Introduction
Osteoporosis is a debilitating disease that
predominantly affects elderly individuals worldwide (1). At present, >200 million patients
suffer from osteoporosis worldwide (2). According to research statistics,
>1.6 million fractures are caused by osteoporosis (3,4).
For women aged >50 years, the risk of suffering from fractures
due to osteoporosis is 40–50% (5). Osteoporosis is a systemic skeletal
disease that is characterized by a decrease in mass and
deterioration of the bone microstructure (6). The consequences brought on by
osteoporosis are bone fragility and susceptibility to fractures,
and this can cause brittle fractures (7). At the cellular level, the
pathogenesis of osteoporosis is primarily related to the disruption
of bone remodeling (8). Bone
remodeling primarily depends on osteoblast osteogenesis and
osteoclast bone absorption, so as to maintain a certain balance
(9). When a disease or aberrant
inflammation disrupt this balance, osteoporosis may occur (10).
In bone tissues, osteoblasts originate from
mesenchymal stem cells (MSCs), whereas osteoclasts are derived from
hematopoietic stem cells (11).
The differentiation of osteoblasts is achieved through the initial
organic phase, which is production of collagen matrix, and the
final inorganic phase, which is mineralization by hydroxyapatite
crystals on the collagen scaffold (12). After the completion of matrix
secretion and mineralization, the osteoblasts, which are embedded
in the matrix, are known as bone cells, whereas the osteoblasts
remaining on the bone surface turn into flattened lining cells or
gradually undergo apoptosis. Importantly, bone marrow MSCs (BMSCs)
are pluripotent. Apart from osteoblasts, BMSCs can differentiate
into several different cell types, including adipocytes, myocytes,
chondrocytes, endothelial cells and vascular smooth muscle cells
(13). Amongst these cell
lineages, adipogenic differentiation is particularly relevant to
the homeostasis of bone, as cells undergoing osteoblast formation
can be diverted to adipocytes, leading to the loss of these cells
from the osteoblast pool and the reduction of overall osteogenic
potential (13).
The imbalance of adipocyte differentiation and
regulation of osteoblasts is commonly seen in aging and diabetic
individuals, which can lead to the impairment of fat bone marrow,
impairment of osteoblast renewal and an increase in the incidence
of chronic bone loss (13–15).
Beyond that, adipocytes secrete a series of biologically active
signaling molecules, which can also influence bone homeostasis. It
has been reported that several of these molecules, including
chemorin, resistin, visfatin, leptin and adiponectin, affect the
development and function of osteoblasts and osteoclasts (16–21).
Omentin-1, a 34 kDa adipokine selectively expressed
in omental adipose tissue, is abundant in the plasma (22,23). Omentin-1 has been demonstrated to
serve as a significant factor in multiple physiological processes,
including insulin action, cardiovascular function and the
inflammatory response (23–25). A previous study demonstrated that
omentin-1 plays an important role in protecting against vascular
calcification (26). A clinical
study has also shown that omentin-1 levels are inversely correlated
with obesity and insulin resistance (27). Regarding its effects on bone, a
previous study reported that circulating omentin-1 levels were
inversely correlated with bone mineral density (BMD) in the lumbar
region of the spine in Iranian postmenopausal women (28). The levels of circulating omentin-1
correlates positively with adiponectin and negatively with body
mass index and the leptin levels (29). In a co-culture system of
osteoblasts and osteoclast precursors, omentin-1 can reduce
osteoclast formation by stimulating osteoprotegerin (OPG) (30). To date, only a few studies have
investigated the association between omentin-1 and osteoblast
proliferation and differentiation. Moreover, the osteogenic effect
induced by omentin-1 remains unclear. The TGF-β/Smad signaling
pathway is involved in the regulation of cell differentiation and
it is a key pathway related to osteogenesis (31–33). To understand the underlying
mechanism driving the effects of omentin-1 on osteogenic
differentiation in MC3T3-E1, the role of the TGF-β/Smad pathway in
this process was investigated. The aim of the present study was to
determine the regulatory effects of omentin-1 on osteoblast
viability and differentiation, as well as to explore the underlying
molecular mechanism.
Materials and methods
Cell culture and treatment
Mouse embryo osteoblast precursor cell line MC3T3-E1
was obtained from American Type Culture Collection (ATCC,
CRL-2593™). The cells were cultured in DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (HyClone; Cytiva) and
1% penicillin/streptomycin (Invitrogen; Thermo Fisher Scientific,
Inc.) at 37°C in a 5% CO2 humidified incubator. In order
to stimulate osteogenic induction, the MC3T3-E1 cells were seeded
in DMEM supplemented with 10% FBS, 1% penicillin/streptomycin, 10
mM β-glycerophosphate, 50 µg/ml ascorbic acid and 10 nM
dexamethasone (all Sigma-Aldrich; Merck KGaA) (34). Subsequently, cells were treated
with different doses of omentin-1 (100, 500 and 1,000 ng/ml; Cell
Science, Inc.) for 24 h at 37°C and/or 3 µM SIS3 HCl (Selleck
Chemicals) for 6 h at 37°C for the following experiments.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). iScript™ Reverse Transcription SuperMix kit (Bio-Rad
Laboratories, Inc.) was used to reverse transcribe 2 µg RNA into
cDNA according to the manufacturer's protocol. qPCR was performed
using SYBR-Green MasterMix (Applied Biosystems; Thermo Fisher
Scientific, Inc.) on an ABI PRISM 7900 Sequence Detection system
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The PCR
amplification program was as follows: 94°C for 60 sec, followed by
40 cycles of 94°C for 30 sec, 60°C for 30 sec and 72°C for 60 sec.
The primer sequences used for PCR were as follows: Bone
morphogenetic protein 2 (BMP2) forward, 5′-AGCCCTTCACTGCCATCCTGT-3′
and reverse, 5′-ATTCTCTCGTTCACCGCCCAC-3′; Runt-related
transcription factor 2 (Runx2) forward, 5′-CCCAACTTCCTGTGCTCC-3′
and reverse, 5′-AGTGAAACTCTTGCCTCGTC-3′; Collagen1 forward,
5′-TAAGGGTGACAGAGGCGATG-3′ and reverse, 5′-GGACCGCTAGGACCAGTTTC-3′;
osteopontin (Opn) forward, 5′-TCCAAAGTCAGCCAGGAATCC-3′ and reverse,
5′-CGGAGTTGTCTGTGCTCTTCA-3′; osteocalcin (Ocn) forward,
5′-CTCCTTACCCGGATCCCCTG-3′ and reverse, 5′-GTAGAAGCGCTGGTAGGCGT-3′;
Osterix forward, 5′-TCCCTGGATATGACTCATCCCT-3′ and reverse,
5′-CCAAGGAGTAGGTGTGTTGCC-3′; and GAPDH forward,
5′-GGGAAACTGTGGCGTGAT-3′ and reverse, 5′-GAGTGGGTGTCGCTGTTGA-3′.
Gene expression analysis was performed using the 2−ΔΔCq
method (35) and normalized to
GAPDH.
Cell viability
The cells were cultured in 96-well culture plates at
a density of 2×104 cells/well. After 3 days, a Cell
Counting Kit-8 (CCK-8) assay (Beyotime Institute of Biotechnology)
was performed to evaluate cell viability. The CCK-8 solution was
added to each group, and the cells were incubated at 37°C for 1 h.
The absorbance of each well was measured at a wavelength of 450 nm
using a microplate spectrophotometer.
Western blot assay
Cells in the different treatment groups were lysed
using RIPA lysis buffer (Beyotime Institute of Biotechnology) and
protein concentrations were measured using Bradford reagent
(Bio-Rad Laboratories, Inc.). Equal quantities of proteins (40
µg/lane) were separated via 10% SDS-PAGE (Bio-Rad Laboratories,
Inc.) and transferred to a PVDF membrane (MilliporeSigma). After
being blocked with 5% non-fat milk in 0.1% tris-buffered saline
with Tween-20 for 1 h at room temperature, the membranes were
incubated with primary antibodies against BMP2 (1:1,000; cat. no.
ab214821; Abcam), phosphorylated (p)-Smad1 (1:1,000; cat. no.
ab226821; Abcam), p-Smad5 (1:2,000; cat. no. ab92698; Abcam), Smad1
(1:1,000; cat. no. ab33902; Abcam), Smad5 (1:1,000; cat. no.
ab40771; Abcam), Runx2 (1:1,000; cat. no. ab236639; Abcam),
collagen1 (1:1,000; cat. no. ab138492; Abcam), Opn (1:1,000; cat.
no. ab63856; Abcam), Ocn (1:1,000; cat. no. ab133612; Abcam) and
osterix (1:1,000; cat. no. ab209484; Abcam) and GAPDH (1:2,500;
cat. no. ab9485; Abcam) overnight at 4°C, and then incubated with
the appropriate horseradish peroxidase-conjugated secondary
antibody (1:1,000; cat. no. #7074; Cell Signaling Technology, Inc.)
for 1 h at room temperature. The blots were then visualized using
an enhanced chemiluminescence system (Beyotime Institute of
Biotechnology). Densitometry analysis was performed using ImageJ
(Version 1.49; National Institutes of Health).
Cell differentiation analysis
Alkaline phosphatase (ALP) activity, an early
biochemical marker widely used to assess osteogenic activity, was
measured. Briefly, the cells were seeded in 12-well culture plates
at a density of 4×104 cells per ml for 7 days.
Subsequently, the cells were fixed in 4% paraformaldehyde for 30
min at room temperature, followed by treatment with nitroblue
tetrazolium (Sigma-Aldrich; Merck KGaA) and
5-bromo-4-chloro-3-indolyl phosphate (Sigma-Aldrich; Merck KGaA)
for 2 h at room temperature. Cells were washed with deionized water
and observed under an inverted light microscope (Nikon Corporation;
×200 magnification).
Cell mineralization analysis
Mineralization was evaluated by quantifying the
formation of calcium phosphate in cells using an Alizarin Red
Staining (ARS) kit (Sigma-Aldrich; Merck KGaA). Briefly, the cells
were cultured in 12-well culture plates at a density of
4×104 cells per ml. After 21 days, the cultured cells
were fixed with 95% ethanol for 10 min at room temperature. To
stain the calcium deposits, 2% ARS solution (Sigma-Aldrich; Merck
KGaA) was applied for 15 min at room temperature. To measure the
degree of mineralization, the ARS released from the cell matrix was
incubated in cetyl pyridinium chloride for 15 min at room
temperature and quantified by spectrophotometry at 540 nm.
Statistical analysis
Data are presented as the mean ± standard deviation
of three independent experiments. Differences between groups were
determined using an unpaired two-tailed Student's t-test for
comparisons between two groups, and one-way ANOVA with a post hoc
Tukey's test was used for comparisons between multiple groups.
Statistical analysis was performed using SPSS 13.0 (SPSS, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
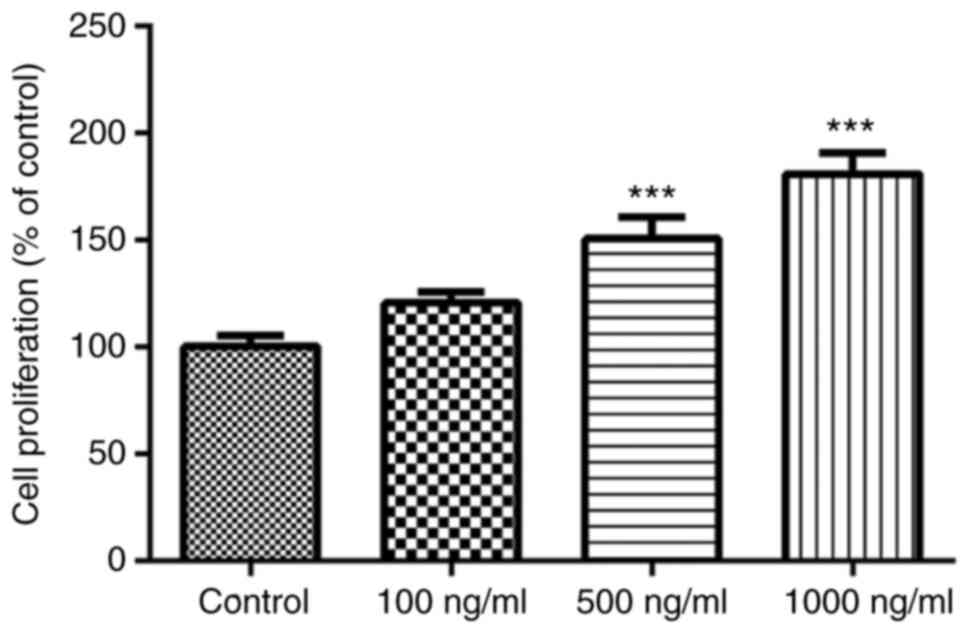
Omentin-1 stimulates viability of
osteoblasts
The effect of omentin-1 on MC3T3-E1 cell viability
was assessed using a CCK-8 assay. As shown in Fig. 1, the viability of osteoblasts was
altered in a dose-dependent manner following treatment with
omentin-1, and the effect was significant when treated with 1,000
ng/ml omentin-1 after 3 days of treatment.
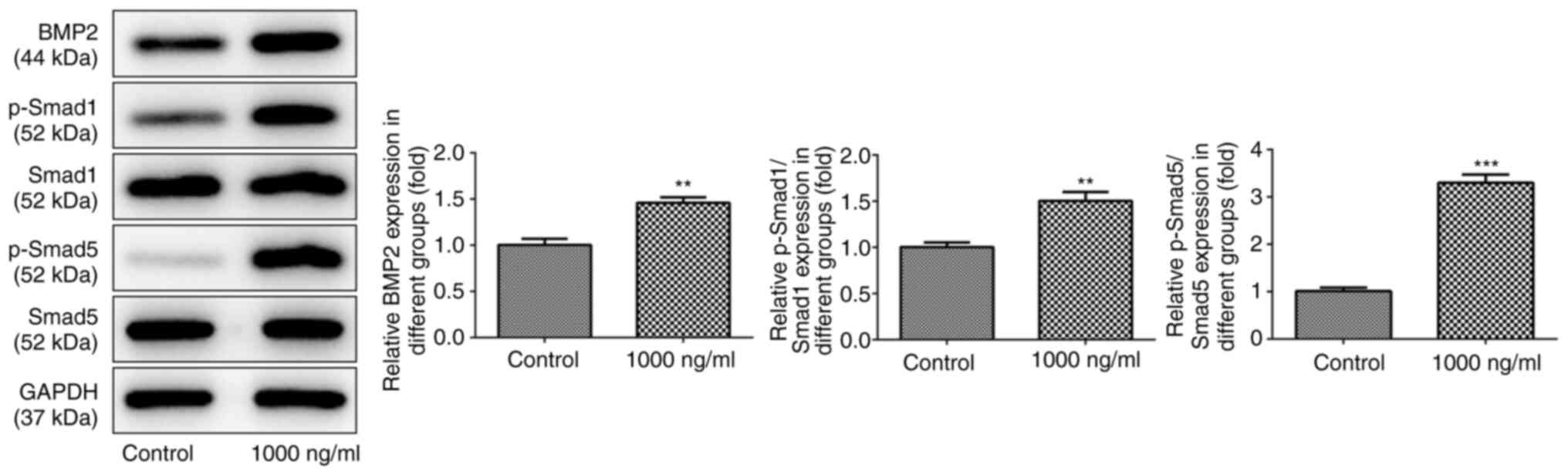
Omentin-1 activates the Smad signaling
pathway and increases BMP2 expression in osteoblasts
To explore the potential mechanisms by which
omentin-1 exerted its effects on osteoblasts, western blotting was
performed. As shown in Fig. 2,
omentin-1 at a dose of 1,000 ng/ml increased the protein expression
levels of BMP2, p-Smad1 and p-Smad5 (P<0.05). However, 1,000
ng/ml omentin-1 did not exert a notable effect on the expression
levels of Smad1 and Smad5.
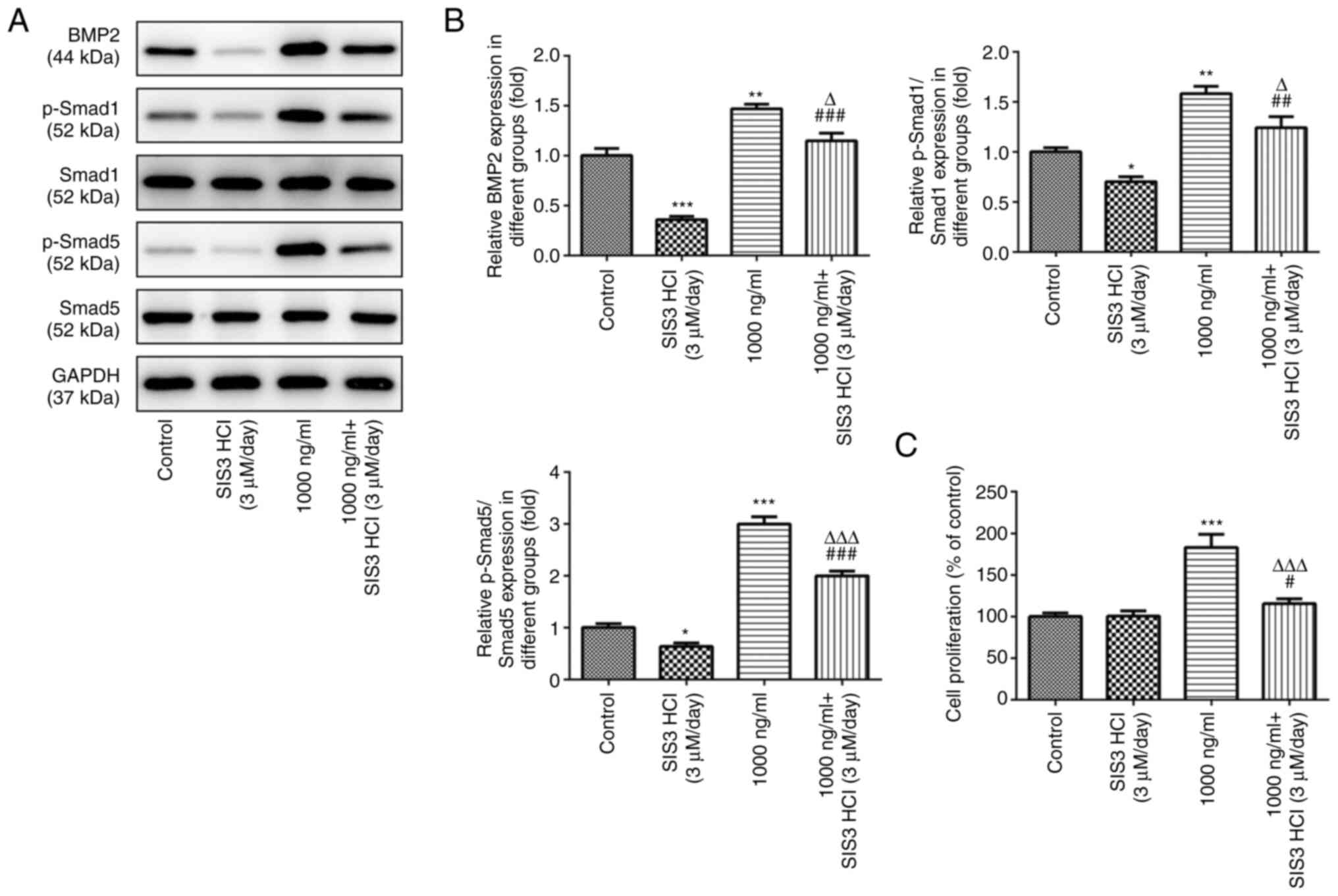
Omentin-1 promotes the viability of
osteoblasts through the TGF-β/Smad signaling pathway
As shown in Fig. 3A
and B, SIS3 HCl, a Smad inhibitor, significantly inhibited BMP2
protein levels and phosphorylation of Smad1 and Smad5 compared with
the control group, while 1,000 ng/ml omentin-1 significantly
increased the expression levels of the three proteins. Meanwhile,
SIS3 HCl attenuated the effects of omentin-1 on the expression of
BMP2, p-Smad1 and p-Smad5 in MC3T3-E1 cells. In addition, CCK-8
analysis showed a significant increase in cell viability in
omentin-1-treated MC3T3-E1 cells, and this increase in viability
was significantly attenuated by SIS3 HCl (Fig. 3C).
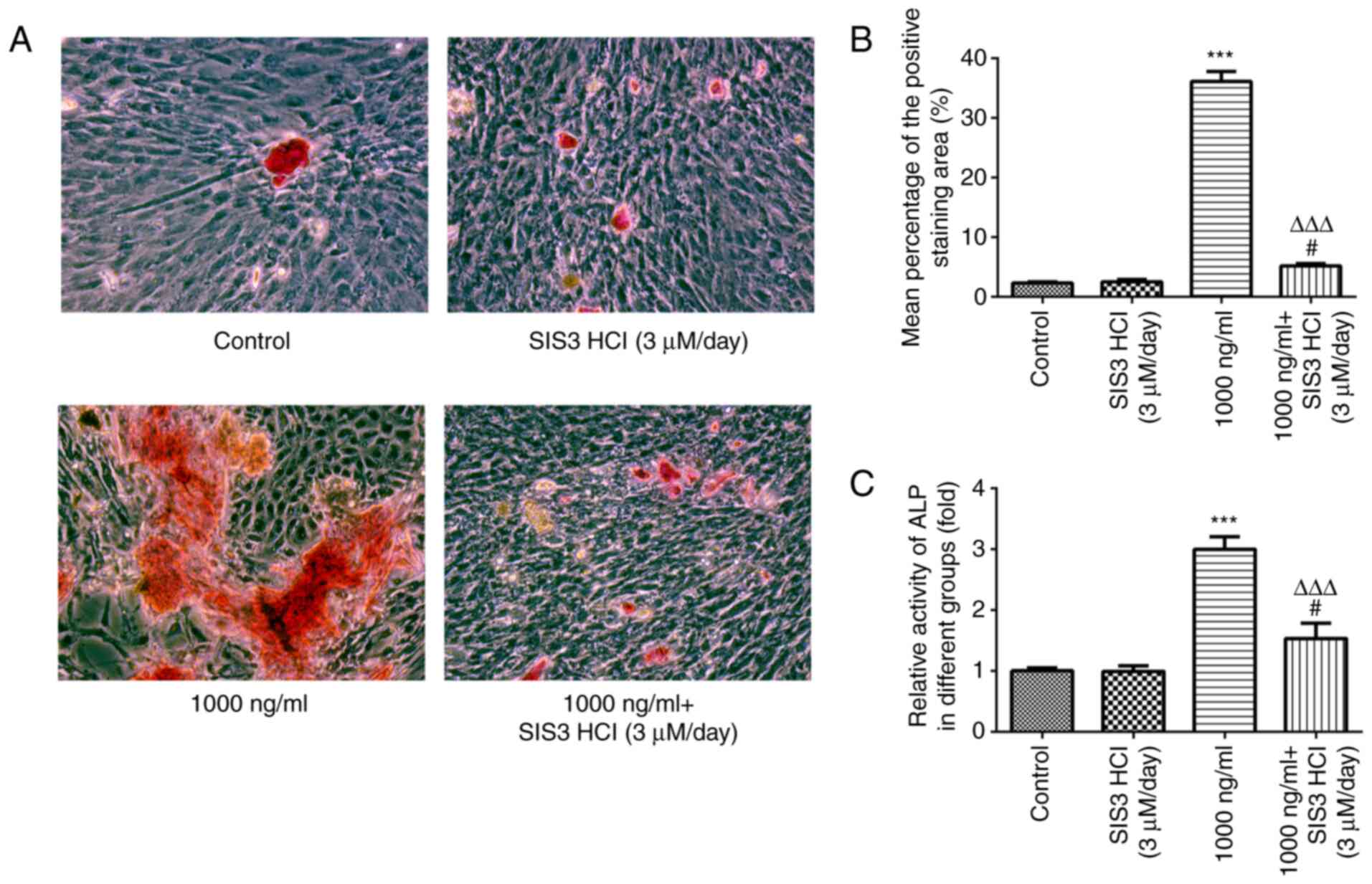
Omentin-1 regulates differentiation
and mineralization of osteoblasts via the TGFβ/Smad signaling
pathway
Next, the effects of omentin-1 on osteoblasts were
assessed. As shown in Fig. 4A and
B, ARS staining in the omentin-1-treated cells showed an
increased mineralization rate compared with the control group, and
SIS3 HCl significantly attenuated the omentin-1-induced
mineralization. In addition, ALP activity was significantly
increased in the omentin-1 group, whereas SIS3 HCl significantly
weakened the activity of ALP in MC3T3-E1 cells (Fig. 4C).
Analysis of cellular Runx2, collagen1,
Opn, Ocn and osterix expression
Data from RT-qPCR and western blot assays showed
that the expression levels of osteogenesis-related proteins,
including BMP2, Runx2, collagen1, Opn, Ocn and osterix, were
significantly upregulated in the omentin-1 group. However, the
expression levels of these proteins in MC3T3-E1 cells treated with
SIS3 HCl were significantly reduced (Fig. 5A-C).
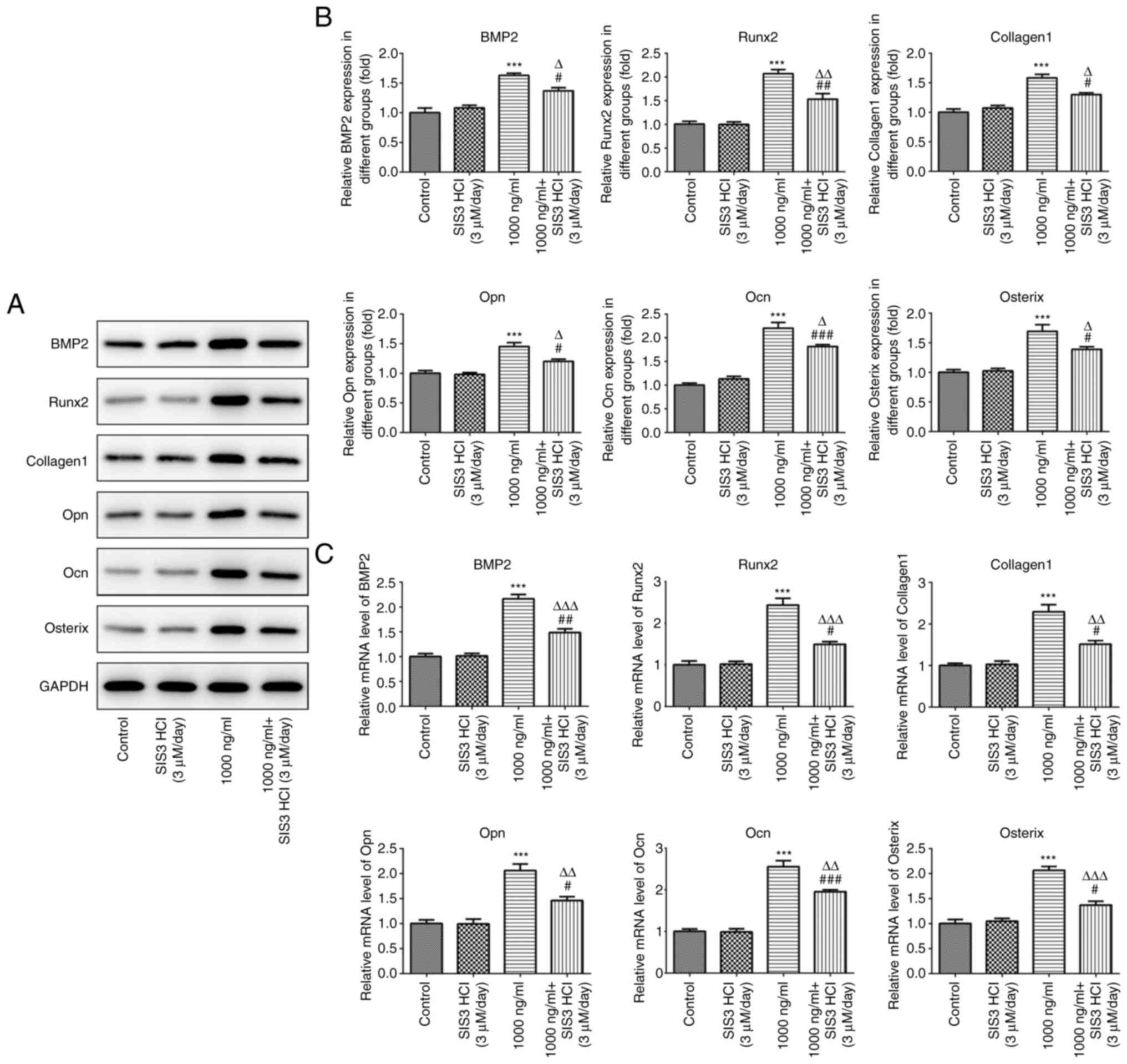 | Figure 5.Effects of omentin-1/transforming
growth factor-β/Smad signaling on the expression of genes related
to bone formation. (A and B) Western blotting analysis was applied
for protein detection of BMP2, Runx2, Collagen1, Opn, Ocn and
Osterix. (C) mRNA levels of BMP2, Runx2, Collagen1, Opn, Ocn and
Osterix were measured by reverse transcription-quantitative PCR.
***P<0.001 vs. control; #P<0.05,
##P<0.01, ###P<0.001 vs. SIS3 HCl
group; ΔP<0.05, ΔΔP<0.01,
ΔΔΔP<0.001 vs. 1,000 ng/ml omentin-1 group. BMP2,
bone morphogenetic protein 2; Runx2, runt-related transcription
factor 2; Opn, osteopontin; Ocn, osteocalcin. |
Discussion
Bone remodeling and skeletal homeostasis are dynamic
life-long processes that rely on the balance and integrated actions
between osteoblastic bone formation and osteoclastic bone
resorption (36–38). Osteoblastic lineage cells interact
with osteoclastic lineage cells, which is necessary for the
formation of functional osteoclasts (39). Certain bone-related diseases, such
as osteoporosis, rheumatoid arthritis and osteoarthritis, are
associated with abnormal bone metabolism and accelerated bone loss
(40). Bone formation and repair
are intrinsically associated with the balanced activity of
osteoblasts (41). Osteoblasts
are responsible for bone matrix secretion and mineralization, and
they tightly regulate osteoclast activation and differentiation
(42). Dysfunctional behaviors of
osteoblasts lead to improper bone matrix deposition and
mineralization, affecting the size, shape and integrity of the
skeletal structures (43).
MC3T3-E1, a popular osteoblast cell line that
represents a pre-osteoblastic phenotype, is a mouse calvaria clonal
cell line that has given rise to several subclones (44). Decreased proliferative activities
have been observed at passage numbers above 36, whereas their
replicative senescence has been displayed when these cells reach a
high passage number of 60, which is similar to that of human cells,
at which point they show inconsistent cell cycling (45). This important feature also makes
these cells suitable as novel models for in vitro bone
research, including bone remodeling and formation (46).
Omentin-1, which was discovered from a human omental
fat cDNA library, is primarily expressed in visceral adipose tissue
(23). Omentin-1 can promote
osteogenic differentiation, although this has been contested
(47–52). Therefore, the present study
investigated the effects of omentin-1 on MC3T3-EI cells by
observing proliferation patterns, with the aim of determining the
regulatory effect of omentin-1 and identifying the underlying
molecular mechanisms. When MC3T3-E1 cells were treated with
different concentrations of omentin-1 (100-1,000 ng/ml) for 3 days,
the viability of MC3T3-E1 cells increased significantly.
Additionally, the effect of omentin-1 was weakened after adding a
BMP inhibitor. This suggested that omentin-1 could stimulate the
viability of osteoblasts. Based on these experiments, 1,000 ng/ml
omentin-1 was used for subsequent experiments, and 0 ng/ml
omentin-1 was used as the control group to further study the
effects of omentin-1 on osteoblasts.
Regulation of gene expression by TGF-β is achieved
primarily through the initiation of intracellular signal
transduction and the activation of the Smad signaling pathway
(53,54). TGF-β promotes osteogenic
differentiation, and at the same time, it also inhibits adipogenic
differentiation of human MSCs (55). The TGF-β/Smad pathway is also key
to adipogenesis (56). TGF-β
inhibits adipogenic differentiation in human preadipocytes, which
is mediated primarily by the Smad family members (57,58). Smad3 inhibits adipogenic
conversion, whereas interfering with Smad3 function confers
resistance to inhibition of adipogenesis by TGF-β (56). The present results showed that
SIS3 HCL treatment significantly inhibited the protein expression
levels of BMP2, p-SMAD1 and p-SMAD5, while 1,000 ng/ml omentin-1
increased the protein expression levels of BMP2 and p-Smad1/5.
However, 1,000 ng/ml omentin-1 exerted no effect on Smad1 and
Smad5, which are also members of the TGF-β/Smad pathway.
To confirm the effects of the omentin-1/TGF-β/Smad
pathway on MC3T3-E1 cells, cells were pretreated with a selective
BMP TGF-β/Smad signaling inhibitor, SIS3 HCl (3 µM per day), 3 days
prior to treatment with 1,000 ng/ml omentin-1. Cell viability was
promoted in the 1,000 ng/ml omentin-1 group, and SIS3 HCl
attenuated the increase in viability mediated by omentin-1. The
results from the CCK-8 assays revealed that omentin-1 significantly
promoted the viability ability of the MC3T3-E1 cells via the
TGF-β/Smad pathway.
In previous years, several studies have investigated
the relationship between omentin-1 and bone metabolism. On the one
hand, adiponectin has been shown to induce osteoblast proliferation
and differentiation (50), as
well as to increase bone mass via osteoclastogenesis suppression
and osteoblastogenesis activation (51), and the results of the present
study were in agreement with these previous findings. Conversely,
adiponectin has been verified to induce osteoclast formation via
stimulation of RANKL and inhibition of OPG production (48,49). This suggests that adipokines also
serve an important role in osteoclastic balance; thus the effects
of adipokines on osteoclasts should form the basis of future
studies. A previous study showed that there was an inverse
correlation between omentin-1 and broadband ultrasound attenuation
levels in postmenopausal women. This association was not mediated
by OPG levels (52). The results
from in vitro and in vivo studies have shown
contradictory results. Results from a meta-analysis of
observational studies demonstrated an inverse relationship between
adiponectin levels and BMD, with five studies showing a negative
association, and another five showing no association (47). The reasons for such discrepant
findings are unclear, but it is conceivable that the
microenvironment in human bodies can be affected by various
factors. The ability of cell culture and animal models to emulate
the complex processes that take place in the human body is limited,
such as possible adiponectin-dependent counter-regulatory
mechanisms.
Osteoporosis, a type of bone-related disease,
results from impaired bone formation and excessive bone resorption.
Current strategies to augment bone formation, such as the use of
growth factors (59), have shown
promising results. However, the osteoblast lineage specification
and bone regeneration remain unclear. An improved understanding of
these may considerably improve the therapeutic strategies
available. In order to determine how omentin-1 guides osteoblast
bone formation, the effects of several potential mediators involved
in the pro-osteogenic pathway were assessed using western blotting
in the current study. Osteoblasts are derived from osteoprogenitors
that differentiate by progressively expressing maturation markers
(60). Runx2 is a transcription
factor that serves a role in osteoblast differentiation. Previous
studies have shown that silencing of Runx2 can block the
differentiation of osteoblasts in mice (61,62). Patients with cleidocranial
dysplasia, which is caused by heterozygous mutations in the Runx2
gene, are characterized by an underdeveloped collarbone, short
stature, excess teeth, fontanelle patency and other bone
growth-related defects (63).
Osterix is a zinc finger transcription factor present in
osteoblasts and serves a leading role in the osteoblast
differentiation process (63). A
previous study showed that Runx2 can regulate the levels of osterix
(64). Collagen-1 is a primary
structural protein present in the extracellular space in the bone,
making up 25–35% of the entire-body protein content (65). Depending upon the degree of
mineralization, collagen tissues may be rigid (bone), compliant
(tendon) or have a gradient from rigid to compliant (cartilage)
(66). As the skeleton is the
main structural component of the body, collagen-1 is responsible
for maintaining the strength of these structures (67). Opn is also known as bone
sialoprotein I, a protein in humans encoded by the SPP1 gene
(68). Ocn is a vitamin
K-dependent calcium-binding protein (69). Opn and Ocn are two typical
biomarkers of osteoblasts, and are involved in osteogenic
differentiation and the mineralization of extracellular matrix
during bone formation and repair (70,71). Previous studies have shown that
Runx2 stimulates the differentiation of MSCs into osteoblasts by
regulating Opn and Ocn activity (61,72). In the present study, it was shown
that omentin-1 significantly upregulated the expression of Runx2,
Collagen-1, Opn, Ocn and osterix, suggesting that it accelerated
bone formation.
Based on the aforementioned results, it was noticed
in the present study that SIS3 HCL treatment alone only affected
the TGF-β/Smad pathway, but had no significant effects on the
viability, differentiation and mineralization of osteoblasts and
the expression of osteogenesis-related proteins. However, SIS3 HCL
exerted obvious roles when the cells were treated with omentin-1.
Hence, we speculated that under normal conditions, osteoblasts were
only partially affected by SIS3 HCL except for the expression of
TGF-β/Smad pathway, but omentin-1 had significant promoting effects
on cell differentiation and mineralization via regulating the
TGF-β/Smad pathway. Thus, there is a noticeable effect of SIS3 HCL
on biological function and signaling and omentin-1-mediated
osteoblasts.
In conclusion, the present study examined the
potential effects of omentin-1 on the viability and differentiation
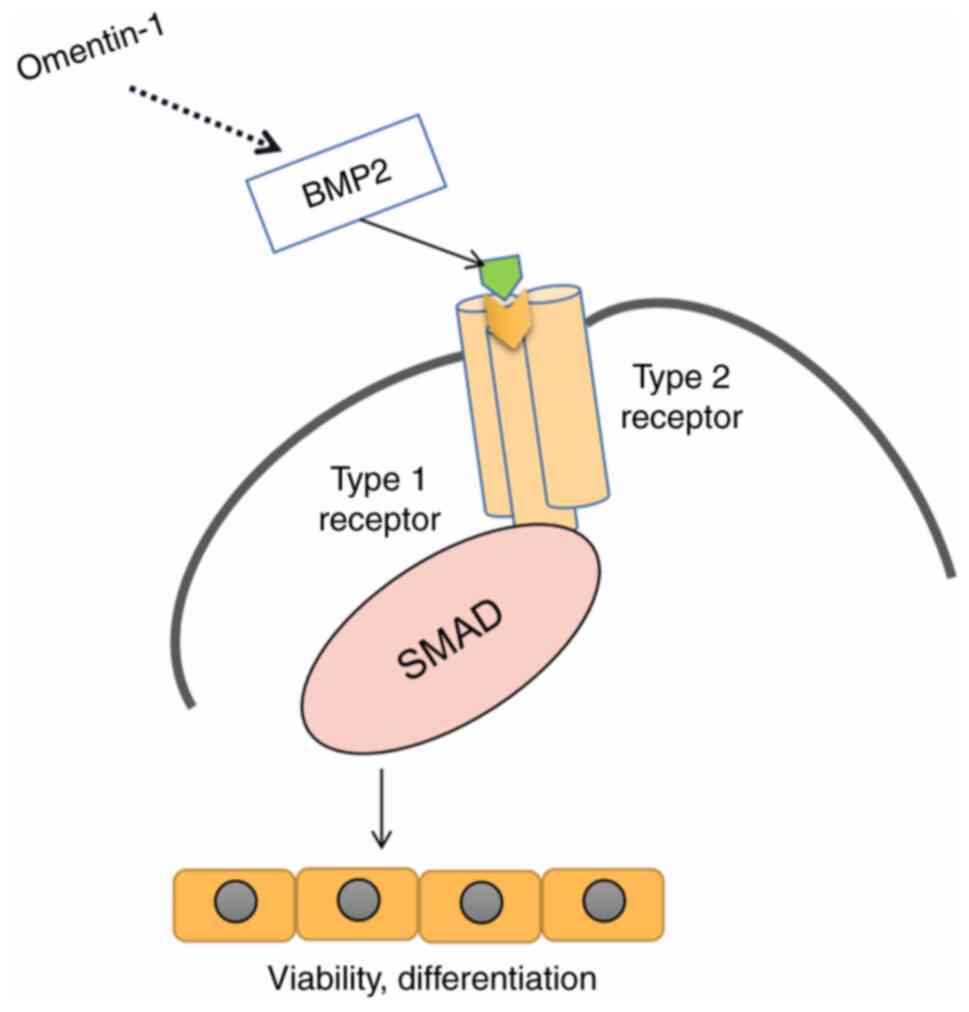
of osteoblasts and the signaling pathways involved (Fig. 6). Omentin-1 promoted osteoblast
viability in a dose-dependent manner. Western blotting revealed
that omentin-1 induced the activation of BMP2 and p-Smad1/5.
Furthermore, omentin-1 promoted osteoblast viability,
differentiation and mineralization, and these effects were impeded
by a TGF-β/Smad inhibitor. These findings indicated that omentin-1
promoted osteoblast viability and differentiation via the
TGF-β/Smad signaling pathway, suggesting that omentin-1 may be a
potential target in the treatment of osteoporosis.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
CT, DL and GT designed the study, drafted and
revised the manuscript. YQ and JZ analyzed the data and searched
the literature. CT, DL, YQ and JZ performed the experiments. CT and
GT confirm the authenticity of all the raw data. All authors have
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Miller PD: Management of severe
osteoporosis. Expert Opin Pharmacother. 17:473–488. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Noh JY, Yang Y and Jung H: Molecular
mechanisms and emerging therapeutics for osteoporosis. Int J Mol
Sci. 21:76232020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Clark S: Osteoporosis-the disease of the
21st century? Lancet. 359:17142002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cooper C: Epidemiology of osteoporosis.
Osteoporos Int. 9 (Suppl 2):S2–S8. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Johnell O and Kanis J: Epidemiology of
osteoporotic fractures. Osteoporos Int. 16 (Suppl 2):S3–S7. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Straka M, Straka-Trapezanlidis M, Deglovic
J and Varga I: Periodontitis and osteoporosis. Neuro Endocrinol
Lett. 36:401–406. 2015.PubMed/NCBI
|
|
7
|
Kurra S, Fink DA and Siris ES:
Osteoporosis-associated fracture and diabetes. Endocrinol Metab
Clin North Am. 43:233–243. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lane NE: Epidemiology, etiology, and
diagnosis of osteoporosis. Am J Obstet Gynecol. 194 (Suppl
2):S3–S11. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Baccaro LF, Conde DM, Costa-Paiva L and
Pinto-Neto AM: The epidemiology and management of postmenopausal
osteoporosis: A viewpoint from Brazil. Clin Interv Aging.
10:583–591. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lane JM, Russell L and Khan SN:
Osteoporosis. Clin Orthop Relat Res. 372:139–150. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen X, Wang Z, Duan N, Zhu G, Schwarz EM
and Xie C: Osteoblast-osteoclast interactions. Connect Tissue Res.
59:99–107. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jayakumar P and Di Silvio L: Osteoblasts
in bone tissue engineering. Proc Inst Mech Eng H. 224:1415–1440.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Malhotra A and Habibovic P: Calcium
phosphates and angiogenesis: Implications and advances for bone
regeneration. Trends Biotechnol. 34:983–992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Leijten J, Chai YC, Papantoniou I, Geris
L, Schrooten J and Luyten FP: Cell based advanced therapeutic
medicinal products for bone repair: Keep it simple? Adv Drug Deliv
Rev. 84:30–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei DX, Dao JW and Chen GQ: A Micro-Ark
for Cells: Highly open porous polyhydroxyalkanoate microspheres as
injectable scaffolds for tissue regeneration. Adv Mater.
30:e18022732018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shen X, Zhang Y, Gu Y, Xu Y, Liu Y, Li B
and Chen L: Sequential and sustained release of SDF-1 and BMP-2
from silk fibroin-nanohydroxyapatite scaffold for the enhancement
of bone regeneration. Biomaterials. 106:205–216. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cahill KS, Chi JH, Day A and Claus EB:
Prevalence, complications, and hospital charges associated with use
of bone-morphogenetic proteins in spinal fusion procedures. JAMA.
302:58–66. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Skovrlj B, Koehler SM, Anderson PA,
Qureshi SA, Hecht AC, Iatridis JC and Cho SK: Association between
BMP-2 and carcinogenicity. Spine (Phila Pa 1976). 40:1862–1871.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu C, Su P, Chen X, Meng Y, Yu W, Xiang AP
and Wang Y: Biocompatibility and osteogenesis of biomimetic
Bioglass-Collagen-Phosphatidylserine composite scaffolds for bone
tissue engineering. Biomaterials. 32:1051–1058. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Fan W, Ma Z, Wu C, Fang W, Liu G
and Xiao Y: The effects of pore architecture in silk fibroin
scaffolds on the growth and differentiation of mesenchymal stem
cells expressing BMP7. Acta Biomater. 6:3021–3028. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Holzwarth JM and Ma PX: Biomimetic
nanofibrous scaffolds for bone tissue engineering. Biomaterials.
32:9622–9629. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schaffler A, Neumeier M, Herfarth H, Furst
A, Scholmerich J and Buchler C: Genomic structure of human omentin,
a new adipocytokine expressed in omental adipose tissue. Biochim
Biophys Acta. 1732:96–102. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang RZ, Lee MJ, Hu H, Pray J, Wu HB,
Hansen BC, Shuldiner AR, Fried SK, McLenithan JC and Gong DW:
Identification of omentin as a novel depot-specific adipokine in
human adipose tissue: Possible role in modulating insulin action.
Am J Physiol Endocrinol Metab. 290:E1253–E1261. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamawaki H, Kuramoto J, Kameshima S, Usui
T, Okada M and Hara Y: Omentin, a novel adipocytokine inhibits
TNF-induced vascular inflammation in human endothelial cells.
Biochem Biophys Res Commun. 408:339–343. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yamawaki H, Tsubaki N, Mukohda M, Okada M
and Hara Y: Omentin, a novel adipokine, induces vasodilation in rat
isolated blood vessels. Biochem Biophys Res Commun. 393:668–672.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Duan XY, Xie PL, Ma YL and Tang SY:
Omentin inhibits osteoblastic differentiation of calcifying
vascular smooth muscle cells through the PI3K/Akt pathway. Amino
Acids. 41:1223–1231. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
de Souza Batista CM, Yang RZ, Lee MJ,
Glynn NM, Yu DZ, Pray J, Ndubuizu K, Patil S, Schwartz A, Kligman
M, et al: Omentin plasma levels and gene expression are decreased
in obesity. Diabetes. 56:1655–1661. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tohidi M, Akbarzadeh S, Larijani B,
Kalantarhormozi M, Ostovar A, Assadi M, Vahdat K, Farrokhnia M,
Sanjdideh Z, Amirinejad R and Nabipour I: Omentin-1, visfatin and
adiponectin levels in relation to bone mineral density in Iranian
postmenopausal women. Bone. 51:876–881. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Moreno-Navarrete JM, Catalán V, Ortega F,
Gómez-Ambrosi J, Ricart W, Frühbeck G and Fernández-Real JM:
Circulating omentin concentration increases after weight loss. Nutr
Metab (Lond). 7:272010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xie H, Xie PL, Wu XP, Chen SM, Zhou HD,
Yuan LQ, Sheng ZF, Tang SY, Luo XH and Liao EY: Omentin-1
attenuates arterial calcification and bone loss in
osteoprotegerin-deficient mice by inhibition of RANKL expression.
Cardiovasc Res. 92:296–306. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang J, Wang M, Chen F, Wei Y, Chen X,
Zhou Y, Yang X, Zhu X, Tu C and Zhang X: Nano-hydroxyapatite
coating promotes porous calcium phosphate ceramic-induced
osteogenesis via BMP/Smad signaling pathway. Int J Nanomedicine.
14:7987–8000. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu Z, Yu Z, Chang H, Wang Y, Xiang H,
Zhang X and Yu B: Strontium-containing α-calcium sulfate
hemihydrate promotes bone repair via the TGF-β/Smad signaling
pathway. Mol Med Rep. 20:3555–3564. 2019.PubMed/NCBI
|
|
33
|
Wang QL, Li HF, Wang DP, Liu ZY, Xiao WW,
Xu LL and Yu S: Effect of GGCX on the differentiation function of
osteoporosis bone marrow mesenchymal stem cells through regulating
TGFβ/smad signaling pathway. Eur Rev Med Pharmacol Sci.
23:7224–7231. 2019.PubMed/NCBI
|
|
34
|
Kim SW, Her SJ, Park SJ, Kim D, Park KS,
Lee HK, Han BH, Kim MS, Shin CS and Kim SY: Ghrelin stimulates
proliferation and differentiation and inhibits apoptosis in
osteoblastic MC3T3-E1 cells. Bone. 37:359–369. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lerner UH, Kindstedt E and Lundberg P: The
critical interplay between bone resorbing and bone forming cells. J
Clin Periodontol. 46 (Suppl 21):33–51. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fili S, Karalaki M and Schaller B:
Mechanism of bone metastasis: The role of osteoprotegerin and of
the host-tissue microenvironment-related survival factors. Cancer
Lett. 283:10–19. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kearns AE, Khosla S and Kostenuik PJ:
Receptor activator of nuclear factor kappaB ligand and
osteoprotegerin regulation of bone remodeling in health and
disease. Endocr Rev. 29:155–192. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Horowitz MC, Xi Y, Wilson K and Kacena MA:
Control of osteoclastogenesis and bone resorption by members of the
TNF family of receptors and ligands. Cytokine Growth Factor Rev.
12:9–18. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wada T, Nakashima T, Hiroshi N and
Penninger JM: RANKL-RANK signaling in osteoclastogenesis and bone
disease. Trends Mol Med. 12:17–25. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Long F: Building strong bones: Molecular
regulation of the osteoblast lineage. Nat Rev Mol Cell Biol.
13:27–38. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nakamichi Y, Udagawa N, Kobayashi Y,
Nakamura M, Yamamoto Y, Yamashita T, Mizoguchi T, Sato M, Mogi M,
Penninger JM and Takahashi N: Osteoprotegerin reduces the serum
level of receptor activator of NF-kappaB ligand derived from
osteoblasts. J Immunol. 178:192–200. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Valenti MT, Dalle Carbonare L and Mottes
M: Osteogenic differentiation in healthy and pathological
conditions. Int J Mol Sci. 18:412016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wan Hasan WN, Abd Ghafar N, Chin KY and
Ima-Nirwana S: Annatto-derived tocotrienol stimulates osteogenic
activity in preosteoblastic MC3T3-E1 cells: A temporal sequential
study. Drug Des Devel Ther. 12:1715–1726. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Czekanska EM, Stoddart MJ, Richards RG and
Hayes JS: In search of an osteoblast cell model for in vitro
research. Eur Cell Mater. 24:1–17. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Aubin JE: Regulation of osteoblast
formation and function. Rev Endocr Metab Disord. 2:81–94. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Biver E, Salliot C, Combescure C, Gossec
L, Hardouin P, Legroux-Gerot I and Cortet B: Influence of
Adipokines and Ghrelin on Bone Mineral Density and Fracture Risk: A
systematic review and meta-analysis. J Clin Endocrinol Metab.
96:2703–2713. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Luo XH, Guo LJ, Xie H, Yuan LQ, Wu XP,
Zhou HD and Liao EY: Adiponectin stimulates RANKL and inhibits OPG
expression in human osteoblasts through the MAPK signaling pathway.
J Bone Miner Res. 21:1648–1656. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang QP, Li XP, Wang M, Zhao LL, Li H, Xie
H and Lu ZY: Adiponectin exerts its negative effect on bone
metabolism via OPG/RANKL pathway: An in vivo study. Endocrine.
47:845–853. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Luo XH, Guo LJ, Yuan LQ, Xie H, Zhou HD,
Wu XP and Liao EY: Adiponectin stimulates human osteoblasts
proliferation and differentiation via the MAPK signaling pathway.
Exp Cell Res. 309:99–109. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Oshima K, Nampei A, Matsuda M, Iwaki M,
Fukuhara A, Hashimoto J, Yoshikawa H and Shimomura I: Adiponectin
increases bone mass by suppressing osteoclast and activating
osteoblast. Biochem Biophys Res Commun. 331:520–526. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wu SS, Liang QH, Liu Y, Cui RR, Yuan LQ
and Liao EY: Omentin-1 stimulates human osteoblast proliferation
through PI3K/Akt signal pathway. Int J Endocrinol. 2013:3689702013.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Miyazono K: TGF-beta signaling by Smad
proteins. Cytokine Growth Factor Rev. 11:15–22. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
ten Dijke P, Miyazono K and Heldin CH:
Signaling inputs converge on nuclear effectors in TGF-beta
signaling. Trends Biochem Sci. 25:64–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
van Zoelen EJ, Duarte I, Hendriks JM and
van der Woning SP: TGFβ-induced switch from adipogenic to
osteogenic differentiation of human mesenchymal stem cells:
Identification of drug targets for prevention of fat cell
differentiation. Stem Cell Res Ther. 7:1232016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Santibanez JF and Kocic J: Transforming
growth factor-β superfamily, implications in development and
differentiation of stem cells. Biomol Concepts. 3:429–445. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Petruschke T, Röhrig K and Hauner H:
Transforming growth factor beta (TGF-beta) inhibits the
differentiation of human adipocyte precursor cells in primary
culture. Int J Obes Relat Metab Disord. 18:532–536. 1994.PubMed/NCBI
|
|
58
|
Choy L, Skillington J and Derynck R: Roles
of autocrine TGF-beta receptor and Smad signaling in adipocyte
differentiation. J Cell Biol. 149:667–682. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
El Bialy I, Jiskoot W and Reza Nejadnik M:
Formulation, delivery and stability of bone morphogenetic proteins
for effective bone regeneration. Pharm Res. 34:1152–1170. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Rutkovskiy A, Stensløkken KO and Vaage IJ:
Osteoblast differentiation at a glance. Med Sci Monit Basic Res.
22:95–106. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yang D, Okamura H and Qiu L: Upregulated
osterix expression elicited by Runx2 and Dlx5 is required for the
accelerated osteoblast differentiation in PP2A Cα-knockdown cells.
Cell Biol Int. 42:403–410. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Komori T: Runx2, an inducer of osteoblast
and chondrocyte differentiation. Histochem Cell Biol. 149:313–323.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Mundlos S, Otto F, Mundlos C, Mulliken JB,
Aylsworth AS, Albright S, Lindhout D, Cole WG, Henn W, Knoll JH, et
al: Mutations involving the transcription factor CBFA1 cause
cleidocranial dysplasia. Cell. 89:773–779. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Nakashima K, Zhou X, Kunkel G, Zhang Z,
Deng JM, Behringer RR and de Crombrugghe B: The novel zinc
finger-containing transcription factor osterix is required for
osteoblast differentiation and bone formation. Cell. 108:17–29.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Varma S, Orgel JP and Schieber JD:
Nanomechanics of type I collagen. Biophys J. 111:50–56. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Murshed M: Mechanism of bone
mineralization. Cold Spring Harb Perspect Med. 8:a0312292018.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Kisling A, Lust RM and Katwa LC: What is
the role of peptide fragments of collagen I and IV in health and
disease? Life Sci. 228:30–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Chen Y, Ou Y, Dong J, Yang G, Zeng Z, Liu
Y, Liu B, Li W, He X and Lan T: Osteopontin promotes collagen I
synthesis in hepatic stellate cells by miRNA-129-5p inhibition. Exp
Cell Res. 362:343–348. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
An J, Yang H, Zhang Q, Liu C, Zhao J,
Zhang L and Chen B: Natural products for treatment of osteoporosis:
The effects and mechanisms on promoting osteoblast-mediated bone
formation. Life Sci. 147:46–58. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Zhou H, Choong P, McCarthy R, Chou ST,
Martin TJ and Ng KW: In situ hybridization to show sequential
expression of osteoblast gene markers during bone formation in
vivo. J Bone Miner Res. 9:1489–1499. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
van Leeuwen JP, van Driel M, van den Bemd
GJ and Pols HA: Vitamin D control of osteoblast function and bone
extracellular matrix mineralization. Crit Rev Eukaryot Gene Expr.
11:199–226. 2001.PubMed/NCBI
|
|
72
|
Takahashi T, Kato S, Suzuki N, Kawabata N
and Takagi M: Autoregulatory mechanism of Runx2 through the
expression of transcription factors and bone matrix proteins in
multipotential mesenchymal cell line, ROB-C26. J Oral Sci.
47:199–207. 2005. View Article : Google Scholar : PubMed/NCBI
|