The miRNAs contained
in exosomes are an important medium for intercellular
communication; after exosomes enter target cells, they are
regulated by degradation and re-expression and alter the genes of
target cells (13). The expression
profile of RNA in cells not only reflects donor specificity, but
the RNA contained in exosomes is affected by the type of
differentiated cells; thus, exosomes from different cell sources
express different biological characteristics (42). Previous studies have also confirmed
that MSC-derived exosomes, including adipose stem cells and
umbilical cord stem cells, have the ability to promote wound
healing in animal models (43,44).
These functions mainly depend on miRNAs carried by exosomes, such
as miR-31 and miR-125a, as stimulation signals, which promote the
migration, proliferation and angiogenesis of endothelial cells
(45,46). It has also been revealed that
exosomes can reduce scar formation and the proliferation of
myofibroblasts in a mouse model of skin defects. High-throughput
sequencing and functional analysis have demonstrated that exosomes
carrying miR-21, miR-23a, miR-125b and miR-145 inhibit the
TGF-β2/Smad2 signaling pathway, thereby inhibiting the formation of
myofibroblasts and preventing scar formation (47). The current study revealed that
hADSCs-exo inhibited pathological scar formation in scalded mice by
transmitting exogenous miR-29a, thus reducing collagen deposition
in tissues.
TGF-β serves an important role and mediates a
complex mechanism in the process of scar formation. It participates
in the regulation of a variety of biological substances and
response processes, including extracellular matrix and proteases,
as well as the proliferation, differentiation and migration of
various cells (47). Currently, it
is considered that during scar formation, TGF-β can regulate the
activation of downstream Smad signaling pathways (48,49).
Moreover, any abnormal expression of the TGF-β/Smad signaling
pathway may cause scar hyperplasia. A variety of molecules and
proteins have been shown to promote or inhibit the transmission of
signals, such as Toll-like receptors, miRNAs, TNF receptor-related
TNF receptor-associated protein 1-like protein and Smad interacting
protein 1. The abnormal expression of miRNAs during fibrosis can
serve a promotive or inhibitory role by participating in various
processes in inflammatory pathways or the immune response. For
example, miRNAs can promote or inhibit various links in the
TGF-β/Smad signaling pathway to promote or inhibit scar formation
(49). The results of the present
study also indicated that miR-29a-modified hADSCs-exo therapy can
inhibit the TGF-β2/Smad3 signaling pathway via its derived miR-29a
to inhibit collagen deposition and extracellular matrix fibrosis to
further inhibit scar formation.
In conclusion, the present study provided evidence
that miR-29a-overexpressing hADSC-derived exosomes can inhibit the
TGF-β2/Smad3 signaling pathway in wound tissues to promote wound
healing and reduce pathological scar formation by transmitting
exogenous miR-29a to wound tissues. These results suggested that
miR-29a-modified hADSCs-exo may be a potential therapeutic for skin
wound healing and pathological scars.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the Yunnan
Science and Technology Department-Kunming Medical University Joint
Fund [grant no. 2017FE467(−051)].
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
All authors contributed substantially to this
manuscript. RY contributed primarily to this manuscript by
participating in all the experiments and writing the manuscript. XD
participated primarily in the in vivo testing. YL
contributed mainly to the in vitro testing. CL contributed
to the data collation and analysis. LL contributed to the
conception and design of this study, and guided the writing of the
manuscript. RY and LL confirmed the authenticity of the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent for
publication
All experimental mouse protocols were approved by
the Animal Ethics Committee of The First Affiliated Hospital of
Kunming Medical University (approval no. kmmu2021188), and the
animals were handled according to the management requirements of
the Animal Management Association of The First Affiliated Hospital
of Kunming Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Schultz G, Davidson J, Kirsner R,
Bornstein P and Herman I: Dynamic reciprocity in the wound
microenvironment. Wound Repair Regen. 19:134–148. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karppinen S, Heljasvaara R, Gullberg D,
Tasanen K and Pihlajaniemi T: Toward understanding scarless skin
wound healing and pathological scarring. F1000Res. 8:F10002019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brown BC, Moss TP, McGrouther DA and Bayat
A: Skin scar preconceptions must be challenged: Importance of
self-perception in skin scarring. J Plast Reconstr Aesthet Surg.
63:1022–1029. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
van den Broek LJ, Limandjaja GC, Niessen
FB and Gibbs S: Human hypertrophic and keloid scar models:
Principles, limitations and future challenges from a tissue
engineering perspective. Exp Dermatol. 23:382–386. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu J, Wang MY, Tai HC and Cheng NC: Cell
sheet composed of adipose-derived stem cells demonstrates enhanced
skin wound healing with reduced scar formation. Acta Biomater.
77:191–200. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu SC, Bamodu OA, Kuo KT, Fong IH, Lin
CC, Yeh CT and Chen SG: Adipose-derived stem cell induced-tissue
repair or wound healing is mediated by the concomitant upregulation
of miR-21 and miR-29b expression and activation of the AKT
signaling pathway. Arch Biochem Biophys. 705:1088952021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ni X, Shan X, Xu L, Yu W, Zhang M, Lei C,
Xu N, Lin J and Wang B: Adipose-derived stem cells combined with
platelet-rich plasma enhance wound healing in a rat model of
full-thickness skin defects. Stem Cell Res Ther. 12:2262021.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cai Y, Li J, Jia C, He Y and Deng C:
Therapeutic applications of adipose cell-free derivatives: A
review. Stem Cell Res Ther. 11:3122020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Basu J and Ludlow JW: Exosomes for repair,
regeneration and rejuvenation. Expert Opin Biol Ther. 16:489–506.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Merino-González C, Zuñiga FA, Escudero C,
Ormazabal V, Reyes C, Nova-Lamperti E, Salomón C and Aguayo C:
Mesenchymal stem cell-derived extracellular vesicles promote
angiogenesis: Potencial clinical application. Front Physiol.
7:242016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Konala VB, Mamidi MK, Bhonde R, Das AK,
Pochampally R and Pal R: The current landscape of the mesenchymal
stromal cell secretome: A new paradigm for cell-free regeneration.
Cytotherapy. 18:13–24. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Casado-Díaz A, Quesada-Gómez JM and Dorado
G: Extracellular vesicles derived from mesenchymal stem cells (MSC)
in regenerative medicine: Applications in skin wound Healing. Front
Bioeng Biotechnol. 8:1462020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rani S and Ritter T: The exosome - A
naturally secreted nanoparticle and its application to wound
healing. Advanced materials (Deerfield Beach Fla.). 28:5542–5552.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rani S, Ryan A, Griffin M and Ritter T:
Mesenchymal stem cell-derived extracellular vesicles: Toward
cell-free therapeutic applications. Molecular therapy : the journal
of the American Society of Gene Therapy. 23:812–823. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shabbir A, Cox A, Rodriguez-Menocal L,
Salgado M and Van Badiavas E: Mesenchymal stem cell exosomes induce
proliferation and migration of normal and chronic wound
fibroblasts, and enhance angiogenesis in vitro. Stem Cells Dev.
24:1635–1647. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Diegelmann R and Evans M: Wound healing:
an overview of acute, fibrotic and delayed healing. Front Biosci.
9:283–289. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yuan H, Guan J, Zhang J, Zhang R and Li M:
Exosomes secreted by human urine-derived stem cells accelerate skin
wound healing by promoting angiogenesis in rat. Cell Biol Int.
41:9332017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu L, Wang J, Zhou X, Xiong Z, Zhao J, Yu
R, Huang F, Zhang H and Chen L: Exosomes derived from human adipose
mensenchymal stem cells accelerates cutaneous wound healing via
optimizing the characteristics of fibroblasts. Sci Rep.
6:329932016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Liu L, Yang J, Yu Y, Chai J, Wang L,
Ma L and Yin H: Exosome derived from human umbilical cord
mesenchymal stem cell mediates MiR-181c attenuating burn-induced
excessive inflammation. EBioMedicine. 8:72–82. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tafrihi M and Hasheminasab E: MiRNAs:
Biology, biogenesis, their Web-based tools, and databases.
MicroRNA. 8:4–27. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ha M and Kim VN: Regulation of microRNA
biogenesis. Nat Rev Mol Cell Biol. 15:509–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dietrich C, Singh M, Kumar N and Singh SR:
The emerging roles of microRNAs in stem cell aging. Adv Exp Med
Biol. 1056:11–26. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheng J, Wang Y, Wang D and Wu Y:
Identification of collagen 1 as a post-transcriptional target of
miR-29b in skin fibroblasts: Therapeutic implication for scar
reduction. Am J Med Sci. 346:98–103. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Murali VP and Holmes CA: Mesenchymal
stromal cell-derived extracellular vesicles for bone regeneration
therapy. Bone Rep. 14:1010932021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ichinohe N, Ishii M, Tanimizu N, Mizuguchi
T, Yoshioka Y, Ochiya T, Suzuki H and Mitaka T: Extracellular
vesicles containing miR-146a-5p secreted by bone marrow mesenchymal
cells activate hepatocytic progenitors in regenerating rat livers.
Stem Cell Res Ther. 12:3122021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Du Y and Ning JZ: MiR-182 Promotes
ischemia/reperfusion-induced acute kidney injury in rat by
targeting FoxO3. Urol Int. 105:687–696. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ouyang Z and Wei K: miRNA in cardiac
development and regeneration. Cell Regen (Lond).
10:142021.PubMed/NCBI
|
|
28
|
Zhou J, Zhang X, Liang P, Ren L, Zeng J,
Zhang M, Zhang P and Huang X: Protective role of microRNA-29a in
denatured dermis and skin fibroblast cells after thermal injury.
Biol Open. 5:211–219. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Guo L and Huang X, Liang P, Zhang P, Zhang
M, Ren L, Zeng J, Cui X and Huang X: Role of XIST/miR-29a/LIN28A
pathway in denatured dermis and human skin fibroblasts (HSFs) after
thermal injury. J Cell Biochem. 119:1463–1474. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zgheib C, Hodges M, Hu J, Beason DP,
Soslowsky LJ, Liechty KW and Xu J: Mechanisms of mesenchymal stem
cell correction of the impaired biomechanical properties of
diabetic skin: The role of miR-29a. Wound Repair Regen. 24:237–246.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang Z, Feng C, Song K, Qi Z, Huang W and
Wang Y: lncRNA-H19/miR-29a axis affected the viability and
apoptosis of keloid fibroblasts through acting upon COL1A1
signaling. J Cell Biochem. 121:4364–4376. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang T, Wang X, Wang Z, Lou D, Fang QQ,
Hu YY, Zhao WY, Zhang LY, Wu LH and Tan WQ: Current potential
therapeutic strategies targeting the TGF-β/Smad signaling pathway
to attenuate keloid and hypertrophic scar formation. Biomed
Pharmacother. 129:1102872020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Guo J, Lin Q, Shao Y, Rong L and Zhang D:
miR-29b promotes skin wound healing and reduces excessive scar
formation by inhibition of the TGF-β1/Smad/CTGF signaling pathway.
Can J Physiol Pharmacol. 95:437–442. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhu Y, Li Z, Wang Y, Li L, Wang D, Zhang
W, Liu L, Jiang H, Yang J and Cheng J: Overexpression of miR-29b
reduces collagen biosynthesis by inhibiting heat shock protein 47
during skin wound healing. Transl Res. 178:38–53.e6. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang SJ, Yun CJ, Liu J, Yao SY, Li Y,
Wang M, Wang C, Bai YY and Xue H: MicroRNA-29a attenuates
angiotensin-II induced-left ventricular remodeling by inhibiting
collagen, TGF-β and SMAD2/3 expression. J Geriatr Cardiol.
17:96–104. 2020.PubMed/NCBI
|
|
36
|
Ragni E, Perucca Orfei C, De Luca P,
Viganò M, Colombini A, Lugano G, Bollati V and de Girolamo L:
miR-22-5p and miR-29a-5p are reliable reference genes for analyzing
extracellular vesicle-associated miRNAs in adipose-derived
mesenchymal stem cells and are stable under inflammatory priming
mimicking osteoarthritis condition. Stem Cell Rev Rep. 15:743–754.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang X, Li Z, Cui Y, Cui X, Chen C and
Wang Z: Exosomes isolated from bone marrow mesenchymal stem cells
exert a protective effect on osteoarthritis via lncRNA
LYRM4-AS1-GRPR-miR-6515-5p. Front Cell Dev Biol. 9:6443802021.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bian W, Meng B, Li X, Wang S, Cao X, Liu
N, Yang M, Tang J, Wang Y and Yang X: OA-GL21, a novel bioactive
peptide from Odorrana andersonii, accelerated the healing of
skin wounds. Biosci Rep. Jun 21–2018.(Epub ahead of print). doi:
10.1042/BSR20180215. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−ΔΔC(T)) method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Bayat A, McGrouther DA and Ferguson MW:
Skin scarring. BMJ. 326:88–92. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
New SE, Alvarez-Gonzalez C, Vagaska B,
Gomez SG, Bulstrode NW, Madrigal A and Ferretti P: A matter of
identity - Phenotype and differentiation potential of human somatic
stem cells. Stem Cell Res (Amst). 15:1–13. 2015. View Article : Google Scholar
|
|
42
|
Ren K: Exosomes in perspective: A
potential surrogate for stem cell therapy. Odontology. 107:271–284.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lopez-Verrilli MA, Caviedes A, Cabrera A,
Sandoval S, Wyneken U and Khoury M: Mesenchymal stem cell-derived
exosomes from different sources selectively promote neuritic
outgrowth. Neuroscience. 320:129–139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Baglio SR, Rooijers K, Koppers-Lalic D,
Verweij FJ, Pérez Lanzón M, Zini N, Naaijkens B, Perut F, Niessen
HW, Baldini N, et al: Human bone marrow- and adipose-mesenchymal
stem cells secrete exosomes enriched in distinctive miRNA and tRNA
species. Stem Cell Res Ther. 6:1272015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kang T, Jones TM, Naddell C, Bacanamwo M,
Calvert JW, Thompson WE, Bond VC, Chen YE and Liu D:
Adipose-derived stem cells induce angiogenesis via microvesicle
transport of miRNA-31. Stem Cells Transl Med. 5:440–450. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liang X, Zhang L, Wang S, Han Q and Zhao
RC: Exosomes secreted by mesenchymal stem cells promote endothelial
cell angiogenesis by transferring miR-125a. J Cell Sci.
129:2182–2189. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Fang S, Xu C, Zhang Y, Xue C, Yang C, Bi
H, Qian X, Wu M, Ji K, Zhao Y, et al: Umbilical cord-derived
mesenchymal stem cell-derived exosomal MicroRNAs suppress
myofibroblast differentiation by inhibiting the transforming growth
factor-β/SMAD2 pathway during wound healing. Stem Cells Transl Med.
5:1425–1439. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Valluru M, Staton CA, Reed MW and Brown
NJ: Transforming growth factor-β and endoglin signaling orchestrate
wound healing. Front Physiol. 2:892011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Gras C, Ratuszny D, Hadamitzky C, Zhang H,
Blasczyk R and Figueiredo C: miR-145 contributes to hypertrophic
scarring of the skin by inducing myofibroblast activity. Mol Med.
21:296–304. 2015. View Article : Google Scholar : PubMed/NCBI
|