Introduction
Heme oxygenase-1 (HO-1), also known as heat shock
protein 32, is the rate-limiting enzyme in the heme degradation
pathway that degrades hemoglobin into biliverdin, free iron and
carbon monoxide (CO) (1).
Biliverdin is subsequently reduced to bilirubin via biliverdin
reductase. Both biliverdin and bilirubin are bile pigments with
antioxidant properties (2).
Endogenous CO can act as a second messenger, thereby affecting a
variety of physiological and pathological processes, including cell
proliferation, inflammation, apoptosis and injury (3). As a cytoprotective enzyme, HO-1
serves an important role in maintaining cell homeostasis.
To date, three HO polymorphic isoenzymes have been
identified, namely HO-1, HO-2 and HO-3, of which HO-1 is inducible.
The HO-1-encoding gene is located on the long arm of chromosome
22q12 in the human body, with a total length of 6.8 kb (4). HO-1 is expressed in a variety of
tissues, including the liver, spleen, kidney, heart, lung, brain,
blood vessels, smooth muscle cells and endothelial cells, among
which the highest expression is observed in the liver, spleen and
kidney (5,6). Under normal physiological
conditions, the expression of HO-1 is relatively low in most
tissues. However, under various pathophysiological stress or
stimulation conditions, such as hypoxia, ultraviolet light,
inflammatory mediators, heme, ischemia and other harmful stimuli,
HO-1 expression is induced to protect cells against oxidative and
inflammatory damage (7,8). In the presence of the aforementioned
stimulatory factors, the induced expression of HO-1 is primarily
affected by redox-sensitive transcription factors, including
nuclear factor erythroid 2-related factor 2 (Nrf2), activator
protein 1 (AP1), hypoxia-inducible factor (HIF) and BTB and CNC
homology 1 (Bach1) (9). In
addition, a large number of epidemiological genetic studies have
shown that the expression of HO-1 is regulated by genetic
polymorphisms in the promoter of its gene, which might be
associated with a variety of diseases, such as coronary
atherosclerosis, rheumatoid arthritis and viral diseases (10–13).
The present review focuses on the clinical evidence
that HO-1 gene polymorphisms are associated with the incidence and
outcome of different diseases. To further understand this
association, a brief discussion of the regulation of HO-1-induced
expression mediated by transcription factors is included.
Regulation of HO-1 expression
As a stress-induced cytoprotective enzyme, the
investigation of the molecular mechanism underlying HO-1 regulation
began with studies on how various stress stimulation conditions,
including stress, hypoxia and small-molecule inducers, could
influence the activity of the enzyme. Under these stress
conditions, the expression of HO-1 is primarily regulated by
redox-sensitive transcription factors, including Nrf2, AP1, HIF and
Bach1. The present review discusses the mechanisms through which
each of these factors regulate HO-1.
Nrf2 is considered to be the primary transcription
factor that regulates the expression of HO-1 (14). Under basic conditions, Nrf2 can
form a protein complex with Kelch-like ECH-associated protein 1
(KEAP1) in the cytoplasm, which inhibits the activity of Nrf2 via
proteasome-mediated degradation of Nrf2 (15,16). When cells are exposed to chemical
or physical stimuli, Nrf2 separates from KEAP1 and transfers to the
nucleus, where it binds to MAF bZIP transcription factor (Maf). The
Nrf2/Maf heterodimers bind the antioxidant response element (ARE),
leading to the transcriptional activation of antioxidant genes,
including the HO-1 gene (17,18). Bach1, a heme sensor, acts as a
major transcriptional repressor of HO-1 expression. Bach1 impairs
the DNA-binding activity of Nrf2 by competing with Nrf2 for binding
to the AREs of numerous antioxidant genes under basal conditions.
High levels of heme in cells can cause Bach1 to dissociate from
AREs, leading to nuclear export and subsequent proteasome-mediated
degradation of Bach1. Under these circumstances, transcriptional
expression of HO-1 occurs (19,20). In general, in response to various
stress stimuli, the feedback loop formed by Nrf2, KEAP1 and Bach1
has a pivotal role in regulating the expression of HO-1.
Other transcriptional factors have been shown to be
involved in the regulation of HO-1 expression, including HIF-1, AP1
and NF-κB (21,22). Similarly to Nrf2, these factors
can combine with the cis-regulatory element of the HO-1
gene, resulting in an increase in HO-1 expression (21). HIF-1, a redox-responsive
transcription factor, induces HO-1 expression in different cell
types under hypoxic conditions (23–26). However, a specific mutant of HIF-1
lost its DNA-binding activity, which resulted in inhibition of HO-1
expression (27). Mechanistic
studies have shown that HIF-1α/HIF-1β heterodimers are necessary
for hypoxia-induced HO-1 expression, since hypoxia induces the
expression of HIF-1α, which binds to specific DNA sequences
dependent on HIF-1β (27). AP1
transcriptional factor is generated by dimers of the Jun and Fos
family proteins. AP1 is able to bind to
tetradecanoylphorbol-13-acetate-responsive elements of the HO-1
gene promoter (28,29), subsequently leading to the
upregulation of HO-1 expression, which has been demonstrated by
functional studies and sequence analysis of the promoter (21,30–32). Similarly to AP1, the NF-κB family
of transcription factors are dimerization complex factors formed by
REL proto-oncogene, NF-κB subunit family proteins. NF-κB homodimers
comprising p52 and p50 are transcription repressors, whereas NF-κB
heterodimers formed by p65 and p50 act primarily as transcription
activators (21,33). AP1 and NF-κB have been shown to
regulate HO-1 expression primarily in immune cells, thereby
fulfilling an important role in protecting against
inflammation-mediated damage, including that mediated by reactive
oxygen species (34).
In addition to the direct regulation mediated by a
range of transcription activators, a large number of HO-1 inducers
are also able to indirectly activate the expression of the protein
through acting on the protein kinase signaling pathway. Numerous
plant-derived polyphenols have been found to induce HO-1 gene
expression by activating the MAPK, PI3K/AKT and AMP-activated
protein kinase (AMPK) signaling pathways (35–37). For example, salvianolic acid A, a
plant-derived polyphenol that has been widely studied, provides
significant protection in several cell types following exposure to
different stimuli by activating the PI3K/Akt/mTOR complex 1 or MAPK
ERK/p38 signaling pathways (38–40). Quercetin, another polyphenol
antioxidant, activates the MAPK signaling pathway, thereby inducing
the expression of HO-1, which provides protection against apoptosis
and inflammation (41,42). Additionally, plant-derived
polyphenols have been reported to show antioxidative stress and
anti-inflammatory properties via AMPK-dependent induction of HO-1
(43,44). The precise mechanism by which the
protein kinase signaling pathway induces the expression of HO-1
involves the cross-talk of transcription factors, including Nrf2,
AP1 and HIF-1, and has been previously reviewed in detail (21,34). In general, the induced expression
of HO-1 is an adaptive strategy that enables cells to achieve
self-protection under stress conditions. However, a variety of
genetic epidemiological studies have shown that there are two
important genetic polymorphisms in the promoter region of the HO-1
gene (45–47). These polymorphisms determine the
expression level of HO-1 and affect several diseases, such as
cardiovascular, pulmonary, liver, renal, cancer and viral
diseases.
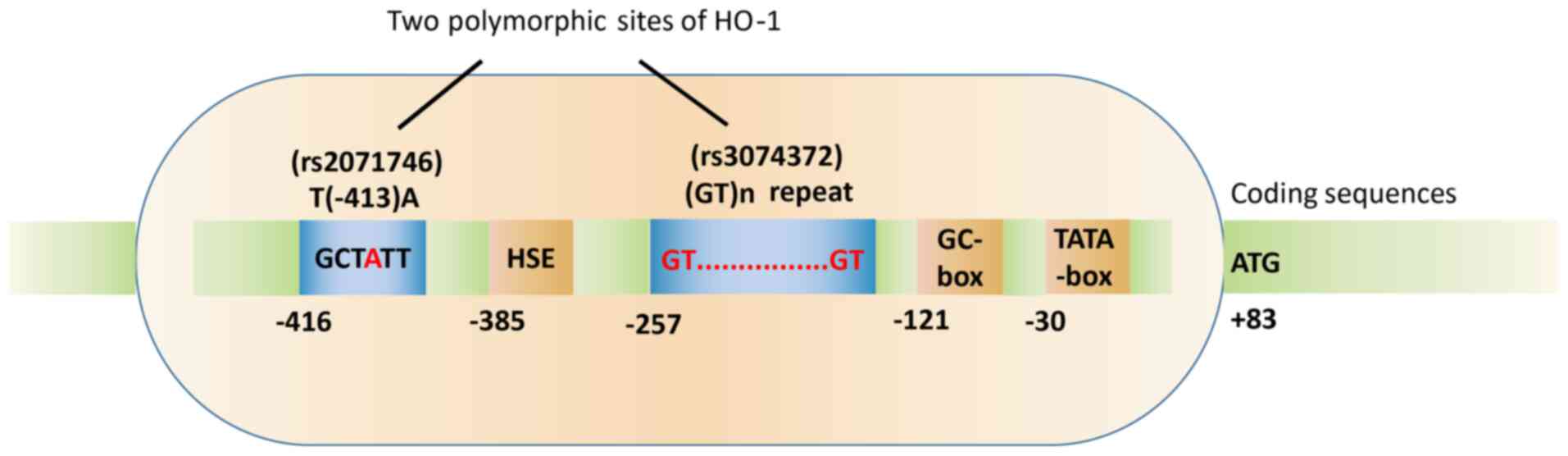
HO-1 promoter polymorphism
HO-1 gene polymorphisms are primarily based on two
sites: The (GT)n double nucleotide polymorphism (also called
microsatellite polymorphism) and the T(−413)A single nucleotide
polymorphism (SNP), both of which can regulate HO-1 gene
transcription under specific conditions (45). These two polymorphic sites on the
HO-1 gene promoter are shown in Fig.
1.
(GT)n double nucleotide
polymorphism
Of the two polymorphism sites found in the HO-1
promoter region, the (GT)n dinucleotide repeat polymorphism
(rs3074372) occurs more commonly. The number of (GT)n repeats
ranges from 12–45 repeats (48),
and the distribution of (GT)n repeats is bimodal, with alleles of
~23 and 30 repeat lengths (5,12,48–52). According to this genetic
distribution in humans and the effect of GTn repeats on the
transcriptional activity of HO-1, short (S) alleles have been
defined as having <25 GTn repeats, whereas those with ≥25
repeats were defined as long (L) alleles (49,53). Alternatively, other studies have
used a different classification method: S alleles have been denoted
by <27 GT, middle alleles (M) as 27–32 GT and L alleles as ≥33
GT (11,54). The biological function generated
through the use of either classification system should be
consistent, taking into account the (GT)n distribution frequency
(5).
Through the use of the luciferase reporter gene
system in vitro, previous studies have confirmed that S
alleles are associated with a higher transcriptional activity of
HO-1 compared with L alleles (51,52). Consistent with this, genetic
epidemiological studies have shown that subjects with S alleles
have higher levels of HO-1 mRNA compared with subjects with L
alleles (12,13). Moreover, when lymphocytes isolated
from humans were exposed to H2O2 stimulation
in vitro, the HO-1 expression level in cells with S alleles
was found to be higher compared with that of cells with L alleles.
Functionally, lymphocytes with S alleles exhibit a stronger
antiapoptotic ability (11). On
the other hand, L (GT)n repeats have been demonstrated to be
associated with a higher susceptibility to the development of
numerous diseases, such as hypertension, atherosclerosis, acute
kidney injury, breast cancer and pancreatitis. Clinical studies
have shown that the (GT)n repeat length of the HO-1 promoter is
correlated with the progression of a variety of diseases, such as
hypertension, atherosclerosis, and acute kidney injury (55–57).
T(−413)A SNP
T(−413)A SNP (rs2071746), another HO-1 promoter
polymorphism, was detected by reverse transcription-quantitative
PCR. Yamada et al (52)
transfected plasmids containing different HO-1 gene promoters into
bovine aortic endothelial cells to detect functional differences
caused by SNPs. The results obtained showed that cells containing
the A(−413)-(GT)30 allele were able to significantly
enhance the biological activity of HO-1. Several studies have
subsequently confirmed that the A allele is also associated with a
high transcription level of HO-1, which correlates closely with the
occurrence and progression of certain human diseases, including
cardiovascular, and liver diseases (58–60). A previous study reported that the
AA genotype of T(−413)A could significantly reduce the occurrence
of myocardial infarction and angina pectoris (61). In addition, the survival rate of
transplanted livers was shown to be significantly higher following
liver transplantation from donors who carried the A allele
(62).
Population characteristics of HO-1
gene promoter polymorphisms
According to a study of HO-1 gene promoter
polymorphism in Caucasian and East Asian populations, the
distribution of (GT)n alleles showed a bimodal pattern with
(GT)23 and (GT)30 being the two most common
alleles in the two groups (50,51). However, the distribution of (GT)n
alleles showed a trimodal pattern with peaks at (GT)23,
(GT)30 and (GT)39 in African-American
populations (50). Compared with
Caucasian populations, meta-analysis showed that East Asian
populations had more S alleles and fewer L alleles (58). (GT)n polymorphism includes three
genotypes, namely SS, SL and LL. East Asian populations showed
higher SS frequencies, ranging from 15 to 25%, whereas Caucasian
populations displayed SS frequencies ranging from 6 to 17%
(58). The SS genotype was
associated with a lower incidence of chronic diseases, such as
coronary heart disease and rheumatoid arthritis (12,58). The incidence of rheumatoid
arthritis in East Asian populations is lower than that in Caucasian
populations, which might be related to the higher SS genotype in
East Asians. In addition, a recent study showed that Caucasian
American populations have a higher SS allele frequency compared
with African American populations, which might be the cause of
higher neurocognitive impairment caused by human immunodeficiency
virus (HIV) infection in African American populations (50). Similarly, the T(−413)A SNP
polymorphism genotype also contains three genotypes, namely AA, AT
and TT. These genotypes also differs between Caucasian and East
Asian populations. Meta-analysis showed that East Asian populations
had a lower AA genotype (~22%) compared with Caucasian populations
(~35%). People with the AA genotype displayed a lower risk of
coronary heart disease (58).
Mechanism of promoter polymorphisms
regulating HO-1 expression
To date, the mechanism through which the T(−413)A
and (GT)n gene polymorphisms regulate the expression of HO-1 has
not been elucidated. A potential explanation may be that (GT)n
microsatellite repeats tend to form a Z-DNA conformation, such as a
left-handed double-helix structure (63). The Z-DNA conformation only occurs
rarely in the genome, although its incidence does increase under
certain conditions, such as the presence of a
purine/pyrimidine-alternating sequence, DNA supercoiling and a high
concentration of salt and certain cations (5,64).
During the transcription process, the local Z-DNA conformation
reduces the transcription activity of DNA. Therefore, the L (GT)n
microsatellite repeats in the promoter tend to form a Z-DNA
conformation, which consequently reduces the transcriptional
activity of HO-1. In addition, a previous study showed that the
T(−413)A and (GT)n polymorphisms may be involved in the splicing
and translation of the HO-1 transcript (65). An extra exon (Exon1a) was
identified for the first time in the 5′-untranslated region
(5′-UTR) of HO-1. Moreover, the T(−413)A and (GT)n loci are located
between Exon1a and Exon1. T(−413)A and (GT)n are located in the
primary transcripts of HO-1 as alternative intronic or exonic
elements. Rapid amplification of cDNA ends and luciferase reporter
assays were subsequently used to confirm that T(−413)A and (GT)n
are involved in 5′-UTR alternative splicing of the HO-1 primary
transcript, which may result in translational control of HO-1
through regulating mRNA stability and translational efficiency
(65). Moreover, expression of
Exon 1a induced by hemin in hepatoma HepG2 cells is shown to
decrease mature HO-1 mRNA isoform translation, which might explain
the previous finding that human HO-1 mRNA and protein levels fail
to correlate (66).
Association between HO-1 gene polymorphism
and clinical diseases
As a cytoprotective enzyme, the intracellular
expression level of HO-1 is influenced by the (GT)n dinucleotide
polymorphism and T(−413)A SNP in its promoter region. Several
studies have shown that these two polymorphisms affect the
occurrence and progression of a variety of diseases, including
liver and kidney, lung, cardiovascular, cancer and viral diseases
(48,52,55–58,67). The association between two HO-1
promoter polymorphisms and diseases are summarized in Table I.
 | Table I.Association between two HO-1 promoter
polymorphisms and a variety of diseases. |
Table I.
Association between two HO-1 promoter
polymorphisms and a variety of diseases.
| A, (GT)n
dinucleotide polymorphism |
|---|
|
|---|
| Disease | Polymorphism | P-value | (Refs.) |
|---|
| Cardiovascular
disease |
|
|
|
|
Coronary heart disease | S (GT)n repeats
reduce the risk of coronary heart disease or restenosis | P<0.05 | (58,68) |
|
Hypertension | Carriers with the L
(GT)n allele have an increased risk of hypertension, whereas the S
(GT)n allele may play a protective role | P<0.05 | (55,70) |
|
Atherosclerosis | Subjects with (GT)n
repeats ≥32 display an increased risk of cardiovascular disease and
enhanced atherosclerosis progression | P<0.01 | (56) |
| Pulmonary
disease |
|
|
|
| Chronic
obstructive pulmonary disease | L (GT)n allele is a
high risk factor for chronic obstructive pulmonary disease | P<0.01 | (52,78,79) |
| Liver disease |
|
|
|
| Liver
transplantation | S (GT)n sequence
(n<25) is beneficial to the functional improvement and survival
of allografts | P<0.05 | (82) |
| Renal disease |
|
|
|
| Acute
kidney injury | Carrying the L
(GT)n allele increases the risk of acute kidney injury after heart
surgery, whereas carrying S (GT)n allele increases the risk of
acute kidney injury in patients with sepsis | P<0.05 | (57,87) |
| Cancer |
|
|
|
|
Pancreatic cancer,
melanoma | The S (GT)n allele
sequence is associated with an increased risk and lesion of
pancreatic cancer and melanoma | P<0.001
(pancreatic cancer) or P<0.05 (melanoma) | (88,89) |
| Acute
lymphoblastic leukemia | Patients with S
(GT)n repeats display chemotherapy resistance and chemo-induced
neutropenia | P<0.05 | (92) |
| Breast
cancer, esophageal squamous cell carcinoma and laryngeal squamous
cell carcinoma | L (GT)n repeats are
associated with a higher risk of these type of cancer | P<0.05 | (67,93–95) |
| Gastric
cancer and lung adenocarcinoma | The effect of HO-1
promoter polymorphism on cancer may depend on a variety of factors,
including tumor type, subject ethnicity and other factors | N/A | (96–98) |
| Viral disease |
|
|
|
|
HIV | S (GT)n allele
inhibits neuroimmune activation, HIV-associated neuroinflammation
and neurocognitive deficits | P<0.05 | (10,48,50) |
|
HCV | S (GT)n allele
decreases the infection titer of HCV | P>0.05 | (101) |
| Other disease |
|
|
|
|
Neonatal hyperbilirubin | S (GT)n repeat gene
promoter results in a higher risk of neonatal
hyperbilirubinemia | P<0.05 | (103) |
|
Rheumatoid arthritis | S (GT)n allele
reduces the risk of rheumatoid arthritis and prevents the
development of joint injury | P<0.05 | (12,104) |
|
Pancreatitis | HO-1 containing LL
allele doubles the risk of pancreatic necrosis in patients with
acute pancreatitis | P<0.05 | (105) |
|
| B, T(−413)A
single nucleotide polymorphism |
|
| Disease |
Polymorphism | P-value | (Refs.) |
|
| Cardiovascular
disease |
|
|
|
|
Coronary heart disease | The A allele
reduces the risk of coronary heart disease | P<0.05 | (58) |
|
Hypertension | AA genotype
increases the incidence of hypertension in a female population | P<0.01 | (59) |
| Liver disease |
|
|
|
| Liver
transplantation | Patients carrying
the T allele are more likely to develop esophageal varices compared
with those carrying the A allele | P<0.001 | (60) |
| Other disease |
|
|
|
| Type II
diabetes | Patients with the
TT genotype are more prone to developing proteinuria | P<0.05 | (106) |
|
Ischemic heart disease | The AA genotype of
HO-1 reduces the incidence of ischemic heart disease | P<0.05 | (61) |
Cardiovascular disease
HO-1 is a protective factor for cardiovascular
diseases (68). Previous studies
have shown that the level of serum bilirubin, a downstream product
of HO-1, is negatively correlated with the occurrence of coronary
heart disease (CHD), and lower levels of serum bilirubin increase
the risk of CHD. Through a correlation analysis, Zhang et al
(58) demonstrated that the HO-1
(GT)n repeat length polymorphism and T(−413)A SNP were both
associated with a risk of CHD following percutaneous coronary
intervention. Patients with the S allele of HO-1 (GT)n had a
reduced risk of restenosis compared with those with the L allele of
HO-1 (GT)n. Similarly, the HO-1 A allele of T(−413)A was associated
with a reduced risk of CHD. Consistent with the aforementioned
results, Liang et al (69)
showed that the HO-1 promoter (GT)n repeat sequence in patients
with CHD with a reduced ejection fraction was longer compared with
that in patients with CHD with moderate ejection fraction.
Ono et al (59) demonstrated that the HO-1 AA
genotype is associated with an increased incidence of hypertension
in a female population. Wenzel et al (55) reported that the individuals
carrying the HO-1 L allele had an increased risk for the prevalence
of arterial hypertension and higher overall mortality. In line with
this, another study demonstrated that the HO-1 S allele may serve a
protective role against environmental stressors in blood pressure
regulation and cardiovascular mortality risk in hypertensive
individuals (70). Therefore, the
(GT)n repeat or SNP polymorphisms may be used as molecular markers
for screening individuals who are at risk of hypertension.
Atherosclerosis is an inflammatory disease (71). As an anti-inflammatory protein,
HO-1 has been confirmed to have a protective effect on blood
vessels and is able to inhibit the formation of atherosclerotic
lesions (72–74) and thrombosis (75). Liang et al (76) also found that the HO-1 promoter
(GT)n repeat sequence was longer in patients with microvascular
angina compared with that in healthy controls. Similarly, Pechlaner
et al (56) demonstrated
that there is a strong correlation between the HO-1(GT)n repeat
polymorphism and cardiovascular disease risk in subjects for whom
the number of (GT)n repeats was ≥32. Taken together, these studies
provided some indicators for how atherosclerosis may be accelerated
and antioxidant defenses decreased in high-risk vascular
groups.
In conclusion, individuals carrying the L allele of
HO-1 (GT)n were shown to have an increased risk of CHD,
hypertension and atherosclerosis, whereas the A allele of T(−413)A
in the HO-1 gene promoter is associated with a lower risk of CHD,
but an increased risk of hypertension. However, these controversial
conclusions might have resulted from the limited number of subjects
and different populations included in the CHD studies, thus these
limitations also need be taken into consideration.
Pulmonary disease
The induction of HO-1 expression is considered to be
an important protective mechanism involved in acute and chronic
lung diseases, and HO-1 is an important enzymes that exerts
antioxidant protective effects in the lung (77). HO-1 gene polymorphism is closely
associated with chronic obstructive pulmonary disease (COPD)
(52,78,79).
A previous study confirmed that insufficient or
inhibited levels of HO-1 expression are an important internal
factor for inducing the occurrence of COPD (80). Zhang et al (81) found that HO-1 exerts a protective
function on the lungs, and the number of GT repeats was shown to
influence the effects of oral N-acetylcysteine treatment. Patients
with L (GT)n alleles were shown to have a much higher rate of
deterioration compared with patients that lacked the L alleles.
From this study, it may be concluded that HO-1 gene polymorphism
has an important role in COPD. Yamada et al (52), Du et al (78) and Zhou et al (79) also demonstrated that the L (GT)n
allele frequency in patients with COPD was significantly higher
compared with that in the control group.
Liver disease
HO-1 has also been shown to protect allografts
against ischemia/reperfusion and immune rejection. By studying 60
patients with allogeneic orthotopic liver transplantation and their
accompanying donors, Zhang et al (82) found that donor-derived HO-1 gene
promoter polymorphism exerted a protective effect on allograft
function following orthotopic transplantation. A short (GT)n
sequence (n<25 repeats) leads to increased HO-1 transcriptional
activity, and a series of transplantation models have demonstrated
that HO-1 upregulation is beneficial to the functional improvement
and survival of the allograft (82–85). In addition, studies have shown
that patients with liver cirrhosis who carry the HO-1 T allele are
more likely to develop esophageal varices than those with liver
cirrhosis who carry the A allele. Therefore, the T allele of the
HO-1 gene SNP is a risk factor for esophageal varices in patients
with cirrhosis (60).
Renal disease
Acute kidney injury (AKI) refers to a multifactorial
clinical syndrome caused by a variety of diseases, such as
hypovolemia, diabetes, heart or lung disease (86). Leaf et al (57) demonstrated that repeated HO-1
polymorphism is associated with the development of AKI after
cardiac surgery in adults, and carrying the HO-1 L (GT)n sequence
(n>27 repeats) increases the risk of AKI after cardiac surgery.
Hemolysis may be the primary cause of AKI, but the factors
associated with cardiac surgery are complex and varied. The most
plausible mechanism that has been proposed to date is that the
expression of HO-1 is reduced in patients with L (GT)n repeats,
resulting in a decrease in the decomposition of heme. Therefore,
patients with reduced HO-1 expression caused by L (GT)n repeats may
have increased susceptibility to heme-related AKI during cardiac
surgery.
In a different study, Vilander et al
(87) classified the HO-1 (GT)n
repeat sequence in 300 patients with severe AKI sepsis and 353
patients without AKI sepsis. The alleles were classified according
to the number of replicates. The short sequence S was defined as
<27 repeats, the long sequence L was defined as ≥27 repeats and
the very long sequence (L2) was defined as ≥34 repeats. Patients
with at least one L2 sequence had lower plasma HO-1 concentrations
compared with those without the L2 sequence. Patients with the S
allele were found to be at an increased risk for AKI, and the
adjusted odds ratio for each S allele was 1.3 in the additive
genetic model. The conclusions of the aforementioned two studies
are different as the susceptibility factors of the study
populations were different. In the study conducted by Leaf et
al, patients underwent cardiopulmonary bypass surgery, whereas
all patients were critically ill with sepsis in the study conducted
by Vilander et al. In addition, urination criteria were not
included in the study by Leaf et al, but patients with mild
AKI were excluded in the study by Vilander et al. In
conclusion, it is necessary to distinguish the different phenotypes
of AKI clinical syndromes when conducting pathophysiological
studies on patients with AKI.
Cancer
Increasing evidence on the association between HO-1
gene promoter polymorphism and cancer has been reported. Previous
studies have shown that S (GT)n repeats in the HO-1 promoter lead
to higher transcriptional activity, which leads to an increased
risk of pancreatic cancer and melanoma (88,89). In pancreatic cancer studies,
patients with HO-1 S (GT)n repeats developed cancer more rapidly
and were more likely to experience recurrence. This finding was
consistent with the fact that HO-1 expression in pancreatic tumors
was higher compared with that in healthy tissues, and that HO-1
overexpression has been shown to accelerate the growth,
angiogenesis and metastasis of pancreatic tumors (90,91). In pediatric patients with acute
lymphoblastic leukemia, patients with HO-1 S (GT)n repeats were
associated with chemotherapy resistance and chemo-induced
neutropenia (92). In other
studies, L (GT)n repeats in the promoter of the HO-1 gene have been
reported to be associated with a higher risk of breast cancer,
esophageal squamous cell carcinoma and laryngeal squamous cell
carcinoma (67,93–95).
However, the effects of HO-1 promoter polymorphism
in certain types of cancer have given rise to inconsistent results.
For example, Lo et al (96) studied 183 patients with gastric
cancer and 250 control patients in the Taiwan area. The results
demonstrated that the L (GT)n repeat of the HO-1 gene promoter was
associated with a high frequency of gastric cancer, and that the M
(GT)n repeat may have a protective effect on gastric adenocarcinoma
with a low frequency of lymphatic vascular invasion. However, Sawa
et al (97) found that, in
the female Japanese population, HO-1 S (GT)n repeats were
associated with a risk of gastric cancer. This difference in the
reported results may have been due to environmental factors and
living habits. In addition, for lung adenocarcinoma, male smokers
with the L allele had a significantly increased risk of lung
adenocarcinoma compared with those with the non-L allele. In female
non-smokers, L allele carriers did not differ between patients and
controls (98). For sporadic
colorectal cancer, studies have shown no association between HO-1
promoter (GT)n repeat polymorphism/T(−413)A SNP and the risk of
disease (99). Taken together,
these findings suggested that the effect of HO-1 gene promoter
polymorphism on cancer may depend on a variety of factors,
including tumor type and subject ethnicity (100).
Viral diseases
Studies have shown that gene polymorphisms in the
HO-1 promoter region are associated with the occurrence and
progression of viral infectious diseases. Gill et al
(48) demonstrated that
individual HO-1 (GT)n allele repeat length polymorphisms have a
unique modifying effect on the risk of HIV-induced central nervous
system neuroinflammation and associated neuropathy. The HO-1 S
(GT)n allele increases HO-1 promoter activity and exerts a
neuroprotective function through inhibiting neuroimmune activation.
Similarly, previous studies have shown that the HO-1 S (GT)n allele
sequence in African American populations reduces HIV-associated
neuroinflammation and limits HIV-induced neurocognitive deficits
(10,50). Therefore, therapeutic strategies
that induce HO-1 expression should be able to reduce HIV-associated
central nervous system inflammation and reduce the risk of HIV
neurological disease development.
Kah et al (101) showed that inoculating mice with
human hepatocytes containing the HO-1 S (GT)n allele led to a
decrease in the infection titer of hepatitis C virus (HCV) compared
with mice inoculated with the HO-1 L (GT)n allele. Therefore,
HO-1-mediated HCV inhibition is (GT)n polymorphism-dependent. In
addition, it has been suggested that HO-1 promoter polymorphism may
influence coronavirus COVID-19 disease severity, such as acute lung
injury and thromboembolism. Therefore, the repeat length of the
HO-1 (GT)n allele in patients with severe COVID-19 should be
investigated (102).
Other diseases
HO-1 gene promoter polymorphism has also been
reported to be associated with neonatal hyperbilirubinemia
(103), rheumatoid arthritis
(12,104), pancreatitis (105), type II diabetes mellitus
(106), heart disease (107) and other diseases to varying
degrees. HO-1 carrying an S (GT)n repeat gene promoter was found to
be associated with a higher risk of neonatal hyperbilirubinemia
(103). By contrast, carrying
the S (GT)n microsatellite allele was found to reduce the risk of
rheumatoid arthritis and prevent the development of joint injury,
whereas carrying the L (GT)n microsatellite allele led to an
increase in the susceptibility to rheumatoid arthritis and promoted
the development of joint injury (12,104). For pancreatitis, the results of
logistic regression analysis revealed that HO-1 containing the L
(GT)n allele doubled the risk of pancreatic necrosis in patients
with acute pancreatitis (105).
However, in patients undergoing heart transplantation, (GT)n
polymorphism was not found to be associated with heart failure or
patient survival. The researchers assumed that lower HO-1 activity
in patients with L (GT)n alleles might be compensated by other
protective genes or enzymes (107). In addition, in a retrospective
analysis of patients with type II diabetes, HO-1 T(−413)A SNP was
significantly associated with the prevalence of low proteinuria,
and patients with the TT genotype were more prone to develop
proteinuria. Furthermore, in vitro experiments revealed that
the promoter activity of HO-1 containing the A allele was higher
than that of T allele (106).
The AA genotype of HO-1 for T(−413)A SNP polymorphism was also
found to reduce the incidence of ischemic heart disease, which
might be associated with the high expression level of HO-1
(61).
Conclusions and prospects
As a stress-induced cytoprotective enzyme, HO-1
expression is regulated by a complex network, including
transcription factors, cytoplasmic signaling pathways,
post-translational modifications and genetic polymorphisms. The
research of HO-1 induced expression primarily focuses on
transcription factors. To treat a variety of diseases related to
inflammation and immune regulation, screening small molecule HO-1
inducers that target transcription factors is the most commonly
used method (108,109). However, this attempt at
translational research was not successful and encountered great
challenges. In addition to toxicity and bioavailability issues,
HO-1 gene polymorphism in the population may also be an important
reason for the difficulty of this translational study. Therefore,
if the HO-1 gene polymorphism is added as a biomarker in clinical
trials, it may improve the success rate of translational research.
The current challenge is that the clinical results of the
correlation between HO-1 gene polymorphism and disease are still
controversial, especially due to the small sample sizes and
inconsistent definition standards. On the other hand, the molecular
mechanism underlying HO-1 polymorphism-mediated effects on HO-1
gene expression under disease conditions has not been fully
revealed, which limits our understanding of the correlation between
HO-1 gene polymorphisms and different diseases. Through the method
of in vitro transient transfection, the relationship between
(GT)n repeats length and HO-1 expression level has been established
at the cellular level; however, the research of (GT)n repeat length
effects on HO-1 expression at the animal level has not yet been
reported. With the emergence of CRISPR technology, it is possible
to construct HO-1 gene polymorphism transgenic mice. Future
research should focus on exploring the molecular mechanisms by
which HO-1 gene polymorphism and transcription factors coregulate
HO-1 expression and affect disease using transgenic mouse systems.
These investigations will provide further support for the
development of targeted HO-1 reagents in different diseases.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 81902052).
Availability of data and materials
Not applicable.
Authors' contributions
LLM and LS wrote and prepared the final version of
the manuscript. YXW and BHS contributed to revising the manuscript.
YFL and YLJ conceptualized the idea of the review. All authors read
and approved the final manuscript. Data authentication is not
applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HO-1
|
heme oxygenase-1
|
|
CO
|
carbon monoxide
|
|
CHD
|
coronary heart disease
|
|
S
|
short
|
|
L
|
long
|
|
COPD
|
chronic obstructive pulmonary
disease
|
|
AKI
|
acute kidney injury
|
References
|
1
|
Gozzelino R, Jeney V and Soares MP:
Mechanisms of cell protection by heme oxygenase-1. Annu Rev
Pharmacol Toxicol. 50:323–354. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wagener FA, Volk HD, Willis D, Abraham NG,
Soares MP, Adema GJ and Figdor CG: Different faces of the heme-heme
oxygenase system in inflammation. Pharmacol Rev. 55:551–571. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim HP, Ryter SW and Choi AM: CO as a
cellular signaling molecule. Annu Rev Pharmacol Toxicol.
46:411–449. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Calabrese V, Butterfield DA, Scapagnini G,
Stella AG and Maines MD: Redox regulation of heat shock protein
expression by signaling involving nitric oxide and carbon monoxide:
Relevance to brain aging, neurodegenerative disorders, and
longevity. Antioxid Redox Signal. 8:444–477. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Exner M, Minar E, Wagner O and Schillinger
M: The role of heme oxygenase-1 promoter polymorphisms in human
disease. Free Radic Biol Med. 37:1097–1104. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Linnenbaum M, Busker M, Kraehling JR and
Behrends S: Heme oxygenase isoforms differ in their subcellular
trafficking during hypoxia and are differentially modulated by
cytochrome P450 reductase. PLoS One. 7:e354832012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shibahara S: The heme oxygenase dilemma in
cellular homeostasis: New insights for the feedback regulation of
heme catabolism. Tohoku J Exp Med. 200:167–186. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhou J, Terluk MR, Basso L, Mishra UR,
Orchard PJ, Cloyd JC, Schröder H and Kartha RV: N-acetylcysteine
provides cytoprotection in murine oligodendrocytes through heme
oxygenase-1 activity. Biomedicines. 8:2402020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jazwa A and Cuadrado A: Targeting heme
oxygenase-1 for neuroprotection and neuroinflammation in
neurodegenerative diseases. Curr Drug Targets. 11:1517–1531. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Seu L, Burt TD, Witte JS, Martin JN, Deeks
SG and McCune JM: Variations in the heme oxygenase-1 microsatellite
polymorphism are associated with plasma CD14 and viral load in
HIV-infected African-Americans. Genes Immun. 13:258–267. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hirai H, Kubo H, Yamaya M, Nakayama K,
Numasaki M, Kobayashi S, Suzuki S, Shibahara S and Sasaki H:
Microsatellite polymorphism in heme oxygenase-1 gene promoter is
associated with susceptibility to oxidant-induced apoptosis in
lymphoblastoid cell lines. Blood. 102:1619–1621. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rueda B, Oliver J, Robledo G, López-Nevot
MA, Balsa A, Pascual-Salcedo D, González-Gay MA, González-Escribano
MF and Martín J: HO-1 promoter polymorphism associated with
rheumatoid arthritis. Arthritis Rheum. 56:3953–3958. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brydun A, Watari Y, Yamamoto Y, Okuhara K,
Teragawa H, Kono F, Chayama K, Oshima T and Ozono R: Reduced
expression of heme oxygenase-1 in patients with coronary
atherosclerosis. Hypertens Res. 30:341–348. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Loboda A, Damulewicz M, Pyza E, Jozkowicz
A and Dulak J: Role of Nrf2/HO-1 system in development, oxidative
stress response and diseases: An evolutionarily conserved
mechanism. Cell Mol Life Sci. 73:3221–3247. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang DD: Mechanistic studies of the
Nrf2-Keap1 signaling pathway. Drug Metab Rev. 38:769–789. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jaramillo MC and Zhang DD: The emerging
role of the Nrf2-Keap1 signaling pathway in cancer. Genes Dev.
27:2179–2191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Motohashi H, Katsuoka F, Engel JD and
Yamamoto M: Small Maf proteins serve as transcriptional cofactors
for keratinocyte differentiation in the Keap1-Nrf2 regulatory
pathway. Proc Natl Acad Sci USA. 101:6379–6384. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Itoh K, Chiba T, Takahashi S, Ishii T,
Igarashi K, Katoh Y, Oyake T, Hayashi N, Satoh K, Hatayama I, et
al: An Nrf2/small Maf heterodimer mediates the induction of phase
II detoxifying enzyme genes through antioxidant response elements.
Biochem Biophys Res Commun. 236:313–322. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dhakshinamoorthy S, Jain AK, Bloom DA and
Jaiswal AK: Bach1 competes with Nrf2 leading to negative regulation
of the antioxidant response element (ARE)-mediated NAD(P)H: Quinone
oxidoreductase 1 gene expression and induction in response to
antioxidants. J Biol Chem. 280:16891–16900. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Reichard JF, Motz GT and Puga A: Heme
oxygenase-1 induction by NRF2 requires inactivation of the
transcriptional repressor BACH1. Nucleic Acids Res. 35:7074–7086.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Medina MV, Sapochnik D, Garcia Solá M and
Coso O: Regulation of the expression of heme oxygenase-1: Signal
transduction, gene promoter activation, and beyond. Antioxid Redox
Signal. 32:1033–1044. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Alam J and Cook JL: How many transcription
factors does it take to turn on the heme oxygenase-1 gene? Am J
Respir Cell Mol Biol. 36:166–174. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dawn B and Bolli R: HO-1 induction by
HIF-1: A new mechanism for delayed cardioprotection? Am J Physiol
Heart Circ Physiol. 289:H522–H524. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li X, Yu J, Gong L, Zhang Y, Dong S, Shi
J, Li C, Li Y, Zhang Y and Li H: Heme oxygenase-1(HO-1) regulates
Golgi stress and attenuates endotoxin-induced acute lung injury
through hypoxia inducible factor-1α (HIF-1α)/HO-1 signaling
pathway. Free Radic Biol Med. 165:243–253. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tong Y, Kai J, Wang S, Yu Y, Xie S, Zheng
H, Wang Y, Liu Y, Zhu K, Guan X, et al: VHL regulates the
sensitivity of clear cell renal cell carcinoma to SIRT4-mediated
metabolic stress via HIF-1α/HO-1 pathway. Cell Death Dis.
12:6212021. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He C, Zhang W, Li S, Ruan W, Xu J and Xiao
F: Edaravone improves septic cardiac function by inducing an
HIF-1α/HO-1 pathway. Oxid Med Cell Longev. 2018:52163832018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee PJ, Jiang B-H, Chin BY, Iyer NV, Alam
J, Semenza GL and Choi AK: Hypoxia-inducible factor-1 mediates
transcriptional activation of the heme oxygenase-1 gene in response
to hypoxia. J Biol Chem. 272:5375–5381. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tanos T, Marinissen MJ, Leskow FC,
Hochbaum D, Martinetto H, Gutkind JS and Coso OA: Phosphorylation
of c-Fos by members of the p38 MAPK family. Role in the AP-1
response to UV light. J Biol Chem. 280:18842–18852. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Monje P, Marinissen MJ and Gutkind JS:
Phosphorylation of the carboxyl-terminal transactivation domain of
c-Fos by extracellular signal-regulated kinase mediates the
transcriptional activation of AP-1 and cellular transformation
induced by platelet-derived growth factor. Mol Cell Biol.
23:7030–7043. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
McBride K and Nemer M: The C-terminal
domain of c-fos is required for activation of an AP-1 site specific
for jun-fos heterodimers. Mol Cell Biol. 18:5073–5081. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yeh CH, Chen TP, Wang YC, Lin YM and Lin
PJ: HO-1 activation can attenuate cardiomyocytic apoptosis via
inhibition of NF-kappaB and AP-1 translocation following cardiac
global ischemia and reperfusion. J Surg Res. 155:147–156. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee PJ, Camhi SL, Chin BY, Alam J and Choi
AM: AP-1 and STAT mediate hyperoxia-induced gene transcription of
heme oxygenase-1. Am J Physiol Lung Cell Mol Physiol.
279:L175–L182. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Paine A, Eiz-Vesper B, Blasczyk R and
Immenschuh S: Signaling to heme oxygenase-1 and its
anti-inflammatory therapeutic potential. Biochem Pharmacol.
80:1895–1903. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Campbell NK, Fitzgerald HK and Dunne A:
Regulation of inflammation by the antioxidant haem oxygenase 1. Nat
Rev Immunol. 21:411–425. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Merecz-Sadowska A, Sitarek P, Kucharska E,
Kowalczyk T, Zajdel K, Cegliński T and Zajdel R: Antioxidant
properties of plant-derived phenolic compounds and their effect on
skin fibroblast cells. Antioxidants (Basel). 10:7262021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yarmohammadi F, Hayes AW and Karimi G:
Natural compounds against cytotoxic drug-induced cardiotoxicity: A
review on the involvement of PI3K/Akt signaling pathway. J Biochem
Mol Toxicol. 35:e226832021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhou Y, Jiang Z, Lu H, Xu Z, Tong R, Shi J
and Jia G: Recent advances of natural polyphenols activators for
Keap1-Nrf2 signaling pathway. Chem Biodivers. 16:e19004002019.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang H, Liu YY, Jiang Q, Li KR, Zhao YX,
Cao C and Yao J: Salvianolic acid A protects RPE cells against
oxidative stress through activation of Nrf2/HO-1 signaling. Free
Radic Biol Med. 69:219–228. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang HF, Wang JH, Wang YL, Gao C, Gu YT,
Huang J, Wang JH and Zhang Z: Salvianolic acid A protects the
kidney against oxidative stress by activating the Akt/GSK-3β/Nrf2
signaling pathway and inhibiting the NF-κB signaling pathway in 5/6
nephrectomized rats. Oxid Med Cell Longev.
2019:28535342019.PubMed/NCBI
|
|
40
|
Gu X, Zheng C, Zheng Q, Chen S, Li W,
Shang Z and Zhang H: Salvianolic acid A attenuates early brain
injury after subarachnoid hemorrhage in rats by regulating
ERK/P38/Nrf2 signaling. Am J Transl Res. 9:5643–5652.
2017.PubMed/NCBI
|
|
41
|
Sun GY, Chen Z, Jasmer KJ, Chuang DY, Gu
Z, Hannink M and Simonyi A: Quercetin attenuates inflammatory
responses in BV-2 microglial cells: Role of MAPKs on the Nrf2
pathway and induction of heme oxygenase-1. PLoS One.
10:e01415092015. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yao P, Nussler A, Liu L, Hao L, Song F,
Schirmeier A and Nussler N: Quercetin protects human hepatocytes
from ethanol-derived oxidative stress by inducing heme oxygenase-1
via the MAPK/Nrf2 pathways. J Hepatol. 47:253–261. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Campbell NK, Fitzgerald HK, Fletcher JM
and Dunne A: Plant-derived polyphenols modulate human dendritic
cell metabolism and immune function via AMPK-dependent induction of
heme oxygenase-1. Front Immunol. 10:3452019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kosuru R, Kandula V, Rai U, Prakash S, Xia
Z and Singh S: Pterostilbene decreases cardiac oxidative stress and
inflammation via activation of AMPK/Nrf2/HO-1 pathway in
fructose-fed diabetic rats. Cardiovasc Drugs Ther. 32:147–163.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nakasone R, Ashina M, Abe S, Tanimura K,
Van Rostenberghe H and Fujioka K: The role of heme oxygenase-1
promoter polymorphisms in perinatal disease. Int J Environ Res
Public Health. 18:35202021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Öllinger R and Pratschke J: Role of heme
oxygenase-1 in transplantation. Transpl Int. 23:1071–1081. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Schulz S, Chisholm KM, Zhao H, Kalish F,
Yang Y, Wong RJ and Stevenson DK: Heme oxygenase-1 confers
protection and alters T-cell populations in a mouse model of
neonatal intestinal inflammation. Pediatr Res. 77:640–648. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gill AJ, Garza R, Ambegaokar SS, Gelman BB
and Kolson DL: Heme oxygenase-1 promoter region (GT)n polymorphism
associates with increased neuroimmune activation and risk for
encephalitis in HIV infection. J Neuroinflammation. 15:702018.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kaplan M, Wong RJ and Stevenson DK: Heme
oxygenase-1 promoter polymorphisms: Do they modulate neonatal
hyperbilirubinemia? J Perinatol. 37:901–905. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Garza R, Gill AJ, Bastien BL, Garcia-Mesa
Y, Gruenewald AL, Gelman BB, Tsima B, Gross R, Letendre SL and
Kolson DL: Heme oxygenase-1 promoter (GT)n polymorphism associates
with HIV neurocognitive impairment. Neurol Neuroimmunol
Neuroinflamm. 7:e7102020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chen YH, Lin SJ, Lin MW, Tsai HL, Kuo SS,
Chen JW, Charng MJ, Wu TC, Chen LC, Ding YA, et al: Microsatellite
polymorphism in promoter of heme oxygenase-1 gene is associated
with susceptibility to coronary artery disease in type 2 diabetic
patients. Hum Genet. 111:1–8. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yamada N, Yamaya M, Okinaga S, Nakayama K,
Sekizawa K, Shibahara S and Sasaki H: Microsatellite polymorphism
in the heme oxygenase-1 gene promoter is associated with
susceptibility to emphysema. Am J Hum Genet. 66:187–195. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Espinoza JA, González PA and Kalergis AM:
Modulation of antiviral immunity by heme oxygenase-1. Am J Pathol.
187:487–493. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhou Y, Wang SN, Li H, Zha W, Peng Q, Li
S, Chen Y and Jin L: Quantitative trait analysis of polymorphisms
in two bilirubin metabolism enzymes to physiologic bilirubin levels
in Chinese newborns. J Pediatr. 165:1154–1160.e1. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wenzel P, Rossmann H, Müller C, Kossmann
S, Oelze M, Schulz A, Arnold N, Simsek C, Lagrange J, Klemz R, et
al: Heme oxygenase-1 suppresses a pro-inflammatory phenotype in
monocytes and determines endothelial function and arterial
hypertension in mice and humans. Eur Heart J. 36:3437–3446. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pechlaner R, Willeit P, Summerer M, Santer
P, Egger G, Kronenberg F, Demetz E, Weiss G, Tsimikas S, Witztum
JL, et al: Heme oxygenase-1 gene promoter microsatellite
polymorphism is associated with progressive atherosclerosis and
incident cardiovascular disease. Arterioscler Thromb Vasc Biol.
35:229–236. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Leaf DE, Body SC, Muehlschlegel JD,
McMahon GM, Lichtner P, Collard CD, Shernan SK, Fox AA and Waikar
SS: Length polymorphisms in heme oxygenase-1 and AKI after cardiac
surgery. J Am Soc Nephrol. 27:3291–3297. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang MM, Zheng YY, Gao Y, Zhang JZ, Liu
F, Yang YN, Li XM, Ma YT and Xie X: Heme oxygenase-1 gene promoter
polymorphisms are associated with coronary heart disease and
restenosis after percutaneous coronary intervention: A
meta-analysis. Oncotarget. 7:83437–83450. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ono K, Mannami T and Iwai N: Association
of a promoter variant of the haeme oxygenase-1 gene with
hypertension in women. J Hypertens. 21:1497–1503. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Ellakany WI, Mahmoud MoheyEldin K,
Invernizzi P, Mahmoud ElKady A, Eldin Fathy Abou Elkheir H, Abdel
Haleem Abo Elwafa R and Ellakany A: Study of the influence of heme
oxygenase 1 gene single nucleotide polymorphism (rs2071746) on
esophageal varices among patients with cirrhosis. Eur J
Gastroenterol Hepatol. 30:888–892. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ono K, Goto Y, Takagi S, Baba S, Tago N,
Nonogi H and Iwai N: A promoter variant of the heme oxygenase-1
gene may reduce the incidence of ischemic heart disease in
Japanese. Atherosclerosis. 173:313–319. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Buis CI, van der Steege G, Visser DS,
Nolte IM, Hepkema BG, Nijsten M, Slooff MJ and Porte RJ: Heme
oxygenase-1 genotype of the donor is associated with graft survival
after liver transplantation. Am J Transplant. 8:377–385. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Rich A, Nordheim A and Wang AH: The
chemistry and biology of left-handed Z-DNA. Annu Rev Biochem.
53:791–846. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Naylor LH and Clark EM: d(TG)n.d(CA)n
sequences upstream of the rat prolactin gene form Z-DNA and inhibit
gene transcription. Nucleic Acids Res. 18:1595–1601. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kramer M, Sponholz C, Slaba M, Wissuwa B,
Claus RA, Menzel U, Huse K, Platzer M and Bauer M: Alternative 5′
untranslated regions are involved in expression regulation of human
heme oxygenase-1. PLoS One. 8:e772242013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Doberer D, Haschemi A, Andreas M, Zapf TC,
Clive B, Jeitler M, Heinzl H, Wagner O, Wolzt M and Bilban M: Haem
arginate infusion stimulates haem oxygenase-1 expression in healthy
subjects. Br J Pharmacol. 161:1751–1762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hong CC, Ambrosone CB, Ahn J, Choi JY,
McCullough ML, Stevens VL, Rodriguez C, Thun MJ and Calle EE:
Genetic variability in iron-related oxidative stress pathways
(Nrf2, NQ01, NOS3, and HO-1), iron intake, and risk of
postmenopausal breast cancer. Cancer Epidemiol Biomarkers Prev.
16:1784–1794. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Abraham NG and Kappas A: Pharmacological
and clinical aspects of heme oxygenase. Pharmacol Rev. 60:79–127.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liang KW, Lee WJ, Lee WL, Wu JP, Lee IT,
Wang JS and Sheu WH: Subjects with coronary artery disease and
reduced ejection fraction have longer (GT)n repeats in the
heme-oxygenase 1 gene promoter. Heart Vessels. 36:615–620. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wu MM, Chiou HY, Chen CL, Hsu LI, Lien LM,
Wang CH, Hsieh YC, Wang YH, Hsueh YM, Lee TC, et al: Association of
heme oxygenase-1 GT-repeat polymorphism with blood pressure
phenotypes and its relevance to future cardiovascular mortality
risk: An observation based on arsenic-exposed individuals.
Atherosclerosis. 219:704–708. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Ross R: Atherosclerosis-an inflammatory
disease. N Engl J Med. 340:115–126. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Duckers HJ, Boehm M, True AL, Yet SF, San
H, Park JL, Clinton Webb R, Lee ME, Nabel GJ and Nabel EG: Heme
oxygenase-1 protects against vascular constriction and
proliferation. Nat Med. 7:693–698. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ishikawa K, Sugawara D, Wang XP, Suzuki K,
Itabe H, Maruyama Y and Lusis AJ: Heme oxygenase-1 inhibits
atherosclerotic lesion formation in ldl-receptor knockout mice.
Circ Res. 88:506–512. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Tulis DA, Durante W, Peyton KJ, Evans AJ
and Schafer AI: Heme oxygenase-1 attenuates vascular remodeling
following balloon injury in rat carotid arteries. Atherosclerosis.
155:113–122. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Lindenblatt N, Bordel R, Schareck W,
Menger MD and Vollmar B: Vascular heme oxygenase-1 induction
suppresses microvascular thrombus formation in vivo. Arterioscler
Thromb Vasc Biol. 24:601–606. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Liang KW, Lee WJ, Lee IT, Lee WL, Wang JS,
Wu JP and Sheu WHH: Subjects with microvascular angina have longer
GT repeats polymorphism in the haem oxygenase-1 gene promoter.
Biomarkers. 25:144–148. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Raval CM and Lee PJ: Heme oxygenase-1 in
lung disease. Curr Drug Targets. 11:1532–1540. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Du Y, Zhang H, Xu Y, Ding Y, Chen X, Mei
Z, Ding H and Jie Z: Association among genetic polymorphisms of
GSTP1, HO-1, and SOD-3 and chronic obstructive pulmonary disease
susceptibility. Int J Chron Obstruct Pulmon Dis. 14:2081–2088.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Zhou H, Ying X, Liu Y, Ye S, Yan J and Li
Y: Genetic polymorphism of heme oxygenase 1 promoter in the
occurrence and severity of chronic obstructive pulmonary disease: A
meta-analysis. J Cell Mol Med. 21:894–903. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Fischer BM, Pavlisko E and Voynow JA:
Pathogenic triad in COPD: Oxidative stress, protease-antiprotease
imbalance, and inflammation. Int J Chron Obstruct Pulmon Dis.
6:413–421. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zhang JQ, Zhang JQ, Fang LZ, Liu L, Fu WP
and Dai LM: Effect of oral N-acetylcysteine on COPD patients with
microsatellite polymorphism in the heme oxygenase-1 gene promoter.
Drug Des Devel Ther. 9:6379–6387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Zhang ZY, Guan J, Li H, Zhou ZQ and Zhou
GW: Heme oxygenase-1 promoter polymorphism protects liver
allograft. Indian J Surg. 78:14–19. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Soares M, Lin Y, Anrather J, Csizmadia E,
Takigami K, Sato K, Grey ST, Colvin RB, Choi AM, Poss KD and Bach
FH: Expression of heme oxygenase-1 can determine cardiac xenograft
survival. Nat Med. 4:1073–1077. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
84
|
Katori M, Buelow R, Ke B, Ma J, Coito AJ,
Iyer S, Southard D, Busuttil RW and Kupiec-Weglinski JW: Heme
oxygenase-1 overexpression protects rat hearts from cold
ischemia/reperfusion injury via an antiapoptotic pathway.
Transplantation. 73:287–292. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Ozaki KS, Marques GM, Nogueira E, Feitoza
RQ, Cenedeze MA, Franco MF, Mazzali M, Soares MP, Pacheco-Silva A
and Câmara NO: Improved renal function after kidney transplantation
is associated with heme oxygenase-1 polymorphism. Clin Transplant.
22:609–616. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Nisula S, Kaukonen KM, Vaara ST, Korhonen
AM, Poukkanen M, Karlsson S, Haapio M, Inkinen O, Parviainen I,
Suojaranta-Ylinen R, et al: Incidence, risk factors and 90-day
mortality of patients with acute kidney injury in Finnish intensive
care units: The FINNAKI study. Intensive Care Med. 39:420–428.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Vilander LM, Vaara ST, Donner KM, Lakkisto
P, Kaunisto MA and Pettilä V; FINNAKI Study Group, : Heme
oxygenase-1 repeat polymorphism in septic acute kidney injury. PLoS
One. 14:e02172912019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Vashist YK, Uzungolu G, Kutup A, Gebauer
F, Koenig A, Deutsch L, Zehler O, Busch P, Kalinin V, Izbicki JR
and Yekebas EF: Heme oxygenase-1 germ line GTn promoter
polymorphism is an independent prognosticator of tumor recurrence
and survival in pancreatic cancer. J Surg Oncol. 104:305–311. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Okamoto I, Krögler J, Endler G, Kaufmann
S, Mustafa S, Exner M, Mannhalter C, Wagner O and Pehamberger H: A
microsatellite polymorphism in the heme oxygenase-1 gene promoter
is associated with risk for melanoma. Int J Cancer. 119:1312–1315.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Berberat PO, Dambrauskas Z, Gulbinas A,
Giese T, Giese N, Künzli B, Autschbach F, Meuer S, Büchler MW and
Friess H: Inhibition of heme oxygenase-1 increases responsiveness
of pancreatic cancer cells to anticancer treatment. Clin Cancer
Res. 11:3790–3798. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Sunamura M, Duda DG, Ghattas MH, Lozonschi
L, Motoi F, Yamauchi J, Matsuno S, Shibahara S and Abraham NG: Heme
oxygenase-1 accelerates tumor angiogenesis of human pancreatic
cancer. Angiogenesis. 6:15–24. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Bukowska-Strakova K, Włodek J, Pitera E,
Kozakowska M, Konturek-Cieśla A, Cieśla M, Gońka M, Nowak W,
Wieczorek A, Pawińska-Wąsikowska K, et al: Role of HMOX1 promoter
genetic variants in chemoresistance and chemotherapy induced
neutropenia in children with acute lymphoblastic leukemia. Int J
Mol Sci. 22:9882021. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Tang D, Tang WJ, Shi XL, Li WP, Zhou H, Lu
LM and Tao L: Association of the microsatellite (GT)n repeat
polymorphisms of the HO-1 gene promoter and corresponding serum
levels with the risk of laryngeal squamous cell carcinoma. Acta
Otolaryngol. 136:806–811. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Zhang L, Song FF, Huang YB, Zheng H, Song
FJ and Chen KX: Association between the (GT)n polymorphism of the
HO-1 gene promoter region and cancer risk: A meta-analysis. Asian
Pac J Cancer Prev. 15:4617–4622. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Hu JL, Li ZY, Liu W, Zhang RG, Li GL, Wang
T, Ren JH and Wu G: Polymorphism in heme oxygenase-1 (HO-1)
promoter and alcohol are related to the risk of esophageal squamous
cell carcinoma on Chinese males. Neoplasma. 57:86–92. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Lo SS, Lin SC, Wu CW, Chen JH, Yeh WI,
Chung MY and Lui WY: Heme oxygenase-1 gene promoter polymorphism is
associated with risk of gastric adenocarcinoma and lymphovascular
tumor invasion. Ann Surg Oncol. 14:2250–2256. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Sawa T, Mounawar M, Tatemichi M, Gilibert
I, Katoh T and Ohshima H: Increased risk of gastric cancer in
Japanese subjects is associated with microsatellite polymorphisms
in the heme oxygenase-1 and the inducible nitric oxide synthase
gene promoters. Cancer Lett. 269:78–84. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Kikuchi A, Yamaya M, Suzuki S, Yasuda H,
Kubo H, Nakayama K, Handa M, Sasaki T, Shibahara S, Sekizawa K and
Sasaki H: Association of susceptibility to the development of lung
adenocarcinoma with the heme oxygenase-1 gene promoter
polymorphism. Hum Genet. 116:354–360. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Jirásková A, Novotný J, Novotný L, Vodicka
P, Pardini B, Naccarati A, Schwertner HA, Hubácek JA, Puncochárová
L, Šmerhovský Z and Vítek L: Association of serum bilirubin and
promoter variations in HMOX1 and UGT1A1 genes with sporadic
colorectal cancer. Int J Cancer. 131:1549–1555. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Podkalicka P, Mucha O, Józkowicz A, Dulak
J and Łoboda A: Heme oxygenase inhibition in cancers: Possible
tools and targets. Contemp Oncol (Pozn). 22:32–23. 2018.PubMed/NCBI
|
|
101
|
Kah J, Volz T, Lütgehetmann M, Groth A,
Lohse AW, Tiegs G, Sass G and Dandri M: Haem oxygenase-1
polymorphisms can affect HCV replication and treatment responses
with different efficacy in humanized mice. Liver Int. 37:1128–1137.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Singh D, Wasan H and Reeta KH: Heme
oxygenase-1 modulation: A potential therapeutic target for COVID-19
and associated complications. Free Radic Biol Med. 161:263–271.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Zhou JF, Luo JY, Zhu WB, Yang CY, Zeng YL
and Qiu XL: Association between genetic polymorphism of heme
oxygenase 1 promoter and neonatal hyperbilirubinemia: A
meta-analysis. J Matern Fetal Neonatal Med. 34:12–23. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Wagener FA, Toonen EJ, Wigman L, Fransen
J, Creemers MC, Radstake TR, Coenen MJ, Barrera P, van Riel PL and
Russel FG: HMOX1 promoter polymorphism modulates the relationship
between disease activity and joint damage in rheumatoid arthritis.
Arthritis Rheum. 58:3388–3393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Gulla A, Gulbinas A, Dambrauskas Z and
Strupas K: Heme oxygenase-1 polymorphism is associated with the
development of necrotic acute pancreatitis via vascular cell
adhesion molecule-1 and the E-selectin expression regulation
pathway. Pancreas. 48:787–791. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Lee EY, Lee YH, Kim SH, Chung KS, Kwon O,
Kim BS, Nam CM, Park CS, Lee BW, Kang ES, et al: Association
between heme oxygenase-1 promoter polymorphisms and the development
of albuminuria in type 2 diabetes: A case-control study. Medicine
(Baltimore). 94:e18252015. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Holweg CT, Balk AH, Uitterlinden AG,
Niesters HG, Maat LP, Weimar W and Baan CC: Functional heme
oxygenase-1 promoter polymorphism in relation to heart failure and
cardiac transplantation. J Heart Lung Transplant. 24:493–497. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Foresti R, Bains SK, Pitchumony TS, de
Castro Brás LE, Drago F, Dubois-Randé JL, Bucolo C and Motterlini
R: Small molecule activators of the Nrf2-HO-1 antioxidant axis
modulate heme metabolism and inflammation in BV2 microglia cells.
Pharmacol Res. 76:132–148. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Funes SC, Rios M, Fernández-Fierro A,
Covián C, Bueno SM, Riedel CA, Mackern-Oberti JP and Kalergis AM:
Naturally derived heme-oxygenase 1 inducers and their therapeutic
application to immune-mediated diseases. Front Immunol.
11:14672020. View Article : Google Scholar : PubMed/NCBI
|