Introduction
The largest protein reservoir in the body is the
muscles. Muscles serve as a source of amino acids, which can be
used for energy production by various organs during catabolic
periods in human diseases (1).
However, skeletal muscle can exhibit excessive protein degradation
and the ensuing muscle loss can lead to muscle atrophy (2). Muscle atrophy, also known as muscle
wasting disease, is a disease characterized by the loss of muscle
tissue and progressive muscle weakening. If not sufficiently
restored upon re-ambulation, a patient may experience muscle
weakness, poorer physical function and eventually a reduced life
span (1). Therefore,
understanding the changes and mechanisms that occur during the
early periods of muscle loss is of crucial importance in developing
novel therapeutic targets.
Chronic inflammation, characterized by higher
systemic cytokine and acute phase protein circulation (3,4),
is not only linked to aging (inflammaging) (5), but also loss of muscle mass
(6). Inflammation impairs
myogenesis and muscle homeostasis, activating the family of
forkhead box O (FOXO) transcription factors, which then further
promote skeletal muscle atrophy (7,8).
Conversely, it is well-established that inflammation serves an
important role in developing insulin resistance (IR) (9–11),
and this suggests that insulin signaling may modulate catabolic and
anabolic pathways of muscle atrophy, potentially explaining the
simultaneous occurrence of IR and muscle atrophy in several
patients with type 2 diabetes (12,13). Insulin-like growth factor 1 (IGF1)
and/or insulin signaling can simultaneously inhibit protein
breakdown and enhance muscle growth (14–16). Nevertheless, studies on AKT [also
known as protein kinase B (PKB)] highlight another aspect of the
IGF1 pathway in regulating muscle atrophy. AKT controls protein
synthesis through the mechanistic target of rapamycin (mTOR) and
regulates protein degradation through transcription factors in the
FOXO family. However, pro-inflammatory cytokines (such as TNF-α)
contribute to inhibition of the IGF1-AKT pathway and IR (17–19). In addition, nuclear transcription
factor nuclear factor (NF)-κB, which plays an important role as a
mediator of immunity and inflammation, leads to an increase in the
expression of inflammatory cytokines, particularly TNF-α, during
muscle wasting and cachexia (20).
Indeed, in human clinical trials, ingestion of
sucrose alone by women results in an elevation of glucose
concentration, followed by a high insulin response, reducing
glucose concentrations to baseline and high glucose levels
accompanied by a compensatory elevation in free fatty acid (FFA)
concentration (21). In addition,
FFAs serve a crucial role in the inflammatory process of
cardiovascular diseases, as they serve as the building blocks for
several components (cytokines) that are released through the portal
vein and into the liver and then to other tissues (21,22). Furthermore, increased circulating
levels of FFAs is associated with a 7-fold increase in the risk of
developing type 2 diabetes (22),
due to impaired glucose-stimulated insulin secretion and resistance
to the anti-lipolytic effect of insulin and increased adipose
tissue mass (23). Therefore,
effective strategies of improving IR and diabetes should include
consideration of reduced intake of sucrose, reducing lipid
metabolite accumulation of peripheral tissues and lowering the FFA
levels. Increasing evidence has shown that sugars added to food
products (including sucrose, glucose and fructose), increase the
risk of development of inflammation, increased oxidative stress,
dyslipidemia, hypertension and IR (24,25). Indeed, high chronic sucrose intake
accelerates sarcopenia through an alteration of postprandial
stimulation of muscle protein synthesis in older male rats
(26).
Herbal products contain several phytochemicals, such
as carotenoids and polyphenols, alkaloids, flavonoids, glycosides,
phenolic acids, saponins and lignans, all of which have proven
health benefits (27–44). In Taiwan, herbal formula ATG-125
has been used in traditional therapy as it is believed to treat
numerous ailments, including inflammation, muscle spasms, oral
ulcers, wounds and rheumatic pain for several decades. However,
there are no extensive studies assessing its effectiveness and its
underlying mechanisms in muscle atrophy, to the best of our
knowledge. ATG-125 is a the ethanolic extract of a herbal mixture
consisting of Artemisia argyi (27), Morus alba L. (28), Leonurus japonicus Houtt.
(29), Capsicum annuum L.
(30,31), Lophatherum gracile Brongn.
(32), Curcuma longa
(33,34) and Glycyrrhiza uralensis
(35). Traditionally, these
plants have been used for centuries for their antioxidant,
anti-inflammatory and healing properties. Different classes of
bioactive constituents are present in ATG-125, including several
pseudoalkaloids, phenols and flavonoids, thus resulting in a
pharmaceutical composition comprised of a mixture of chlorogenic
acid (36–38), leonurine (29,39), schaftoside (40), rutin (36), isoschaftoside (41), isochlorogenic acid (38), quercetin (42,43), apigenin (43,44), glycyrrhizic acid (35), curcumin (33,34) and artemisetin (27), all of which contribute to its
medicinal properties.
In the present study, using transcriptomic
screening/biomedical analysis for identification of potential
muscle atrophy regulators in the phytochemical-rich formula
ATG-125, the up- and downregulated genes in a high-sucrose intake
model of muscle atrophy in mice were identified. The role of
ATG-125 in the muscle response to sucrose was explored, with the
present study being focused on muscle homeostasis and
Akt/mitochondria biogenesis signaling in young adult mice.
Materials and methods
Plant materials and extracts
ATG-125 is composed of seven herbal ingredients: i)
Artemisia argyi leaves (154 g, Kuang Ying); ii) Morus
alba L. leaves (154 g, Kuang Ying); iii) Leonurus
japonicus Houtt. leaves (154 g, Kuang Ying); iv) Capsicum
annuum L. leaves (77 g, Kuang Ying); v) Lophatherum
gracile Brongn. leaves (77 g, Kuang Ying); vi) Curcuma
longa root (31 g, San Hsing); and vii) Glycyrrhiza
uralensis root (31 g, Jin Yi Hang). These herbs were purchased
from local herbal medicine stores. To prepare the ethanolic
extracts, 678 g of the herbal ingredients was soaked in 5 liters
95% ethanol (Sigma-Aldrich; Merck KGaA) at room temperature for 1
day and filtered, then the filter procedure was repeated twice.
Extraction was performed for 4 h at 70°C in a water bath and
filtered, which was then repeated twice. All separate extractions
were mixed and concentrated under reduced pressure. The yield was
99.66 g (14.7%) and ATG was stored at 4°C until required. Prior to
use in the in vivo experiments, 1.5 g ATG-125 was dissolved
in 100 ml solvent (95% olive oil and 5% glycerol).
High-performance liquid chromatography
(HPLC)
HPLC-UV analysis was performed using a Waters HPLC
System (Waters Corporation), including a 600 quaternary pump, a
Sugai U-620 column temperature controller, a 717 plus autosampler
and a 996-photodiode array detector. The analytical conditions were
set as follows: gradient elution by the mixture of mobile phases A
(0.085% phosphoric acid) and B (acetonitrile) at 0–10 min with a
ratio of 85–90% A and 10–15% B; 10–20 min with a ratio of 80–85% A
and 15–20% B; 20–30 min with a ratio of 60–80% A and 20–40% B;
30–55 min with a ratio of 35–60% A and 40–65% B; 55–65 min with a
ratio of 0–35% A and 65–100% B; and 65–80 min with the ratio of
0–90% A and 10–100% B. The flow rate was kept constant at 1.0
ml/min. The column temperature was controlled at 35°C, the post
time was 15 min and the injection volume was 20 µl. The UV
wavelength for full detection was set at 200–600 nm. The analytical
column was a Cosmosil 5C18-MS-II column (particle size, 5 µm;
column size, 4.6 mm internal diameter) ×250 mm; Nacalai Tesque,
Inc.) and a Lichrospher RP-18 end-capped column (particle size, 5
µm; column size, 4.0 mm internal diameter ×10 mm; Merck KGaA) was
used as a guard column.
Liquid chromatography-mass
spectrometry (LC-MS) analysis and instrumental conditions
LC-MS was used to identify the major markers of
bioactive substances. The system for analysis consisted of a
LC-20AD UFLC system (Shimadzu Corporation) linked to a LCMS-2020
mass spectrometer. The running conditions were designed as follows:
Gradient elution by the mixture of mobile phases A (0.1% formic
acid in water) and B (Acetonitrile) at minutes 0–65, with the ratio
of 90–0% in A and 10–100% in B; at minutes 65–80 with the ratio of
0–90% in A and 100–10% in B. The flow rate was fixed at 1 ml/min
and the column temperature was maintained at 35°C. The injection
volume was adjusted to 20 µl and the analytical column used was a
Cosmosil 5C18-MS-II (5 µm, 4.6×250 mm; Nacalai Tesque, Inc.). Dual
ion modes [electrospray ionization, (ESI)(+) and (−)] were used in
MS detection and the transmission of (M+H)+ and (M-H)-was set as
the optimum condition. The MS detection was set to full scan range
(50-1,200 m/z). The interface voltages were set at 4.5 kV for
ESI(+) and −4.5 kV for ESI(−). With nitrogen as the nebulizing and
drying gas, the flow rate was set at 1.5 and 15 l/min,
respectively. Nitrogen gas temperature was set at 350°C.
Desolvation line temperature was set at 300°C and heat block
temperature was maintained at 500°C. Chromatographic fingerprint
analysis was performed using 3D HPLC, which was primarily performed
as described previously (45,46).
Animals
A total of 15 male young adult C57BL/6J mice (age, 6
months; weight, 24–26 g) were obtained from the National Laboratory
Animal Center (Taiwan) and provided ad libitum access to
standard chow and water and housed (40–60% humidity) with a
controlled temperature (26°C) and a 12-h light/dark cycle (lights
off at 7 pm). Animal studies were performed according to previous
studies (47,48) and were approved by the Chang Gung
University Animal Care and Use Committee (approval no. CGU108-111;
Taoyuan, Taiwan). Mice were randomized into three groups (n=5 per
group): i) Young adult mice; ii) young adult mice fed with 30%
sucrose for 28 days; and iii) young adult mice fed with 30% sucrose
and ATG-125 (0.2 ml applied each day to the abdominal skin) on days
15–28. All mice were maintained on their assigned diet until they
were sacrificed by CO2 (CO2 flow rate, volume
displacement at 70% of the cage vol/min).
Western blot analysis
Gastrocnemius muscles were harvested immediately,
flash frozen in liquid nitrogen and stored at −80°C. Protein was
extracted from muscle tissues using distilled water using a T-PER™
Tissue Protein Extraction Reagent (0.30 mg tissue/200 µl; cat. no.
78510; Thermo Fisher Scientific, Inc.) containing protease
inhibitors (1 µl/ml; cat. no. SI-P8340; Sigma-Aldrich; Merck KGaA),
and the protein concentration was determined using a Bio-Rad
protein assay (Bio-Rad Laboratories, Inc.). Blocking was performed
using 5% w/v nonfat dry milk, 1X TBS with 0.1% Tween-20 at room
temperature for 1 h. Protein (50 µg/lane) was run on a 10% SDS-PAGE
gel and subsequently transferred to a PVDF membrane. For
immunoblotting, membranes were incubated with diluted primary
antibody in 5% w/v nonfat dry milk, 1X TBS with 0.1% Tween-20 at
4°C with gentle shaking overnight. The following primary antibodies
were used: Phosphorylated (p)-AMP-activated protein kinase (1:500;
p-AMPK; Thr 172; cat. no. sc-33524; Santa Cruz Biotechnology,
Inc.), AMPK (1:500; cat. no. sc-74461; Santa Cruz Biotechnology,
Inc.), p-FOXO3a (1:500; S253; cat. no. ab47285; Abcam), FOXO3a
(1:500; cat. no. ab17026; Abcam), muscle RING-finger protein-1
(MuRF1; 1:1,000; cat. no. ab77577; Abcam), β-actin (1:1,000; cat.
no. MAB1501; MilliporeSigma), peroxisome proliferator-activated
receptor (PPAR)γ coactivator 1-α (PGC1α; 1:1,000; cat. no. ab54481;
Abcam), Sirtuin 1 (SIRT1; 1:1,000; cat. no. ab110304; Abcam),
uncoupling protein (UCP)1 (1:1,000; cat. no. ab10983; Abcam), UCP2
(1:500; cat. no. ab203244; Abcam) and UCP3 (1:500; cat. no.
ab10985; Abcam). The primary incubation was followed by incubation
with secondary antibodies in 5% w/v nonfat dry milk, 1X TBS with
0.1% Tween-20 at room temperature with gentle shaking for 1 h. The
following secondary antibodies were used: Goat anti-mouse IgG
antibody, HRP-conjugated (1:1,000; cat. no. Ap124P;
MilliporeSigma), goat anti-rabbit IgG antibody, biotin-SP (long
spacer)-conjugated (1:1,000; cat. no. 111-065-003; Jackson
ImmunoResearch Laboratories, Inc.) and goat anti-rat IgG antibody,
biotin-SP-conjugated (1:1,000; cat. no. 112-065-003; Jackson
ImmunoResearch Laboratories, Inc.). Signals were visualized using a
chemiluminescence system (Amersham; Cytiva) according to the
manufacturer's instructions. The optical densities of the bands
were semi-quantified using ImageJ version 1.53 (National Institutes
of Health).
Histology, immunohistochemistry and
immunofluorescence
Muscle tissue was fixed in 4% paraformaldehyde
solution for 3 days, processed for paraffin embedding and cut into
5-µm thick sections at room temperature. For histology, sections
were stained with hematoxylin (2 g/l) for 15 min and with eosin
(0.1% in 0.0003% acetic acid) for 10 min at room temperature
(49). Before immunostaining,
slides were heated in an oven at 65°C for 15 min and the sections
were deparaffinized using xylene and placed in a series of graded
alcohol solutions, as routine dehydrated processing. The endogenous
peroxidase activity was blocked by hydrogen peroxidase treatment
(3%). The sections were then washed with distilled water and
transferred to TBST (0.5% Tween-20; pH 7.4) containing 5% normal
goat serum (cat. no. G9023; Sigma-Aldrich; Merck KGaA) for 30–50
min at room temperature. Next, sections were incubated at room
temperature for 2 h with antibodies against p-AMPK (1:100), glucose
transporter type 4 insulin-responsive (Glut4; 1:80; cat. no.
ab33780; Abcam), p-FOXO3a (1:100), FOXO3A (1:100), MuRF1(1:100),
NF-κB (1:100; cat. no. sc-8008; Santa Cruz Biotechnology, Inc.),
hypoxia inducible factor-1α (HIF-1α; 1:100; cat. no. ab179483;
Abcam), vascular endothelial growth factor (VEGF; 1:100; cat. no.
ab69479; Abcam), TGFβ receptor type-II (TGFβRII; 1:100; cat. no.
sc-17791; Santa Cruz Biotechnology, Inc.), p-p53 (S15; 1:100; cat.
no. ab1431; Abcam), Ki67 (1:100; cat. no. AB9260; MilliporeSigma),
PGC1α (1:100), UCP1 (1:100), UCP2 (1:100; cat. no. ab97931; Abcam)
UCP3 (1:100), p-insulin-like growth factor 1 receptor (IGF1R;
Y1161; 1:100; cat. no. ab39398; Abcam), p-insulin receptor
substrate 1 (IRS1; Tyr 989; 1:100; cat. no. sc-17200; Santa Cruz
Biotechnology, Inc.), p-PI3K (p85; 1:100; cat. no. GTX111068;
GeneTex, Inc.), p-AKT (Ser473; 1:100; cat. no. sc-7985-R; Santa
Cruz Biotechnology, Inc.), p-mTOR (Ser 2448; 1:100; cat. no.
ab109268; Abcam), p-ribosomal S6 kinase (S6K; Thr389; 1:100; cat.
no. 9234; Cell Signaling Technology, Inc.), p-eukaryotic
translation initiation factor 4E-binding protein 1 (4EBP1;
Thr37/46l; 1:100; cat. no. 2855; Cell Signaling Technology, Inc.)
and MitoTracker™ Red CMXRos (1:100; cat. no. M7512; Thermo Fisher
Scientific, Inc.). Sections were incubated with the appropriate
secondary antibodies, including Alexa Fluor 488-conjugated rabbit
IgG (heavy chain + light chain) highly cross adsorbed (1:100; cat.
no. A11034; Thermo Fisher Scientific, Inc.), Alexa Fluor Plus
647-conjugated donkey anti-rabbit (1:100; cat. no. A32795; Thermo
Fisher Scientific, Inc.), goat-anti-mouse IgG HRP-conjugated (H+L;
1:100; cat. no. G-21040; Thermo Fisher Scientific, Inc.) or goat
anti-rabbit IgG HRP-conjugated (H+L; 1:100; cat. no. G-21234;
Thermo Fisher Scientific, Inc.) antibodies for 1 h at room
temperature. All antibodies were diluted in 2% non-immune goat
serum (cat. no. G9023; Sigma-Aldrich; Merck KGaA) in PBS with
Tween-20 (0.05%) buffer at room temperature. Sections were
incubated with DAB (MilliporeSigma) for 5–10 min, and hematoxylin
(Sigma-Aldrich; Merck KGaA) for 3 min, or DAPI (1:10,000; cat. no.
62248; Thermo Fisher Scientific, Inc.) was used for nuclear
staining, and both staining incubations were performed at room
temperature. The optical observed of the stain using CellSens
Dimension software package (Olympus Corporation) and an inverted
fluorescence microscope (Olympus IX71; Olympus Corporation).
RNA isolation and reverse
transcription-quantitative (RT-q)PCR
Total RNA was isolated from 0.2 g muscle tissues
using TRIzol® (Thermo Fisher Scientific, Inc.) and an
RNeasy kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. For qPCR, equal quantities of RNA were
reverse transcribed to cDNA using a High-Capacity cDNA Reverse
Transcription kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The following thermocycling conditions
were used for qPCR: Initial denaturation at 95°C for 5 min; 35 of
cycles of denaturation at 94°C for 30 sec, annealing at 55.8°C for
30 sec 0.5 min, elongation at 72°C for 0.5 min; final extension at
72°C for 30 sec 0.5 min); and holding at 4°C. The expression levels
of Atrogin-1, NADPH oxidase (Nox)−1, Sterol
regulatory element-binding protein (Srebp)−1,
Carnitine palmitoyltransferase I (Cpt-1), Pparα,
Pparγ, acyl-CoA oxidase (Aco), mitochondrial
transcription factor A (Tfam), Sirt1, Pgc1α, Nuclear
respiratory factor 1 (Nrf1), Igf1r, Irs1, Pi3k, Akt,
mTOR, S6k, 4ebp1 and Gapdh were evaluated using SYBR
Green PCR MasterMix (Roche Diagnostics GmbH). Gapdh was used
as the loading control. Relative expression was calculated using
the 2−∆∆Cq method with normalization to constitutive
genes (50). The sequences of the
primers used for RT-qPCR are listed in Table SI (51–77).
RNA isolation and library preparation
for RNA-Seq
Total RNA was extracted from gastrocnemius muscles
using a Qiagen RNeasy Mini Kit (cat. no. 74004; Qiagen GmbH). RNA
purity and quality were evaluated using a SimpliNano™ Biochrom
Spectrophotometer (cat. no. 29061712; Biochrom, Ltd.). A total of 1
µg total RNA/sample was used as input material for the RNA sample
preparations. In order to select cDNA fragments of ~150 bp in
length, the library fragments were purified using the KAPA Pure
Beads System (cat. no. 07983298001; KAPA Biosystems; Roche
Diagnostics GmbH). cDNA was synthesized using a KAPA mRNA HyperPrep
Kit (cat. no. 08098123702; Kapa Biosystems; Roche Diagnostics GmbH)
according to the manufacturer's protocol and index codes. The
clustering of the index-coded samples was performed on a cBot
Cluster Generation System using PE Cluster Kit cBot-HS (cat. no.
PE-401-3001; Illumina, Inc.) according to the manufacturer's
instructions. Following cluster generation, the library
preparations were sequenced using an Illumina platform (NovaSeq
6000 S4 Reagent Kit v1.5; 300 cycles; cat. no. 20028312; Illumina,
Inc.) and 125 bp/150 bp paired-end reads were generated. The
original data obtained by high-throughput sequencing (Illumina
NovaSeq 6000 platform; Illumina, Inc.) were transformed into raw
sequenced reads by CASAVA base calling and stored in the FASTQ
format, and FastQC, MultiQC and ReSeqtools were used to check FASTQ
files for quality (78). The
final library qubit concentration was 3.74 ng/µl.
Raw paired-end reads were filtered by Trimmomatic
(version 0.38) (79). Next, the
genome alignment methods from hierarchical indexing for spliced
alignment of transcripts (HISAT2; version 2.1.0) (80), RSeQC (version 4.0.0) (81), Qualimap (version 2.2.1) (82) and featureCounts (version 2.0.0)
(83) were used. Gene expression
values were represented using the normalization techniques provided
by each algorithm: Fragments per Kilobase of Mapped reads (FPKM),
Transcripts per Million (TPM), Trimmed Mean of M values (TMM from
edgeR) and Relative Log Expression (RLE from DESeq2) (78,84). Gene function of the mapped reads
(unique transcripts) was annotated based on the NCBI non-redundant
protein sequences database (85).
Resulting P-values were adjusted using the Benjamini and Hochberg's
approach for controlling the false discovery rate (FDR) (86). Differentially expressed genes
(DEGs) with a threshold FDR adjusted, P.adjust <0.05 and
fold-change 2 (log2>±1) were selected for further analysis.
Principal component analysis (PCA) (84), volcano plots ggplot2 version 3.3.3
(R package) (87), Gene Ontology
(GO) (88,89), Kyoto Encyclopedia of Genes and
Genomes (KEGG) (89), Disease
Ontology (DO) (90) and Gene Set
Enrichment Analysis (GSEA) (91)
enrichment of the DEGs were analyzed. ClusterProfiler package
(version 3.14.3) supported the enrichment analysis of GO and KEGG
with either hypergeometric tests or GSEA (91). Furthermore, the function emapplot
and heatmap diagram were also from the enrichplot package of
ClusterProfiler (91).
Statistical analysis
All data are presented as the mean ± standard error
of the mean. Statistical analysis was performed using SPSS version
21.0 (IBM Corp.). All experiments were repeated three times for
data analysis. All data were analyzed using a one-way ANOVA
followed by a Bonferroni's post hoc test. P<0.05 was considered
to indicate a statistically significant difference.
Results
LC-MS analysis of the composition of
ATG-125
A typical LC-MS chromatographic fingerprint profile
of ATG-125 was obtained (Fig. 1)
and 10 major components were identified. The well-separated peaks
with retention times of 60 min were identified as follows: i)
Chlorogenic acid (12.82 min); ii) Leonurine (20.375 min); iii)
Rutin (27.060 min); iv) Isoschaftoside (31.078 min); v)
Isochlorogenic acid (3,5-Dicaffeoylquinic acid, 31.52 min); vi)
Quercetin (36.836 min); vii) Apigenin (39.271 min); viii)
Glycyrrhizic acid (41.667 min); ix) Curcumin (50.946 min); and x)
Artemisetin (52.064) (Table I).
The structure of compounds (Fig.
1A-D) and the 3D-HPLC fingerprint (Fig. 1E and F) are shown.
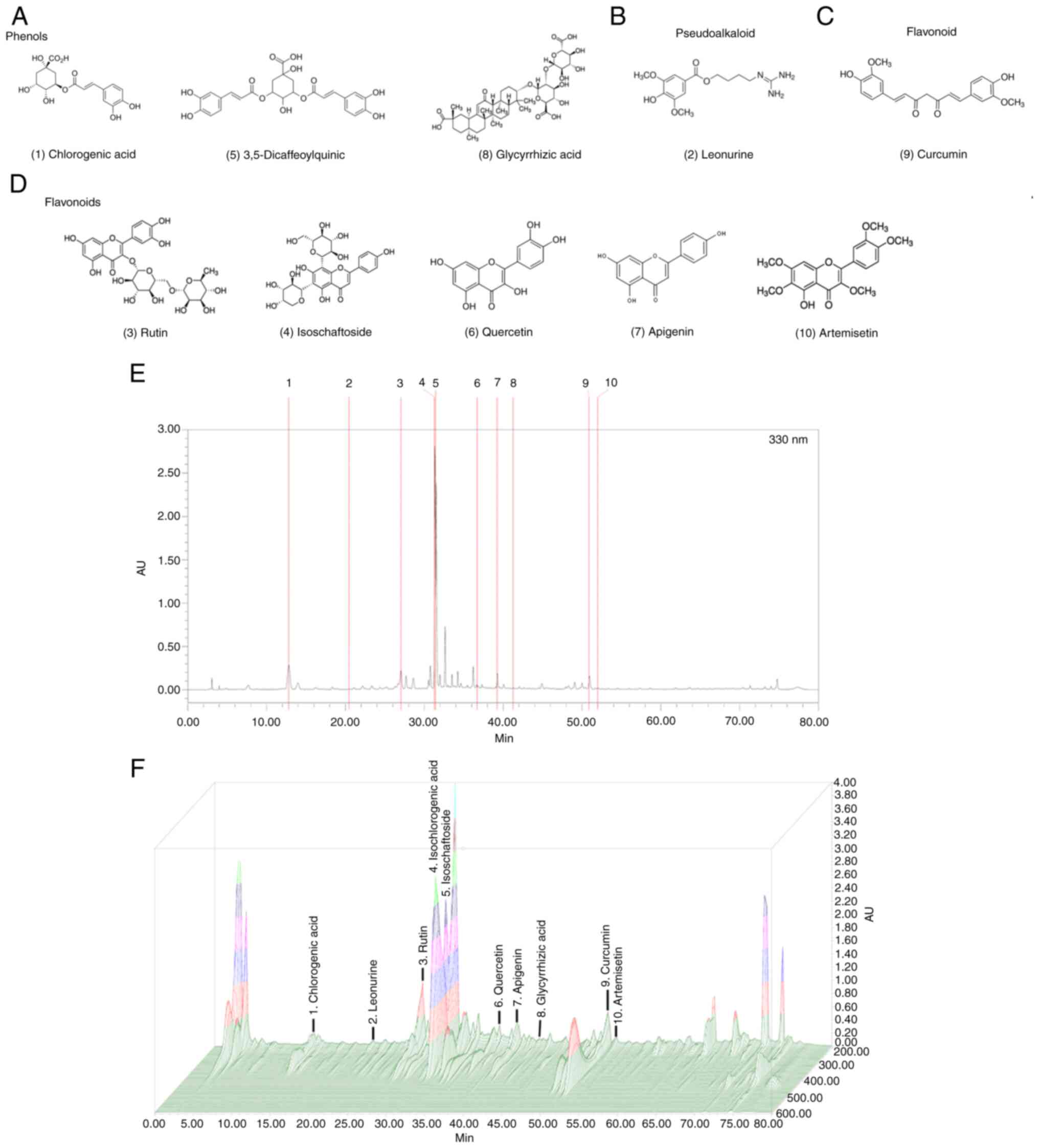 | Figure 1.Chromatogram of the herbal formula
ATG-125 obtained by LC-MS. (A-D) Structures of the compounds
identified by LC-MS. Chlorogenic acid, Leonurine, Rutin,
Isoschaftoside, Isochlorogenic acid (3,5-Dicaffeoylquinic acid),
Quercetin, Apigenin, Glycyrrhizic acid, Curcumin and Artemisetin.
(E) HPLC and (F) 3D-HPLC fingerprints. Chromatographic fingerprint
analysis was performed using HPLC and LC-MS. HPLC, high performance
liquid chromatography; LC-MS, liquid chromatography-mass
spectrometry. |
 | Table I.Percentage of chlorogenic acid,
leonurine, rutin, isoschaftoside, isochlorogenic acid
(3,5-dicaffeoylquinic acid), quercetin, apigenin, glycyrrhizic
acid, curcumin and artemisetin. |
Table I.
Percentage of chlorogenic acid,
leonurine, rutin, isoschaftoside, isochlorogenic acid
(3,5-dicaffeoylquinic acid), quercetin, apigenin, glycyrrhizic
acid, curcumin and artemisetin.
| No. | Retention time
(min) | Compound | Wavelength
(nm) | Weight of compound
in ATG-125 (mg/g) | Percentage of
compound in ATG-125 (%) |
|---|
| 1 |
12.82 | Chlorogenic
acid | 327 |
3.82 | 0.38 |
| 2 | 20.375 | Leonurine | 280 |
0.40 | 0.04 |
| 3 | 27.060 | Rutin | 350 |
3.64 | 0.36 |
| 4 | 31.078 | Isoschaftoside | 327 | 20.34 | 2.03 |
| 5 |
31.52 | Isochlorogenic acid
(3,5-Dicaffeoylquinic acid) | 327 | 11.83 | 1.18 |
| 6 | 36.836 | Quercetin | 370 |
0.06 | 0.01 |
| 7 | 39.271 | Apigenin | 330 |
0.64 | 0.06 |
| 8 | 41.667 | Glycyrrhizic
acid | 250 |
0.89 | 0.09 |
| 9 | 50.946 | Curcumin | 430 |
2.98 | 0.30 |
| 10 | 52.064 | Artemisetin | 350 |
0.11 | 0.01 |
Global transcriptome characteristics
of ATG-125 on sucrose--induced young adult muscle atrophy in
mice
To investigate the molecular-level changes in the
gastrocnemius muscles of sucrose-induced muscle atrophy in mice
treated with or without ATG-125, RNA-seq analysis of the
gastrocnemius muscle lysates was performed (Fig. 2A). Principal Component Analysis
revealed distinct gene expression profiles for the three different
groups (Fig. 2B). The gene count
data were used to analyze differences in gene expression using
DESeq2. A total of 136 DEGs were identified, including 18
significantly upregulated and 118 significantly downregulated genes
between the ATG-125 treated and untreated sucrose-induced groups. A
volcano plot of the DEGs is shown in Fig. 2C. The top 18 upregulated DEGs were
identified: Acyl-CoA thioesterase 2, angiopoietin-like 4,
adipocyte-related X-chromosome expressed sequence 2, expressed
sequence BB365896, bone morphogenetic protein 3, CDC14 cell
division cycle 14A, ChaC cation transport regulator 1, cell
death-inducing DFFA-like effector c, RIKEN cDNA E230001N04 gene,
growth arrest and DNA-damage-inducible 45 b, G protein-coupled
receptor 160, methyltransferase like 21E, Nmrk2, pyruvate
dehydrogenase kinase isoenzyme 4, pyruvate dehydrogenase
phosphatase catalytic subunit 2, solute carrier family 7 (cationic
amino acid transporter, y+ system) member 10, Ucp3 and zinc
finger and BTB domain-containing protein 16, all of which were
significantly increased in the ATG-125 treated sucrose-induced mice
compared with the sucrose challenged mice (Table II). The top 14 downregulated DEGs
were α-2-HS-glycoprotein, apolipoprotein (Apo)a1, Apoa2,
Apoc3, cytochrome P450 family 3 subfamily A polypeptide 11,
fatty acid binding protein 1, fibrinogen β chain, methionine
adenosyltransferase 1a, major urinary protein 3, serine (or
cysteine) peptidase inhibitor clade A member (Serpina)1b,
Serpina1d, Serpina1e, Serpina3k and solute carrier organic
anion transporter family member 1B2, all of which were
significantly decreased in the ATG-125 treated sucrose induced mice
compared with the sucrose-challenged mice (Table III).
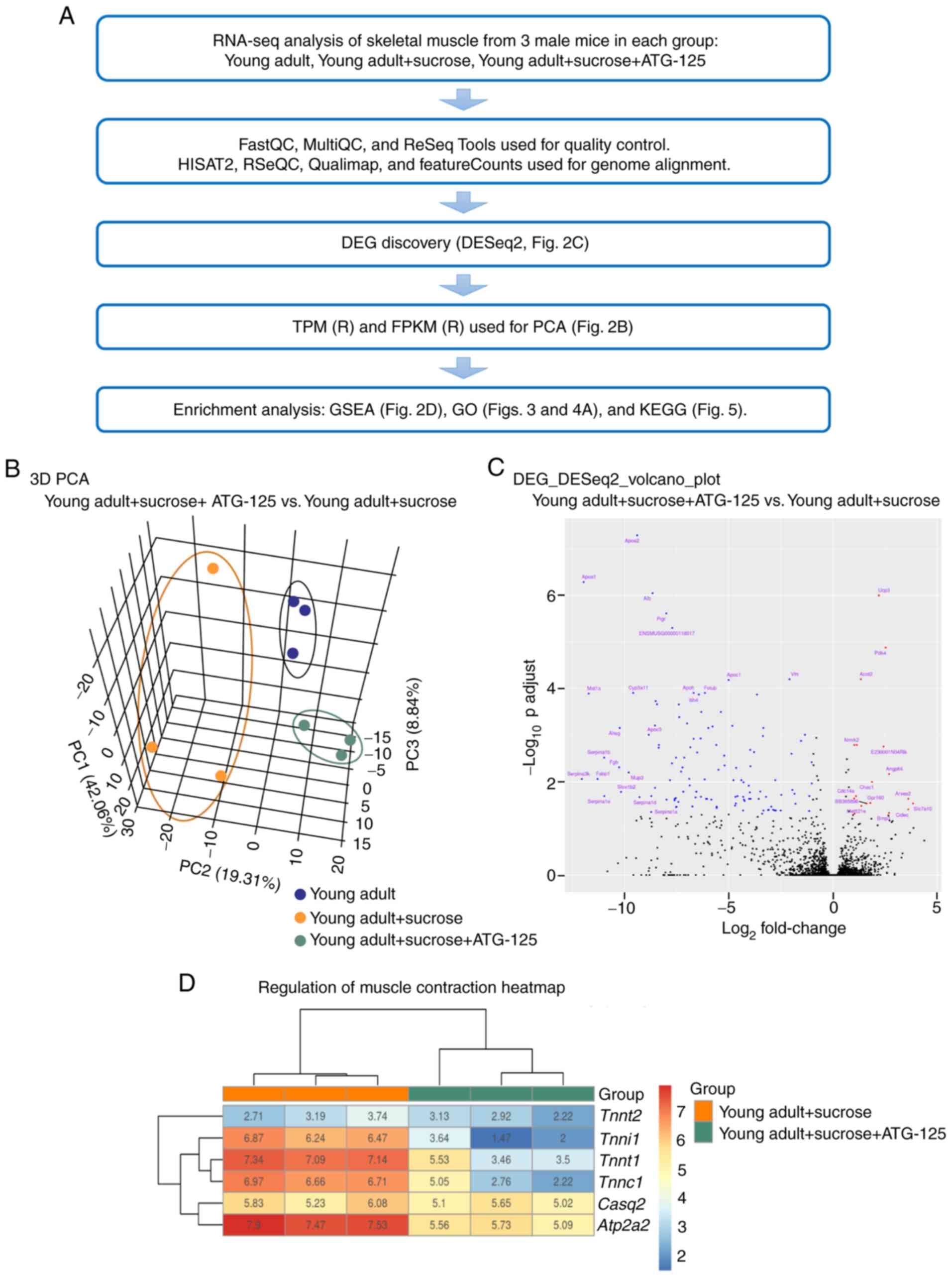 | Figure 2.Transcriptome characteristics of mice
muscles in the sucrose-induced muscle atrophy group with or without
ATG-125 treatment. (A) RNA-seq analysis workflow. (B) PCA showing
overall gene expression patterns in young adult mice,
sucrose-induced muscle atrophy mice and ATG-125 treated
sucrose-induced muscle atrophy mice. (C) Volcano plot of the DEGs.
(D) Heatmap showing genes associated with muscle contraction. PCA,
Principle Component Analysis; DEG, differentially expressed gene;
FPKM, Fragments per Kilobase of Mapped reads; TPM, Transcripts per
Million; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and
Genomes; GSEA, Gene Set Enrichment Analysis; Tnnt2, troponin
T cardiac muscle; Tnnt1, troponin T slow skeletal muscle;
Tnni1, troponin I slow skeletal muscle; Casq2,
calsequestrin 2; Atp2a2, ATPase sarcoplasmic/endoplasmic
reticulum Ca2+ transporting 2. |
| Table II.Top 18 upregulated differentially
expressed genes. |
Table II.
Top 18 upregulated differentially
expressed genes.
| Symbol | Description | log2
Fold-change | P-value | padj |
|---|
| Acot2 | acyl-CoA
thioesterase 2 | 1.34541 |
3.05×10−8 |
6.31×10−5 |
| Angptl4 | angiopoietin-like
4 | 2.70437 |
2.45×10−5 | 0.00680 |
| Arxes2 | adipocyte-related
X-chromosome expressed sequence 2 | 3.62662 | 0.00014 | 0.02329 |
|
BB365896 | expressed sequence
BB365896 | 1.61957 | 0.00020 | 0.02922 |
| Bmp3 | bone morphogenetic
protein 3 | 2.69999 | 0.00037 | 0.04651 |
| Cdc14a | CDC14 cell division
cycle 14A | 1.10848 | 0.00011 | 0.02031 |
| Chac1 | ChaC cation
transport regulator 1 | 1.87761 |
4.18×10−5 | 0.00997 |
| Cidec | cell death-inducing
DFFA-like effector c | 3.62151 | 0.00029 | 0.03906 |
|
E230001N04Rik | RIKEN cDNA
E230001N04 gene | 2.43998 |
3.86×10−6 | 0.00175 |
| Gadd45b | growth arrest and
DNA-damage-inducible 45 b | 1.03662 | 0.00039 | 0.04830 |
| Gpr160 | G protein-coupled
receptor 160 | 1.79903 | 0.00019 | 0.02845 |
|
Mettl21e | methyltransferase
like 21E | 1.37094 | 0.00023 | 0.03285 |
| Nmrk2 | nicotinamide
riboside kinase 2 | 1.15367 |
3.47×10−6 | 0.00162 |
| Pdk4 | pyruvate
dehydrogenase kinase isoenzyme 4 | 2.53921 |
4.99×10−9 |
1.33×10−5 |
| Pdp2 | pyruvate
dehydrogenase phosphatase catalytic subunit 2 | 1.02102 | 0.00013 | 0.02283 |
| Slc7a10 | solute carrier
family 7 (cationic amino acid transporter, y+ system) member
10 | 3.84436 | 0.00020 | 0.02922 |
| Ucp3 | uncoupling protein
3 | 2.20918 |
2.18×10−10 |
1.01×10−6 |
| Zbtb16 | zinc finger and BTB
domain containing 16 | 1.03085 |
3.44×10−6 | 0.00162 |
| Table III.Top 14 downregulated differentially
expressed genes. |
Table III.
Top 14 downregulated differentially
expressed genes.
| Symbol | Description | log2
Fold-change | P-value | padj |
|---|
| Ahsg |
α-2-HS-glycoprotein | −10.24080 |
1.09×10−6 | 0.00070 |
| Apoa1 | apolipoprotein
A-I | −11.97320 |
5.62×10−11 |
5.23×10−7 |
| Apoa2 | apolipoprotein
A-II | −9.40264 |
2.80×10−12 |
5.21×10−8 |
| Apoc3 | apolipoprotein
C-III | −8.84633 |
1.61×10−6 | 0.00098 |
| Cyp3a11 | cytochrome P450
family 3 subfamily a polypeptide 11 | −9.59534 |
8.70×10−8 | 0.00012 |
| Fabp1 | fatty acid binding
protein 1 | −11.30380 |
3.29×10−5 | 0.00867 |
| Fgb | fibrinogen β
chain | −10.26910 |
1.55×10−5 | 0.00487 |
| Mat1a | methionine
adenosyltransferase I α | −11.72980 |
9.67×10−8 | 0.00013 |
| Mup3 | major urinary
protein 3 | −9.80745 |
2.20×10−5 | 0.00628 |
|
Serpina1b | serine (or
cysteine) peptidase inhibitor clade A member 1B | −10.98830 |
8.24×10−6 | 0.00301 |
|
Serpina1d | serine (or
cysteine) peptidase inhibitor clade A member 1D | −9.29357 | 0.00012 | 0.02139 |
|
Serpina1e | serine (or
cysteine) peptidase inhibitor clade A member 1E | −10.98070 | 0.00010 | 0.02026 |
|
Serpina3k | serine (or
cysteine) peptidase inhibitor clade A member 3K | −12.06030 |
3.31×10−5 | 0.00867 |
| Slco1b2 | solute carrier
organic anion transporter family member 1b2 | −10.17570 |
7.99×10−5 | 0.01651 |
Given that the distinction between the ATG-125
treated and untreated clusters were most evident in the
gastrocnemius muscle, the muscle contraction genes in gastrocnemius
muscle that varied according to ATG-125 treatment were probed. To
further determine the differences between the two biological
states, GSEA was performed using the ClusterProfiler package. A
heatmap of the muscle contraction genes showed that they displayed
a steady decreasing trend with ATG-125 treatment. Of these, ATG-125
treatment significantly downregulated the mRNA levels of ATPase
sarcoplasmic/endoplasmic reticulum Ca2+ transporting 2,
troponin I slow skeletal muscle (Tnni1), troponin T cardiac
muscle (Tnnt2) and troponin T slow skeletal muscle
(Tnnt1) in gastrocnemius muscle compared with the sucrose
group (Fig. 2D).
Effect of ATG-125 on DEGs and
functional variation in sucrose-induced young adult muscle atrophy
in mice
To further analyze and classify the biological
function of DEGs, GO functional analysis was performed using the
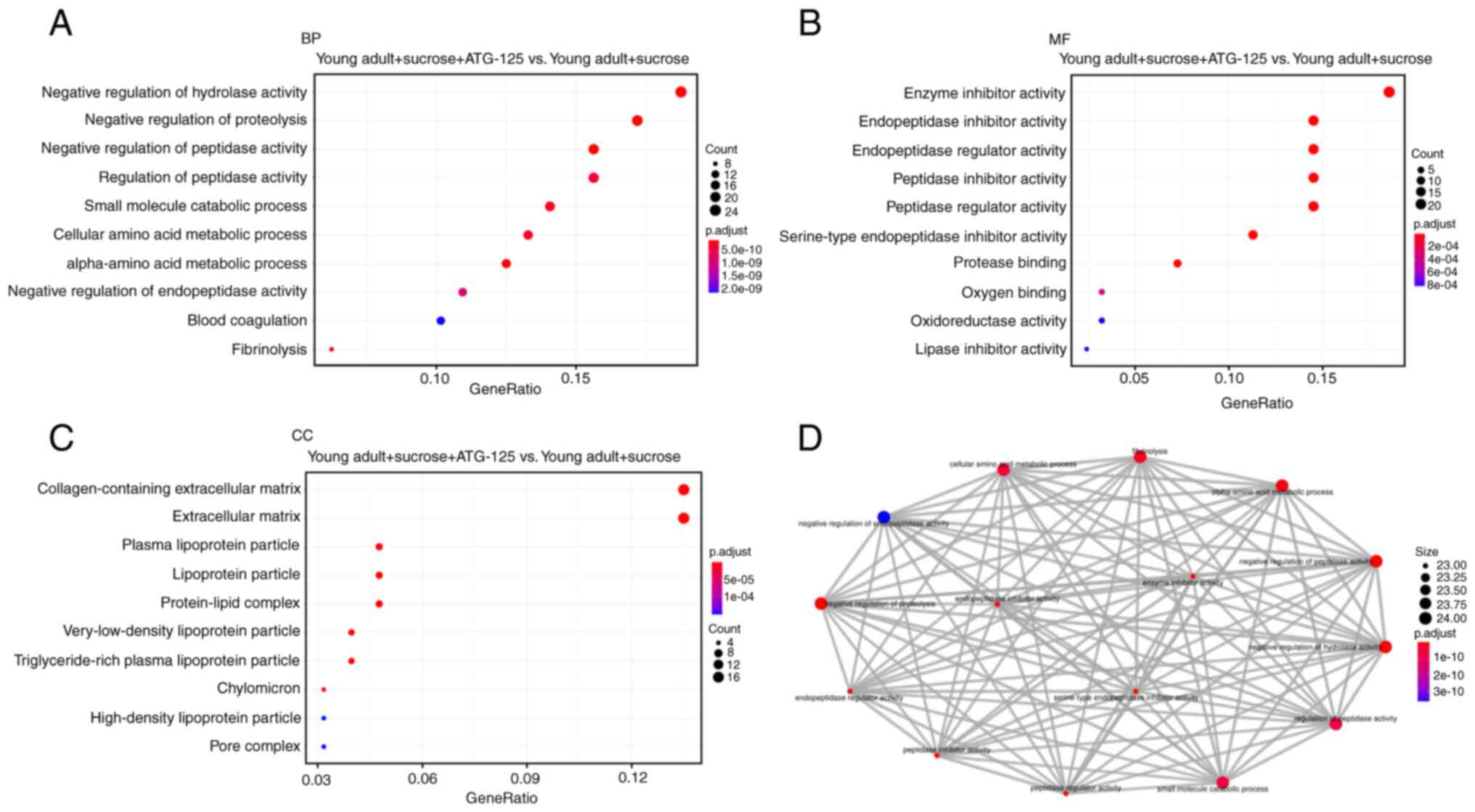
ClusterProfiler package. GO analysis of the DEGs were sub-divided
into three categories: Biological process (BP; Fig. 3A), molecular function (MF;
Fig. 3B) and cellular component
(CC; Fig. 3C) and only
significant GO terms with P-adjust <1 were considered. Top 10
(including downregulated and upregulated) gene-enriched GO terms
were identified and are shown in Fig.
3. In the BP group, the top 2 GO terms were ‘negative
regulation of hydrolase activity’ and ‘negative regulation of
proteolysis’ (Fig. 3A). In the MF
group the top GO term was ‘enzyme inhibitor activity’ (Fig. 3B). In the CC group, the top 2 GO
terms were ‘collagen-containing extracellular matrix’ and
‘extracellular matrix’ (Fig.
3C).
To analyze the functional significance of the
proteins with altered expression corresponding to the ATG-125
treatment, GO Enrichment Map (emapplot) was used to identify
functional networks, which were visualized using ClusterProfiler.
Fig. 3D provides a graphical
overview of the GO Enrichment Map results and indicates that the
altered behavior in response to ATG-125 treatment was observed for
different groups of proteins. Using the GO Enrichment Map plugin
gene function prediction, the gene interaction network was divided
into seven subgroups, which were associated with: i) ‘α-amino acid
metabolic process’; ii) ‘cellular amino acid metabolic process’;
iii) ‘fibrinolysis’; iv) ‘negative regulation of hydrolase
activity’; v) ‘negative regulation of peptidase activity’; vi)
‘negative regulation of proteolysis’; and vii) ‘small molecule
catabolic process’. This category emapplot was supported by the GO
analysis (Fig. 3D).
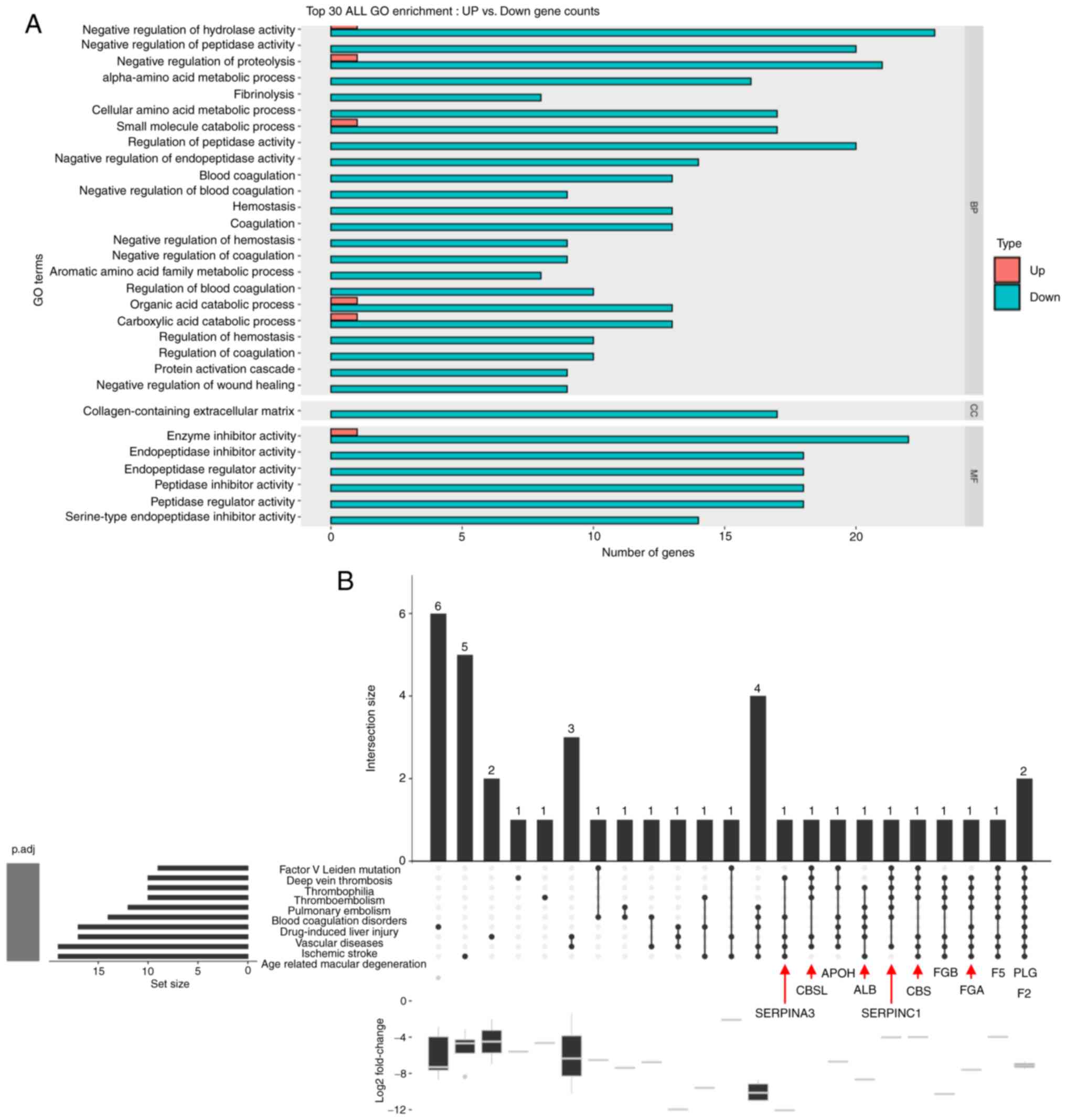
GO enrichment and DO analysis of
ATG-125-regulated DEGs
For GO analysis, 2,203 DEGs from gastrocnemius
muscle were classified into three GO categories (BP, CC and MF).
The top 30 (downregulated and upregulated) gene-enriched GO terms
were identified and are shown in Fig.
4A. GO enrichment analysis was performed to determine whether
the genes in the GO terms were enriched in ‘negative regulation of
hydrolase activity’, ‘negative regulation of peptidase activity’,
‘enzyme inhibitor activity’, ‘endopeptidase inhibitor activity’,
‘negative regulation of proteolysis’, ‘endopeptidase regulator
activity’, ‘peptidase inhibitor activity’, ‘peptidase regulator
activity’ and ‘serine-type endopeptidase inhibitor activity’.
Next, a single asserted classification was used to
include multiple-inferred mechanistic disease classifications from
DO analysis of DEGs, which was performed using the top 10 DO terms
in Fig. 4B. The results showed
that Serpina3 and its interacting genes were significantly
enriched in deep vein thrombosis, blood coagulation disorders,
vascular diseases, ischemic stroke and age-related macular
degeneration based on DO analysis (Fig. 4B).
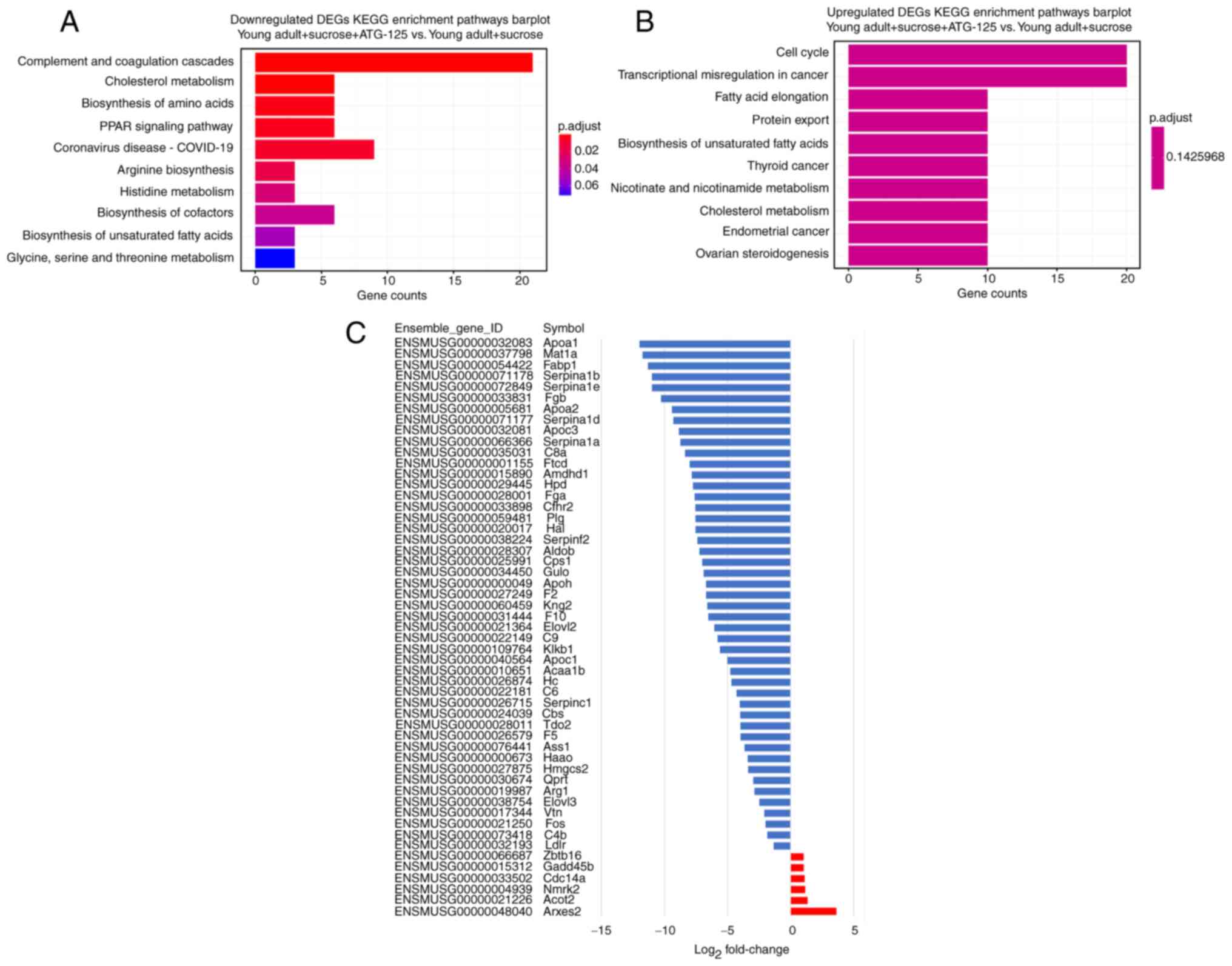
KEGG analysis of DEGs following
ATG-125 treatment of sucrose-induced muscle atrophy
The KEGG enrichment analysis classified the top 10
downregulated and upregulated pathways of the DEGs (Fig. 5A and B). The top downregulated 4
enriched pathways in the gastrocnemius muscle were ‘complement and
coagulation cascades’, ‘cholesterol metabolism’, ‘biosynthesis of
amino acids’ and ‘PPAR signaling pathway’. The top upregulated 4
enriched pathways in the gastrocnemius muscle were ‘cell cycle’,
‘transcriptional misregulation in cancer’, ‘fatty acid elongation’
and ‘protein export’ (Fig. 5A and
B). Examination of the KEGG enrichment analysis showed that
85–90% of the expressed genes were significantly downregulated in
the ATG-125 treated sucrose-induced mice (Fig. 5C). These enriched terms
highlighted the mechanisms regulated by ATG-125 and provide
direction for further research.
ATG-125 attenuates the
AMPK/FOXO3a/MuRF1 signaling pathway in sucrose-induced
gastrocnemius muscle atrophy
In the present study, the body weight of mice fed
sucrose did not differ significantly compared with the young adult
group (Fig. 6A). Mice fed sucrose
exhibited a significant decrease in gastrocnemius muscle weight, as
well as in the ratio of gastrocnemius muscle to body weight
compared with the young adult group (Fig. 6B and C), whereas ATG-125 treatment
significantly increased muscle weight and the ratio of body weight
to muscle compared with the sucrose group (Fig. 6B and C). Mice fed sucrose
exhibited increased quantities of pre-necrotic hyaline and necrotic
fibers, and ATG-125 treatment notably reduced sucrose-induced
muscle atrophy (Fig. 6D). Sucrose
markedly increased gastrocnemius muscle GLUT-4 content but this
markedly decreased after ATG-125 treatment (Fig. 6D).
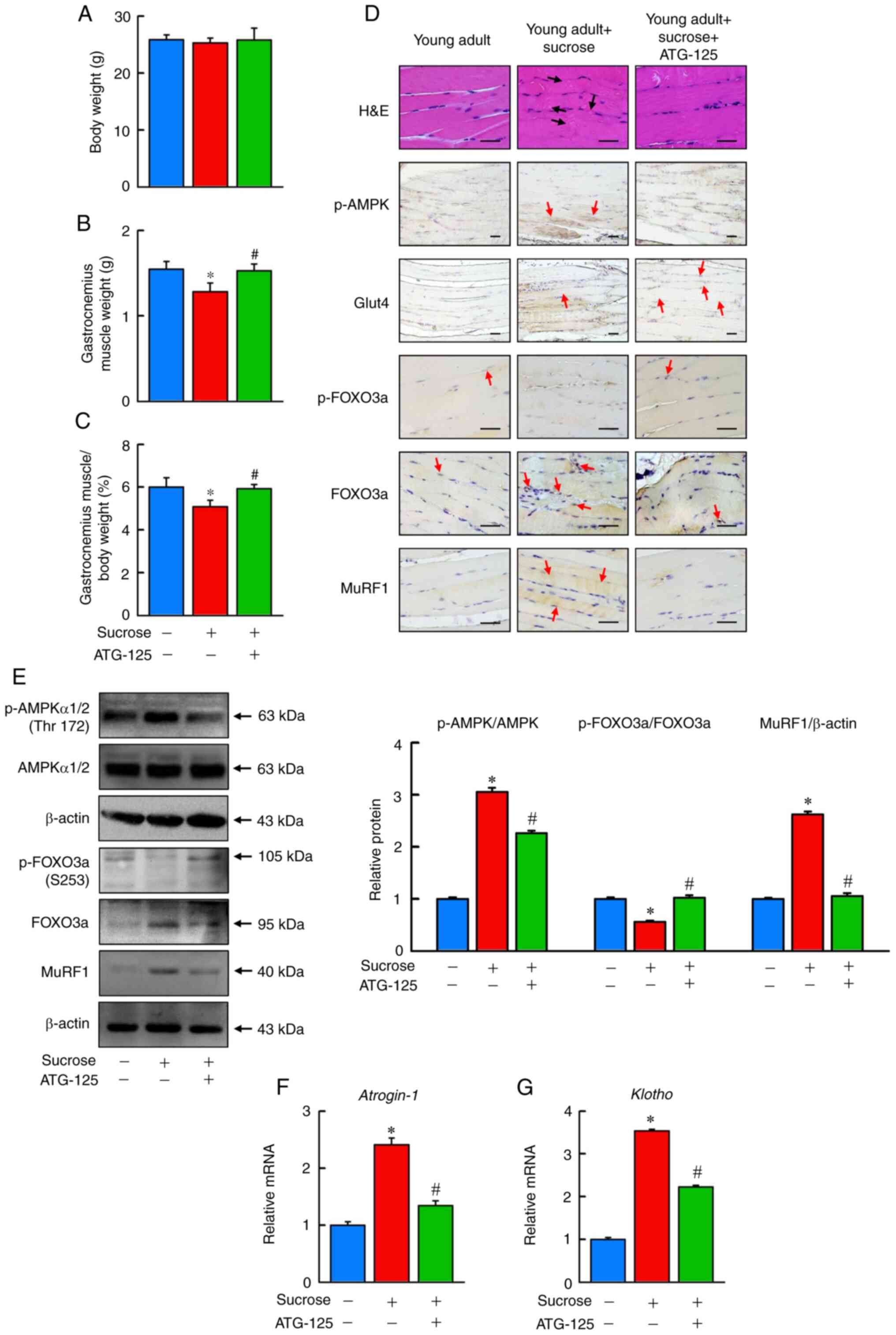 | Figure 6.ATG-125 improves sucrose-induced
muscle atrophy in mice. The (A) body weight, (B) muscle weight and
(C) gastrocnemius muscle/body weight ratio between the young adult
mice, sucrose-induced muscle atrophy mice and ATG-125 treated
sucrose-induced muscle atrophy mice. (D) Representative H&E
staining and immunohistochemical analysis of p-AMPK, Glut4,
p-FOXO3a, FOXO3a and MuRF1 in muscle sections of the young adult
mice, sucrose-induced muscle atrophy mice and ATG-125 treated
sucrose-induced muscle atrophy mice. Red arrows highlight the
positive staining. Scale bar, 100 µm. (E) p-AMPK, AMPK, p-FOXO3a,
FOXO3a and MuRF1 protein expression levels were determined by
western blotting. (F and G) Atrogin-1 and Klotho mRNA
expression levels were determined by reverse
transcription-quantitative PCR relative to Gapdh. Data are
presented as the mean ± standard error of the mean of five
independent experiments. *P<0.05 vs. Young adult mice;
#P<0.05 vs. Sucrose-induced muscle atrophy mice.
H&E, hematoxylin and eosin; p-, phosphorylated; AMPK,
AMP-activated protein kinase; FOXO3a, forkhead box O3a; Glut4,
glucose transporter type 4 insulin-responsive; MuRF1, E3
ubiquitin-protein ligase TRIM63. |
To further confirm whether sucrose-induced
gastrocnemius muscle weight loss was associated with protein
degradation, the activity of the AMPK/FOXO3a/MuRF1 signaling
pathway in gastrocnemius muscle was assessed. Immunohistochemical
and western blot analysis showed p-FOXO3a/OXO3a expression was
significantly reduced in the sucrose fed group compared with the
control, and increased in the ATG-125 treatment group. Furthermore,
high p-AMPK and MuRF1 protein levels, and high Atrogin−1 and
Klotho mRNA expression levels in the gastrocnemius muscle of
young adult mice fed sucrose compared with the young adult group
(Fig. 6D-G). ATG-125
significantly reduced AMPK/FOXO3a/MuRF1 signaling pathway activity
compared with the sucrose group, suggesting that ATG-125 improved
protein degradation through inhibiting AMPK/FOXO3a/MuRF1 signaling
in sucrose-induced muscle atrophy.
ATG-125 improves chronic inflammation
in sucrose-induced gastrocnemius muscle atrophy
To determine the dynamics of recovery from
sucrose-induced gastrocnemius muscle atrophy, the mechanisms of
chronic inflammation in sucrose-fed young adult mice treated with
ATG-125 were determined. Immunohistochemical analysis showed
HIF-1α, VEGF, TGFβRII, NF-κB, p-p53 and Ki67 protein levels, as
well as Nox2, Srebp-1, Cpt-1, Pparα, Ppar-γ, Aco, Tnfα, Il-6,
Il-1β and Crp mRNA expression levels were higher in the
sucrose-induced mice (Fig. 7A-E).
ATG-125 significantly decreased HIF-1α and NF-κB mediated
inflammation signaling compared with sucrose-challenged mice. Thus,
ATG-125 is an important regulator of chronic inflammation.
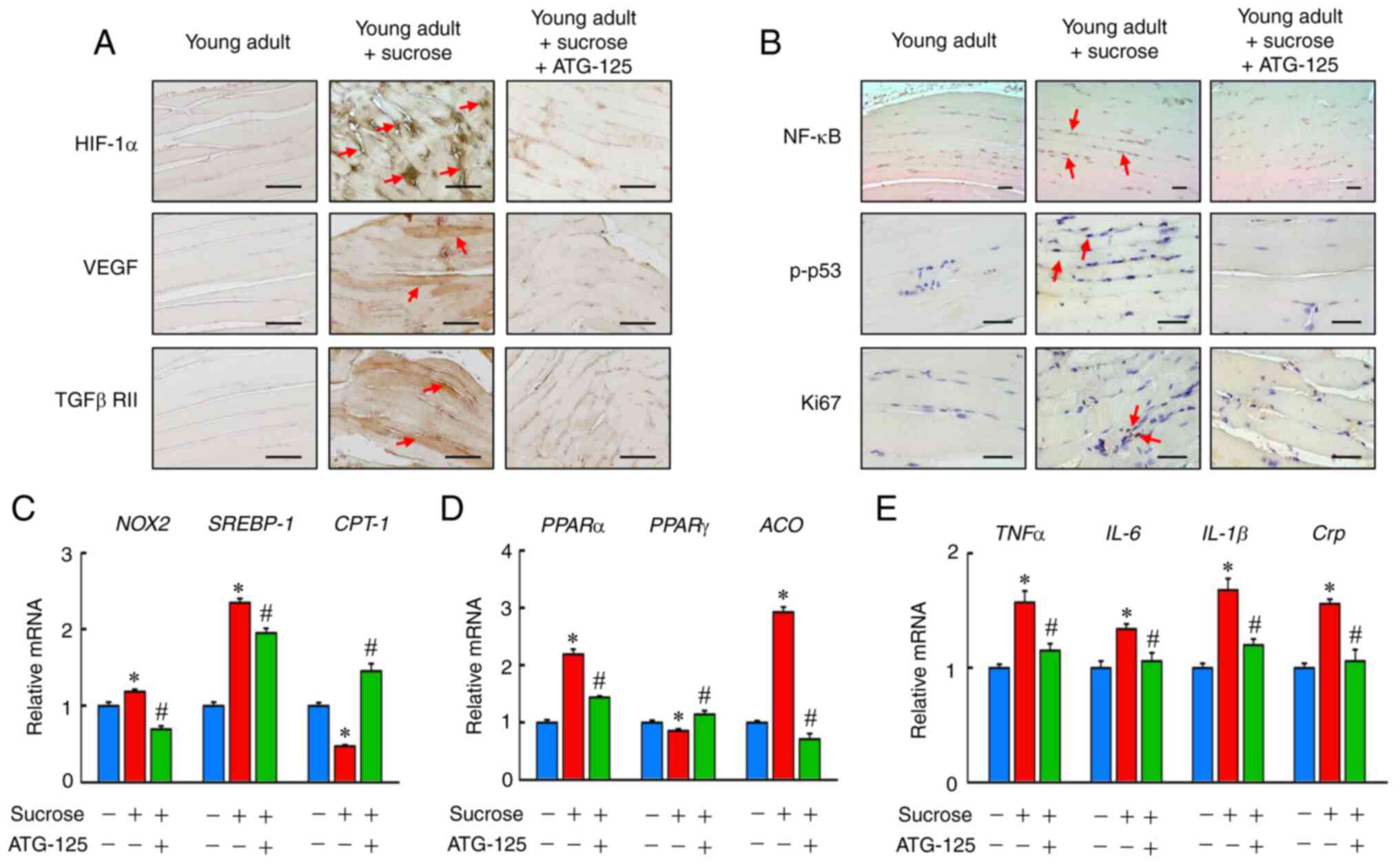 | Figure 7.ATG-125 improves sucrose-induced
inflammation in mice muscle atrophy. (A) Immunohistochemistry
analysis of HIF-1α, VEGF, TGFβRII, (B) NF-κB, p-p53 and Ki67
staining of the muscle sections. Red arrows highlight the positive
staining. Scale bar, 100 µm. (C) Nox2, Srebp-1, Cpt-1, (D)
Pparα, Pparγ, Aco, (E) Tnfα, Il-6, Il-1β and
Crp mRNA expression levels were determined by reverse
transcription-quantitative PCR. Data are presented as the mean ±
the standard error of the mean of five independent repeats.
*P<0.05 vs. Young adult mice; #P<0.05 vs.
Sucrose-induced muscle atrophy mice. HIF-1α, hypoxia inducible
factor-1α; VEGF, vascular endothelial growth factor; TGFβRII,
transforming growth factor β receptor type-II; NF-κB, nuclear
factor-κB; p-, phosphorylated; Nox2, NADPH oxidase 2;
Srebp-1, sterol regulatory element-binding protein 1;
Cpt−1, carnitine palmitoyltransferase 1; Ppar,
peroxisome proliferator-activated receptor; Aco, acyl-CoA
oxidase; Tnfα, tumor necrosis factor α; Il-6, interleukin-6;
Crp, C-reactive protein. |
ATG-125 improves sucrose-induced
mitochondrial dysfunction during muscle atrophy in young adult
mice
Next, the mechanistic underpinnings of the
sucrose-induced mitochondrial dysfunction observed in muscle tissue
of sucrose-fed mice under ATG-125 treatment were investigated. Mice
fed sucrose exhibited lower SIRT1, PGC1α, UCP1, UCP2 and UCP3
protein expression levels (Fig. 8A
and B), as well as reduced mRNA expression of Sirt1, Pgc1α,
Nrf1, Tfam and Ucp2 compared with the young adult group
(Fig. 8C and D). Moreover, PGC1α
was increased in ATG-125 treated young adult mice challenged with
sucrose as well as Pgc1α, Nrf1 and Tfam expression
levels. These results therefore indicated that the mitochondrial
biogenesis may be replenished in sucrose-induced mice.
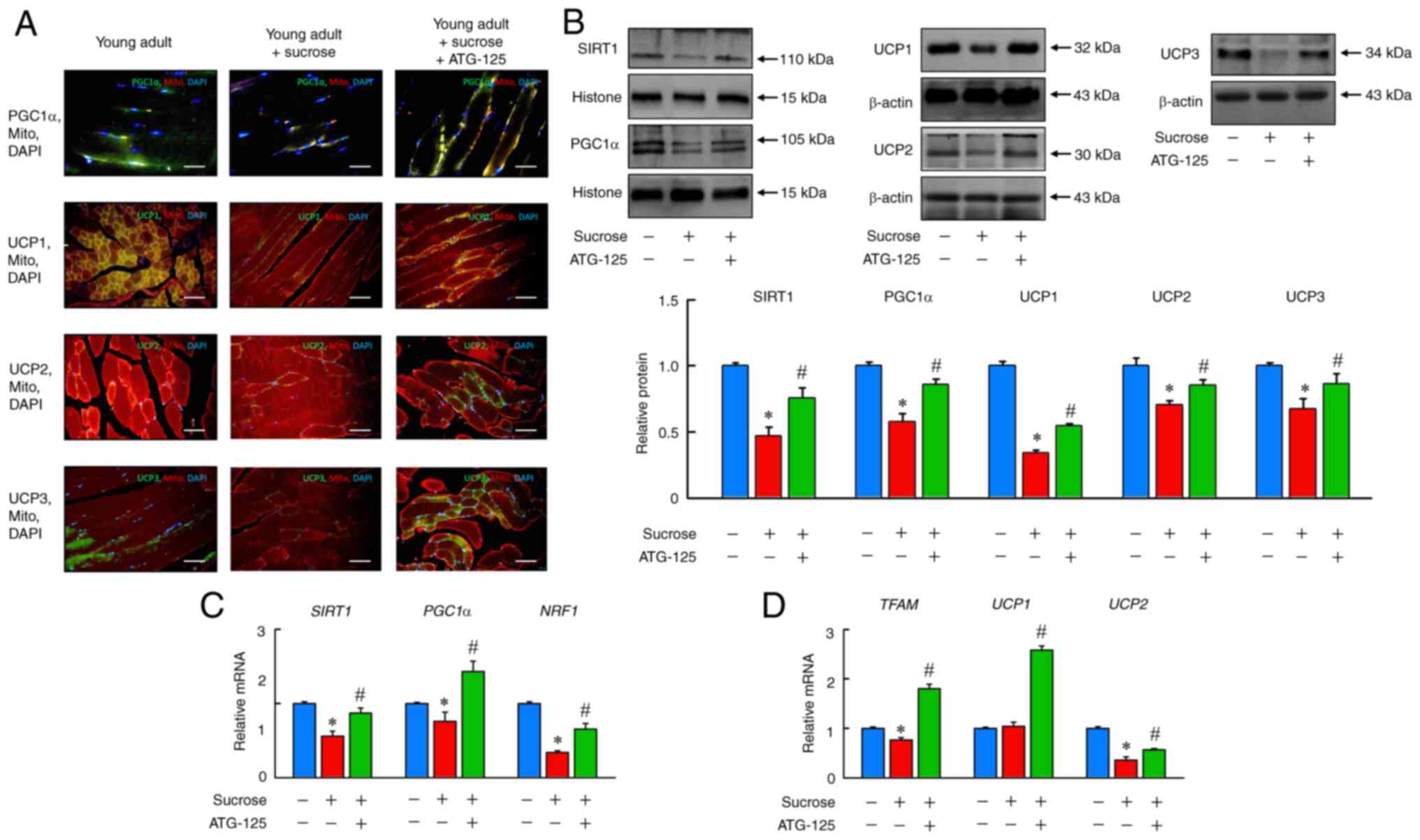 | Figure 8.ATG-125 improves sucrose-induced
mitochondrial dysfunction in mice muscle atrophy. (A)
Immunofluorescence analysis of PGC1α, UCP1, UCP2 and UCP3
expression, as well as Mito tracker staining for muscle sections.
Scale bar, 100 µm. (B) SIRT1, PGC1α, UCP1, UCP2 and UCP3 protein
expression levels were determined by western blotting. (C)
Sirt1, Pgc1α, Nrf1, (D) Tfam, Ucp1 and Ucp2
mRNA expression levels were determined by reverse
transcription-quantitative PCR. Data are presented as the mean ±
the standard error of the mean of five independent repeats.
*P<0.05 vs. Young adult mice; #P<0.05 vs.
Sucrose-induced muscle atrophy mice. SIRT1, sirtuin-1; PGC-1,
peroxisome proliferator-activated receptor γ coactivator 1-α;
Nrf1, nuclear respiratory factor 1; Tfam,
mitochondrial transcription factor A; Ucp1, uncoupling
protein 1. |
ATG-125 enhances IGF-AKT-mTOR
phosphorylation pathways during muscle atrophy in young adult
mice
To identify the role of ATG-125 on protein synthesis
via regulation of insulin and mTOR signaling in sucrose-induced
muscle atrophy, the activity of the IGF-AKT-mTOR phosphorylation
pathway in gastrocnemius muscle was assessed. p-IGF1R, p-IRS1,
p-PI3K, p-AKT, p-mTOR, p-S6K and p-4EBP1 protein levels and
Igf1r, Irs1, Pi3k, Akt, mtor, S6k and 4ebp1 mRNA
expression levels were significantly reduced in the sucrose-induced
young adult mice compared with the ATG-125 treated mice, and
ATG-125 increased the phosphorylation of members of the
IGF-AKT-mTOR pathway compared with the sucrose-induced mice,
suggesting increased activity of this pathway in the gastrocnemius
muscle (Fig. 9A and B). These
data suggested that ATG-125 promoted protein synthesis by enhancing
IGF-AKT-mTOR pathway activity in sucrose-induced muscle
atrophy.
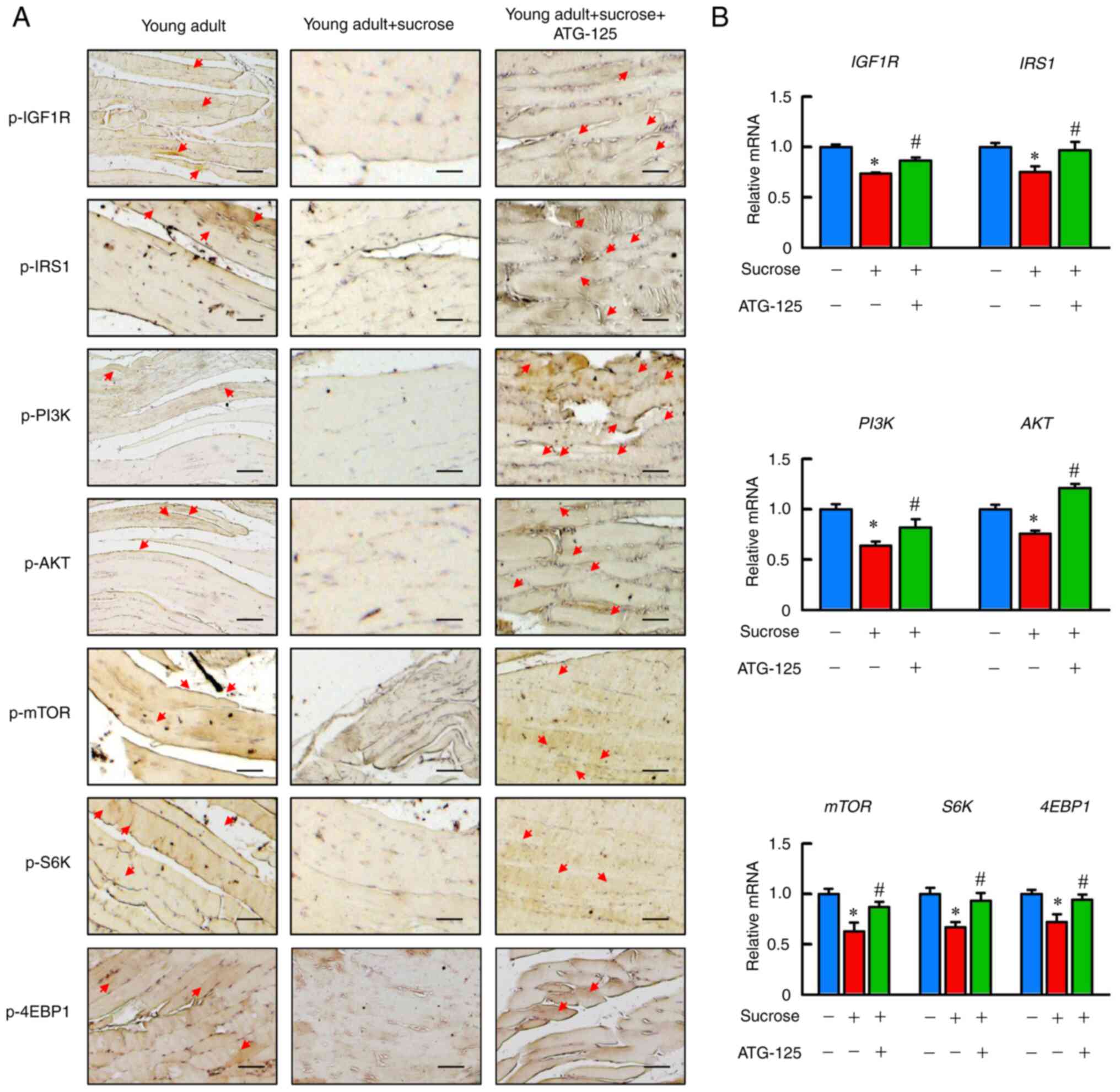 | Figure 9.ATG-125 reverses the impaired
sucrose-induced insulin signaling in muscle atrophy in young adult
mice. (A) Immunohistochemical analysis of p-IGF1R, p-IRS1, p-PI3K,
p-AKT, p-mTOR, p-S6K and p-4EBP1 staining in muscle sections are
shown. Red arrows highlight the positive staining. Scale bar, 100
µm. (B) Igf1r, Irs1, Pi3k, Akt, mtor, S6k and 4ebp1
mRNA expression levels were determined by reverse
transcription-quantitative PCR. Data are presented as the mean ±
the standard error of the mean of five independent repeats.
*P<0.05 vs. Young adult mice; #P<0.05 vs.
Sucrose-induced muscle atrophy mice. p-, phosphorylated; IGF1R,
insulin-like growth factor 1 receptor; IRS1, insulin receptor
substrate 1; PI3K, phospho-inositide 3-kinase; AKT, protein kinase
B; mTOR, mechanistic target of rapamycin; S6K, ribosomal protein S6
kinase; 4EBP1, eukaryotic translation initiation factor 4E-binding
protein 1. |
Discussion
Several studies have analyzed the effects of sugar
additives on metabolic syndromes in humans as well as in animal
models (92–94). The hypothesis of the present study
was that as sugar additives can induce inflammation, oxidative
stress and IR, this could in turn accelerate muscle atrophy.
Sugar-induced metabolic disturbances are reduced by
polyphenols/antioxidants, such as resveratrol (92), quercetin (93) and rutin (94), supplementation of which restores
muscle protein synthesis sensitivity in older rats. Thus, it was
proposed that ATG-125 may also be efficient in counteracting the
hypothetical accelerating effect sugar additives have on muscle
atrophy; thus, the progression of muscle weight, markers of
inflammation and IGF/AKT/mTOR pathways of muscle protein synthesis
were assessed.
A comprehensive understanding of the effects of
ATG-125 on sucrose-induced young adult muscle atrophy was performed
in the present study, thus the network of genes involved in this
modulation was determined to illustrate the potential advantage of
ATG-125, with the aim of identifying the core candidate genes and
pathways involved using a sucrose-induced muscle atrophy mouse
model treated with ATG-125. The DEGs were filtered using R and GO
and KEGG pathway analysis were performed. First, DEGs (18
upregulated and 14 downregulated) were identified, followed by GO
analysis, which revealed that the majority of the DEGs were
significantly enriched in ‘negative regulation of proteolysis’,
‘negative regulation of peptidase activity’, ‘cellular amino acid
metabolic process’, ‘small molecule catabolic process’, ‘blood
coagulation’ and ‘fibrinolysis’ in the BP category. Signaling
pathway analysis indicated that the DEGs shared common pathways in
‘biosynthesis of amino acids’, ‘fatty acid elongation’,
‘cholesterol metabolism’, and ‘cell cycle’.
The presence of Serpina genes as a DEG facilitates
the identification of potential underlying mechanisms involved in
disease occurrence/progression. In the present study,
Serpina3 was downregulated in the mouse model of muscle
atrophy treated with ATG-125 in DO and KEGG pathway enrichment
analyses. The DO results showed that Serpina3 was
significantly enriched in vascular diseases, blood coagulation
disorders, deep vein thrombosis, ischemic stroke and age-related
macular degeneration pathways. Therefore, it was hypothesized that
SERPINA3 may play an important role in high sucrose intake via
regulation of signaling pathways involved in muscle oxidative
stress and inflammation. Indeed, SERPINA3 is involved in several
complex vascular diseases, where it is always involved in
inflammation, activating thrombin biological processes in various
biological networks (95).
SERPINA3, also known as α1-antichymotrypsin, is an acute phase
protein, the levels of which increase in acute and chronic
inflammation (95).
Increasing evidence points to extensive crosstalk
between pro-inflammatory cytokines and other mediators that lead to
activation of the coagulation system and downregulation of
important physiological anticoagulant pathways (96–98), and coagulation also considerably
affects inflammatory activity (98). The expression levels of SERPINA3
in thick choroids was found to be higher in the eyes of patients
with early age-related macular degeneration (99), in which SERPINA3 expression may
increase oxidative stress and inflammation, a leading cause of
blindness, highlighting its potential as a biomarker of macular
degeneration. In the present study, Serpina3 was one of the
top 30 DEGs and was significantly downregulated in the ATG-125
treated mice. Therefore, it was hypothesized that ATG-125 may play
a protective role in the aforementioned vascular-associated
diseases when challenged with high sucrose levels.
In sucrose challenged mice, associations between the
abundance of mRNA transcripts associated with mitochondrial
biogenesis, such as Nmrk2, Ucp3, Slc7a10 (a cationic amino
acid transporter) and pyruvate dehydrogenases (Pdk4 and
Pdp2) pathways were observed, consistent with the potential
protective role of ATG-125 in muscles from metabolic and functional
degeneration. Conversely, in the sucrose-induced gastrocnemius
muscle atrophy mouse model, it was observed that ATG-125
significantly downregulated serine (or cysteine) peptidase
inhibitor (Serpina 3k, 1b, 1d and 1e) levels,
concurrently, all of which are closely associated with specific
vascular functions in the DO list, including ‘deep vein
thrombosis’, ‘blood coagulation disorders’, ‘vascular diseases’,
‘ischemic stroke’ and ‘age-related macular degeneration’ based on
DO analysis. In addition, ATG-125 reduced the expression of
apolipoproteins A-I, A-II and C-III, the major genes involved in
high density lipoprotein particle crosstalk with lipid-induced
cardiovascular disease risk factors and FFAs, together stimulating
inflammation (100–102). Furthermore, the gastrocnemius
muscle transcriptomes in the mouse muscular dystrophy model were
enriched for various amino acid metabolic processes, lipoprotein
particles and protein-lipid complex metabolism-related genes, a
number of which were modulated by ATG-125, and were involved in
proteolytic events, including inflammation.
Replenishing mitochondrial biogenesis with daily
ATG-125 intervention increased the activity of improved muscle
IGF-AKT-mTOR, as determined by the increased phosphorylation of its
members, in sucrose-induced young adult mice, and reversed the
NF-κB-HIF-1α-TGFβRII inflammatory cascade in the model of muscle
atrophy. The effects of mitochondrial energy repletion in the
sucrose-induced mouse model relied on the improvement in
mitochondrial function and on the reduction in general proteolysis,
inflammation and degradation.
In summary, the results of the present study
suggested that the administration of ATG-125 may benefit mice with
muscular dystrophies or other neuromuscular degenerative conditions
characterized by the aforementioned gene expression signatures.
Mitochondria play a crucial role in the maintenance of ionic
balance and other organelles as a source of energy. PGC-1α plays a
crucial role in regulating mitochondrial biogenesis and function,
particularly in skeletal muscle (103). SIRT1 modulates PGC-1α expression
and activity (104), which
regulates mitochondrial biogenesis and function through the
induction and activation of several downstream nuclear
transcription factors (including TFAM and NRF1) (104,105). In the present study, the results
showed that ATG-125 significantly increased the PGC-1α, SIRT1
protein and Nrf1 and Tfam mRNA expression levels and
the mitochondrial UCP1, 2 and 3 levels, suggesting that ATG-125
played a crucial role in regulating mitochondrial biogenesis and
function. Moreover, studies have demonstrated that enhanced
mitochondrial biogenesis and function may play a primary role in
elevated type I muscle fiber formation and may be a novel mediator
of skeletal muscle fiber type (106,107). Although the present study did
not determine whether type I muscle fibers were more abundant in
the ATG-125 treated mice, it was shown that ATG-125 downregulated
the mRNA expression levels of type I muscle fiber-related genes
(Tnni1, Tnnc1 and Tnnt1) in the sucrose-challenged
gastrocnemius muscle. The transcriptomic data suggested that, at
least in part, ATG-125 may promote the formation of type I muscle
fibers in mice muscles, which is dependent on the improvement of
mitochondrial biogenesis.
Indeed, PGC-1α plays a crucial role in the control
of mitochondrial biogenesis, which is intricately involved in core
muscle physiology and exercise physiology through its interaction
with FOXO and NF-kB and its regulation by the SIRT pathway and
apoptotic cascades (108–110).
The activation of FOXO3 enhances atrogin-1 and MuRF-1
transcriptional activity (111,112). Furthermore, NF-κB activation
promotes pro-inflammatory cytokine and myokine production, such as
IL-1β, IL-6, TNF-α and monocyte chemoattractant protein (113,114). Following ATG-125 treatment,
there was a reduction in NF-κB expression, a known deacetylation
target of SIRT1, suggesting that a decrease in
mitochondria-dependent SIRT1 activity was involved in the improved
inflammatory state of the skeletal muscle in the sucrose-challenged
mice. In the muscle, myofibrillar proteins serve as the major amino
acid reservoir in the organism, therefore, protein degradation
serves a physiological role distinct from proteolysis in other
tissues. While proteasomes degrade myofibrillar and most soluble
proteins (115), organelles
(especially the mitochondria) are degraded primarily in lysosomes
(116). Although atrogin-1 and
MuRF1 proteolytic pathways are activated by FOXO3, and this results
in reduced strength due to loss of myofibrillar components, as well
as in reduced endurance due to loss of the mitochondria, it
presumably ensures that the loss of different cell components is
coordinated upon fasting or disuse, allowing the muscle to maintain
a relatively normal composition. This difference may explain why
proteasome-dependent proteolysis accounts for only a small fraction
of the FOXO3-induced proteolysis in myotubes, whereas this system
accounts for the majority of the accelerated proteolysis in normal
and atrophying adult muscles (117). The analysis performed in the
present study, combining unbiased transcriptomics and biomedical
analysis approaches, identified an important mechanism for
protecting the muscle from wasting during excessive sucrose intake.
IGF1-AKT-mTOR phosphorylation was strongly downregulated by
sucrose, which led to loss of muscle mass by concurrently
activating proteolysis and disrupting homeostatic muscle growth.
ATG-125 may exert its protective effects by regulating the balance
between distinct growth-associated mTOR complexes, which strongly
inhibit IGF1-AKT-mTOR phosphorylation, while supporting
mitochondrial biogenesis. Various physiological or pathological
hypertrophic growth stimuli lead to activation of AKT by
phosphorylation, which causes inactivation of FOXO factors via
phosphorylation and nuclear exclusion, thus limiting the expression
of atrophy-related genes (118).
The present study demonstrated that sucrose inactivates
phosphorylation of the PI3K-AKT pathway, thus IGF-1-stimulated
protein synthesis is inhibited, resulting in degradation of
myofibrillar proteins and expression of Atrogen-1 and MuRF1.
The ATG-125-induced inhibition of proteolysis appears to contribute
to muscle growth via inactivation of FOXO3 through AKT
phosphorylation, thus maintaining proteostasis and organelle
quality for homeostatic preservation of muscle mass.
Loss of muscle mass and force significantly affects
the health, quality of life and even survival of an individual.
Given its beneficial effects on reducing muscle wasting, ATG-125
should be regarded as a novel regulator of mammalian muscle growth
with potentially broad applications in the treatment of common
catabolic conditions. However, the present study has certain
limitations: i) By mixing all the phytochemicals together, the
focused insights in single compound characteristics on muscle
atrophy were not assessed. ii) Bioinformatics was only performed in
young adult mice and the mechanisms identified may not apply to the
muscle atrophy observed in aging mice or type 2 diabetic animals.
iii) ATG-125 is a phytochemicals-rich herbal formula that contains
multiple antimicrobial and antioxidant properties herbs. In
addition to their diverse bioactive properties in animal models,
they have potential effects in the maintenance of human health. Due
to this relevance, highly-specific qualitative and quantitative
methods of analysis are necessary to accurately analyze bioactive
substance in different matrices derived from ATG-125. In future
studies, a sensitive and specific multiple reaction monitoring or
parallel reaction monitoring analysis method is required to detect
the verification of screened proteins that may be involved in
relation to muscle atrophy signaling. iv) This study did not
observe the dose-dependent effects of ATG-125 at high or low doses,
which leads to unclear estimates of the acceptable potency of
ATG-125. In other words, the current results just raised a
‘reference dose’ or ‘acceptable concentration’ of ATG-125. The lack
of dose response and positive controls are major limitations of
this study. Additional databases should be used for validation of
the effects of ATG-125 in future studies.
In conclusion, the results of the present study
showed that ATG-125 may exert protective effects against
sucrose-induced muscle atrophy via regulation of the AKT/FOXO3
pathways, which regulates proteolysis, as well as by improving
mitochondrial dysfunction.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by funding from Chang Gung
University (grant nos. SCRPD1J0141, SCRPD1J0221 and
SCRPD1K0651).
Availability of data and materials
The datasets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request. Sequence Read Archive (SRA; http://www.ncbi.nlm.nih.gov/sra) stores raw DNA
sequencing data and alignment information from high-throughput
sequencing platforms. BioSample (accession no. SAMN22871017)
records are indexed and searchable. The datasets generated and/or
analyzed during the current study are available in the BioProjects
(accession no. PRJNA777698) repository (https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA777698),
as well as with derived experimental data in NCBI's primary
archives, including SRA (accession no. SRR16767413-SRR16767421),
which all identify genes from muscle tissue samples.
Authors' contributions
CCY, HML, HHC and TYL designed the experiments.
CCY, HML, MCL, YLL and TYL performed the experiments. CCY and HML
analyzed the data. CCY, HML, HHC and TYL wrote the manuscript. All
authors have read and approved the final manuscript. WHC supervised
the study and interpreted the data. MCL and YLL confirm the
authenticity of all the raw data.
Ethics approval and consent to
participate
All animal experiments were performed in accordance
with the Guide for the Care and Use of Laboratory Animals and were
approved by the Chang Gung University Animal Care and Use Committee
(approval no. CGU108-111; Taoyuan, Taiwan).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lynch GS, Schertzer JD and Ryall JG:
Therapeutic approaches for muscle wasting disorders. Pharmacol
Ther. 113:461–487. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bonaldo P and Sandri M: Cellular and
molecular mechanisms of muscle atrophy. Dis Model Mech. 6:25–39.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ferrucci L and Guralnik JM: Inflammation,
hormones, and body composition at a crossroad. Am J Med.
115:501–502. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pedersen BK and Bruunsgaard H: Possible
beneficial role of exercise in modulating low-grade inflammation in
the elderly. Scand J Med Sci Sports. 13:56–62. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Franceschi C, Bonafè M, Valensin S,
Olivieri F, De Luca M, Ottaviani E and De Benedictis G:
Inflamm-aging. An evolutionary perspective on immunosenescence. Ann
N Y Acad Sci. 908:244–254. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ali S and Garcia JM: Sarcopenia, cachexia
and aging: Diagnosis, mechanisms and therapeutic options-a
mini-review. Gerontology. 60:294–305. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Costamagna D, Costelli P, Sampaolesi M and
Penna F: Role of Inflammation in muscle homeostasis and myogenesis.
Mediators Inflamm. 2015:8051722015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Milan G, Romanello V, Pescatore F, Armani
A, Paik JH, Frasson L, Seydel A, Zhao J, Abraham R, Goldberg AL, et
al: Regulation of autophagy and the ubiquitin-proteasome system by
the FoxO transcriptional network during muscle atrophy. Nat Commun.
6:66702015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gregor MF and Hotamisligil GS:
Inflammatory mechanisms in obesity. Annu Rev Immunol. 29:415–445.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Glass CK and Olefsky JM: Inflammation and
lipid signaling in the etiology of insulin resistance. Cell Metab.
15:635–645. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Olefsky JM and Glass CK: Macrophages,
inflammation, and insulin resistance. Annu Rev Physiol. 72:219–246.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Perry BD, Caldow MK, Brennan-Speranza TC,
Sbaraglia M, Jerums G, Garnham A, Wong C, Levinger P, Asrar Ul Haq
M, Hare DL, et al: Muscle atrophy in patients with Type 2 Diabetes
Mellitus: Roles of inflammatory pathways, physical activity and
exercise. Exerc Immunol Rev. 22:94–109. 2016.PubMed/NCBI
|
|
13
|
Maliszewska K, Adamska-Patruno E and
Krętowski A: The interplay between muscle mass decline, obesity,
and type 2 diabetes. Pol Arch Intern Med. 129:809–816.
2019.PubMed/NCBI
|
|
14
|
Monier S, Le Cam A and Le Marchand-Brustel
Y: Insulin and insulin-like growth factor I. Effects on protein
synthesis in isolated muscles from lean and goldthioglucose-obese
mice. Diabetes. 32:392–397. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rommel C, Bodine SC, Clarke BA, Rossman R,
Nunez L, Stitt TN, Yancopoulos GD and Glass DJ: Mediation of
IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and
PI(3)K/Akt/GSK3 pathways. Nat Cell Biol. 3:1009–1013. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sacheck JM, Ohtsuka A, McLary SC and
Goldberg AL: IGF-I stimulates muscle growth by suppressing protein
breakdown and expression of atrophy-related ubiquitin ligases,
atrogin-1 and MuRF1. Am J Physiol Endocrinol Metab. 287:E591–E601.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
de Alvaro C, Teruel T, Hernandez R and
Lorenzo M: Tumor necrosis factor alpha produces insulin resistance
in skeletal muscle by activation of inhibitor kappaB kinase in a
p38 MAPK-dependent manner. J Biol Chem. 279:17070–17078. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dogra C, Changotra H, Wedhas N, Qin X,
Wergedal JE and Kumar A: TNF-related weak inducer of apoptosis
(TWEAK) is a potent skeletal muscle-wasting cytokine. FASEB J.
21:1857–1869. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hirosumi J, Tuncman G, Chang L, Görgün CZ,
Uysal KT, Maeda K, Karin M and Hotamisligil GS: A central role for
JNK in obesity and insulin resistance. Nature. 420:333–336. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peterson JM, Bakkar N and Guttridge DC:
NF-κB signaling in skeletal muscle health and disease. Curr Top Dev
Biol. 96:85–119. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Törrönen R, Kolehmainen M, Sarkkinen E,
Mykkänen H and Niskanen L: Postprandial glucose, insulin, and free
fatty acid responses to sucrose consumed with blackcurrants and
lingonberries in healthy women. Am J Clin Nutr. 96:527–533. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Abdullah A, Peeters A, de Courten M and
Stoelwinder J: The magnitude of association between overweight and
obesity and the risk of diabetes: A meta-analysis of prospective
cohort studies. Diabetes Res Clin Pract. 89:309–319. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Boden G: Obesity and free fatty acids.
Endocrinol Metab Clin North Am. 37:635–646, viii-ix. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
K S, Senthilkumar GP, Sankar P and Bobby
Z: Attenuation of oxidative stress, inflammation and insulin
resistance by allium sativum in fructose-fed male rats. J Clin
Diagn Res. 7:1860–1862. 2013.PubMed/NCBI
|
|
25
|
Hwang IS, Ho H, Hoffman BB and Reaven GM:
Fructose-induced insulin resistance and hypertension in rats.
Hypertension. 10:512–516. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gatineau E, Savary-Auzeloux I, Migné C,
Polakof S, Dardevet D and Mosoni L: Chronic intake of sucrose
accelerates sarcopenia in older male rats through alterations in
insulin sensitivity and muscle protein synthesis. J Nutr.
145:923–930. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shin MS, Lee J, Lee JW, Park SH, Lee IK,
Choi JA, Lee JS and Kang KS: Anti-Inflammatory effect of
Artemisia argyi on Ethanol-Induced gastric ulcer: Analytical
in vitro and in vivo studies for the identification of action
mechanism and active compounds. Plants (Basel). 10:3322021.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gao XH, Zhang SD, Wang LT, Yu L, Zhao XL,
Ni HY, Wang YQ, Wang JD, Shan CH and Fu YJ: Anti-Inflammatory
effects of neochlorogenic acid extract from mulberry leaf (Morus
alba L.) against LPS-Stimulated inflammatory response through
mediating the AMPK/Nrf2 signaling pathway in A549 Cells. Molecules.
25:13852020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hu PF, Sun FF and Qian J: Leonurine exerts
Anti-Catabolic and Anti-apoptotic effects via nuclear factor kappa
B (NF-κB) and mitogen-activated protein kinase (MAPK) signaling
pathways in chondrocytes. Med Sci Monit. 25:6271–6280. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cho SY, Kim HW, Lee MK, Kim HJ, Kim JB,
Choe JS, Lee YM and Jang HH: Antioxidant and Anti-Inflammatory
activities in relation to the flavonoids composition of pepper
(Capsicum annuum L.). Antioxidants (Basel). 9:9862020.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hernández-Ortega M, Ortiz-Moreno A,
Hernández-Navarro MD, Chamorro-Cevallos G, Dorantes-Alvarez L and
Necoechea-Mondragón H: Antioxidant, antinociceptive, and
anti-inflammatory effects of carotenoids extracted from dried
pepper (Capsicum annuum L.). J Biomed Biotechnol.
2012:5240192012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lai KH, Chen PJ, Chen CC, Yang SH,
El-Shazly M, Chang YC, Wu YH, Wu YH, Wang YH, Hsieh HL and Hwang
TL: Lophatherum gracile Brongn. attenuates neutrophilic
inflammation through inhibition of JNK and calcium. J
Ethnopharmacol. 264:1132242021. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ti H, Mai Z, Wang Z, Zhang W, Xiao M, Yang
Z and Shaw P: Bisabolane-type sesquiterpenoids from Curcuma
longa L. exert anti-influenza and anti-inflammatory activities
through NF-κB/MAPK and RIG-1/STAT1/2 signaling pathways. Food
Funct. 12:6697–6711. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Memarzia A, Khazdair MR, Behrouz S,
Gholamnezhad Z, Jafarnezhad M, Saadat S and Boskabady MH:
Experimental and clinical reports on anti-inflammatory,
antioxidant, and immunomodulatory effects of Curcuma longa
and curcumin, an updated and comprehensive review. Biofactors.
47:311–350. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yue SJ, Qin YF, Kang A, Tao HJ, Zhou GS,
Chen YY, Jiang JQ, Tang YP and Duan JA: Total Flavonoids of
Glycyrrhiza uralensis Alleviates irinotecan-induced colitis
via modification of gut microbiota and fecal metabolism. Front
Immunol. 12:6283582021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li M, Chai X, Wang L, Yang J and Wang Y:
Study of the variation of phenolic acid and flavonoid content from
fresh artemisiae argyi folium to moxa wool. Molecules. 24:46032019.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhao L, Wang D, Liu J, Yu X, Wang R, Wei
Y, Wen C and Ouyang Z: Transcriptomic analysis of key genes
involved in chlorogenic acid biosynthetic pathway and
characterization of MaHCT from Morus alba L. Protein Expr
Purif. 156:25–35. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chang Y, Zhang D, Yang G, Zheng Y and Guo
L: Screening of Anti-Lipase components of Artemisia argyi
leaves based on Spectrum-Effect relationships and HPLC-MS/MS. Front
Pharmacol. 12:6753962021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wen YQ, Gong LY, Wang L, Zhao N, Sun Q,
Kamara MO, Ma HY and Meng FH: Comparative pharmacokinetics study of
leonurine and stachydrine in normal rats and rats with
Cold-Stagnation and Blood-Stasis primary dysmenorrhoea after the
administration of Leonurus japonicus houtt electuary. J Sep
Sci. 42:1725–1732. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dan-Dan L, Hua-Sheng P, Yuan Z, Xian-Zhang
H, Zhi-Lai Z, Da-Hui L, Li-Ping K and Lu-Qi H: Comparison of
chemical components between Artemisia stolonifera and Artemisia
argyi using UPLC-Q-TOF-MS. Zhongguo Zhong Yao Za Zhi.
45:4057–4064. 2020.(In Chinese). PubMed/NCBI
|
|
41
|
Wang ZL, Gao HM, Wang S, Zhang M, Chen K,
Zhang YQ, Wang HD, Han BY, Xu LL, Song TQ, et al: Dissection of the
general two-step di-C-glycosylation pathway for the biosynthesis of
(iso)schaftosides in higher plants. Proc Natl Acad Sci USA.
117:30816–30823. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Suntornsuk L, Kasemsook S and Wongyai S:
Quantitative analysis of aglycone quercetin in mulberry leaves
(Morus alba L.) by capillary zone electrophoresis.
Electrophoresis. 24:1236–1241. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim GD, Lee YS, Cho JY, Lee YH, Choi KJ,
Lee Y, Han TH, Lee SH, Park KH and Moon JH: Comparison of the
content of bioactive substances and the inhibitory effects against
rat plasma oxidation of conventional and organic hot peppers
(Capsicum annuum L.). J Agric Food Chem. 58:12300–12306.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li S, Zhou S, Yang W and Meng D:
Gastro-protective effect of edible plant Artemisia argyi in
ethanol-induced rats via normalizing inflammatory responses and
oxidative stress. J Ethnopharmacol. 214:207–217. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chuang TY, Lien CY, Tsai YC, Lin KF, Hsu
CH, Wu WJ, Su LY, Lu CW and Wu CH: Oral treatment with the Chinese
herbal supplements B307 enhances muscle endurance of ICR mice after
exhaustive swimming via suppressing fatigue, oxidative stress, and
inflammation. Food Sci Nutr. 8:3682–3691. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Lien CY, Lu CW, Lin YH, Wu WJ, Hsu CH,
Chuang TY, Lin KF, Chuang WC, Lee MC and Wu CH: Chinese herbal
medicine, guilu erxian glue, as alternative medicine for adverse
side effects of chemotherapy in Doxorubicin-Treated cell and mouse
models. Evid Based Complement Alternat Med. 2021:55489682021.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kato T, Iizuka K, Takao K, Horikawa Y,
Kitamura T and Takeda J: ChREBP-Knockout mice show sucrose
intolerance and fructose malabsorption. Nutrients. 10:3402018.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Burke SJ, Batdorf HM, Martin TM, Burk DH,
Noland RC, Cooley CR, Karlstad MD, Johnson WD and Collier JJ:
Liquid sucrose consumption promotes obesity and impairs glucose
tolerance without altering circulating insulin levels. Obesity
(Silver Spring). 26:1188–1196. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chen YS, Liu HM and Lee TY:
Ursodeoxycholic acid regulates hepatic energy homeostasis and white
adipose tissue macrophages polarization in Leptin-Deficiency obese
mice. Cells. 8:2532019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Moylan JS, Smith JD, Chambers MA,
McLoughlin TJ and Reid MB: TNF induction of atrogin-1/MAFbx mRNA
depends on Foxo4 expression but not AKT-Foxo1/3 signaling. Am J
Physiol Cell Physiol. 295:C986–C993. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Qian Y, Guo X, Che L, Guan X, Wu B, Lu R,
Zhu M, Pang H, Yan Y, Ni Z and Gu L: Klotho reduces necroptosis by
targeting oxidative stress involved in renal Ischemic-Reperfusion
injury. Cell Physiol Biochem. 45:2268–2282. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Omede F, Zhang S, Johnson C, Daniel E,
Zhang Y, Fields TA, Boulanger J, Liu S, Ahmed I, Umar S, et al:
Dietary phosphate restriction attenuates polycystic kidney disease
in mice. Am J Physiol Renal Physiol. 318:F35–F42. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Murdoch CE, Alom-Ruiz SP, Wang M, Zhang M,
Walker S, Yu B, Brewer A and Shah AM: Role of endothelial Nox2
NADPH oxidase in angiotensin II-induced hypertension and vasomotor
dysfunction. Basic Res Cardiol. 106:527–538. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sag CM, Schnelle M, Zhang J, Murdoch CE,
Kossmann S, Protti A, Santos CXC, Sawyer G, Zhang X, Mongue-Din H,
et al: Distinct regulatory effects of myeloid cell and endothelial
cell NAPDH oxidase 2 on blood pressure. Circulation. 135:2163–2177.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yates Z, Tarling EJ, Langley-Evans SC and
Salter AM: Maternal undernutrition programmes atherosclerosis in
the ApoE*3-Leiden mouse. Br J Nutr. 101:1185–1194. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang CH, Liu HM, Chang ZY, Huang TH and
Lee TY: Losartan prevents hepatic steatosis and macrophage
polarization by inhibiting HIF-1α in a murine model of NAFLD. Int J
Mol Sci. 22:78412021. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Huang Y, Wang J, Qua G, Wang X, Yang L and
Zhong L: Lactobacillus acidophilus ATCC 4356 prevents
atherosclerosis via inhibition of intestinal cholesterol absorption
in apolipoprotein E-knockout mice. Appl Environ Microbiol.
80:7496–7504. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Jin YN, Hwang WY, Jo C and Johnson GV:
Metabolic state determines sensitivity to cellular stress in
Huntington disease: Normalization by activation of PPARγ. PLoS One.
7:e304062012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Shimano H, Yahagi N, Amemiya-Kudo M, Hasty
AH, Osuga J, Tamura Y, Shionoiri F, Iizuka Y, Ohashi K, Harada K,
et al: Sterol regulatory element-binding protein-1 as a key
transcription factor for nutritional induction of lipogenic enzyme
genes. J Biol Chem. 274:35832–35839. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sung YY, Yoon T, Jang S and Kim HK:
Forsythia suspensa suppresses house dust mite
Extract-Induced atopic dermatitis in NC/Nga Mice. PLoS One.
11:e01676872016. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Saba E, Lee YS, Yang WK, Lee YY, Kim M,
Woo SM, Kim K, Kwon YS, Kim TH, Kwak D, et al: Effects of a herbal
formulation, KGC3P, and its individual component, nepetin, on coal
fly dust-induced airway inflammation. Sci Rep. 10:140362020.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Kagari T, Doi H and Shimozato T: The
importance of IL-1 beta and TNF-alpha, and the noninvolvement of
IL-6, in the development of monoclonal antibody-induced arthritis.
J Immunol. 169:1459–1466. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Choi BH, Ahn IS, Kim YH, Park JW, Lee SY,
Hyun CK and Do MS: Berberine reduces the expression of adipogenic
enzymes and inflammatory molecules of 3T3-L1 adipocyte. Exp Mol
Med. 38:599–605. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yang H, Liao D, Tong L, Zhong L and Wu K:
MiR-373 exacerbates renal injury and fibrosis via
NF-κB/MatrixMetalloproteinase-9 signaling by targeting Sirtuin1.
Genomics. 111:786–792. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Shin H, Ma Y, Chanturiya T, Cao Q, Wang Y,
Kadegowda AKG, Jackson R, Rumore D, Xue B, Shi H, et al: Lipolysis
in brown adipocytes is not essential for Cold-Induced thermogenesis
in mice. Cell Metab. 26:764–777.e5. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Thornton SM, Krolopp JE and Abbott MJ:
IL-15 mediates mitochondrial activity through a PPAR
δ-Dependent-PPAR α-Independent mechanism in skeletal muscle cells.
PPAR Res. 2016:54658042016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Luan G, Li G, Ma X, Jin Y, Hu N, Li J,
Wang Z and Wang H: Dexamethasone-Induced mitochondrial dysfunction
and insulin Resistance-Study in 3T3-L1 adipocytes and mitochondria
isolated from mouse liver. Molecules. 24:19822019. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Dusaulcy R, Rancoule C, Grès S, Wanecq E,
Colom A, Guigné C, van Meeteren LA, Moolenaar WH, Valet P and
Saulnier-Blache JS: Adipose-specific disruption of autotaxin
enhances nutritional fattening and reduces plasma lysophosphatidic
acid. J Lipid Res. 52:1247–1255. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Bernardi S, Toffoli B, Tisato V, Bossi F,
Biffi S, Lorenzon A, Zauli G, Secchiero P and Fabris B: TRAIL
reduces impaired glucose tolerance and NAFLD in the high-fat diet
fed mouse. Clin Sci (Lond). 132:69–83. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Bermúdez-Muñoz JM, Celaya AM,
Hijazo-Pechero S, Wang J, Serrano M and Varela-Nieto I: G6PD
overexpression protects from oxidative stress and age-related
hearing loss. Aging Cell. 19:e132752020. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Wang Y, Sakata T, Elalieh HZ, Munson SJ,
Burghardt A, Majumdar S, Halloran BP and Bikle DD: Gender
differences in the response of CD-1 mouse bone to parathyroid
hormone: Potential role of IGF-I. J Endocrinol. 189:279–287. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Al-Dwairi A, Brown AR, Pabona JM, Van TH,
Hamdan H, Mercado CP, Quick CM, Wight PA, Simmen RC and Simmen FA:
Enhanced gastrointestinal expression of cytosolic malic enzyme
(ME1) induces intestinal and liver lipogenic gene expression and
intestinal cell proliferation in mice. PLoS One. 9:e1130582014.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhao L, Li C, Liu F, Zhao Y, Liu J, Hua Y,
Liu J, Huang J and Ge C: A blockade of PD-L1 produced antitumor and
antimetastatic effects in an orthotopic mouse pancreatic cancer
model via the PI3K/Akt/mTOR signaling pathway. Onco Targets Ther.
10:2115–2126. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Deng L, Huang L, Sun Y, Heath JM, Wu H and
Chen Y: Inhibition of FOXO1/3 promotes vascular calcification.
Arterioscler Thromb Vasc Biol. 35:175–183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Choi DH, Yang J and Kim YS: Rapamycin
suppresses postnatal muscle hypertrophy induced by
myostatin-inhibition accompanied by transcriptional suppression of
the Akt/mTOR pathway. Biochem Biophys Rep. 17:182–190.
2019.PubMed/NCBI
|
|
77
|
Roselló-Díez A, Madisen L, Bastide S, Zeng
H and Joyner AL: Cell-nonautonomous local and systemic responses to
cell arrest enable long-bone catch-up growth in developing mice.
PLoS Biol. 16:e20050862018. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Prieto C and Barrios D: RaNA-Seq:
Interactive RNA-Seq analysis from FASTQ files to functional
analysis. Bioinformatics. Nov 15–2019.(Epub ahead of print).
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Bolger AM, Lohse M and Usadel B:
Trimmomatic: A flexible trimmer for Illumina sequence data.
Bioinformatics. 30:2114–2120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Sahraeian SME, Mohiyuddin M, Sebra R,
Tilgner H, Afshar PT, Au KF, Bani Asadi N, Gerstein MB, Wong WH,
Snyder MP, et al: Gaining comprehensive biological insight into the
transcriptome by performing a broad-spectrum RNA-seq analysis. Nat
Commun. 8:592017. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wang L, Wang S and Li W: RSeQC: Quality
control of RNA-seq experiments. Bioinformatics. 28:2184–2185. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Okonechnikov K, Conesa A and
García-Alcalde F: Qualimap 2: Advanced multi-sample quality control
for high-throughput sequencing data. Bioinformatics. 32:292–294.
2046.PubMed/NCBI
|
|
83
|
Liao Y, Smyth GK and Shi W: FeatureCounts:
An efficient general purpose program for assigning sequence reads
to genomic features. Bioinformatics. 30:923–930. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Varet H, Brillet-Guéguen L, Coppée JY and
Dillies MA: SARTools: A DESeq2- and EdgeR-Based R pipeline for
comprehensive differential analysis of RNA-Seq data. PLoS One.
11:e01570222016. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Holm L and Sander C: Removing
near-neighbour redundancy from large protein sequence collections.
Bioinformatics. 14:423–429. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Stat Soc Ser C Appl Stat. 57:289–300.
1995.
|
|
87
|
Liu S, Cai Y, Changyong E, Sheng J and
Zhang X: Screening and validation of independent predictors of poor
survival in pancreatic cancer. Pathol Oncol Res. 27:16098682021.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Gene Ontology Consortium: The Gene
Ontology project in 2008. Nucleic Acids Res. 36:D440–D444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Zhu HM, Fei Q, Qian LX, Liu BL, He X and
Yin L: Identification of key pathways and genes in nasopharyngeal
carcinoma using bioinformatics analysis. Oncol Lett. 17:4683–4694.
2019.PubMed/NCBI
|
|
90
|
Deng YJ, Ren FH, Yuan WH, Zhang GZ, Wu ZL
and Xie QQ: GRB10 and E2F3 as diagnostic markers of osteoarthritis
and their correlation with immune infiltration. Diagnostics
(Basel). 10:1712020. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Wu T, Hu E, Xu S, Chen M, Guo P, Dai J,
Feng T, Zhou L, Tang W, Zhan L, et al: clusterProfiler 4.0: A
universal enrichment tool for interpreting omics data. Innovation
(N Y). 2:1001412021.PubMed/NCBI
|
|
92
|
Bagul PK, Middela H, Matapally S, Padiya
R, Bastia T, Madhusudana K, Reddy BR, Chakravarty S and Banerjee
SK: Attenuation of insulin resistance, metabolic syndrome and
hepatic oxidative stress by resveratrol in fructose-fed rats.
Pharmacol Res. 66:260–268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Phuwamongkolwiwat P, Suzuki T, Hira T and
Hara H: Fructooligosaccharide augments benefits of
quercetin-3-O-β-glucoside on insulin sensitivity and plasma total
cholesterol with promotion of flavonoid absorption in sucrose-fed
rats. Eur J Nutr. 53:457–468. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Marzani B, Balage M, Vénien A, Astruc T,
Papet I, Dardevet D and Mosoni L: Antioxidant supplementation
restores defective leucine stimulation of protein synthesis in
skeletal muscle from old rats. J Nutr. 138:2205–2211. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Slowik A, Borratynska A, Turaj W, Pera J,
Dziedzic T, Figlewicz DA, Betlej M, Krzyszkowski T, Czepko R and
Szczudlik A: Alpha1-antichymotrypsin gene (SERPINA3) A/T
polymorphism as a risk factor for aneurysmal subarachnoid
hemorrhage. Stroke. 36:737–740. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
O'Brien M: The reciprocal relationship
between inflammation and coagulation. Top Companion Anim Med.
27:46–52. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Levi M: The coagulant response in sepsis
and inflammation. Hamostaseologie. 30:10–12. 14–16. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Levi M and van der Poll T: Inflammation
and coagulation. Crit Care Med. 38 (Suppl 2):S26–S34. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Sohn EH, Khanna A, Tucker BA, Abràmoff MD,
Stone EM and Mullins RF: Structural and biochemical analyses of
choroidal thickness in human donor eyes. Invest Ophthalmol Vis Sci.
55:1352–1360. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Nazir S, Jankowski V, Bender G, Zewinger
S, Rye KA and van der Vorst EPC: Interaction between high-density
lipoproteins and inflammation: Function matters more than
concentration! Adv Drug Deliv Rev. 159:94–119. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Maïga SF, Kalopissis AD and Chabert M:
Apolipoprotein A-II is a key regulatory factor of HDL metabolism as
appears from studies with transgenic animals and clinical outcomes.
Biochimie. 96:56–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
da Silva IT, de Almeida-Pititto B and
Ferreira SRG: Reassessing lipid metabolism and its potentialities
in the prediction of cardiovascular risk. Arch Endocrinol Metab.
59:171–180. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Lehman JJ, Barger PM, Kovacs A, Saffitz
JE, Medeiros DM and Kelly DP: Peroxisome proliferator-activated
receptor gamma coactivator-1 promotes cardiac mitochondrial
biogenesis. J Clin Invest. 106:847–856. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Price NL, Gomes AP, Ling AJ, Duarte FV,
Martin-Montalvo A, North BJ, Agarwal B, Ye L, Ramadori G, Teodoro
JS, et al: SIRT1 is required for AMPK activation and the beneficial
effects of resveratrol on mitochondrial function. Cell Metab.
15:675–690. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Vazquez F, Lim JH, Chim H, Bhalla K,
Girnun G, Pierce K, Clish CB, Granter SR, Widlund HR, Spiegelman BM
and Puigserver P: PGC1α expression defines a subset of human
melanoma tumors with increased mitochondrial capacity and
resistance to oxidative stress. Cancer Cell. 23:287–301. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Zhang L, Zhou Y, Wu W, Hou L, Chen H, Zuo
B, Xiong Y and Yang J: Skeletal muscle-specific overexpression of
PGC-1α induces Fiber-type conversion through enhanced mitochondrial
respiration and fatty acid oxidation in mice and pigs. Int J Biol
Sci. 13:1152–1162. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Lin J, Wu H, Tarr PT, Zhang CY, Wu Z, Boss
O, Michael LF, Puigserver P, Isotani E, Olson EN, et al:
Transcriptional co-activator PGC-1 alpha drives the formation of
slow-twitch muscle fibres. Nature. 418:797–801. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Puigserver P, Wu Z, Park CW, Graves R,
Wright M and Spiegelman BM: A cold-inducible coactivator of nuclear
receptors linked to adaptive thermogenesis. Cell. 92:829–839. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Eisele PS, Salatino S, Sobek J, Hottiger
MO and Handschin C: The peroxisome proliferator-activated receptor
γ coactivator 1α/β (PGC-1) coactivators repress the transcriptional
activity of NF-κB in skeletal muscle cells. J Biol Chem.
288:2246–2260. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Lira VA, Benton CR, Yan Z and Bonen A:
PGC-1alpha regulation by exercise training and its influences on
muscle function and insulin sensitivity. Am J Physiol Endocrinol
Metab. 299:E145–E161. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Sandri M, Sandri C, Gilbert A, Skurk C,
Calabria E, Picard A, Walsh K, Schiaffino S, Lecker SH and Goldberg
AL: Foxo transcription factors induce the atrophy-related ubiquitin
ligase atrogin-1 and cause skeletal muscle atrophy. Cell.
117:399–412. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Stitt TN, Drujan D, Clarke BA, Panaro F,
Timofeyva Y, Kline WO, Gonzalez M, Yancopoulos GD and Glass DJ: The
IGF-1/PI3K/Akt pathway prevents expression of muscle
atrophy-induced ubiquitin ligases by inhibiting FOXO transcription
factors. Mol Cell. 14:395–403. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Guttridge DC, Mayo MW, Madrid LV, Wang CY
and Baldwin AS Jr: NF-kappaB-induced loss of MyoD messenger RNA:
possible role in muscle decay and cachexia. Science. 289:2363–2366.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Kang C, Shin WS, Yeo D, Lim W, Zhang T and
Ji LL: Anti-inflammatory effect of avenanthramides via NF-κB
pathways in C2C12 skeletal muscle cells. Free Radic Biol Med.
117:30–36. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Solomon V and Goldberg AL: Importance of
the ATP-ubiquitin-proteasome pathway in the degradation of soluble
and myofibrillar proteins in rabbit muscle extracts. J Biol Chem.
271:26690–26697. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Scott SV and Klionsky DJ: Delivery of
proteins and organelles to the vacuole from the cytoplasm. Curr
Opin Cell Biol. 10:523–529. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Lecker SH, Goldberg AL and Mitch WE:
Protein degradation by the ubiquitin-proteasome pathway in normal
and disease states. J Am Soc Nephrol. 17:1807–1819. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Papanicolaou KN, Izumiya Y and Walsh K:
Forkhead transcription factors and cardiovascular biology. Circ
Res. 102:16–31. 2008. View Article : Google Scholar : PubMed/NCBI
|